
Impact of Chitooligosaccharide Conjugated Epigallocatechin Gallate and Non-Thermal High-Voltage Atmospheric Cold Plasma on Vibrio parahaemolyticus: An In Vitro Study and the Use in Blood Clam Meat

 ,
,  , ,
, ,  , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Chemicals and Microbial Media
2.2. Preparation of Bacterial Strain
2.3. High Voltage Cold Atmospheric Plasma (HVACP) in the Package Treatment
2.4. Effect of Combined Treatment of COS-EGCG Conjugate and HVACP on V. parahaemolyticus
2.4.1. Time-Kill Analysis
2.4.2. Cell Membrane Permeability
2.4.3. Protein and Nucleic Acid Leakage
2.4.4. Biofilm Formation
2.4.5. Structure and Morphology of Cells
2.5. Effect of Combined Treatment of COS-EGCG Conjugate and HVACP on V. parahaemolyticus Inoculated on Shucked Blood Clam Meat
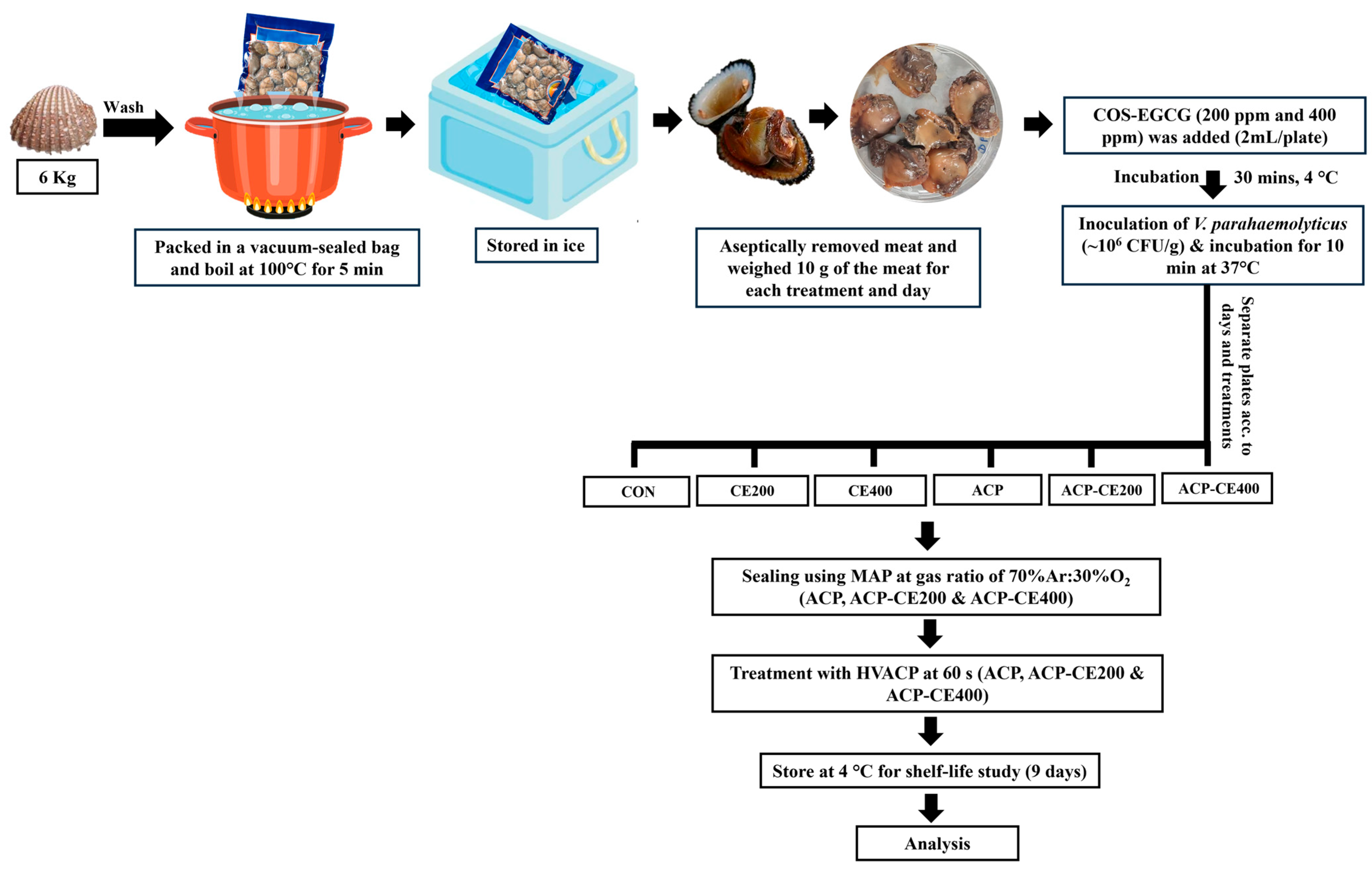
2.5.1. Preparation of Blood Clam Meat
2.5.2. V. parahaemolyticus Count
2.5.3. Scanning Electron Microscopy (SEM)
2.6. Statistical Analysis
3. Results and Discussion
3.1. Effect of Argon-Oxygen Gas Composition on Ozone Generation
3.2. Effects of COS-EGCG Conjugate Without and with HVACP Treatment at Selected Conditions on V. parahaemolyticus Cells
3.2.1. Time-Killing Analysis
3.2.2. Effects of COS-EGCG Conjugate Without and with HVACP Treatment on Cell Membrane Permeability of V. parahaemolyticus
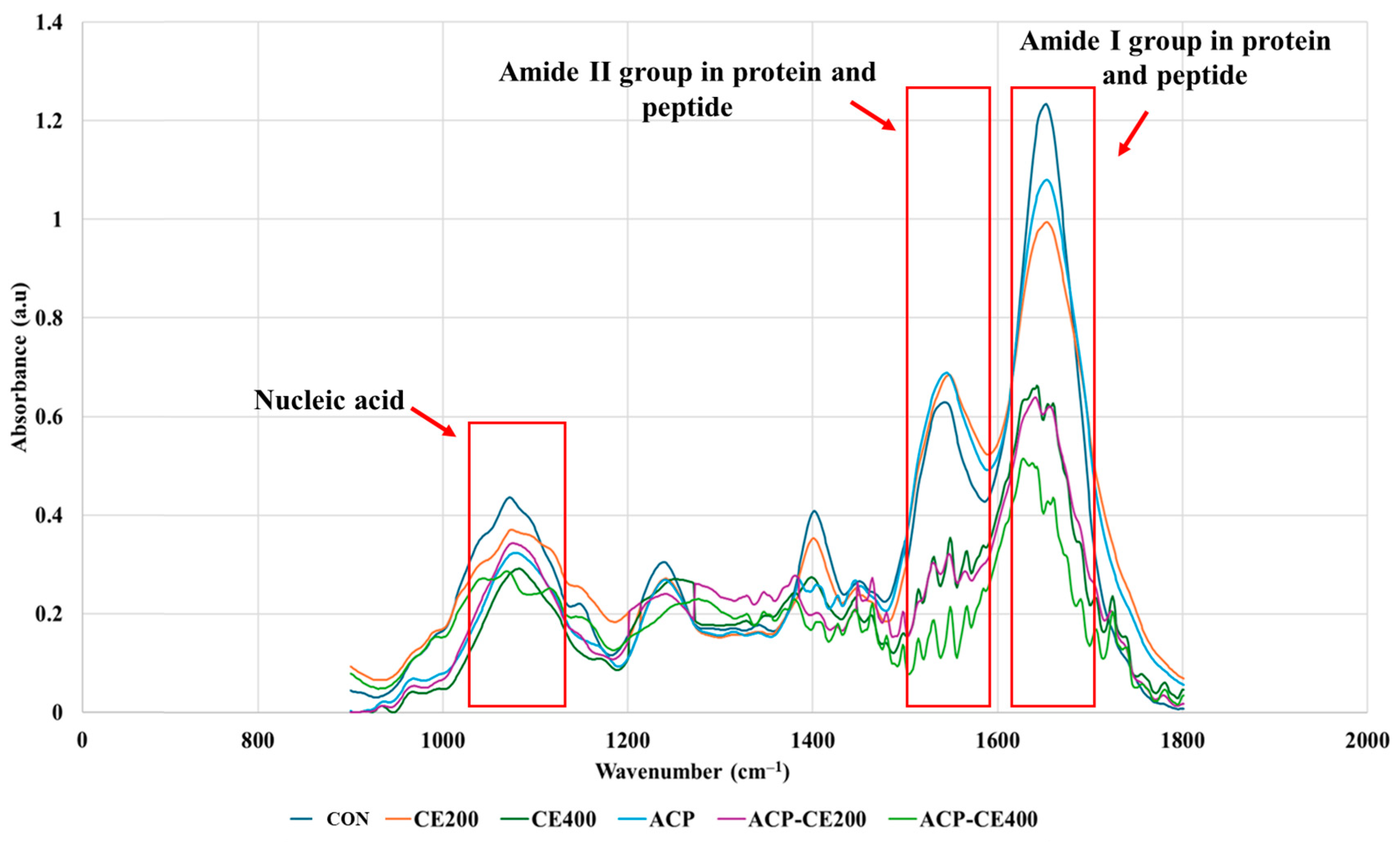
3.2.3. Protein and Nucleic Acid Leakage of V. parahaemolyticus Cells Treated With COS-EGCG Conjugate in Combination With and Without HVACP
3.2.4. Inhibition of Biofilm Formation
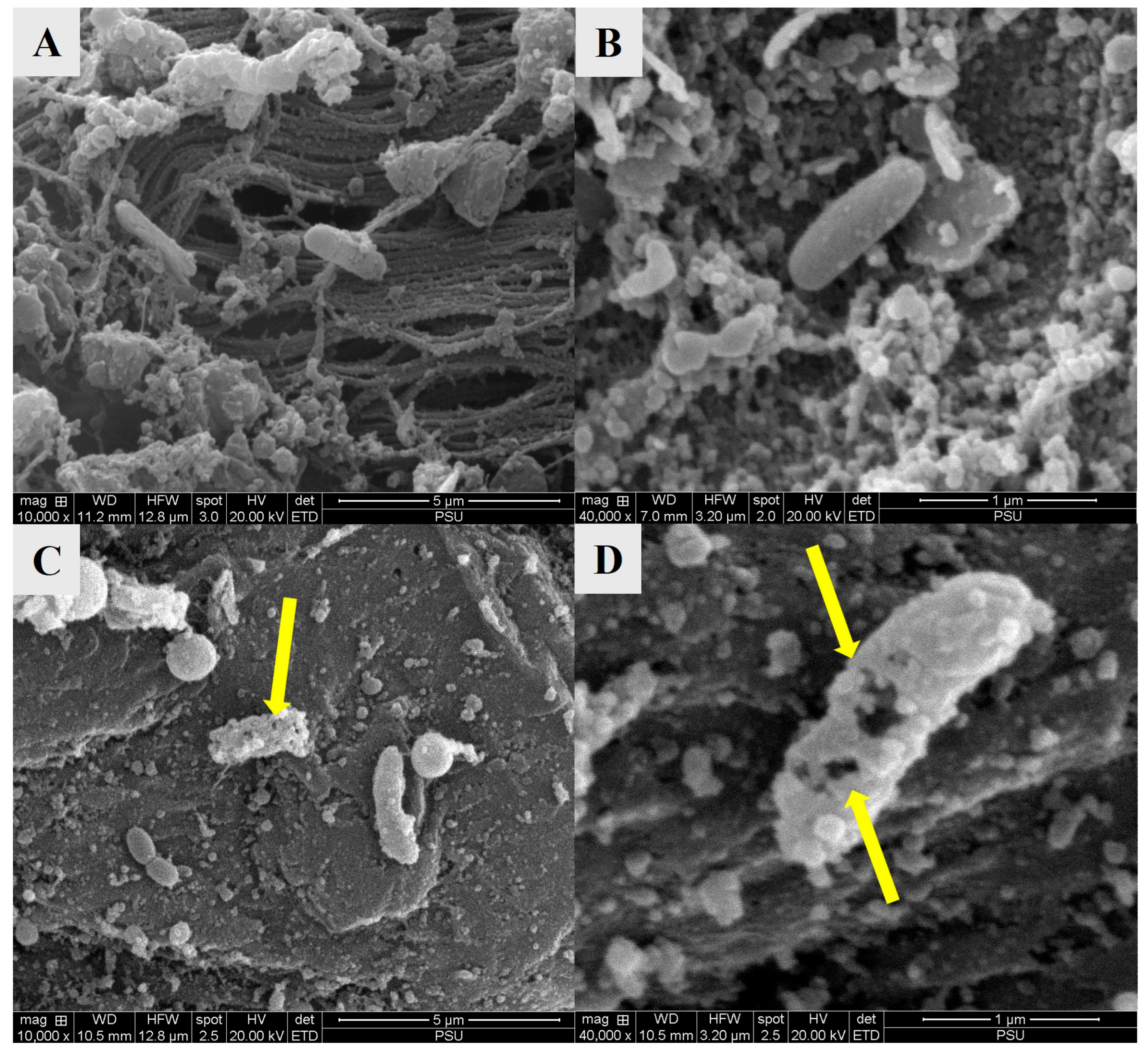
3.2.5. Structural and Morphological Changes Observed by SEM and TEM
3.3. Impact of COS-EGCG Conjugate in Combination with HVACP on V. parahaemolyticus Cells Inoculated on Blood Clam Meat
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Desdouits, M.; Reynaud, Y.; Philippe, C.; Guyader, F.S.L. A comprehensive review for the surveillance of human pathogenic microorganisms in shellfish. Microorganisms 2023, 11, 2218. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.C.L.; Goncalves, A.; Fernandes, C.; Cabecinha, E.; Monteiro, S.; Guedes, H.; Almeida, G.; Garcia, J.; da Silva, G.J.; Varandas, S.; et al. Multidrug-resistant Escherichia coli accumulated by freshwater bivalves: An underestimated risk for public health? Pathogens 2024, 13, 617. [Google Scholar] [CrossRef] [PubMed]
- Kijewska, A.; Koroza, A.; Grudlewska-Buda, K.; Kijewski, T.; Wiktorczyk-Kapischke, N.; Zorena, K.; Skowron, K. Molluscs-A ticking microbial bomb. Front. Microbiol. 2022, 13, 1061223. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.J.; Aleya, A.Y.; Zahan, N.; Paul, C.; Yeasmine, S. Annual reproductive phenology and condition index of blood cockle Tegillarca granosa (L., 1758) collected from the west coast of Moheshkhali Island, Cox’s Bazar, Bangladesh. Ocean Sci. J. 2024, 59, 25. [Google Scholar] [CrossRef]
