
An Overview of Seafood Allergens: Structure–Allergenicity Relationship and Allergenicity Elimination Processing Techniques


Abstract
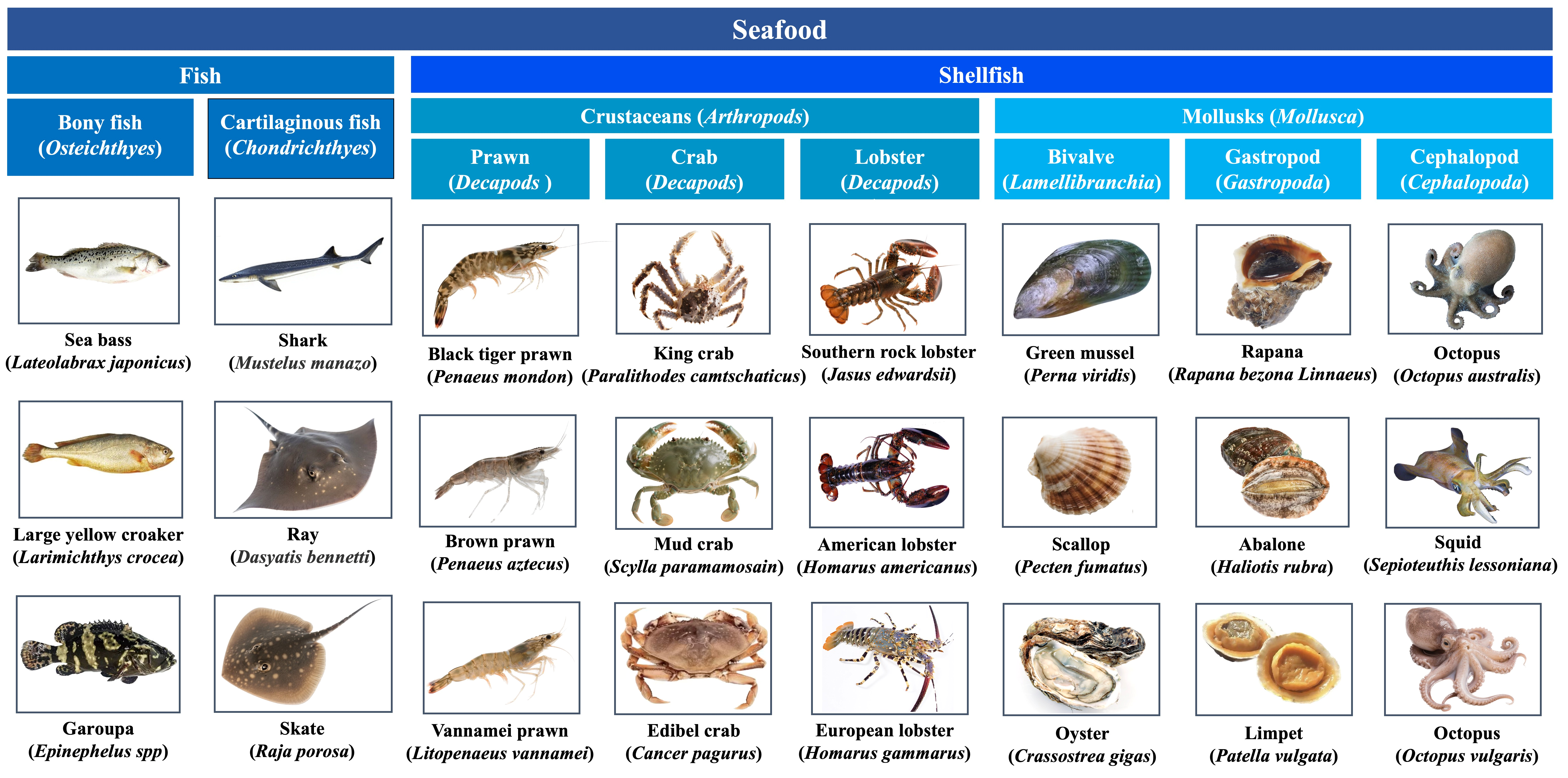
1. Introduction
2. Seafood Allergens and Their Molecular Properties
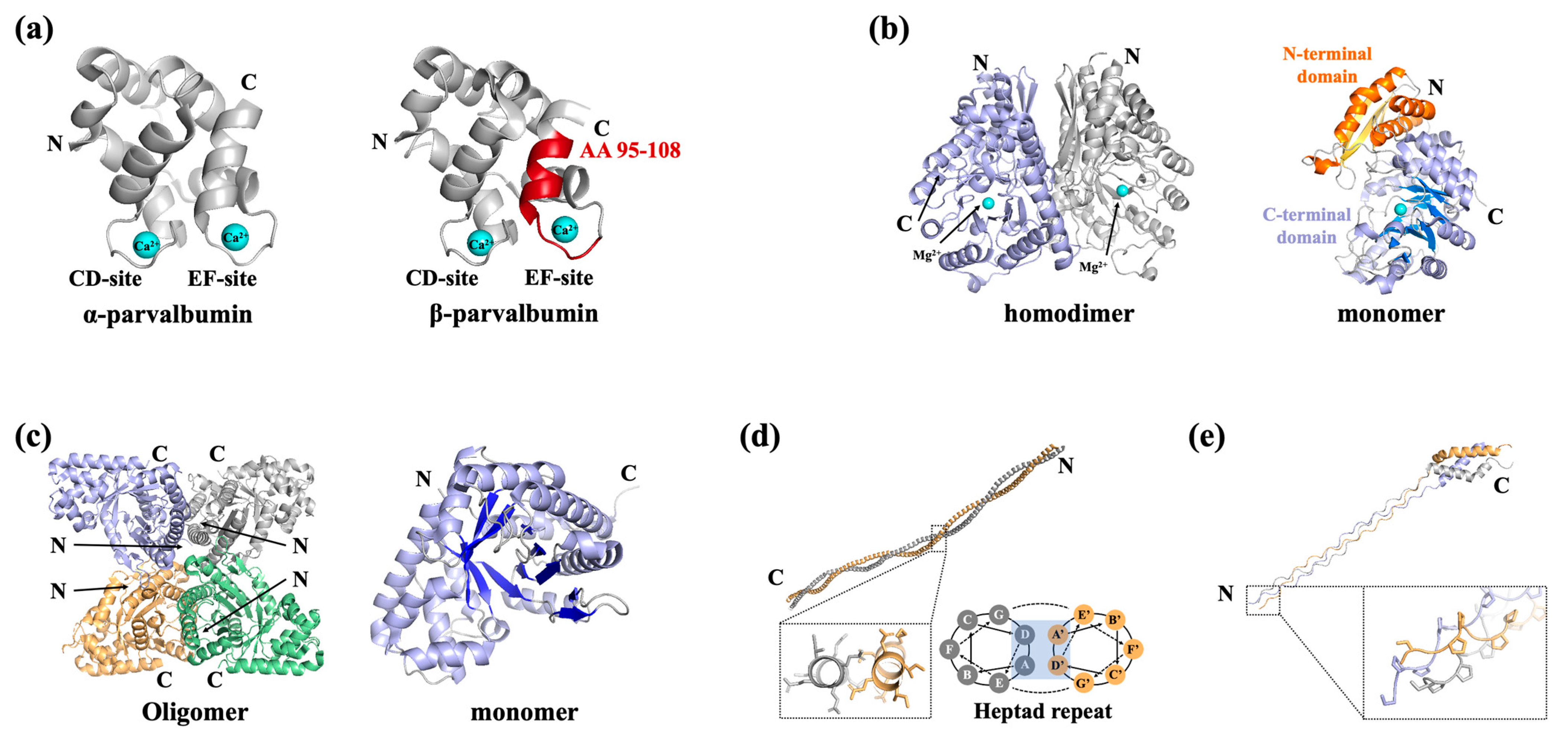
2.1. Fish Allergens
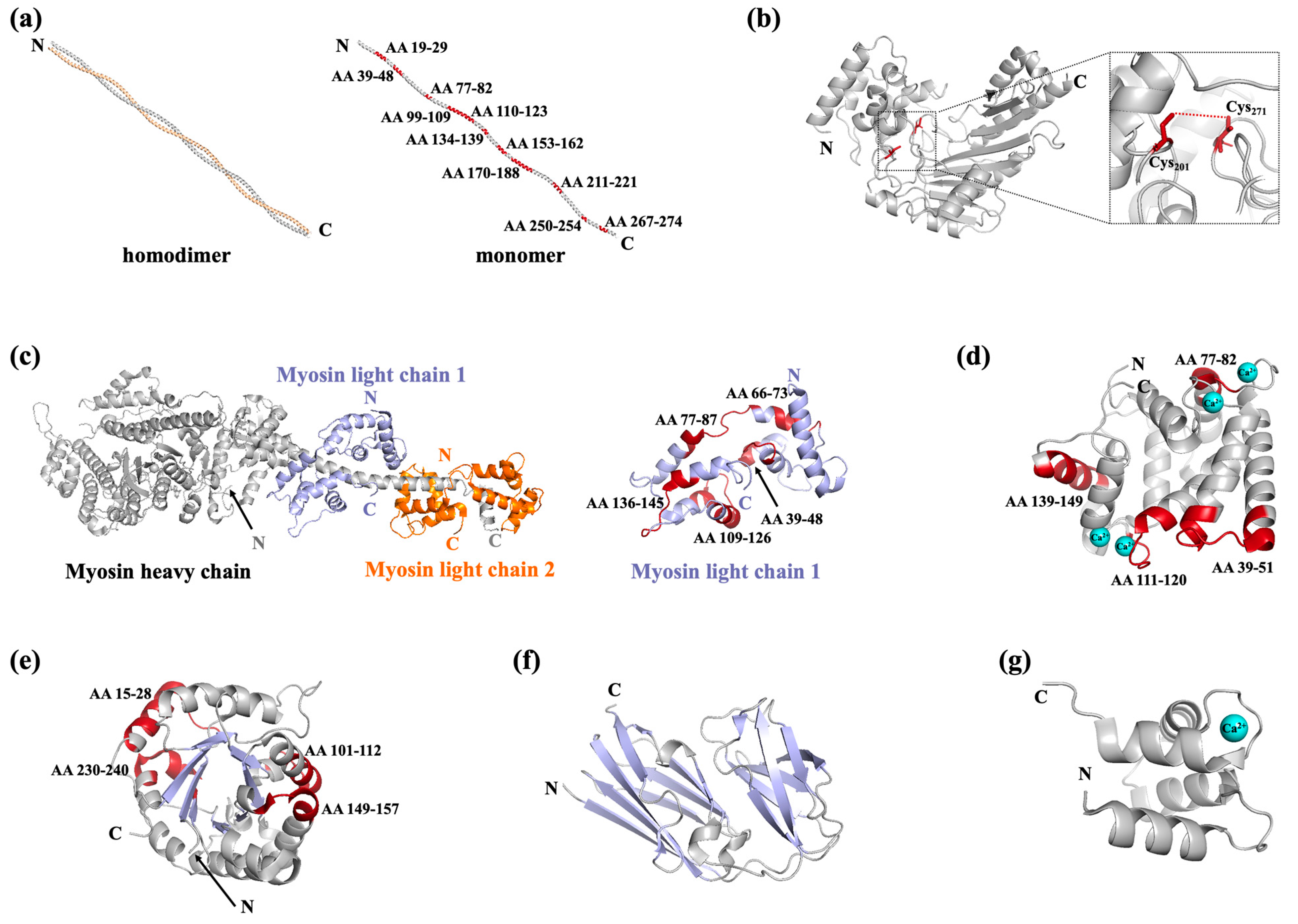
2.2. Shellfish Allergens
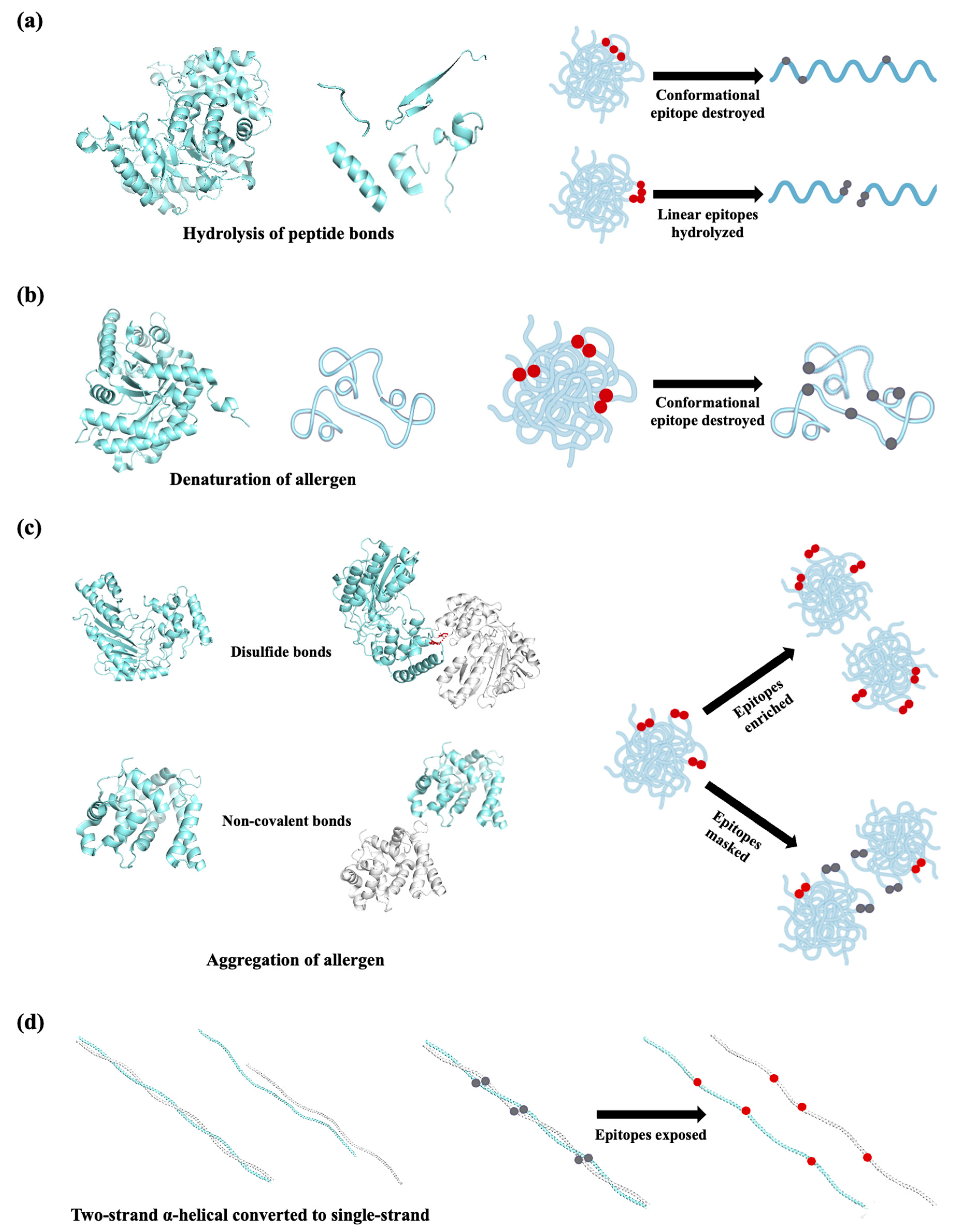
3. Effect of Processing Techniques on Seafood Allergens
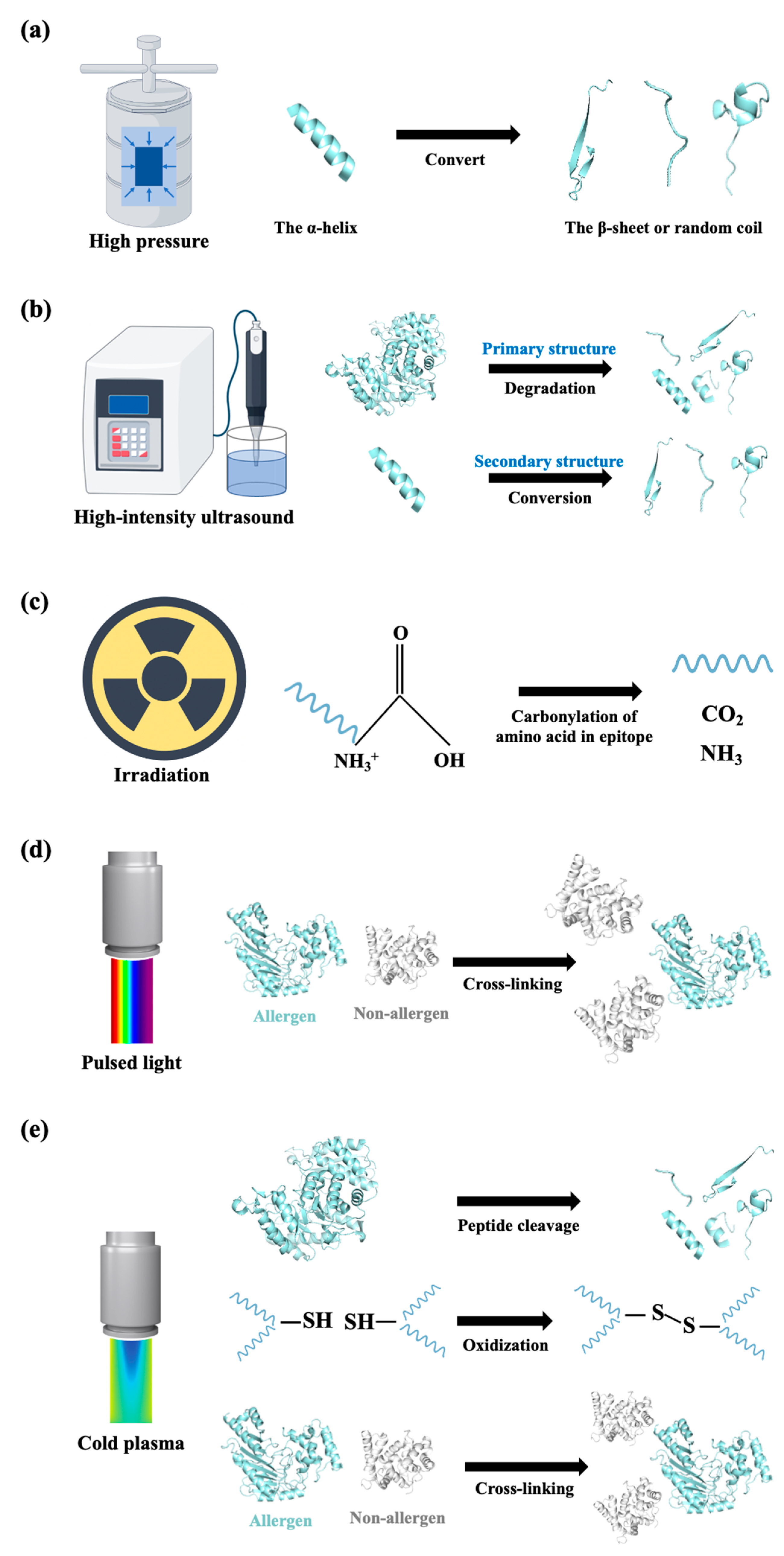
3.1. Physical Processing Techniques
3.1.1. Thermal Treatments
3.1.2. Non-Thermal Treatments
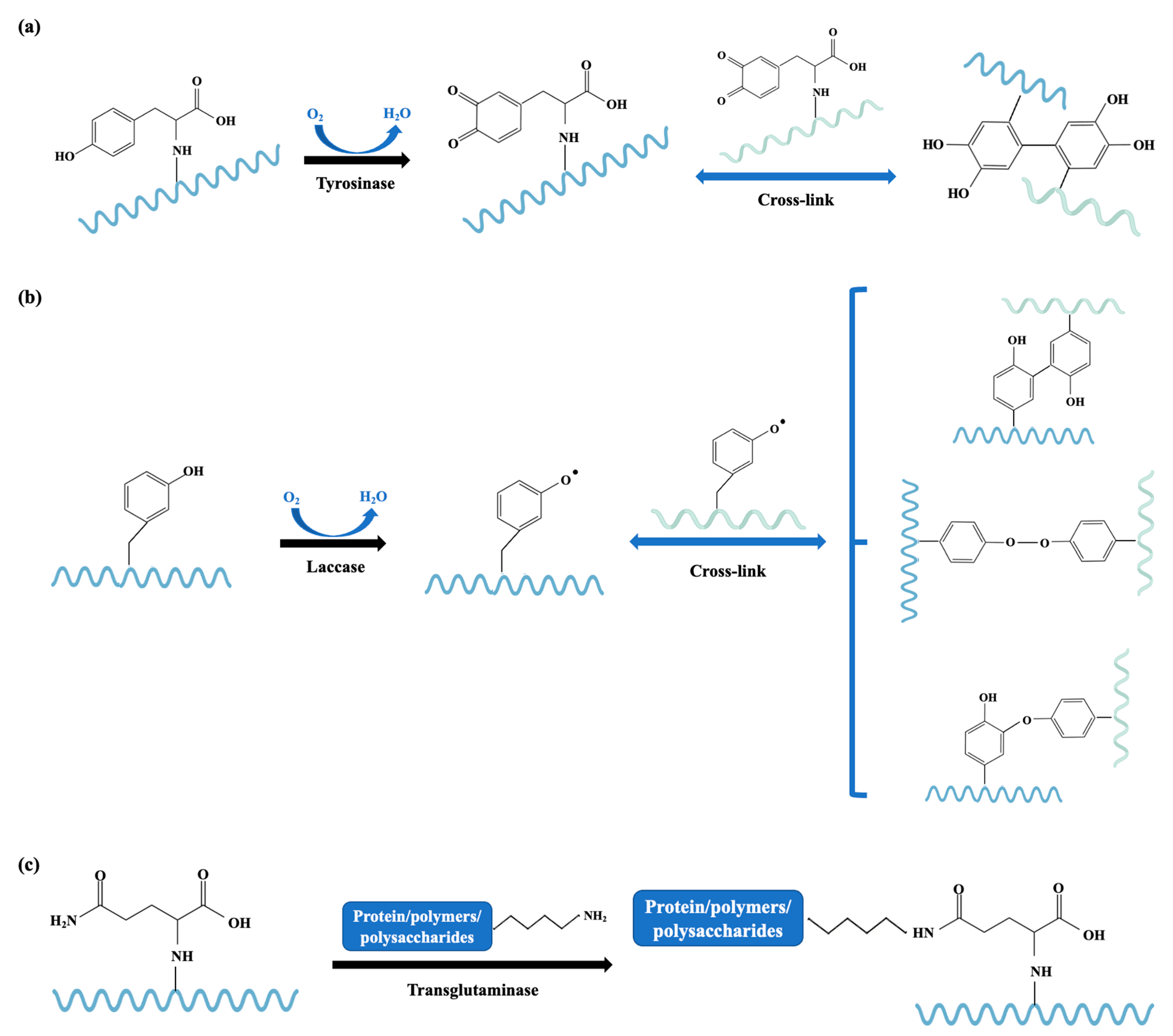
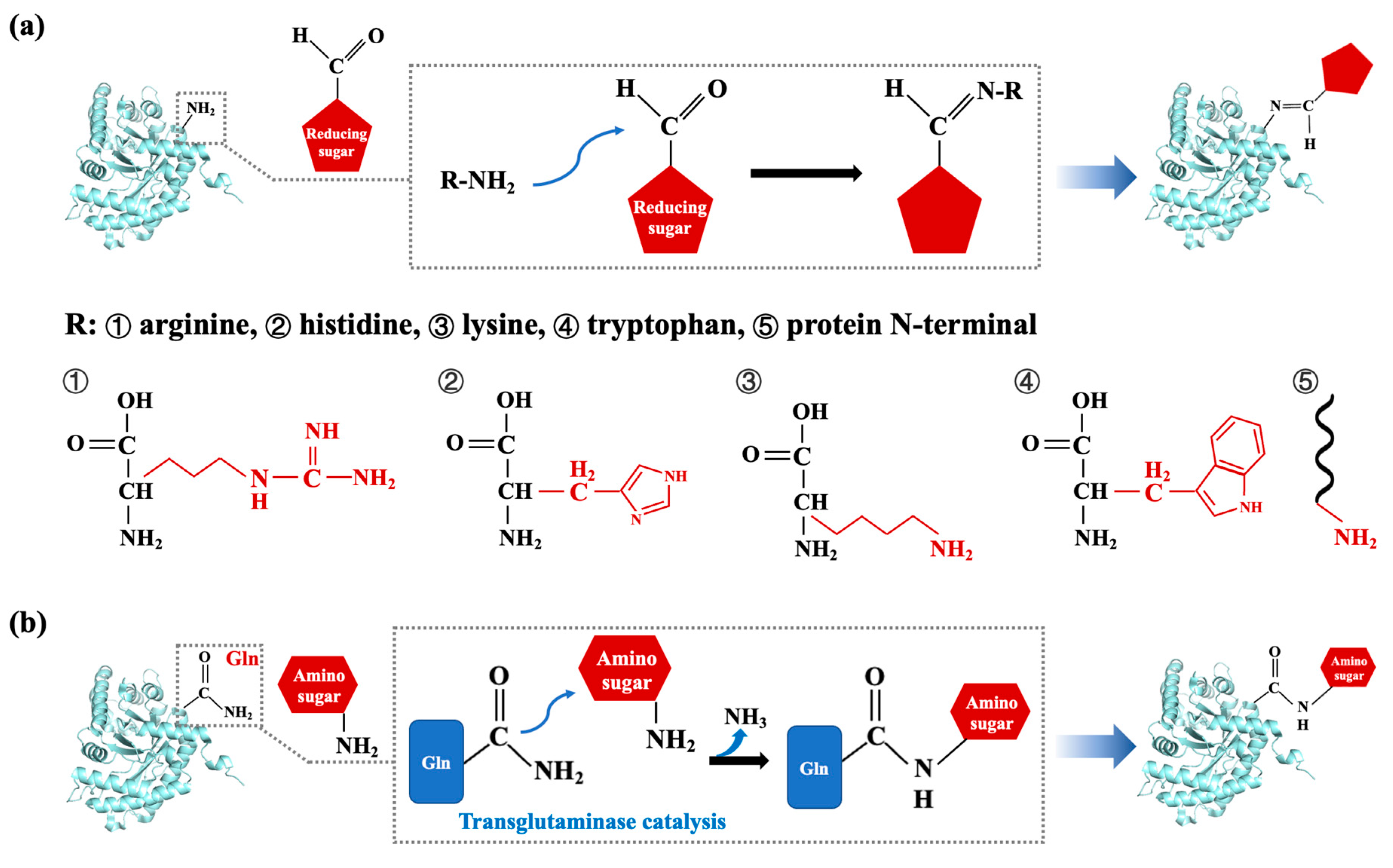
3.2. Chemical Processing Techniques
3.2.1. Enzymatic-Catalyzed Cross-Linking
3.2.2. Glycation Modification
3.3. Biological Processing Techniques
3.3.1. Enzymatic Hydrolysis
3.3.2. Fermentation Treatment
3.4. Combination Processing
4. Challenges and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| WHO/IUIS | World Health Organization and International Union of Immunological Societies |
| MW | Molecular weight |
| PDB | Protein data bank |
References
- Frischmeyer-Guerrerio, P.A.; Young, F.D.; Aktas, O.N.; Haque, T. Insights into the clinical, immunologic, and genetic underpinnings of food allergy. Immunol. Rev. 2024, 326, 162–172. [Google Scholar] [CrossRef]
- Bartha, I.; Almulhem, N.; Santos, A.F. Feast for thought: A comprehensive review of food allergy 2021–2023. J. Allergy Clin. Immunol. 2024, 153, 576–594. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Summary Report of the Ad Hoc Joint FAO/WHO Expert Consultation on Risk Assessment of Food Allergens. Part 1: Review and Validation of Codex Priority Allergen List Through Risk Assessment. 2022. Available online: https://www.fao.org/3/cb4653en/cb4653en.pdf (accessed on 1 May 2025).
- Kamath, S.D.; Bublin, M.; Kitamura, K.; Matsui, T.; Ito, K.; Lopata, A.L. Cross-reactive epitopes and their role in food allergy. J. Allergy Clin. Immunol. 2023, 151, 1178–1190. [Google Scholar] [CrossRef] [PubMed]
- Kvasnicka, J.; Stylianou, K.S.; Nguyen, V.K.; Huang, L.; Chiu, W.A.; Burton, G.A., Jr.; Semrau, J.; Jolliet, O. Human Health Benefits from Fish Consumption vs. Risks from inhalation exposures associated with contaminated sediment remediation: Dredging of the Hudson river. Environ. Health Perspect. 2019, 127, 127004. [Google Scholar] [CrossRef] [PubMed]
- Wai, C.Y.Y.; Leung, N.Y.H.; Chu, K.H.; Leung, P.S.C.; Leung, A.S.Y.; Wong, G.W.K.; Leung, T.F. Overcoming shellfish allergy: How far have we come? Int. J. Mol. Sci. 2020, 21, 2234. [Google Scholar] [CrossRef]
- Spolidoro, G.C.I.; Ali, M.M.; Amera, Y.T.; Nyassi, S.; Lisik, D.; Ioannidou, A.; Rovner, G.; Khaleva, E.; Venter, C.; van Ree, R.; et al. Prevalence estimates of eight big food allergies in Europe: Updated systematic review and meta-analysis. Allergy 2023, 78, 2361–2417. [Google Scholar] [CrossRef]
- Gupta, R.S.; Warren, C.M.; Smith, B.M.; Jiang, J.; Blumenstock, J.A.; Davis, M.M.; Schleimer, R.P.; Nadeau, K.C. Prevalence and severity of food allergies among US adults. JAMA Netw. Open 2019, 2, e185630. [Google Scholar] [CrossRef]
- Davis, C.M.; Gupta, R.S.; Aktas, O.N.; Diaz, V.; Kamath, S.D.; Lopata, A.L. Clinical management of seafood allergy. J. Allergy Clin. Immunol.-Pract. 2020, 8, 37–44. [Google Scholar] [CrossRef]
- Wai, C.Y.Y.; Leung, N.Y.H.; Leung, A.S.Y.; Wong, G.W.K.; Leung, T.F. Seafood allergy in Asia: Geographical Specificity and beyond. Front. Allergy 2021, 2, 676903. [Google Scholar] [CrossRef]
- Feng, H.; Zhou, J.; Lu, Y.; Zhao, Q.; Yang, Z.; Xiong, X.; Luo, N.; Chen, Y.; Wang, L.; Wu, Y. Prevalence of self-reported food allergy among adults in Jiangxi, China. World Allergy Organ. J. 2023, 16, 100773. [Google Scholar] [CrossRef]
