
Bioactive Polysaccharides Prevent Lipopolysaccharide-Induced Intestinal Inflammation via Immunomodulation, Antioxidant Activity, and Microbiota Regulation

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Immune Organ Indices
2.4. Antioxidant Capacity Assessment
2.5. Immune Parameter Analysis
2.6. Histological Characterization of Intestinal Tissues
2.7. Immune and Tight Junction-Related Gene Expression
2.8. Analysis of Gut Microbiota Composition
2.9. Statistical Analysis and Data Visualization
3. Results
3.1. Relief of Inflammatory Symptoms by Five Polysaccharides
3.2. Effects of Five Polysaccharides on Immune Organ Indices
3.3. Effects of Five Polysaccharides on Antioxidant Parameters
3.4. Effects of Five Polysaccharides on the Levels of Immune-Related Cytokines and Inflammatory Factors
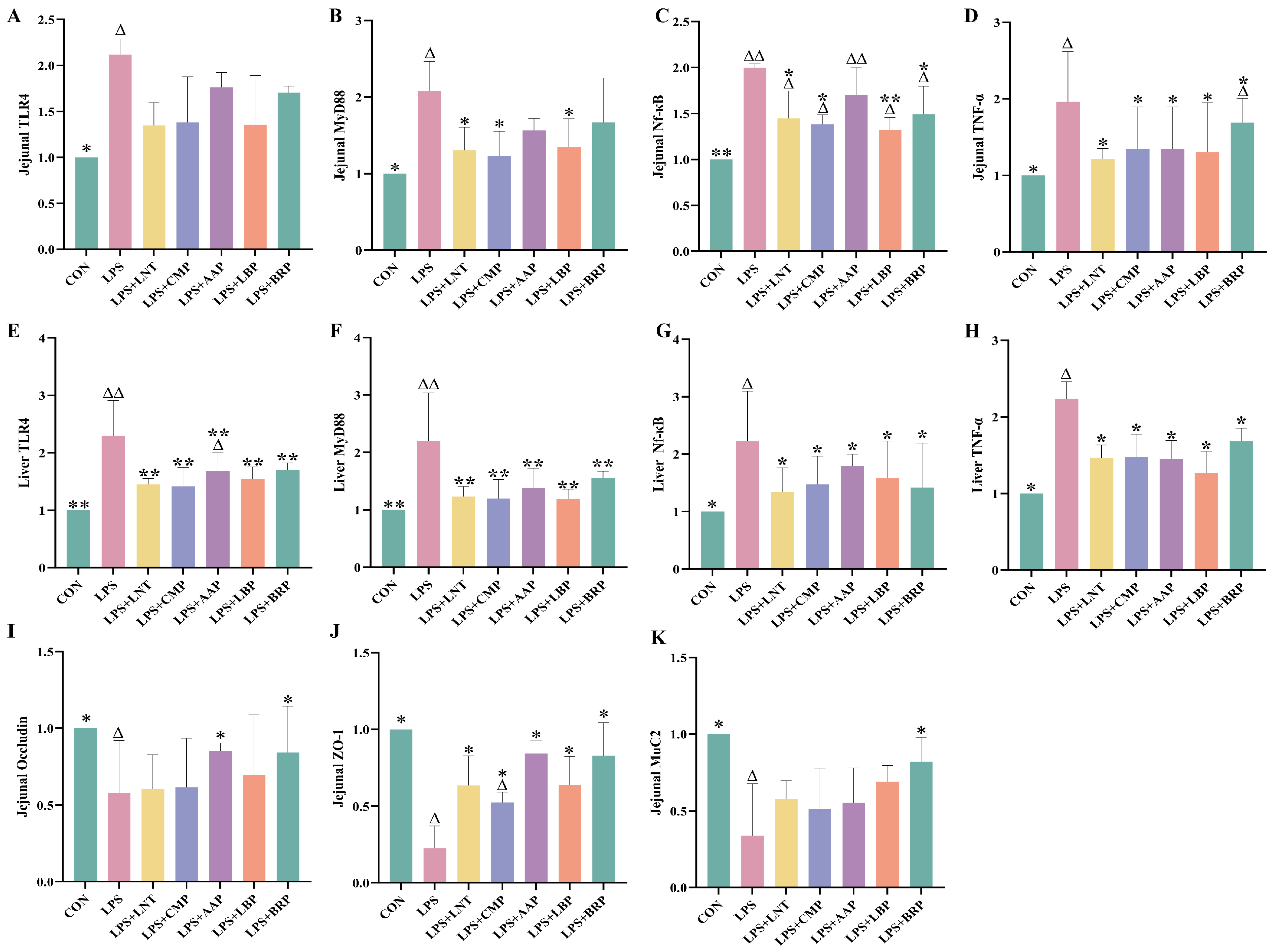
3.5. Effects of Five Polysaccharides on TLR4/NF-κB Signaling Pathway-Associated Gene Expression
3.6. Effects of Five Polysaccharides on the Expression of Genes Related to Jejunal Intestinal Barrier
3.7. Effects of Five Polysaccharides on Jejunal Tissue Morphology
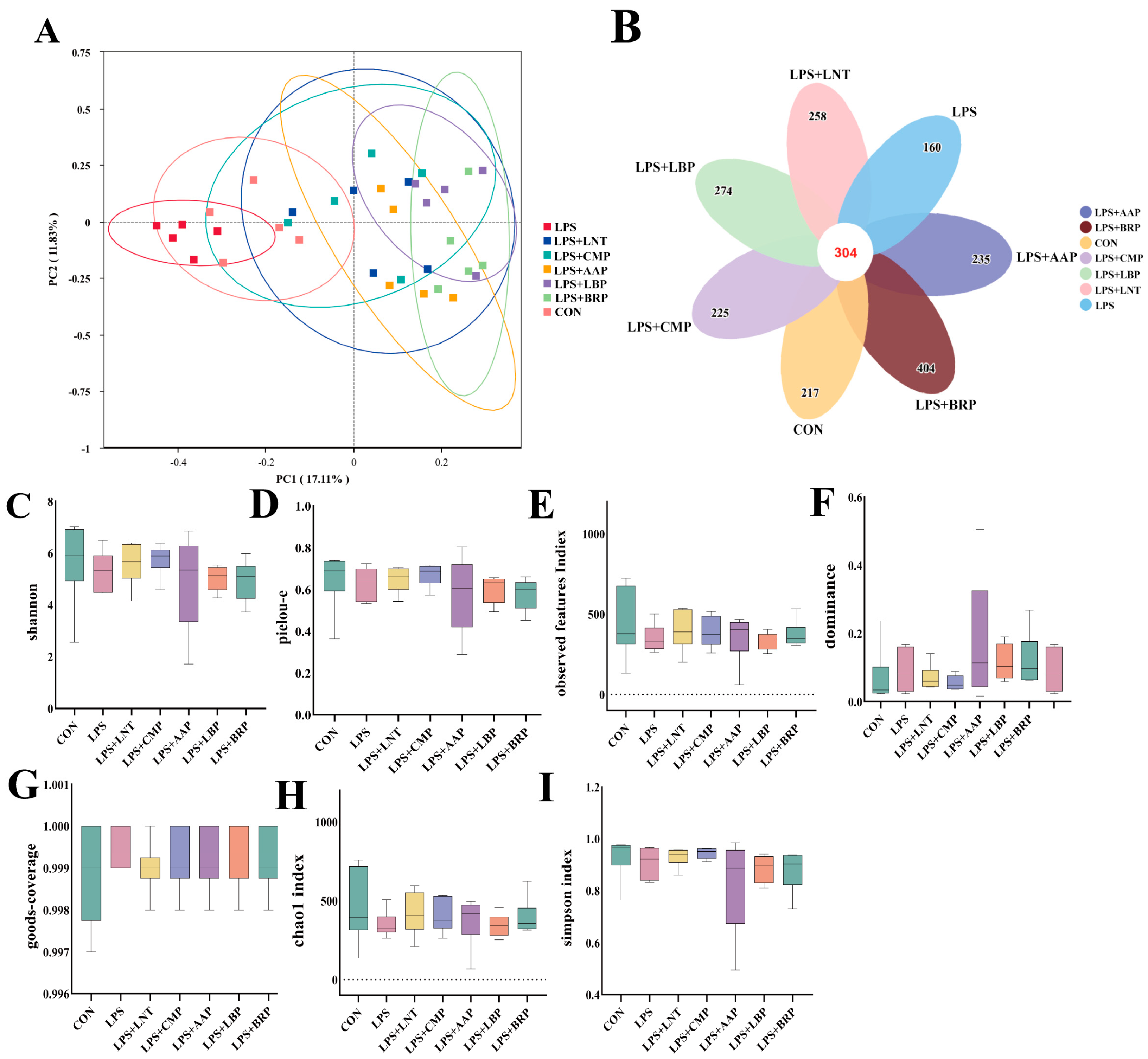
3.8. Analysis of Diversity of Cecal Flora by Five Polysaccharides
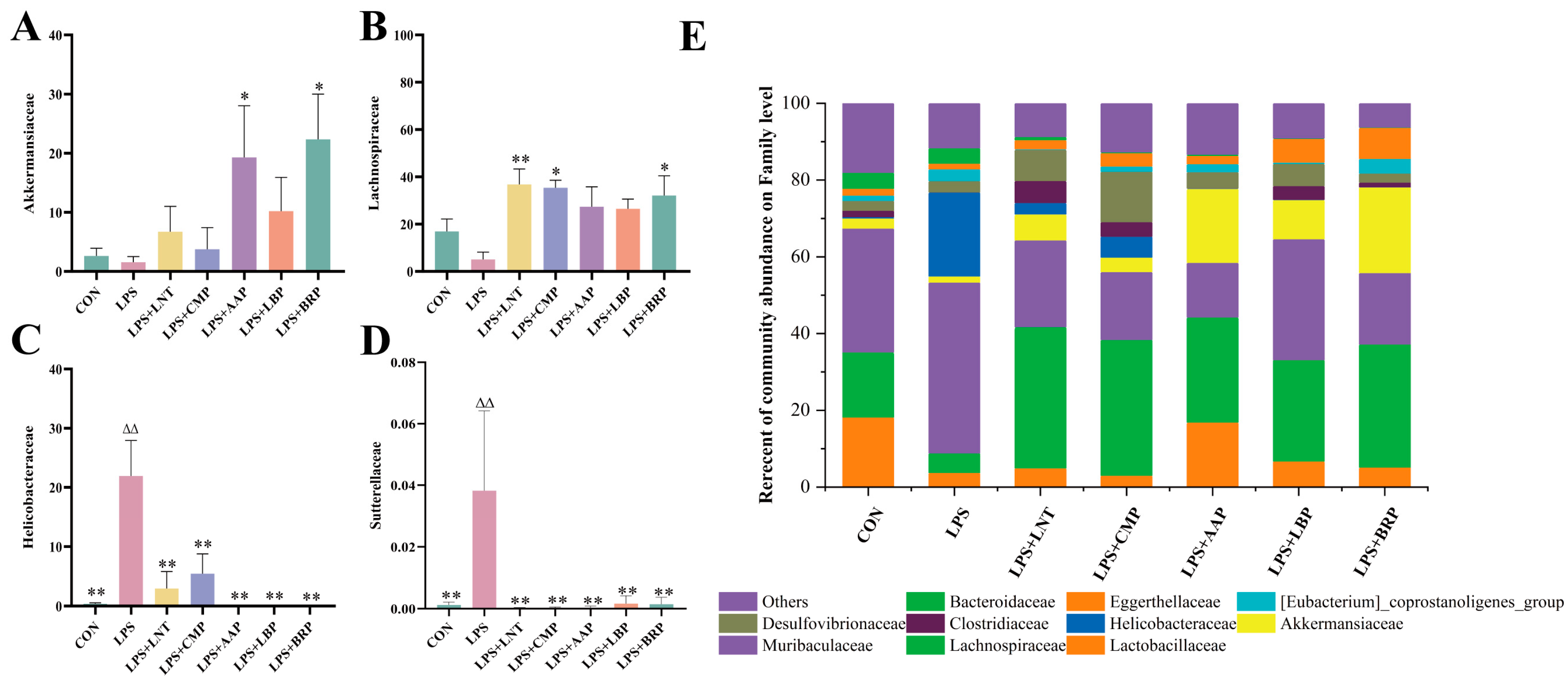
3.9. Effect of Five Polysaccharides on the Microbial Composition of Mouse Cecum
3.10. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tinazzi, M.; Sacilotto, A.; Cocetta, V.; Giacomini, I.; Raso, F.; Bulferi, G.; De Togni, H.; Lanza, R.; Consolo, P.; Berretta, M.; et al. Bowel Inflammation and Nutrient Supplementation: Effects of a Fixed Combination of Probiotics, Vitamins, and Herbal Extracts in an In Vitro Model of Intestinal Epithelial Barrier Dysfunction. Yale J. Biol. Med. 2024, 97, 297–308. [Google Scholar] [CrossRef]
- Gold, S.L.; Raman, M. Malnutrition Assessment in Patients with Inflammatory Bowel Disease. Can. Ibd Today 2023, 1, 32–38. [Google Scholar] [CrossRef]
- Hyder, C.S.; Uddin, M.A.; Jalal, M.T.; Karim, S.S.; Taher, M.A.; Nahar, K.; Hossain, M.S.; Sheikh, M.S.H.; Islam, M.S. Frequency and Pattern of Premalignant and Malignant Lesions among Fecal Immunochemical Test Positive Patients. Mymensingh Med. J. 2025, 34, 745–751. [Google Scholar]
- Giri, B.; Holubar, S.D.; Liska, D.; Lavryk, O.; Cohen, B.L.; Valente, M.A.; Steele, S.R.; Duraes, L.C. Biologic Therapy Is Associated with Improved Oncologic Outcomes in Crohn’s Disease-Associated Colorectal Cancer. Dis. Colon. Rectum. 2025, 68, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Roy, V.; Kunhipurayil, S.; Patel, K.M.; Elassar, S.; Sojitra, V.; Desai, H.D. Trends of Inflammatory Bowel Disease Burden in the United States (1990–2019), Projections of Deaths to 2040: Insights from the 2019 Global Burden of Disease Study. Inflamm. Bowel Dis. 2024, 30, S41–S42. [Google Scholar] [CrossRef]
- Qin, Z.; Chu, Q.; Ding, A.; Li, C.; Zhang, M. Sirtuin 3 ameliorates inflammatory bowel disease via inhibiting intestinal inflammation and oxidative stress. J. Clin. Biochem. Nutr. 2024, 74, 235–244. [Google Scholar] [CrossRef]
- Huang, W.N.; Lin, C.C. AB0079 Theaflavin Alleviates Collagen-Induced Arthritis in Mice by Decreasing Reactive Oxygen Species and Pro-Inflammatory Cytokines. Ann. Rheum. Dis. 2023, 82, 1218. [Google Scholar] [CrossRef]
- Cherukury, H.M.; Argueta, D.A.; Garcia, N.; Fouda, R.; Kiven, S.B.; Velasco, G.; Gupta, K. Cannabidiol Attenuates Chronic and Acute Hyperalgesia in Sickle Mice by Reducing Inflammation and Oxidative Stress. Blood 2022, 140, 8226–8227. [Google Scholar] [CrossRef]
- Bezerra, F.S.; Lanzetti, M.; Nesi, R.T.; Nagato, A.C.; Silva, C.P.; Kennedy-Feitosa, E.; Melo, A.C.; Cattani-Cavalieri, I.; Porto, L.C.; Valenca, S.S. Oxidative Stress and Inflammation in Acute and Chronic Lung Injuries. Antioxidants 2023, 12, 548. [Google Scholar] [CrossRef]
- Taherkhani, S.; Suzuki, K.; Castell, L. A Short Overview of Changes in Inflammatory Cytokines and Oxidative Stress in Response to Physical Activity and Antioxidant Supplementation. Antioxidants 2020, 9, 886. [Google Scholar] [CrossRef]
- Wu, X.; Ma, Y.; Li, X.; He, N.; Zhang, T.; Liu, F.; Feng, H.; Dong, J. Molecular mechanism of kidney damage caused by abamectin in carp: Oxidative stress, inflammation, mitochondrial damage, and apoptosis. Toxicology 2023, 494, 153599. [Google Scholar] [CrossRef]
- López-Moreno, A.; Carbonne, C.; Kropp, C.; Rios-Covian, D.; Pepke, F.; Langella, P.; Aguilera, M.; Martin, R. Characterisation of potential anti-inflammatory next-generation probiotics resistant to bisphenol A. Benef. Microbes. 2024, 16, 91–107. [Google Scholar] [CrossRef]
- Pertiwi, R.B.; Setiabudi, Y.C.; Mayangsari, Y.; Suroto, D.A.; Rahayu, E.S. Probiotic Lactiplantibacillus plantarum subsp. plantarum Dad-13 Alleviates 2, 4, 6-Trinitrobenzene Sulfonic Acid-Induced Colitis Through Short-Chain Fatty Acid Production and Inflammatory Cytokine Regulation. Prev. Nutr. Food Sci. 2024, 29, 270–278. [Google Scholar] [CrossRef]