- Lai, Z.W.; Teoh, H.W.; Lee, C.W.; Lee, S.L.; Saito, H.; Chong, V.C. Macrobenthic community associated with semi-cultured blood cockles (Tegillarca granosa) in tropical mudflats. Cont. Shelf Res. 2020, 195, 104061. [Google Scholar] [CrossRef]
- Sudsandee, S.; Tantrakarnapa, K.; Tharnpoophasiam, P.; Limpanont, Y.; Mingkhwan, R.; Worakhunpiset, S. Evaluating health risks posed by heavy metals to humans consuming blood cockles (Anadara granosa) from the Upper Gulf of Thailand. Environ. Sci. Pollut. Res. Int. 2017, 24, 14605–14615. [Google Scholar] [CrossRef] [PubMed]
- Palamae, S.; Temdee, W.; Buatong, J.; Suyapoh, W.; Sornying, P.; Tsai, Y.-H.; Benjakul, S. Use of high pressure processing in combination with acidic electrolyzed water depuration for the shelf-life extension of blood clam (Tegillarca granosa). Food Control 2024, 156, 110160. [Google Scholar] [CrossRef]
- Lopatek, M.; Wieczorek, K.; Osek, J. Prevalence and antimicrobial resistance of bacterial foodborne pathogens isolated from raw bivalve molluscs subjected to consumption in Poland during a ten-year period. Foods 2022, 11, 3521. [Google Scholar] [CrossRef] [PubMed]
- Ngasotter, S.; Mukherjee, S.; Singh, S.K.; Bharti, D.; Haque, R.; Varshney, S.; Nanda, C.; Waikhom, D.; Devi, M.S.; Singh, A.S. Prevalence, virulence and antibiotic resistance profiles of Vibrio parahaemolyticus from seafood and its environment: An updated review. Mediterr. J. Infect. Microbes Antimicrob. 2022, 11, 1. [Google Scholar] [CrossRef]
- Siddique, A.B.; Moniruzzaman, M.; Ali, S.; Dewan, M.N.; Islam, M.R.; Islam, M.S.; Amin, M.B.; Mondal, D.; Parvez, A.K.; Mahmud, Z.H. Characterization of pathogenic Vibrio parahaemolyticus isolated from fish aquaculture of the southwest coastal area of Bangladesh. Front. Microbiol. 2021, 12, 635539. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.W.; Rukayadi, Y.; Hasan, H.; Thung, T.Y.; Lee, E.; Rollon, W.D.; Hara, H.; Kayali, A.Y.; Nishibuchi, M.; Radu, S. Prevalence and antibiotic resistance patterns of Vibrio parahaemolyticus isolated from different types of seafood in Selangor, Malaysia. Saudi J. Biol. Sci. 2020, 27, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Chinnadurai, S.; Elavarasan, K.; Geethalakshmi, V.; Kripa, V.; Mohamed, K.S. Development of a depuration protocol for commercially important edible bivalve molluscs of India: Ensuring microbiological safety. Food Microbiol. 2023, 110, 104172. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, J.A.; Lamarao, C.V.; Sanches, E.A.; Rodrigues, S.; Fernandes, F.A.N.; Ramos, G.; Esmerino, E.A.; Cruz, A.G.; Campelo, P.H. Cold plasma as a pre-treatment for processing improvement in food: A review. Food Res. Int. 2023, 167, 112663. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, Y.; Pan, W.; Han, Q.; Wei, Y.; Li, Y.; Hu, Y.; Ying, X.; Armani, A.; Guidi, A.; et al. Antibacterial mechanism of atmospheric cold plasma against Pseudomonas fluorescens and Pseudomonas putida and its preservation application on in-packaged red shrimp paste. Food Chem. 2025, 464, 141590. [Google Scholar] [CrossRef] [PubMed]
- Olatunde, O.O.; Shiekh, K.A.; Benjakul, S. Pros and cons of cold plasma technology as an alternative non-thermal processing technology in seafood industry. Trends Food Sci. Technol. 2021, 111, 617–627. [Google Scholar] [CrossRef]
- Laroque, D.A.; Seó, S.T.; Valencia, G.A.; Laurindo, J.B.; Carciofi, B.A.M. Cold plasma in food processing: Design, mechanisms, and application. J. Food Eng. 2022, 312, 110748. [Google Scholar] [CrossRef]
- Oner, M.E.; Gultekin Subasi, B.; Ozkan, G.; Esatbeyoglu, T.; Capanoglu, E. Efficacy of cold plasma technology on the constituents of plant-based food products: Principles, current applications, and future potentials. Food Res. Int. 2023, 172, 113079. [Google Scholar] [CrossRef] [PubMed]
- Nwabor, O.F.; Onyeaka, H.; Miri, T.; Obileke, K.; Anumudu, C.; Hart, A. A cold plasma technology for ensuring the microbiological safety and quality of foods. Food Eng. Rev. 2022, 14, 535–554. [Google Scholar] [CrossRef] [PubMed]
- Nikzadfar, M.; Kazemi, A.; Abooei, R.; Abbaszadeh, R.; Firouz, M.S.; Akbarnia, A.; Rashvand, M. Application of cold plasma technology on the postharvest preservation of in-packaged fresh fruit and vegetables: Recent challenges and development. Food Bioprocess Technol. 2024, 17, 4473–4505. [Google Scholar] [CrossRef]
- Hernandez-Torres, C.J.; Reyes-Acosta, Y.K.; Chavez-Gonzalez, M.L.; Davila-Medina, M.D.; Kumar Verma, D.; Martinez-Hernandez, J.L.; Narro-Cespedes, R.I.; Aguilar, C.N. Recent trends and technological development in plasma as an emerging and promising technology for food biosystems. Saudi J. Biol. Sci. 2022, 29, 1957–1980. [Google Scholar] [CrossRef] [PubMed]
- Giannoglou, M.; Dimitrakellis, P.; Efthimiadou, A.; Gogolides, Ε.; Katsaros, G. Comparative study on the effect of cold atmospheric plasma, ozonation, pulsed electromagnetic fields and high-pressure technologies on sea bream fillet quality indices and shelf Life. Food Eng. Rev. 2020, 13, 175–184. [Google Scholar] [CrossRef]
- Cheng, J.-H.; Lv, X.; Pan, Y.; Sun, D.-W. Foodborne bacterial stress responses to exogenous reactive oxygen species (ROS) induced by cold plasma treatments. Trends Food Sci. Technol. 2020, 103, 239–247. [Google Scholar] [CrossRef]