- Le, T.T.K.; Tran, T.T.B.; Ho, H.T.M.; Vu, A.T.L.; McBryde, E.; Lopata, A.L. The predominance of seafood allergy in Vietnamese adults: Results from the first population-based questionnaire survey. World Allergy Organ. J. 2020, 13, 100102. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.S.Y.; Wai, C.Y.Y.; Leung, N.Y.H.; Ngai, N.A.; Chua, G.T.; Ho, P.K.; Lam, I.C.S.; Cheng, J.W.C.H.; Chan, O.M.; Li, P.F.; et al. Real-world sensitization and tolerance pattern toseafood in fish-allergic individuals. J. Allergy Clin. Immunol.-Pract. 2024, 12, 633–642.e9. [Google Scholar] [CrossRef]
- Zotova, V.; Clarke, A.E.; Chan, E.S.; Asai, Y.; Chin, R.; Van Lambalgen, C.; Harada, L.; Ben-Shoshan, M. Low resolution rates of seafood allergy. J. Allergy Clin. Immunol. Pract. 2019, 7, 690–692. [Google Scholar] [CrossRef] [PubMed]
- Sudharson, S.; Kalic, T.; Hafner, C.; Breiteneder, H. Newly defined allergens in the WHO/IUIS Allergen Nomenclature Database during 01/2019–03/2021. Allergy 2021, 76, 3359–3373. [Google Scholar] [CrossRef] [PubMed]
- Ruethers, T.; Taki, A.C.; Karnaneedi, S.; Nie, S.; Kalic, T.; Dai, D.; Daduang, S.; Leeming, M.; Williamson, N.A.; Breiteneder, H.; et al. Expanding the allergen repertoire of salmon and catfish. Allergy 2021, 76, 1443–1453. [Google Scholar] [CrossRef]
- Fu, L.L.; Wang, C.; Zhu, Y.; Wang, Y. Seafood allergy: Occurrence, mechanisms and measures. Trends Food Sci. Technol. 2019, 88, 80–92. [Google Scholar] [CrossRef]
- Wu, Y.T.; Lu, Y.Y.; Huang, Y.H.; Lin, H.; Dang, X.; Jing, Y.; Meng, Z.; Wang, X.; Li, Z. Insight analysis of the cross-sensitization of multiple fish parvalbumins via the Th1/Th2 immunological balance and cytokine release from the perspective of safe consumption of fish. Food Qual. Saf. 2022, 6, fyac056. [Google Scholar] [CrossRef]
- Huang, Y.; Li, Z.; Wu, Y.; Li, Y.; Pramod, S.; Chen, G.; Zhu, W.; Zhang, Z.; Wang, H.; Lin, H. Comparative analysis of allergenicity and predicted linear epitopes in α and β parvalbumin from turbot (Scophthalmus maximus). J. Sci. Food Agric. 2023, 103, 2313–2324. [Google Scholar] [CrossRef]
- Yang, R.Q.; Chen, Y.L.; Chen, F.; Wang, H.; Zhang, Q.; Liu, G.M.; Jin, T.; Cao, M.J. Purification, characterization, and crystal structure of parvalbumins, the major allergens in Mustelus griseus. J. Agric. Food Chem. 2018, 66, 8150–8159. [Google Scholar] [CrossRef]
- Perez-Gordo, M.; Pastor-Vargas, C.; Lin, J.; Bardina, L.; Cases, B.; Ibanez, M.D.; Vivanco, F.; Cuesta-Herranz, J.; Sampson, H.A. Epitope mapping of the major allergen from Atlantic cod in Spanish population reveals different IgE-binding patterns. Mol. Nutr. Food Res. 2013, 57, 1283–1290. [Google Scholar] [CrossRef]
- Morales-Amparano, M.B.; Huerta-Ocampo, J.A.; Pastor-Palacios, G.; Teran, L.M. The Role of enolases in allergic disease. J. Allergy Clin. Immunol. Pract. 2021, 9, 3026–3032. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, A.; Hilger, C.; Lehners-Weber, C.; Codreanu-Morel, F.; Morisset, M.; Metz-Favre, C.; Pauli, G.; de Blay, F.; Revets, D.; Muller, C.P.; et al. Identification of enolases and aldolases as important fish allergens in cod, salmon and tuna: Component resolved diagnosis using parvalbumin and the new allergens. Clin. Exp. Allergy 2013, 43, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Wai, C.Y.Y.; Leung, N.Y.H.; Leung, A.S.Y.; Fusayasu, N.; Sato, S.; Xu, K.J.Y.; Yau, Y.S.; Rosa Duque, J.S.; Kwan, M.Y.W.; Cheng, J.; et al. Differential patterns of fish sensitization in Asian populations: Implication for precision diagnosis. Allergol. Int. 2023, 72, 458–465. [Google Scholar] [CrossRef]