- Ning, S.; Zhang, Z.; Zhou, C.; Wang, B.; Liu, Z.; Feng, B. Cross-talk between macrophages and gut microbiota in inflammatory bowel disease: A dynamic interplay influencing pathogenesis and therapy. Front. Med. 2024, 11, 1457218. [Google Scholar] [CrossRef]
- Li, L.; Liu, T.; Gu, Y.; Wang, X.; Xie, R.; Sun, Y.; Wang, B.; Cao, H. Regulation of gut microbiota-bile acids axis by probiotics in inflammatory bowel disease. Front. Immunol. 2022, 13, 974305. [Google Scholar] [CrossRef]
- Kunst, C.; Schmid, S.; Michalski, M.; Tümen, D.; Buttenschön, J.; Müller, M.; Gülow, K. The Influence of Gut Microbiota on Oxidative Stress and the Immune System. Biomedicines 2023, 11, 1388. [Google Scholar] [CrossRef]
- Li, R.; Yang, P.; Liu, B.; Ye, Z.; Zhang, P.; Li, M.; Gong, Y.; Huang, Y.; Yang, L.; Li, M. Lycium barbarum polysaccharide remodels colon inflammatory microenvironment and improves gut health. Heliyon 2024, 10, e30594. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, M.; Li, W.; Ma, L.; Liu, X.; Ding, Q.; Yu, W.; Yu, T.; Ding, C.; Liu, W. Research progress of natural plant polysaccharides inhibiting inflammatory signaling pathways and regulating intestinal flora and metabolism to protect inflammatory bowel disease. Int. J. Biol. Macromol. 2023, 253, 126799. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, J.; Zhao, Y.; Zong, S.; Tian, Y.; Chen, S.; Li, M.; Liu, H.; Zhang, Q.; Jing, X.; et al. Therapeutic effects of lentinan on inflammatory bowel disease and colitis-associated cancer. J. Cell Mol. Med. 2019, 23, 750–760. [Google Scholar] [CrossRef]
- Hou, C.; Chen, L.; Yang, L.; Ji, X. An insight into anti-inflammatory effects of natural polysaccharides. Int. J. Biol. Macromol. 2020, 153, 248–255. [Google Scholar] [CrossRef]
- Huo, J.; Wu, Z.; Sun, W.; Wang, Z.; Wu, J.; Huang, M.; Wang, B.; Sun, B. Protective Effects of Natural Polysaccharides on Intestinal Barrier Injury: A Review. J. Agric. Food Chem. 2022, 70, 711–735. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, W.; Wu, E.; Wang, K.; Chen, X.; Cui, Y.; Zhang, G.; Lv, F.; Wang, Y.; Peng, X.; et al. Polysaccharides From Abrus cantoniensis Hance Modulate Intestinal Microflora and Improve Intestinal Mucosal Barrier and Liver Oxidative Damage Induced by Heat Stress. Front. Vet. Sci. 2022, 9, 868433. [Google Scholar] [CrossRef]
- Kuang, Z.; Jin, T.; Wu, C.; Zong, Y.; Yin, P.; Dong, W.; Lin, X.; You, W.; Zhang, C.; Wang, L.; et al. Lentinan Attenuates Damage of the Small Intestinal Mucosa, Liver, and Lung in Mice with Gut-Origin Sepsis. J. Immunol. Res. 2021, 2021, 2052757. [Google Scholar] [CrossRef]
- Mongmonsin, U.; Rungsa, P.; Teajaroen, W.; Somdee, T.; Daduang, J.; Daduang, S. Effect of Cordyceps militaris extract on T-lymphocyte, Th2, and Th17 cytokines in vitro and in vivo. Asian J. Agric. Biol. 2024. [Google Scholar] [CrossRef]
- Li, W.; Gao, M.; Han, T. Lycium barbarum polysaccharides ameliorate intestinal barrier dysfunction and inflammation through the MLCK-MLC signaling pathway in Caco-2 cells. Food Funct. 2020, 11, 3741–3748. [Google Scholar] [CrossRef]
- Gao, Z.; Hu, N.; Cao, P.; Cheng, J.; Qiu, W.; Wang, Q.; Chen, X.; Chang, C.; Cai, C.; Meng, Y.; et al. The sol–gel property of a triple-helix polysaccharide from Auricularia auricula-judae and the protective effect on DSS-induced ulcerative colitis. Food Front. 2024, 5, 867–879. [Google Scholar] [CrossRef]
- Liu, T.; Wang, S.; Yan, J.; Ma, Y.; Wang, Z.; Zhu, J.; Chen, S.; Gao, M.Y.; Shen, H.; Wang, J. Effects of Qiamagu (Brassica rapa L.) polysaccharide on growth performance, immunity and gut health of yellow-feathered broilers. BMC Vet. Res. 2025, 21, 127. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, J.; Hao, H. Antitumor immunostimulatory activity of the traditional Chinese medicine polysaccharide on hepatocellular carcinoma. Front. Immunol. 2024, 15, 1369110. [Google Scholar] [CrossRef]
- Li, X.; Zhu, R.; Liu, Q.; Sun, H.; Sheng, H.; Zhu, L. Effects of traditional Chinese medicine polysaccharides on chronic diseases by modulating gut microbiota: A review. Int. J. Biol. Macromol. 2024, 282, 136691. [Google Scholar] [CrossRef]
- Guo, Y.; Wei, Y.; Liu, C.; Li, H.; Du, X.; Meng, J.; Liu, J.; Li, Q. Elucidation of antioxidant activities of intracellular and extracellular polysaccharides from Cordyceps militaris in vitro and their protective effects on ulcerative colitis in vivo. Int. J. Biol. Macromol. 2024, 267, 131385. [Google Scholar] [CrossRef]
- Song, Y.; Feng, Y.; Liu, G.; Duan, Y.; Zhang, H. Research progress on edible mushroom polysaccharides as a novel therapeutic strategy for inflammatory bowel disease. Int. J. Biol. Macromol. 2025, 305, 140994. [Google Scholar] [CrossRef]