- Sreelakshmi, V.P.; Vendan, S.E.; Negi, P.S. The effect of cold plasma treatment on quality attributes and shelf life of apples. Postharvest Biol. Technol. 2024, 214, 112975. [Google Scholar] [CrossRef]
- Dai, W.; Wang, W.; Gu, S.; Xu, M.; Yao, H.; Zhou, X.; Ding, Y. Effect of chitosan-epigallocatechin gallate coating on volatile flavor compounds retention in bighead carp (Aristichthys nobilis) fillets during chilled storage. LWT 2022, 169, 114027. [Google Scholar] [CrossRef]
- Qi, Z.; Tan, W.; Guo, Z.; Jiang, A. Preparation and characterization of polyphenol-functionalized chitooligosaccharide pyridinium salts with antioxidant activity. Mar. Drugs 2025, 23, 150. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Benjakul, S. The combined effect of squid pen chitooligosaccharides and high voltage cold atmospheric plasma on the shelf-life extension of Asian sea bass slices stored at 4 °C. Innov. Food Sci. Emerg. Technol. 2020, 64, 102339. [Google Scholar] [CrossRef]
- Bourke, P.; Ziuzina, D.; Han, L.; Cullen, P.J.; Gilmore, B.F. Microbiological interactions with cold plasma. J. Appl. Microbiol. 2017, 123, 308–324. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.H.; Palamae, S.; Yingkajorn, M.; Benjakul, S.; Singh, A.; Buatong, J. Multidrug-resistance of Vibrio species in bivalve mollusks from southern Thailand: Isolation, identification, pathogenicity, and their sensitivity toward chitooligosaccharide-epigallocatechin-3-gallate conjugate. Foods 2024, 13, 2375. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Singh, A.; Zhang, B.; Visessanguan, W.; Benjakul, S. Chitooligosaccharide conjugates prepared using several phenolic compounds via ascorbic acid/H2O2 free radical grafting: Characteristics, antioxidant, antidiabetic, and antimicrobial activities. Foods 2022, 11, 920. [Google Scholar] [CrossRef] [PubMed]
- Palamae, S.; Mittal, A.; Buatong, J.; Zhang, B.; Hong, H.; Benjakul, S. Chitooligosaccharide-catechin conjugate: Antimicrobial mechanisms toward Vibrio parahaemolyticus and its use in shucked Asian green mussel. Food Control 2023, 151, 109794. [Google Scholar] [CrossRef]
- Wang, R.; Hu, X.; Agyekumwaa, A.K.; Li, X.; Xiao, X.; Yu, Y. Synergistic effect of kojic acid and tea polyphenols on bacterial inhibition and quality maintenance of refrigerated sea bass (Lateolabrax japonicus) fillets. LWT 2021, 137, 110452. [Google Scholar] [CrossRef]