- Sariyer, E.; Yakarsonmez, S.; Danis, O.; Turgut-Balik, D. A study of Bos taurus muscle specific enolase; biochemical characterization, homology modelling and investigation of molecular interaction using molecular docking and dynamics simulations. Int. J. Biol. Macromol. 2018, 120 Pt B, 2346–2353. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, R.; Satoh, R.; Nakajima, Y.; Kawasaki, N.; Yamaguchi, T.; Sawada, J.; Nagoya, H.; Teshima, R. Comparative study of GH-transgenic and non-transgenic amago salmon (Oncorhynchus masou ishikawae) allergenicity and proteomic analysis of amago salmon allergens. Regul. Toxicol. Pharmacol. 2009, 55, 300–308. [Google Scholar] [CrossRef]
- Ruethers, T.; Taki, A.C.; Nugraha, R.; Cao, T.T.; Koeberl, M.; Kamath, S.D.; Williamson, N.A.; O’Callaghan, S.; Nie, S.; Mehr, S.S.; et al. Variability of allergens in commercial fish extracts for skin prick testing. Allergy 2019, 74, 1352–1363. [Google Scholar] [CrossRef]
- Boucher, L.E.; Bosch, J. Structure of Toxoplasma gondii fructose-1,6-bisphosphate aldolase. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2014, 70 Pt 9, 1186–1192. [Google Scholar] [CrossRef]
- Liu, R.; Holck, A.L.; Yang, E.; Liu, C.; Xue, W. Tropomyosin from tilapia (Oreochromis mossambicus) as an allergen. Clin. Exp. Allergy 2013, 43, 365–377. [Google Scholar] [CrossRef]
- Li, M.-S.; Xia, F.; Liu, Q.-M.; Zheng, J.-F.; Li, T.-Q.; Liao, Y.-N.; Chen, G.-X.; Luo, L.-Z.; Liu, Y.-X.; Liu, G.-M. Identification and allergenicity analysis of tropomyosin: A heat-stable allergen in Lateolabrax japonicus. J. Agric. Food Chem. 2024, 73, 822–834. [Google Scholar] [CrossRef]
- Whitby, F.G.; Phillips, G.N. Crystal structure of tropomyosin at 7 Angstroms resolution. Proteins-Struct. Funct. Genet. 2000, 38, 49–59. [Google Scholar] [CrossRef]
- Cheng, J.-H.; Wang, H.; Sun, D.-W. An overview of tropomyosin as an important seafood allergen: Structure, cross-reactivity, epitopes, allergenicity, and processing modifications. Compr. Rev. Food Sci. Food Saf. 2022, 21, 127–147. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, X.M.; Wen, Y.Q.; Zhao, J.L.; Xu, T.C.; Yong, L.; Lin, H.; Zhang, H.W.; Li, Z.X. Comparison of tropomyosin released peptide and epitope mapping after in vitro digestion from fish (Larimichthys crocea), shrimp (Litopenaeus vannamei) and clam (Ruditapes philippinarum) through SWATH-MS based proteomics. Food Chem. 2023, 403, 134314. [Google Scholar] [CrossRef]
- Hamada, Y.; Nagashima, Y.; Shiomi, K. Identification of collagen as a new fish allergen. Biosci. Biotechnol. Biochem. 2001, 65, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Akiyama, H.; Huge, J.; Kubota, H.; Chikazawa, S.; Satoh, T.; Miyake, T.; Uhara, H.; Okuyama, R.; Nakagawara, R.; et al. Fish collagen is an important panallergen in the Japanese population. Allergy 2016, 71, 720–723. [Google Scholar] [CrossRef]
- Shimojo, N.; Yagami, A.; Ohno, F.; Tsurumi, Y.; Nakamura, M.; Suzuki, K.; Kuwabara, K.; Futamura, K.; Ohno, S.; Yokogawa, T.; et al. Fish collagen as a potential indicator of severe allergic reactions among patients with fish allergies. Clin. Exp. Allergy 2022, 52, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Bella, J. Collagen structure: New tricks from a very old dog. Biochem. J. 2016, 473, 1001–1025. [Google Scholar] [CrossRef]
- Boudko, S.P.; Bachinger, H.P. Structural insight for chain selection and stagger control in collagen. Sci. Rep. 2016, 6, 37831. [Google Scholar] [CrossRef]
- Kalic, T.; Kamath, S.D.; Ruethers, T.; Taki, A.C.; Nugraha, R.; Le, T.T.K.; Humeniuk, P.; Williamson, N.A.; Hira, D.; Rolland, J.M.; et al. Collagen—An important fish allergen for improved diagnosis. J. Allergy Clin. Immunol. Pract. 2020, 8, 3084–3092.e10. [Google Scholar] [CrossRef]
- Valverde-Monge, M.; Pastor-Vargas, C.; Rodriguez Del Rio, P.; Escudero, C.; Sanchez-Garcia, S.; Mendez Brea, P.; Ibanez, M.D. Anaphylaxis by exclusive allergy to swordfish and identification of a new fish allergen. Pediatr. Allergy Immunol. 2018, 29, 563–565. [Google Scholar] [CrossRef]