- Fan, S.; Nie, S.; Huang, X.; Wang, S.; Hu, J.; Xie, J.; Nie, Q.; Xie, M. Protective properties of combined fungal polysaccharides from Cordyceps sinensis and Ganoderma atrum on colon immune dysfunction. Int. J. Biol. Macromol. 2018, 114, 1049–1055. [Google Scholar] [CrossRef]
- Ji, X.; Su, L.; Zhang, P.; Yue, Q.; Zhao, C.; Sun, X.; Li, K.; Liu, X.; Zhang, S.; Zhao, L. Lentinan improves intestinal inflammation and gut dysbiosis in antibiotics-induced mice. Sci. Rep. 2022, 12, 19609. [Google Scholar] [CrossRef]
- Zhou, W.; Kan, X.; Chen, G.; Sun, Y.; Ran, L.; Yan, Y.; Mi, J.; Lu, L.; Zeng, X.; Cao, Y. The polysaccharides from the fruits of Lycium barbarum L. modify the gut community profile and alleviate dextran sulfate sodium-induced colitis in mice. Int. J. Biol. Macromol. 2022, 222, 2244–2257. [Google Scholar] [CrossRef]
- Islam, T.; Xu, B.; Bian, Z. Anti-inflammatory and gut microbiota regulatory effects of ultrasonic degraded polysaccharides from Auricularia auricula-judae in DSS-induced colitis mice. Ultrason. Sonochem. 2025, 117, 107339. [Google Scholar] [CrossRef]
- Song, S.; Zhang, X.; Zheng, H.; Liao, Y.; Tang, P.; Liu, Y.; Tang, A.; Ran, P.; Sun, X.; Yang, P. Intermittent Fasting Reduces Intestinal Inflammation in Dextran Sulfate Sodium-Induced Colitis of Mice. Food Sci. Nutr. 2025, 13, e70014. [Google Scholar] [CrossRef]
- Anhê, F.F.; Barra, N.G.; Cavallari, J.F.; Henriksbo, B.D.; Schertzer, J.D. Metabolic endotoxemia is dictated by the type of lipopolysaccharide. Cell Rep. 2021, 36, 109691. [Google Scholar] [CrossRef]
- Wan, F.; Wang, M.; Zhong, R.; Chen, L.; Han, H.; Liu, L.; Zhao, Y.; Lv, H.; Hou, F.; Yi, B.; et al. Supplementation With Chinese Medicinal Plant Extracts From Lonicera hypoglauca and Scutellaria baicalensis Mitigates Colonic Inflammation by Regulating Oxidative Stress and Gut Microbiota in a Colitis Mouse Model. Front. Cell Infect. Microbiol. 2021, 11, 798052. [Google Scholar] [CrossRef]
- Sánchez-Bonet, D.; Padrón-Sanz, C.; Lloris-Cejalvo, J.M.; Lloris-Carsí, J.M.; Cejalvo-Lapeña, D. Antioxidant Effect of a Fucus vesiculosus Extract on Intestinal Ischemia/Reperfusion Injury in Rats: A Biochemical and Histological Study. Antioxidants 2025, 14, 624. [Google Scholar] [CrossRef]
- Hao, W.; Chen, Z.; Yuan, Q.; Ma, M.; Gao, C.; Zhou, Y.; Zhou, H.; Wu, X.; Wu, D.; Farag, M.A.; et al. Ginger polysaccharides relieve ulcerative colitis via maintaining intestinal barrier integrity and gut microbiota modulation. Int. J. Biol. Macromol. 2022, 219, 730–739. [Google Scholar] [CrossRef]
- Wan, L.; Qian, C.; Yang, C.; Peng, S.; Dong, G.; Cheng, P.; Zong, G.; Han, H.; Shao, M.; Gong, G.; et al. Ginseng polysaccharides ameliorate ulcerative colitis via regulating gut microbiota and tryptophan metabolism. Int. J. Biol. Macromol. 2024, 265, 130822. [Google Scholar] [CrossRef]
- Wu, S.; Chen, X.; Cai, R.; Chen, X.; Zhang, J.; Xie, J.; Shen, M. Sulfated Chinese Yam Polysaccharides Alleviate LPS-Induced Acute Inflammation in Mice through Modulating Intestinal Microbiota. Foods 2023, 12, 1772. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Yang, D.; Xiao, J.; Hong, W.; Sun, H.; Xie, Q.; Zeng, C. Artemisia argyi polysaccharide alleviates osmotic diarrhea by enhancing intestinal barrier protection and anti-inflammation. Int. J. Biol. Macromol. 2024, 282, 136779. [Google Scholar] [CrossRef] [PubMed]
- Jędrzejewski, T.; Pawlikowska, M.; Piotrowski, J.; Kozak, W. Protein-bound polysaccharides from Coriolus versicolor attenuate LPS-induced synthesis of pro-inflammatory cytokines and stimulate PBMCs proliferation. Immunol. Lett. 2016, 178, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, D.; Zhao, F.; Yang, Z.; Wang, D.; Qiao, M.; Fang, Y.; Li, W.; Wu, R.; He, P.; et al. Quantification of Dynamic Protein Interactions and Phosphorylation in LPS Signaling Pathway by SWATH-MS. Mol. Cell Proteom. 2019, 18, 1054–1069. [Google Scholar] [CrossRef]
- Chen, X.; Wu, J.; Fu, X.; Wang, P.; Chen, C. Fructus mori polysaccharide alleviates diabetic symptoms by regulating intestinal microbiota and intestinal barrier against TLR4/NF-κB pathway. Int. J. Biol. Macromol. 2023, 249, 126038. [Google Scholar] [CrossRef]
- Zheng, J.; Gong, S.; Han, J. Arabinogalactan Alleviates Lipopolysaccharide-Induced Intestinal Epithelial Barrier Damage through Adenosine Monophosphate-Activated Protein Kinase/Silent Information Regulator 1/Nuclear Factor Kappa-B Signaling Pathways in Caco-2 Cells. Int. J. Mol. Sci. 2023, 24, 15337. [Google Scholar] [CrossRef]
- Lian, Y.Z.; Chang, C.; Chen, Y.; Tinkov, A.A.; Skalny, A.V.; Chao, J.C. Lycium barbarum polysaccharides and capsaicin modulate inflammatory cytokines and colonic microbiota in colitis rats induced by dextran sulfate sodium. J. Clin. Biochem. Nutr. 2022, 71, 229–237. [Google Scholar] [CrossRef]
- Chen, S.; Wang, J.; Dong, N.; Fang, Q.; Zhang, Y.; Chen, C.; Cui, S.W.; Nie, S. Polysaccharides from natural Cordyceps sinensis attenuated dextran sodium sulfate-induced colitis in C57BL/6J mice. Food Funct. 2023, 14, 720–733. [Google Scholar] [CrossRef]