- Wang, L.H.; Chen, L.; Zhao, S.; Huang, Y.; Zeng, X.A.; Aadil, R.M. Inactivation efficacy and mechanisms of atmospheric cold plasma on Alicyclobacillus acidoterrestris: Insight into the influence of growth temperature on survival. Front. Nutr. 2022, 9, 1012901. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Mei, J.; Xie, J. Elucidating antibacterial activity and mechanism of daphnetin against Pseudomonas fluorescens and Shewanella putrefaciens. J. Food Qual. 2020, 2020, 6622355. [Google Scholar] [CrossRef]
- Feng, J.; Wang, T.; Zhang, S.; Shi, W.; Zhang, Y. An optimized SYBR Green I/PI assay for rapid viability assessment and antibiotic susceptibility testing for Borrelia burgdorferi. PLoS ONE 2014, 9, e111809. [Google Scholar] [CrossRef] [PubMed]
- Buatong, J.; Mittal, A.; Mittraparp-Arthorn, P.; Palamae, S.; Saetang, J.; Benjakul, S. Bactericidal action of shrimp shell chitooligosaccharide conjugated with epigallocatechin gallate (COS-EGCG) against Listeria monocytogenes. Foods 2023, 12, 634. [Google Scholar] [CrossRef] [PubMed]
- Buatong, J.; Rukachaisirikul, V.; Sangkanu, S.; Surup, F.; Phongpaichit, S. Antifungal metabolites from marine-derived streptomyces sp. AMA49 against Pyricularia oryzae. J. Pure Appl. Microbiol. 2019, 13, 653–665. [Google Scholar] [CrossRef]
- Mallick, S.R.; Sarangi, S.C.; Xess, I.; Sood, S.; Das, B.K.; Kapil, A.; Singh, G.; Gautam, H.; Bhatt, S.; Kar, S.; et al. Antimicrobial efficacy of argon cold atmospheric pressure plasma jet on clinical isolates of multidrug-resistant ESKAPE bacteria. TechRxiv 2023, 7, 421–428. [Google Scholar] [CrossRef]
- Hu, J.; Chen, J.; Zheng, Y.; Huang, J.; Xie, K.; Li, Y.; Peng, S.; Chen, J.; Deng, S. Effect of atmospheric cold plasma treatment modes on the quality of red shrimp (Solenocera crassicornis) during cold chain storage. LWT 2023, 190, 115543. [Google Scholar] [CrossRef]
- Dharini, M.; Jaspin, S.; Mahendran, R. Cold plasma reactive species: Generation, properties, and interaction with food biomolecules. Food Chem. 2023, 405, 134746. [Google Scholar] [CrossRef] [PubMed]
- Olatunde, O.O.; Benjakul, S.; Vongkamjan, K. High voltage cold atmospheric plasma: Antibacterial properties and its effect on quality of Asian sea bass slices. Innov. Food Sci. Emerg. Technol. 2019, 52, 305–312. [Google Scholar] [CrossRef]
- Hemmati, V.; Garavand, F.; Khorshidian, N.; Cacciotti, I.; Goudarzi, M.; Chaichi, M.; Tiwari, B.K. Impact of cold atmospheric plasma on microbial safety, total phenolic and flavonoid contents, antioxidant activity, volatile compounds, surface morphology, and sensory quality of green tea powder. Food Biosci. 2021, 44, 101348. [Google Scholar] [CrossRef]
- Niveditha, A.; Pandiselvam, R.; Prasath, V.A.; Singh, S.K.; Gul, K.; Kothakota, A. Application of cold plasma and ozone technology for decontamination of Escherichia coli in foods—A review. Food Control 2021, 130, 108338. [Google Scholar] [CrossRef]