- Xia, F.; Li, M.S.; Liu, Q.M.; Liu, M.; Yang, Y.; Cao, M.J.; Chen, G.X.; Jin, T.; Liu, G.M. Crystal structure analysis and conformational epitope mutation of triosephosphate isomerase, a mud crab allergen. J. Agric. Food Chem. 2019, 67, 12918–12926. [Google Scholar] [CrossRef]
- Larco-Rojas, X.; Gonzalez-Gutierrez, M.L.; Vazquez-Cortes, S.; Bartolome, B.; Pastor-Vargas, C.; Fernandez-Rivas, M. Occupational asthma and urticaria in a fishmonger due to creatine kinase, a cross-reactive fish allergen. J. Investig. Allergol. Clin. Immunol. 2017, 27, 386–388. [Google Scholar] [CrossRef]
- Yang, Y.; He, X.R.; Huan, F.; Bai, T.L.; Zhang, Q.H.; Li, F.J.; Chen, G.X.; Zheng, P.Y.; Xu, L.M.; Liu, G.M. Crystal structure of Pro c 2 provides insights into cross-reactivity of aquatic allergens from the phosphagen kinase family. J. Agric. Food Chem. 2024, 72, 28400–28411. [Google Scholar] [CrossRef]
- Hanaoka, K.; Takahagi, S.; Ishii, K.; Nakano, M.; Chinuki, Y.; Tanaka, A.; Yanase, Y.; Hide, M. Type-I-hypersensitivity to 15 kDa, 28 kDa and 54 kDa proteins in vitellogenin specific to Gadus chalcogrammus roe. Allergol. Int. 2020, 69, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.R.; Day, E.D., Jr.; Miller, J.S. The major heat stable allergen of shrimp. Ann. Allergy 1981, 47, 17–22. [Google Scholar]
- Nugraha, R.; Kamath, S.D.; Johnston, E.; Karnaneedi, S.; Ruethers, T.; Lopata, A.L. Conservation analysis of B-cell allergen epitopes to predict clinical cross-reactivity between shellfish and inhalant invertebrate allergens. Front. Immunol. 2019, 10, 2676. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.J.; He, H.; Wang, X.; Wu, S.; Wang, Q.; Zhang, J.; Qiao, Y.; Liu, H. Research progress on shrimp allergens and allergenicity reduction methods. Foods 2025, 14, 895. [Google Scholar] [CrossRef] [PubMed]
- James, J.K.; Pike, D.H.; Khan, I.J.; Nanda, V. Structural and dynamic properties of allergen and non-allergen forms of tropomyosin. Structure 2018, 26, 997. [Google Scholar] [CrossRef]
- Liu, M.; Huan, F.; Li, M.; Han, T.; Xia, F.; Yang, Y.; Liu, Q.; Chen, G.; Cao, M.; Liu, G. Mapping and IgE-binding capacity analysis of heat/digested stable epitopes of mud crab allergens. Food Chem. 2021, 344, 128735. [Google Scholar] [CrossRef]
- Lv, L.T.; Wei, F.L.; Liu, L.; Song, F.; Hou, X.; Yang, Q. Study on the allergenicity of tropomyosin from different aquatic products based on conformational and linear epitopes analysis. J. Agric. Food Chem. 2025, 73, 4936–4946. [Google Scholar] [CrossRef]
- Li, M.S.; Xia, F.; Liu, Q.; Chen, Y.; Yun, X.; Liu, M.; Chen, G.-x.; Wang, L.; Cao, M.-J.; Liu, G.-M. IgE Epitope analysis for Scy p 1 and Scy p 3, the heat-stable myofibrillar allergens in mud crab. J. Agric. Food Chem. 2022, 70, 12189–12202. [Google Scholar] [CrossRef]
- Yu, C.J.; Lin, Y.F.; Chiang, B.L.; Chow, L.P. Proteomics and immunological analysis of a novel shrimp allergen, Pen m 2. J. Immunol. 2003, 170, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, G.Y.; Yang, H.; Hu, M.J.; Cao, M.J.; Su, W.J.; Jin, T.; Liu, G.M. Crystal structure determination of Scylla paramamosain arginine kinase, an allergen that may cause cross-reactivity among invertebrates. Food Chem. 2019, 271, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cao, M.J.; Alcocer, M.; Liu, Q.M.; Fei, D.X.; Mao, H.Y.; Liu, G.M. Mapping and characterization of antigenic epitopes of arginine kinase of Scylla paramamosain. Mol. Immunol. 2015, 65, 310–320. [Google Scholar] [CrossRef]
- Chen, H.L.; Mao, H.Y.; Cao, M.J.; Cai, Q.F.; Su, W.J.; Zhang, Y.X.; Liu, G.M. Purification, physicochemical and immunological characterization of arginine kinase, an allergen of crayfish (Procambarus clarkii). Food Chem. Toxicol. 2013, 62, 475–484. [Google Scholar] [CrossRef]