- Wang, J.; Gao, H.; Xie, Y.; Wang, P.; Li, Y.; Zhao, J.; Wang, C.; Ma, X.; Wang, Y.; Mao, Q.; et al. Lycium barbarum polysaccharide alleviates dextran sodium sulfate-induced inflammatory bowel disease by regulating M1/M2 macrophage polarization via the STAT1 and STAT6 pathways. Front. Pharmacol. 2023, 14, 1044576. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.; Ma, M.; Zhang, J.; Liu, M.; Mosenthin, R.; Zhao, L.; Huang, S.; Ma, Q. Dietary arabinogalactan modulates immunity and improves gut barrier in broilers via regulating metabolome and gut microbiome. Carbohydr. Polym. 2025, 352, 123223. [Google Scholar] [CrossRef]
- Zhao, T.; Liu, S.; Ma, X.; Shuai, Y.; He, H.; Guo, T.; Huang, W.; Wang, Q.; Liu, S.; Wang, Z.; et al. Lycium barbarum arabinogalactan alleviates intestinal mucosal damage in mice by restoring intestinal microbes and mucin O-glycans. Carbohydr. Polym. 2024, 330, 121882. [Google Scholar] [CrossRef] [PubMed]
- Maity, P.; Sen, I.K.; Chakraborty, I.; Mondal, S.; Bar, H.; Bhanja, S.K.; Mandal, S.; Maity, G.N. Biologically active polysaccharide from edible mushrooms: A review. Int. J. Biol. Macromol. 2021, 172, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Huang, X.; Wen, J.; Chen, S.; Wu, X.; Ma, W.; Cui, S.W.; Xie, M.; Nie, S. Innate immune receptors co-recognition of polysaccharides initiates multi-pathway synergistic immune response. Carbohydr. Polym. 2023, 305, 120533. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Tang, X.; Shuai, X.; Jiang, C.; Liu, X.; Wang, L.; Yao, Y.; Nie, S.; Xie, M. Mannose Receptor Mediates the Immune Response to Ganoderma atrum Polysaccharides in Macrophages. J. Agric. Food Chem. 2017, 65, 348–357. [Google Scholar] [CrossRef]
- Jiang, S.; Yin, H.; Li, R.; Shi, W.; Mou, J.; Yang, J. The activation effects of fucoidan from sea cucumber Stichopus chloronotus on RAW264.7 cells via TLR2/4-NF-κB pathway and its structure-activity relationship. Carbohydr. Polym. 2021, 270, 118353. [Google Scholar] [CrossRef]
- Nguepi Tsopmejio, I.S.; Yuan, J.; Diao, Z.; Fan, W.; Wei, J.; Zhao, C.; Li, Y.; Song, H. Auricularia polytricha and Flammulina velutipes reduce liver injury in DSS-induced Inflammatory Bowel Disease by improving inflammation, oxidative stress, and apoptosis through the regulation of TLR4/NF-κB signaling pathways. J. Nutr. Biochem. 2023, 111, 109190. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Escrivá, C.; Dromant, M.; Borrás, C.; Viña, J. Lipid peroxidation as measured by chromatographic determination of malondialdehyde. Human plasma reference values in health and disease. Arch. Biochem. Biophys. 2021, 709, 108941. [Google Scholar] [CrossRef]
- Cai, Y.; Liu, S.; Ge, X.; Cheng, L.; Zhang, X. Inhibitory effect of tea flower polysaccharides on oxidative stress and microglial oxidative damage in aging mice by regulating gut microbiota. Food Funct. 2024, 15, 11444–11457. [Google Scholar] [CrossRef]
- Ji, X.; Hou, H.; Wang, X.; Qiu, Y.; Ma, Y.; Wang, S.; Guo, S.; Huang, S.; Zhang, C. Effect of dietary Glycyrrhiza polysaccharides on growth performance, hepatic antioxidant capacity and anti-inflammatory capacity of broiler chickens. Res. Vet. Sci. 2024, 167, 105114. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Xue, J.; Yu, C.; Tang, Y.; Mo, W.; Tang, Z.; Sheng, W.; Jiao, Y.; Zhu, W.; Cao, J. NF-E2-Related Factor 2 (Nrf2) Ameliorates Radiation-Induced Skin Injury. Front. Oncol. 2021, 11, 680058. [Google Scholar] [CrossRef]
- Ju, S.; Singh, M.K.; Han, S.; Ranbhise, J.; Ha, J.; Choe, W.; Yoon, K.; Yeo, S.G.; Kim, S.S.; Kang, I. Oxidative Stress and Cancer Therapy: Controlling Cancer Cells Using Reactive Oxygen Species. Int. J. Mol. Sci. 2024, 25, 12387. [Google Scholar] [CrossRef]
- Fan, X.; Xiao, X.; Yu, W.; Yu, B.; He, J.; Zheng, P.; Yu, J.; Luo, J.; Luo, Y.; Yan, H.; et al. Yucca schidigera purpurea-sourced arabinogalactan polysaccharides augments antioxidant capacity facilitating intestinal antioxidant functions. Carbohydr. Polym. 2024, 326, 121613. [Google Scholar] [CrossRef]
- Jiang, S.; Xiao, X.; Li, J.; Mu, Y. Lycium barbarum polysaccharide-glycoprotein ameliorates ionizing radiation-induced epithelial injury by regulating oxidative stress and ferroptosis via the Nrf2 pathway. Free Radic. Biol. Med. 2023, 204, 84–94. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, Y.; Feng, Y.; Jiao, F.; Jia, L. Lentinus edodes Polysaccharides Alleviate Acute Lung Injury by Inhibiting Oxidative Stress and Inflammation. Molecules 2022, 27, 7328. [Google Scholar] [CrossRef]
- Wen, C.; Ye, Z.; Liu, G.; Liang, L.; Liu, X.; Li, Y.; Xu, X.; Zhang, J. Isolation, Purification, and Characterization of Lentinus edodes Polysaccharides Extracted With Subcritical Water Enhanced With Deep Eutectic Solvent. Chem. Biodivers. 2025, 22, e202402658. [Google Scholar] [CrossRef]