- Mayookha, V.P.; Pandiselvam, R.; Kothakota, A.; Padma Ishwarya, S.; Chandra Khanashyam, A.; Kutlu, N.; Rifna, E.J.; Kumar, M.; Panesar, P.S.; Abd El-Maksoud, A.A. Ozone and cold plasma: Emerging oxidation technologies for inactivation of enzymes in fruits, vegetables, and fruit juices. Food Control 2023, 144, 109399. [Google Scholar] [CrossRef]
- Moreno-Vasquez, M.J.; Plascencia-Jatomea, M.; Sanchez-Valdes, S.; Tanori-Cordova, J.C.; Castillo-Yanez, F.J.; Quintero-Reyes, I.E.; Graciano-Verdugo, A.Z. Characterization of epigallocatechin-gallate-grafted chitosan nanoparticles and evaluation of their antibacterial and antioxidant potential. Polymers 2021, 13, 1375. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, S.Z.; Chen, J.Y.; Chen, D.Z.; Deng, S.G.; Xu, B. Effect of cold plasma on maintaining the quality of chub mackerel (Scomber japonicus): Biochemical and sensory attributes. J. Sci. Food Agric. 2019, 99, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, J.; Su, W.; Chen, G.; Zhang, J. Effects of mango and partridge tea extracts on microbial, physical, and chemical properties of tilapia fillets treated with in-package cold plasma during refrigerator storage. Food Qual. Saf. 2024, 8, fyae001. [Google Scholar] [CrossRef]
- Gao, Y.; Zhuang, H.; Yeh, H.-Y.; Bowker, B.; Zhang, J. Effect of rosemary extract on microbial growth, pH, color, and lipid oxidation in cold plasma-processed ground chicken patties. Innov. Food Sci. Emerg. Technol. 2019, 57, 102168. [Google Scholar] [CrossRef]
- Rosenberg, M.; Azevedo, N.F.; Ivask, A. Propidium iodide staining underestimates viability of adherent bacterial cells. Sci. Rep. 2019, 9, 6483. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Luo, L.; Chen, J.; Sun, H.; Wang, X.; Yi, Y.; Lv, X. Cell wall and DNA damage of Staphylococcus aureus by bacteriocin BM1157. LWT 2020, 134, 109842. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, C.; Han, Q. Mechanisms of bacterial inhibition and tolerance around cold atmospheric plasma. Appl. Microbiol. Biotechnol. 2023, 107, 5301–5316. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Patil, S.; Boehm, D.; Milosavljevic, V.; Cullen, P.J.; Bourke, P. Mechanisms of inactivation by high-voltage atmospheric cold plasma differ for Escherichia coli and Staphylococcus aureus. Appl. Environ. Microbiol. 2016, 82, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Shimatani, K.; Ozawa, T.; Shigemune, N.; Tomiyama, D.; Yui, K.; Katsuki, M.; Ikeda, K.; Nonaka, A.; Miyamoto, T. Mechanism for the antibacterial action of epigallocatechin gallate (EGCg) on Bacillus subtilis. Biosci. Biotechnol. Biochem. 2015, 79, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Bayati, M.; Lund, M.N.; Tiwari, B.K.; Poojary, M.M. Chemical and physical changes induced by cold plasma treatment of foods: A critical review. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13376. [Google Scholar] [CrossRef] [PubMed]