- Mao, H.-Y.; Cao, M.-J.; Maleki, S.J.; Cai, Q.-F.; Su, W.-J.; Yang, Y.; Liu, G.-M. Structural characterization and IgE epitope analysis of arginine kinase from Scylla paramamosain. Mol. Immunol. 2013, 56, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Gao, X.; Lin, H.; Lin, H.; Zhang, Z.; Khan, M.U.; Li, Y.; Chen, Y.; Li, Z. Identification and amino acid analysis of allergenic epitopes of a novel allergen paramyosin (Rap v 2) from Rapana venosa. J. Agric. Food Chem. 2021, 69, 5381–5391. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Ding, X.; Gao, X.; Lin, H.; Khan, M.U.; Lin, H.; Dang, X.; Li, Z. Immunological cross-reactivity involving mollusc species and mite-mollusc and cross-reactive allergen PM are risk factors of mollusc allergy. J. Agric. Food Chem. 2022, 70, 360–372. [Google Scholar] [CrossRef]
- Yu, C.; Gao, X.; Lin, H.; Xu, L.; Ahmed, I.; Khan, M.U.; Xu, M.; Chen, Y.; Li, Z. Purification, characterization, and three-dimensional structure prediction of paramyosin, a novel allergen of Rapana venosa. J. Agric. Food Chem. 2020, 68, 14632–14642. [Google Scholar] [CrossRef]
- Yang, Y.T.; Gourinath, S.; Kovacs, M.; Nyitray, L.; Reutzel, R.; Himmel, D.M.; O’Neall-Hennessey, E.; Reshetnikova, L.; Szent-Gyorgyi, A.G.; Brown, J.H.; et al. Rigor-like structures from muscle myosins reveal key mechanical elements in the transduction pathways of this allosteric motor. Structure 2007, 15, 553–564. [Google Scholar] [CrossRef]
- Conti, A.; Alqassir, N.; Breda, D.; Zanardi, A.; Alessio, M.; Burastero, S.E. Serological proteome analysis identifies crustacean myosin heavy chain type 1 protein and house dust mite Der p 14 as cross-reacting allergens. Adv. Clin. Exp. Med. 2023, 32, 107–112. [Google Scholar] [CrossRef]
- Saenz de San Pedro, B.; Lopez Guerrero, A.; Navarrete del Pino, M.A.; Alcantara Villar, M.; Alvarez, F.; Lopez-Matas, M.A.; Carnes, J. Myosin heavy chain: An allergen involved in anaphylaxis to shrimp head. J. Investig. Allergol. Clin. Immunol. 2023, 33, 66–68. [Google Scholar]
- Shibata, Y.; Satoshi, S.; Oishi, T.; Ohko, K.; Fujieda, M.; Naka, T.; Sano, S. Myosin heavy chain, a novel allergen for fish allergy in patients with atopic dermatitis. J. Investig. Dermatol. 2019, 139, S223. [Google Scholar] [CrossRef]
- Ayuso, R.; Grishina, G.; Bardina, L.; Carrillo, T.; Blanco, C.; Ibanez, M.D.; Sampson, H.A.; Beyer, K. Myosin light chain is a novel shrimp allergen, Lit v 3. J. Allergy Clin. Immunol. 2008, 122, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.M.A.; Kamath, S.; Lopata, A.L.; Helleur, R.J. Analysis of the allergenic proteins in black tiger prawn (Penaeus monodon) and characterization of the major allergen tropomyosin using mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2462–2470. [Google Scholar] [CrossRef]
- Bauermeister, K.; Wangorsch, A.; Garoffo, L.P.; Reuter, A.; Conti, A.; Taylor, S.L.; Lidholm, J.; DeWitt, A.M.; Enrique, E.; Vieths, S.; et al. Generation of a comprehensive panel of crustacean allergens from the North Sea Shrimp Crangon crangon. Mol. Immunol. 2011, 48, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Li, M.S.; Xia, F.; Liu, M.; He, X.R.; Chen, Y.Y.; Bai, T.L.; Chen, G.X.; Wang, L.; Cao, M.J.; Liu, G.M. Cloning, expression, and epitope identification of myosin light chain 1: An allergen in mud crab. J. Agric. Food Chem. 2019, 67, 10458–10469. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, H.-F.; Zhang, Y.-X.; Chen, H.-L.; Cao, M.-J.; Li, M.-S.; Zhang, M.-L.; He, X.-R.; Liu, G.-M. Expression and epitope identification of myosin light chain isoform 1, an allergen in Procambarus clarkii. Food Chem. 2020, 317, 126422. [Google Scholar] [CrossRef]