- Liu, X.; Huang, L.; Zhang, X.; Xu, X. Polysaccharides with antioxidant activity: Extraction, beneficial roles, biological mechanisms, structure-function relationships, and future perspectives: A review. Int. J. Biol. Macromol. 2025, 300, 140221. [Google Scholar] [CrossRef]
- Yoon, J.W.; Shin, S.; Park, J.; Lee, B.R.; Lee, S.I. TLR/MyD88-Mediated Inflammation Induced in Porcine Intestinal Epithelial Cells by Ochratoxin a Affects Intestinal Barrier Function. Toxics 2023, 11, 437. [Google Scholar] [CrossRef]
- Saito, A.C.; Endo, C.; Fukazawa, Y.; Higashi, T.; Chiba, H. Effects of TAMP family on the tight junction strand network and barrier function in epithelial cells. Ann. N. Y. Acad. Sci. 2022, 1517, 234–250. [Google Scholar] [CrossRef]
- Kuo, W.; Zuo, L.; Odenwald, M.A.; Madha, S.; Singh, G.; Gurniak, C.B.; Abraham, C.; Turner, J.R. The Tight Junction Protein ZO-1 Is Dispensable for Barrier Function but Critical for Effective Mucosal Repair. Gastroenterology 2021, 161, 1924–1939. [Google Scholar] [CrossRef]
- Yu, J.; Zhao, J.; Xie, H.; Cai, M.; Yao, L.; Li, J.; Han, L.; Chen, W.; Yu, N.; Peng, D. Dendrobium huoshanense polysaccharides ameliorate ulcerative colitis by improving intestinal mucosal barrier and regulating gut microbiota. J. Funct. Foods 2022, 96, 105231. [Google Scholar] [CrossRef]
- Luo, L.; Liu, Y.; Cai, X.; Wang, Y.; Xue, J.; Zhang, J.; Yang, F. Bletilla striata polysaccharides ameliorates lipopolysaccharide-induced injury in intestinal epithelial cells. Saudi J. Gastroenterol. 2019, 25, 302–308. [Google Scholar] [CrossRef]
- Bhadoriya, P.; Jatley, A.; Singh, A.; Mehrotra, R.; Jain, M.; Mohammed, A.; Hamzah, Z.; Bukke, S.P.N. Exploring gut microbiota’s influence on cognitive health and neurodegenerative disorders: Mechanistic insights and therapeutic approaches. Discov. Immun. 2025, 2, 3. [Google Scholar] [CrossRef]
- Zhao, Z.; Tong, Y.; Kang, Y.; Qiu, Z.; Li, Q.; Xu, C.; Wu, G.; Jia, W.; Wang, P. Sodium butyrate (SB) ameliorated inflammation of COPD induced by cigarette smoke through activating the GPR43 to inhibit NF-κB/MAPKs signaling pathways. Mol. Immunol. 2023, 163, 224–234. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, R.; Yu, J.; Bao, T.; Liu, Y.; Bai, Z.; Wang, T.; Li, Y.; Zhang, X.; Wang, H.; et al. Butyrate ameliorates inflammation of alcoholic liver disease by suppressing the LPS-TLR4-NF-κB/NLRP3 axis via binding GPR43-β-arrestin2. J. Funct. Foods 2022, 99, 105351. [Google Scholar] [CrossRef]
- Luo, Q.; Sun, M.; Guo, Y.; Tan, S.; Wu, X.; Abassa, K.; Lin, L.; Liu, H.; Jiang, J.; Wei, X. Sodium butyrate protects against lipopolysaccharide-induced liver injury partially via the GPR43/ β-arrestin-2/NF-κB network. Gastroenterol. Rep. 2021, 9, 154–165. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, M.; Wang, X.; Ren, Y.; Yue, T.; Wang, Z.; Gao, Z. Edible fungal polysaccharides, the gut microbiota, and host health. Carbohydr. Polym. 2021, 273, 118558. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, Q.; Liu, Y.; Wei, Y.; He, S.; Lin, W.; Wang, Y.; Li, L.; Xu, Y. Vancomycin-induced gut microbiota dysbiosis aggravates allergic rhinitis in mice by altered short-chain fatty acids. Front. Microbiol. 2022, 13, 1002084. [Google Scholar] [CrossRef]
- Rodriguez-Palacios, A.; Harding, A.; Menghini, P.; Himmelman, C.; Retuerto, M.; Nickerson, K.P.; Lam, M.; Croniger, C.M.; McLean, M.H.; Durum, S.K.; et al. The Artificial Sweetener Splenda Promotes Gut Proteobacteria, Dysbiosis, and Myeloperoxidase Reactivity in Crohn’s Disease-Like Ileitis. Inflamm. Bowel Dis. 2018, 24, 1005–1020. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, H.; Tu, X.; Gao, Z. The Role of Short-Chain Fatty Acids of Gut Microbiota Origin in Hypertension. Front. Microbiol. 2021, 12, 730809. [Google Scholar] [CrossRef]
- Strati, F.; Pujolassos, M.; Burrello, C.; Giuffrè, M.R.; Lattanzi, G.; Caprioli, F.; Troisi, J.; Facciotti, F. Antibiotic-associated dysbiosis affects the ability of the gut microbiota to control intestinal inflammation upon fecal microbiota transplantation in experimental colitis models. Microbiome 2021, 9, 39. [Google Scholar] [CrossRef]
- Xu, H.; Wang, S.; Jiang, Y.; Wu, J.; Chen, L.; Ding, Y.; Zhou, Y.; Deng, L.; Chen, X. Poria cocos Polysaccharide Ameliorated Antibiotic-Associated Diarrhea in Mice via Regulating the Homeostasis of the Gut Microbiota and Intestinal Mucosal Barrier. Int. J. Mol. Sci. 2023, 24, 1423. [Google Scholar] [CrossRef]
- Hiippala, K.; Kainulainen, V.; Kalliomäki, M.; Arkkila, P.; Satokari, R. Mucosal Prevalence and Interactions with the Epithelium Indicate Commensalism of Sutterella spp. Front. Microbiol. 2016, 7, 1706. [Google Scholar]
- Jauvain, M.; Courtois, S.; Lehours, P.; Bessède, E. Metformin Modifies the Gut Microbiota of Mice Infected with Helicobacter pylori. Pharmaceuticals 2021, 14, 329. [Google Scholar] [CrossRef]