- De Meutter, J.; Goormaghtigh, E. Evaluation of protein secondary structure from FTIR spectra improved after partial deuteration. Eur. Biophys. J. 2021, 50, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiedz, I.; Wasko, A.; Pawlat, J.; Polak-Berecka, M. The state of research on antimicrobial activity of cold plasma. Pol. J. Microbiol. 2019, 68, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Yu, H.; Qiu, W.; Mei, J.; Xie, J. Antimicrobial effect of epigallocatechin gallate against Shewanella putrefaciens ATCC 8071: A study based on cell membrane and biofilm. Curr. Microbiol. 2022, 79, 297. [Google Scholar] [CrossRef] [PubMed]
- Saharan, B.S.; Beniwal, N.; Duhan, J.S. From formulation to function: A detailed review of microbial biofilms and their polymer-based extracellular substances. Microbe 2024, 5, 100194. [Google Scholar] [CrossRef]
- Hao, S.; Yang, D.; Zhao, L.; Shi, F.; Ye, G.; Fu, H.; Lin, J.; Guo, H.; He, R.; Li, J.; et al. EGCG-mediated potential inhibition of biofilm development and quorum sensing in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 4946. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Li, C.; Cui, H.; Lin, L. Feasibility of cold plasma for the control of biofilms in food industry. Trends Food Sci. Technol. 2020, 99, 142–151. [Google Scholar] [CrossRef]
- Ignasimuthu, K.; Prakash, R.; Murthy, P.S.; Subban, N. Enhanced bioaccessibility of green tea polyphenols and lipophilic activity of EGCG octaacetate on gram-negative bacteria. LWT 2019, 105, 103–109. [Google Scholar] [CrossRef]
- Das, S.; Gajula, V.P.; Mohapatra, S.; Singh, G.; Kar, S. Role of cold atmospheric plasma in microbial inactivation and the factors affecting its efficacy. Health Sci. Rev. 2022, 4, 100037. [Google Scholar] [CrossRef]
- Ravash, N.; Hesari, J.; Feizollahi, E.; Dhaliwal, H.K.; Roopesh, M.S. Valorization of cold plasma technologies for eliminating biological and chemical food hazards. Food Eng. Rev. 2023, 16, 22–58. [Google Scholar] [CrossRef]
- Bae, J.Y.; Seo, Y.H.; Oh, S.W. Antibacterial activities of polyphenols against foodborne pathogens and their application as antibacterial agents. Food Sci. Biotechnol. 2022, 31, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pipliya, S.; Srivastav, P.P.; Srivastava, B. Exploring the role of various feed gases in cold plasma technology: A comprehensive review. Food Bioprocess Technol. 2023, 17, 3367–3407. [Google Scholar] [CrossRef]