- Zhang, Y.-X.; Chen, H.-L.; Maleki, S.J.; Cao, M.-J.; Zhang, L.-J.; Su, W.-J.; Liu, G.-M. Purification, characterization, and analysis of the allergenic properties of myosin light chain in Procambarus clarkii. J. Agric. Food Chem. 2015, 63, 6271–6282. [Google Scholar] [CrossRef]
- Ayuso, R.; Grishina, G.; Dolores Ibanez, M.; Blanco, C.; Carrillo, T.; Bencharitiwong, R.; Sanchez, S.; Nowak-Wegrzyn, A.; Sampson, H.A. Sarcoplasmic calcium-binding protein is an EF-hand-type protein identified as a new shrimp allergen. J. Allergy Clin. Immunol. 2009, 124, 114–120. [Google Scholar] [CrossRef]
- Han, T.J.; Huan, F.; Liu, M.; Li, M.S.; Yang, Y.; Chen, G.X.; Lai, D.; Cao, M.J.; Liu, G.M. IgE epitope analysis of sarcoplasmic-calcium-binding protein, a heat-resistant allergen in Crassostrea angulata. Food Funct. 2021, 12, 8570–8582. [Google Scholar] [CrossRef]
- Hu, M.J.; Liu, G.Y.; Yang, Y.; Pan, T.M.; Liu, Y.X.; Sun, L.C.; Cao, M.J.; Liu, G.M. Cloning, expression, and the effects of processing on sarcoplasmic calcium-binding protein: An important allergen in mud crab. J. Agric. Food Chem. 2017, 65, 6247–6257. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Jin, T.; Li, M.; Yun, X.; Huan, F.; Liu, Q.; Hu, M.; Wei, X.; Zheng, P.; Liu, G. Crystal structure analysis of sarcoplasmic-calcium-binding protein: An allergen in Scylla paramamosain. J. Agric. Food Chem. 2023, 71, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, Y.-X.; Liu, M.; Maleki, S.J.; Zhang, M.-L.; Liu, Q.-M.; Cao, M.-J.; Su, W.-J.; Liu, G.-M. Triosephosphate isomerase and filamin C share common epitopes as novel allergens of Procambarus clarkii. J. Agric. Food Chem. 2017, 65, 950–963. [Google Scholar] [CrossRef]
- Xia, F.; Li, M.; Liu, Q.; Liu, H.; Yang, Y.; Liu, M.; Chen, G.; Luo, L.; Liu, Y.; Liu, G. Allergenicity and linear epitope analysis of Scy p 8, an allergen from mud crab. J. Agric. Food Chem. 2024, 72, 13402–13414. [Google Scholar] [CrossRef]
- He, X.-R.; Yang, Y.; Chen, Y.-X.; Kang, S.; Li, F.-J.; Li, D.-X.; Liu, Q.-M.; Chen, G.-X.; Chen, X.-M.; Liu, G.-M. Immunoglobulin E epitope mapping and structure-allergenicity relationship analysis of crab allergen Scy p 9. J. Agric. Food Chem. 2023, 71, 17379–17390. [Google Scholar] [CrossRef]
- He, X.R.; Yang, Y.; Kang, S.; Chen, Y.X.; Zheng, P.Y.; Chen, G.X.; Chen, X.M.; Cao, M.J.; Jin, T.; Liu, G.M. Crystal structure analysis and ige epitope mapping of allergic predominant region in Scylla paramamosain filamin C, Scy p 9. J. Agric. Food Chem. 2022, 70, 1282–1292. [Google Scholar] [CrossRef]
- Karnaneedi, S.; Huerlimann, R.; Johnston, E.B.; Nugraha, R.; Ruethers, T.; Taki, A.C.; Kamath, S.D.; Wade, N.M.; Jerry, D.R.; Lopata, A.L. Novel allergen discovery through comprehensive de novo transcriptomic analyses of five shrimp species. Int. J. Mol. Sci. 2021, 22, 32. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.S.; Yumoto, F.; Tanaka, H.; Miyakawa, T.; Miyauchi, Y.; Takeshita, D.; Sawano, Y.; Ojima, T.; Ohtsuki, I.; Tanokura, M. Structure of the Ca2+-saturated C-terminal domain of scallop troponin C in complex with a troponin I fragment. Biol. Chem. 2013, 394, 55–68. [Google Scholar] [CrossRef]
- Giuffrida, M.G.; Villalta, D.; Mistrello, G.; Amato, S.; Asero, R. Shrimp allergy beyond tropomyosin in Italy: Clinical relevance of arginine kinase, sarcoplasmic calcium binding protein and hemocyanin. Eur. Ann. Allergy Clin. Immunol. 2014, 46, 172–177. [Google Scholar]