- Li, S.; Wu, D.; Cao, M.; Yu, Z.; Wu, M.; Liu, Y.; Zhou, J.; Yan, S.; Chen, J.; Huang, M.; et al. Effects of choline supplementation on liver biology, gut microbiota, and inflammation in Helicobacter pylori-infected mice. Life Sci. 2020, 259, 118200. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, F.; Mao, L.; Feng, T.; Wang, K.; Xu, M.; Lv, B.; Wang, X. Bifico relieves irritable bowel syndrome by regulating gut microbiota dysbiosis and inflammatory cytokines. Eur. J. Nutr. 2023, 62, 139–155. [Google Scholar] [CrossRef]
- Yu, Z.; Cao, M.; Peng, J.; Wu, D.; Li, S.; Wu, C.; Qing, L.; Zhang, A.; Wang, W.; Huang, M.; et al. Lacticaseibacillus casei T1 attenuates Helicobacter pylori-induced inflammation and gut microbiota disorders in mice. BMC Microbiol. 2023, 23, 39. [Google Scholar] [CrossRef]
- Fayyaz, I.; Zahoor, M.A.; Shahid, M.; Rasool, M.H.; Nawaz, Z. Effect of Lactobacillus casei on serum interleukins following enteropathogenic E. coli infection in experimental rabbits. Pak. J. Pharm. Sci. 2018, 31, 2131–2136. [Google Scholar]
- Huang, Q.; Liang, J.; Yang, C.; Li, K.; Niu, M.; Fan, J.; Zhang, X. Stimulation-responsive mucoadhesive probiotics for inflammatory bowel disease treatment by scavenging reactive oxygen species and regulating gut microbiota. Biomaterials 2023, 301, 122274. [Google Scholar] [CrossRef]
- Zhou, R.; Wang, Y.; Wang, Z.; Liu, K.; Wang, Q.; Bao, H. Effects of Auricularia auricula-judae polysaccharide on pasting, gelatinization, rheology, structural properties and in vitro digestibility of kidney bean starch. Int. J. Biol. Macromol. 2021, 191, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Kang, Y. Synergistic interaction of Auricularia auricula-judae polysaccharide with yam starch: Effects on physicochemical properties and in vitro starch digestibility. Food Sci. Biotechnol. 2018, 27, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Chaves, L.S.; Oliveira, A.C.P.; Oliveira, A.P.; Lopes, A.L.F.; Araujo, A.K.S.; Pacheco, G.; Silva, K.C.; Martins, F.E.C.; Gomes, I.A.B.; Ramos, S.V.S.; et al. Cashew gum fractions protect intestinal mucosa against shiga toxin-producing Escherichia coli infection: Characterization and insights into microbiota modulation. Int. J. Biol. Macromol. 2025, 311, 143916. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Li, J.; Zhang, S.; Zhang, Q.; Duan, J.; Guo, J. Abelmoschus manihot polysaccharide fortifies intestinal mucus barrier to alleviate intestinal inflammation by modulating Akkermansia muciniphila abundance. Acta Pharm. Sin. B 2024, 14, 3901–3915. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, J.; Yi, K.; Peng, L.; Xie, J.; Gou, X.; Peng, T.; Tang, L. Phlorizin ameliorates obesity-associated endotoxemia and insulin resistance in high-fat diet-fed mice by targeting the gut microbiota and intestinal barrier integrity. Gut Microbes 2020, 12, 1842990. [Google Scholar] [CrossRef]
- Liu, M.; Yang, J.; Yan, Z.; Hu, S.; Li, J.; Xu, Z.; Jian, Y. Recent findings in Akkermansia muciniphila-regulated metabolism and its role in intestinal diseases. Clin. Nutr. 2022, 41, 2333–2344. [Google Scholar] [CrossRef]
- Xi, M.; Li, J.; Hao, G.; An, X.; Song, Y.; Wei, H.; Ge, W. Stachyose increases intestinal barrier through Akkermansia muciniphila and reduces gut inflammation in germ-free mice after human fecal transplantation. Food Res. Int. 2020, 137, 109288. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Z.; Gui, Z.; Yang, B.; You, F.; Yang, G.; Zhang, X.; Chang, X.; Meng, X. Supplementation with Akkermansia muciniphila improved intestinal barrier and immunity in zebrafish (Danio rerio). Fish Shellfish Immunol. 2024, 154, 109935. [Google Scholar] [CrossRef]
- Kotsiliti, E. Phage therapy suppresses gut inflammation in IBD. Nat. Biotechnol. 2022, 40, 1327. [Google Scholar] [CrossRef]
- Liu, S.; Xu, A.; Gao, Y.; Xie, Y.; Liu, Z.; Sun, M.; Mao, H.; Wang, X. Graphene oxide exacerbates dextran sodium sulfate-induced colitis via ROS/AMPK/p53 signaling to mediate apoptosis. J. Nanobiotechnol. 2021, 19, 85. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, S.; Zhang, W.; Cui, H.; Zhang, J.; Yin, X.; Zheng, X.; Shen, T.; Ying, H.; Chen, L.; et al. Cordycepin mitigates dextran sulfate sodium-induced colitis through improving gut microbiota composition and modulating Th1/Th2 and Th17/Treg balance. Biomed. Pharmacother. 2024, 180, 117394. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, M.; Xu, R.; Yu, Y.; Feng, H.; Li, D.; Li, L.; Zhang, B.; Liu, G.; Wang, Y.; et al. SDH, a novel diarylheptane compound, alleviates dextran sulfate sodium (DSS)-induced colitis by reducing Th1/Th2/Th17 induction and regulating the gut microbiota in mice. Int. Immunopharmacol. 2024, 134, 112234. [Google Scholar] [CrossRef] [PubMed]
- Mathur, R.; Alam, M.M.; Zhao, X.; Huang, Y.; Zhu, X. Mechanistic Insight into the Development of TNBS-Mediated Intestinal Fibrosis and Evaluating the Inhibitory Effects of Rapamycin. J. Vis. Exp. 2019, 12, e60067. [Google Scholar] [CrossRef]