- Rathod, N.B.; Ranveer, R.C.; Bhagwat, P.K.; Ozogul, F.; Benjakul, S.; Pillai, S.; Annapure, U.S. Cold plasma for the preservation of aquatic food products: An overview. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4407–4425. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Cheng, J.H. Evaluation of the effects of cold plasma on cell membrane lipids and oxidative injury of Salmonella typhimurium. Molecules 2022, 27, 640. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Details of the Treatments |
|---|---|
| Control | bacterial inoculum without any treatment |
| CE200 | bacterial inoculum treated with 200 ppm of COS-EGCG conjugate |
| CE400 | bacterial inoculum treated with 400 ppm of COS-EGCG conjugate |
| ACP | bacterial inoculum treated with HVACP using gas mixture ratio of 70% Ar:30% O2 for 60 s |
| ACP-CE200 | bacterial inoculum treated with 200 ppm of COS-EGCG conjugate combined with HVACP using gas mixture ratio of 70% Ar:30% O2 for 60 s |
| ACP-CE400 | bacterial inoculum treated with 400 ppm of COS-EGCG conjugate combined with HVACP using gas mixture ratio of 70% Ar:30% O2 for 60 s |
| Gas Composition | Ozone Concentration (ppm) | |||
|---|---|---|---|---|
| 70% Ar + 30% O2 | 80% Ar + 20% O2 | 90% Ar + 10% O2 | ||
| Time | 20 s | 200 | 80 | 100 |
| 60 s | 240 | 20 | 20 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, M.H.; Singh, A.; Singh, A.; Benjakul, S.; Palamae, S.; Mittal, A.; Buatong, J. Impact of Chitooligosaccharide Conjugated Epigallocatechin Gallate and Non-Thermal High-Voltage Atmospheric Cold Plasma on Vibrio parahaemolyticus: An In Vitro Study and the Use in Blood Clam Meat. Foods 2025, 14, 2577. https://doi.org/10.3390/foods14152577
Sharma MH, Singh A, Singh A, Benjakul S, Palamae S, Mittal A, Buatong J. Impact of Chitooligosaccharide Conjugated Epigallocatechin Gallate and Non-Thermal High-Voltage Atmospheric Cold Plasma on Vibrio parahaemolyticus: An In Vitro Study and the Use in Blood Clam Meat. Foods. 2025; 14(15):2577. https://doi.org/10.3390/foods14152577
Chicago/Turabian StyleSharma, Mruganxi Harshad, Avtar Singh, Ankita Singh, Soottawat Benjakul, Suriya Palamae, Ajay Mittal, and Jirayu Buatong. 2025. "Impact of Chitooligosaccharide Conjugated Epigallocatechin Gallate and Non-Thermal High-Voltage Atmospheric Cold Plasma on Vibrio parahaemolyticus: An In Vitro Study and the Use in Blood Clam Meat" Foods 14, no. 15: 2577. https://doi.org/10.3390/foods14152577
APA StyleSharma, M. H., Singh, A., Singh, A., Benjakul, S., Palamae, S., Mittal, A., & Buatong, J. (2025). Impact of Chitooligosaccharide Conjugated Epigallocatechin Gallate and Non-Thermal High-Voltage Atmospheric Cold Plasma on Vibrio parahaemolyticus: An In Vitro Study and the Use in Blood Clam Meat. Foods, 14(15), 2577. https://doi.org/10.3390/foods14152577