- Lu, J.; Luan, H.; Wang, C.; Zhang, L.; Shi, W.; Xu, S.; Jin, Y.; Lu, Y. Molecular and allergenic properties of natural hemocyanin from Chinese mitten crab (Eriocheir sinensis). Food Chem. 2023, 424, 136422. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Y.; Zhang, S.; Zhang, L.; Lan, F. The effect of immunotherapy on cross-reactivity between house dust mite and other allergens in house dust mite -sensitized patients with allergic rhinitis. Expert Rev. Clin. Immunol. 2021, 17, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; She, T.; Zhang, Y.; Li, S.; Xu, Z.; Yan, J.; Li, H. Identification and characterization of ovary development-related protein EJO1 (Eri s 2) from the ovary of Eriocheir sinensis as a new food allergen. Mol. Nutr. Food Res. 2016, 60, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Munera, M.; Martinez, D.; Wortmann, J.; Zakzuk, J.; Keller, W.; Caraballo, L.; Puerta, L. Structural and allergenic properties of the fatty acid binding protein from shrimp Litopenaeus vannamei. Allergy 2022, 77, 1534–1544. [Google Scholar] [CrossRef]
- Wai, C.Y.Y.; Leung, N.Y.H.; Leung, A.S.Y.; Ngai, S.M.; Pacharn, P.; Yau, Y.S.; Rosa Duque, J.S.D.; Kwan, M.Y.W.; Jirapongsananuruk, O.; Chan, W.H.; et al. Comprehending the allergen repertoire of shrimp for precision molecular diagnosis of shrimp allergy. Allergy 2022, 77, 3041–3051. [Google Scholar] [CrossRef]
- Khan, M.U.; Ahmed, I.; Lin, H.; Li, Z.; Costa, J.; Mafra, I.; Chen, Y.; Wu, Y.-N. Potential efficacy of processing technologies for mitigating crustacean allergenicity. Crit. Rev. Food Sci. Nutr. 2019, 59, 2807–2830. [Google Scholar] [CrossRef] [PubMed]
- Rosmilah, M.; Shahnaz, M.; Meinir, J.; Masita, A.; Noormalin, A.; Jamaluddin, M. Identification of parvalbumin and two new thermolabile major allergens of Thunnus tonggol using a proteomics approach. Int. Arch. Allergy Immunol. 2013, 162, 299–309. [Google Scholar] [CrossRef]
- Kubota, H.; Kobayashi, A.; Kobayashi, Y.; Shiomi, K.; Hamada-Sato, N. Reduction in IgE reactivity of Pacific mackerel parvalbumin by heat treatment. Food Chem. 2016, 206, 78–84. [Google Scholar] [CrossRef]
- Liang, J.; Taylor, S.L.; Baumert, J.; Lopata, A.L.; Lee, N.A. Effects of thermal treatment on the immunoreactivity and quantification of parvalbumin from Southern hemisphere fish species with two anti-parvalbumin antibodies. Food Control 2021, 121, 107675. [Google Scholar] [CrossRef]
- van der Ventel, M.L.; Nieuwenhuizen, N.E.; Kirstein, F.; Hikuam, C.; Jeebhay, M.F.; Swoboda, I.; Brombacher, F.; Lopata, A.L. Differential responses to natural and recombinant allergens in a murine model of fish allergy. Mol. Immunol. 2011, 48, 637–646. [Google Scholar] [CrossRef]
- Chen, Y.-T.; Hsieh, Y.-H.P. Characterization of a 36 kDa antigenic protein of fish-specific monoclonal-antibody 8F5. Food Chem. 2022, 379, 132149. [Google Scholar] [CrossRef]
- Fang, L.; Li, G.M.; Zhang, J.T.; Gu, R.; Cai, M.; Lu, J. Identification and mutational analysis of continuous, immunodominant epitopes of the major oyster allergen Crag 1. Clin. Immunol. 2019, 201, 20–29. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Y.; Xu, L.; Zeng, J.; Liu, Y.; Timira, V.; Zhang, Z.; Lin, H.; Li, Z. Thermal induced the structural alterations, increased IgG/IgE binding capacity and reduced immunodetection recovery of tropomyosin from shrimp (Litopenaeus vannamei). Food Chem. 2022, 391, 133215. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.L.; Zhu, W.Y.; Zeng, J.H.; Liu, Y.; Li, H.; Wang, H.; Zhang, Z.; Lin, H.; Li, Z. Insight into the mechanism of allergenicity decreasing in recombinant sarcoplasmic calcium-binding protein from shrimp (Litopenaeus vannamei) with thermal processing via spectroscopy and molecular dynamics simulation techniques. Food Res. Int. 2022, 157, 111427. [Google Scholar] [CrossRef]
- Cabanillas, B.; Novak, N. Effects of daily food processing on allergenicity. Crit. Rev. Food Sci. Nutr. 2019, 59, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Ekezie, F.-G.C.; Cheng, J.-H.; Sun, D.-W. Effects of nonthermal food processing technologies on food allergens: A review of recent research advances. Trends Food Sci. Technol. 2018, 74, 12–25. [Google Scholar] [CrossRef]