- Chi, S.; Rong, L.; Zhang, M.; Li, Y.; Zhou, Y.; Li, X.; Zhang, X.; Luo, Z.; Li, S.; Cao, G.; et al. Biliary atresia: Rotavirus amplification of lipopolysaccharide/toll-like receptor 4 by mediating MMP7 upregulation through NF-κB. Pediatr. Res. 2025, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Cheng, A.; Qiu, M.; Zang, G. Allicin Improves Lung Injury Induced by Sepsis via Regulation of the Toll-Like Receptor 4 (TLR4)/Myeloid Differentiation Primary Response 88 (MYD88)/Nuclear Factor kappa B (NF-κB) Pathway. Med. Sci. Monitor. 2019, 25, 2567–2576. [Google Scholar] [CrossRef]
- Zhang, X.; Li, M.; Wang, H. Astragaloside IV Alleviates the Myocardial Damage Induced by Lipopolysaccharide via the Toll-Like Receptor 4 (TLR4)/Nuclear Factor kappa B (NF-κB)/Proliferator-Activated Receptor α (PPARα) Signaling Pathway. Med. Sci. Monitor. 2019, 25, 7158–7168. [Google Scholar] [CrossRef]
- Gampierakis, I.; Koutmani, Y.; Semitekolou, M.; Morianos, I.; Polissidis, A.; Katsouda, A.; Charalampopoulos, I.; Xanthou, G.; Gravanis, A.; Karalis, K.P. Hippocampal neural stem cells and microglia response to experimental inflammatory bowel disease (IBD). Mol. Psychiatry 2021, 26, 1248–1263. [Google Scholar] [CrossRef]
- Silva, I.; Solas, J.; Pinto, R.; Mateus, V. Chronic Experimental Model of TNBS-Induced Colitis to Study Inflammatory Bowel Disease. Int. J. Mol. Sci. 2022, 23, 4739. [Google Scholar] [CrossRef]
- Zhou, J.; Yan, X.; Bi, X.; Lu, S.; Liu, X.; Yang, C.; Shi, Y.; Luo, L.; Yin, Z. γ-Glutamylcysteine rescues mice from TNBS-driven inflammatory bowel disease through regulating macrophages polarization. Inflamm. Res. 2023, 72, 603–621. [Google Scholar] [CrossRef]
- Tam, J.S.Y.; Coller, J.K.; Hughes, P.A.; Prestidge, C.A.; Bowen, J.M. Toll-like receptor 4 (TLR4) antagonists as potential therapeutics for intestinal inflammation. Indian J. Gastroenterol. 2021, 40, 5–21. [Google Scholar] [CrossRef]
- Zhang, W.; He-Yang, J.; Tu, W.; Zhou, X. Sialylated human milk oligosaccharides prevent intestinal inflammation by inhibiting toll like receptor 4/NLRP3 inflammasome pathway in necrotizing enterocolitis rats. Nutr. Metab. 2021, 18, 5. [Google Scholar] [CrossRef]
- Xu, X.; Li, K.; Liu, Q.; Zhang, H.; Li, L. Epigallocatechin Gallate Alleviates Lipopolysaccharide-Induced Intestinal Inflammation in Wenchang Chicken by Inhibiting the TLR4/MyD88/NF-κB Signaling Pathway. Vet. Sci. 2025, 12, 225. [Google Scholar] [CrossRef]
- Hassan, N.F.; Nada, S.A.; Hassan, A.; El-Ansary, M.R.; Al-Shorbagy, M.Y.; Abdelsalam, R.M. Saroglitazar Deactivates the Hepatic LPS/TLR4 Signaling Pathway and Ameliorates Adipocyte Dysfunction in Rats with High-Fat Emulsion/LPS Model-Induced Non-alcoholic Steatohepatitis. Inflammation 2019, 42, 1056–1070. [Google Scholar] [CrossRef]
- Huang, J.; Li, J.; Geng, Z.; Yin, L.; Niu, M.; Li, Q.; Liu, X.; Cheng, X.; Zhang, X.; Song, X.; et al. Cynaroside ameliorates TNBS-induced colitis by inhibiting intestinal epithelial cell apoptosis via the PI3K/AKT signalling pathway. Front. Pharmacol. 2024, 15, 1496068. [Google Scholar]
- Zhang, Z.; Kong, L.; Lv, M.; Yao, Y.; Gao, L.; Zhou, R.; Ma, W.; Li, J. PVA enema ameliorates DSS-induced acute colitis in mice. BMC Gastroenterol. 2023, 23, 368. [Google Scholar] [CrossRef]
- Wen, X.; Wan, F.; Wu, Y.; Liu, L.; Liu, Y.; Zhong, R.; Chen, L.; Zhang, H. Caffeic acid supplementation ameliorates intestinal injury by modulating intestinal microbiota in LPS-challenged piglets. Food Funct. 2023, 14, 7705–7717. [Google Scholar] [CrossRef]
- Wen, X.; Zhong, R.; Dang, G.; Xia, B.; Wu, W.; Tang, S.; Tang, L.; Liu, L.; Liu, Z.; Chen, L.; et al. Pectin supplementation ameliorates intestinal epithelial barrier function damage by modulating intestinal microbiota in lipopolysaccharide-challenged piglets. J. Nutr. Biochem. 2022, 109, 109107. [Google Scholar] [CrossRef]
- Gautam, S.; Lapcik, L.; Lapcikova, B.; Repka, D.; Szyk-Warszyńska, L. Physicochemical Characterisation of Polysaccharide Films with Embedded Bioactive Substances. Foods 2023, 12, 4454. [Google Scholar] [CrossRef]
- Yang, Y.; Khan, B.M.; Zhang, X.; Zhao, Y.; Cheong, K.; Liu, Y. Advances in Separation and Purification of Bioactive Polysaccharides through High-speed Counter-Current Chromatography. J. Chromatogr. Sci. 2020, 58, 992–1000. [Google Scholar] [CrossRef]
- Cui, M.; Zhang, M.; Liu, K. Colon-targeted drug delivery of polysaccharide-based nanocarriers for synergistic treatment of inflammatory bowel disease: A review. Carbohydr. Polym. 2021, 272, 118530. [Google Scholar] [CrossRef]
- Pulvirenti, A.; Boccia, A.C.; Constantin, C.; Surcel, M.; Munteanu, A.; Peteu, V.; Neagu, M. Single-Component Starch-Based Hydrogels for Therapeutic Delivery. Molecules 2024, 29, 5463. [Google Scholar] [CrossRef]
- Jakubczyk, K.; Drużga, A.; Katarzyna, J.; Skonieczna-Żydecka, K. Antioxidant Potential of Curcumin-A Meta-Analysis of Randomized Clinical Trials. Antioxidants 2020, 9, 1092. [Google Scholar] [CrossRef]
- Caballero, S.; Li, Y.O.; McClements, D.J.; Davidov-Pardo, G. Encapsulation and delivery of bioactive citrus pomace polyphenols: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 8028–8044. [Google Scholar] [CrossRef] [PubMed]
- Periferakis, A.; Periferakis, A.; Troumpata, L.; Periferakis, K.; Georgatos-Garcia, S.; Touriki, G.; Dragosloveanu, C.D.; Caruntu, A.; Savulescu-Fiedler, I.; Dragosloveanu, S.; et al. Pinosylvin: A Multifunctional Stilbenoid with Antimicrobial, Antioxidant, and Anti-Inflammatory Potential. Curr. Issues Mol. Biol. 2025, 47, 204. [Google Scholar] [CrossRef] [PubMed]
- Dvorakova, M.; Landa, P. Anti-inflammatory activity of natural stilbenoids: A review. Pharmacol. Res. 2017, 124, 126–145. [Google Scholar] [CrossRef] [PubMed]
- Dalla Costa, V.; Piovan, A.; Brun, P.; Filippini, R. Morus alba L. Cell Cultures as Sources of Antioxidant and Anti-Inflammatory Stilbenoids for Food Supplement Development. Molecules 2025, 30, 2073. [Google Scholar] [CrossRef]
- Gao, J.; Hu, D.; Shen, Y.; Zheng, Y.; Liang, Y. Optimization of ultrasonic-assisted polysaccharide extraction from Hyperici Perforati Herba using response surface methodology and assessment of its antioxidant activity. Int. J. Biol. Macromol. 2023, 225, 255–265. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Primer Sequence(5′-3′) |
|---|---|
| TLR4 | F:GCACTGTTCTTCTCCTGCCT |
| R:AGAGGTGGTGTAAGCCATGC | |
| MyD88 | F:GCATGGTGGTGGTTGTTTCTG |
| R:GAATCAGTCGCTTCTGTTGG | |
| NF-κB | F:ACACTGGAAGCACGGATGAC |
| R:TGTCTGTGAGTTGCCGGTCT | |
| TNF-α | F:CAAAATTCGAGTGACAAGCC |
| R:TTGTCCCTTGAAGAGAACCT | |
| ZO-1 | F:GCCTAAAGCTGTTCCTGTGAGTCC |
| R:ACCCCGCCGTTGCTGTTAAAC | |
| Occludin | F:TGGCTGCCTTCTGCTTCATTGC |
| R:GAACACCATCACACCCAGGATAGC | |
| MUC2 | F:CCACAACGACTCCTACGCCATC |
| R:CGCTGCCGTCCGACTTGAAG | |
| β-actin | F:AATCCTGCGGCATCCACGAAAC |
| R:CAGCACCGTGTTGGCGTAGAG |
| Item | Group | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| CON | LPS | LPS+LNT | LPS+CMP | LPS+AAP | LPS+LBP | LPS+BRP | ||
| Spleen index | 0.33 ± 0.02 b | 0.33 ± 0.02 b | 0.49 ± 0.02 a | 0.41 ± 0.02 | 0.50 ± 0.01 a | 0.47 ± 0.03 a | 0.50 ± 0.01 a | 0.003 |
| Thymus index | 0.66 ± 0.02 Aa | 0.46 ± 0.02 Bb | 0.56 ± 0.01 | 0.58 ± 0.02 | 0.56 ± 0.01 | 0.49 ± 0.03 Bb | 0.55 ± 0.04 | 0.003 |
| Liver index | 5.62 ± 0.14 Aa | 4.08 ± 0.25 Cc | 5.35 ± 0.10 ABab | 4.86 ± 0.31 ABb | 4.73 ± 0.24 Bb | 5.21 ± 0.17 ABab | 4.92 ± 0.21 ABb | <0.001 |
| Item | Group | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | LPS | LPS+LNT | LPS+CMP | LPS+AAP | LPS+LBP | LPS+BRP | |||
| Duodenum | Velvet length | 176.36 ± 16.95 Aa | 58.96 ± 8.56 De | 99.98 ± 11.58 BCcd | 85.78 ± 2.40 DCd | 133.56 ± 11.95 Bb | 104.65 ± 3.97 BCcd | 118.88 ± 6.44 BCbc | <0.001 |
| Crypt depth | 62.98 ± 3.43 Aa | 27.41 ± 3.89 Dc | 47.37 ± 4.47 BCb | 43.12 ± 3.95 Cb | 59.54 ± 7.38 ABa | 57.66 ± 6.22 ABa | 55.67 ± 5.32 ABa | <0.001 | |
| Jejunum | Velvet length | 154.12 ± 14.11 Aa | 53.53 ± 2.12 De | 84.87 ± 8.96 Ccd | 79.41 ± 4.75 Cd | 101.73 ± 3.30 Cc | 90.54 ± 14.94 Ccd | 124.52 ± 16.62 Bb | <0.001 |
| Crypt depth | 61.71 ± 3.71 Aa | 29.74 ± 0.66 Cc | 49.04 ± 9.37 Bb | 39.48 ± 3.05 BCb | 44.53 ± 5.95 Bb | 44.42 ± 5.78 Bb | 48.82 ± 2.55 Bb | <0.001 | |
| Ileum | Velvet length | 116.41 ± 6.52 Aa | 63.33 ± 3.75 Ee | 78.62 ± 4.73 Dd | 77.85 ± 8.95 Dd | 97.04 ± 1.60 BCc | 88.35 ± 1.33 CDc | 107.29 ± 7.14 ABb | <0.001 |
| Crypt depth | 69.62 ± 5.88 Aa | 31.38 ± 1.06 Cc | 43.17 ± 2.98 BCb | 44.92 ± 4.73 Bb | 45.60 ± 2.38 Bb | 41.71 ± 6.18 BCb | 44.9175 ± 2.58 Bb | <0.001 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, M.; Zhang, W.; Ma, Y.; Liu, T.; Wang, S.; Chen, S.; Wang, Z.; Shen, H. Bioactive Polysaccharides Prevent Lipopolysaccharide-Induced Intestinal Inflammation via Immunomodulation, Antioxidant Activity, and Microbiota Regulation. Foods 2025, 14, 2575. https://doi.org/10.3390/foods14152575
Gao M, Zhang W, Ma Y, Liu T, Wang S, Chen S, Wang Z, Shen H. Bioactive Polysaccharides Prevent Lipopolysaccharide-Induced Intestinal Inflammation via Immunomodulation, Antioxidant Activity, and Microbiota Regulation. Foods. 2025; 14(15):2575. https://doi.org/10.3390/foods14152575
Chicago/Turabian StyleGao, Mingyang, Wanqing Zhang, Yan Ma, Tingting Liu, Sijia Wang, Shuaihu Chen, Zhengli Wang, and Hong Shen. 2025. "Bioactive Polysaccharides Prevent Lipopolysaccharide-Induced Intestinal Inflammation via Immunomodulation, Antioxidant Activity, and Microbiota Regulation" Foods 14, no. 15: 2575. https://doi.org/10.3390/foods14152575
APA StyleGao, M., Zhang, W., Ma, Y., Liu, T., Wang, S., Chen, S., Wang, Z., & Shen, H. (2025). Bioactive Polysaccharides Prevent Lipopolysaccharide-Induced Intestinal Inflammation via Immunomodulation, Antioxidant Activity, and Microbiota Regulation. Foods, 14(15), 2575. https://doi.org/10.3390/foods14152575