- Jin, Y.F.; Deng, Y.; Qian, B.J.; Zhang, Y.; Liu, Z.; Zhao, Y. Allergenic response to squid (Todarodes pacificus) tropomyosin Tod p1 structure modifications induced by high hydrostatic pressure. Food Chem. Toxicol. 2015, 76, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Carrera, M.; Fidalgo, L.G.; Saraiva, J.A.; Aubourg, S.P. Effects of high-pressure treatment on the muscle proteome of hake by bottom-up proteomics. J. Agric. Food Chem. 2018, 66, 4559–4570. [Google Scholar] [CrossRef]
- Zhang, H.E.; Liao, H.Q.; Lu, Y.B.; Hu, Y.; Yang, H.; Cao, S.; Qi, X. Effects of high hydrostatic pressure on the structural characteristics of parvalbumin of cultured large yellow croaker (Larimichthys crocea). J. Food Process. Preserv. 2020, 44, e14911. [Google Scholar] [CrossRef]
- Chen, W.Q.; Ma, H.L.; Wang, Y.Y. Recent advances in modified food proteins by high intensity ultrasound for enhancing functionality: Potential mechanisms, combination with other methods, equipment innovations and future directions. Ultrason. Sonochem. 2022, 85, 105993. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Zhang, X.F.; Chen, W.; Zhou, P. Conformation stability, in vitro digestibility and allergenicity of tropomyosin from shrimp (Exopalaemon modestus) as affected by high intensity ultrasound. Food Chem. 2018, 245, 997–1009. [Google Scholar] [CrossRef]
- Li, Z.X.; Lin, H.; Cao, L.M.; Jameel, K. Effect of high intensity ultrasound on the allergenicity of shrimp. J. Zhejiang Univ. Sci. B 2006, 7, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Vanga, S.K.; Singh, A.; Raghavan, V. Review of conventional and novel food processing methods on food allergens. Crit. Rev. Food Sci. Nutr. 2017, 57, 2077–2094. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.F.; Yang, J.Y.; Liu, K.X.; Xie, X.; Hong, L.; Wang, S.; Wang, S. Irradiation technology: An effective and promising strategy for eliminating food allergens. Food Res. Int. 2021, 148, 110578. [Google Scholar] [CrossRef]
- Li, Z.X.; Lu, Z.C.; Khan, M.N.; Lin, H.; Zhang, L. Protein carbonylation during electron beam irradiation may be responsible for changes in IgE binding to turbot parvalbumin. Food Chem. Toxicol. 2014, 69, 32–37. [Google Scholar] [CrossRef]
- Guan, A.; Mei, K.; Lv, M.; Lu, J.; Lou, Q.; Yang, W. The effect of electron beam irradiation on IgG binding capacity and conformation of tropomyosin in shrimp. Food Chem. 2018, 264, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.C.; Mei, K.L.; Zhang, H.; Xu, D.; Yang, W. Effects of electron beam irradiation on the biochemical properties and structure of myofibrillar protein from Tegillarca granosa meat. Food Chem. 2018, 254, 64–69. [Google Scholar] [CrossRef]
- Salazar, F.; Pizarro-Oteiza, S.; Kasahara, I.; Labbe, M.; Estay, C.; Tarnok, M.E.; Aguilar, L.; Ibanez, R.A. Effect of pulsed ultraviolet light on the degree of antigenicity and hydrolysis in cow’s milk proteins. J. Food Process Eng. 2024, 47, e14629. [Google Scholar] [CrossRef]
- Kaur, S.; Kumar, Y.; Singh, V.; Kaur, J.; Panesar, P.S. Cold plasma technology: Reshaping food preservation and safety. Food Control 2024, 163, 110537. [Google Scholar] [CrossRef]
- Ekezie, F.G.C.; Sun, D.-W.; Cheng, J.H. Altering the IgE binding capacity of king prawn (Litopenaeus vannamei) tropomyosin through conformational changes induced by cold argon-plasma jet. Food Chem. 2019, 300, 125143. [Google Scholar] [CrossRef]
- Shan, C.Y.; Wu, G.Q.; Bu, Y.; Zhu, W.; Li, J.; Li, X. Effect of transglutaminase cross-linking on debittering of Alaska pollock frame hydrolysate. Int. J. Food Sci. Technol. 2024, 59, 438–449. [Google Scholar] [CrossRef]
- Zeng, J.; Wang, Q.; Yi, H.; Chen, C.; Du, C.; Xiong, G.; Wang, B.; Zhao, J.; Zhang, L.; Gong, P. Recent insights in cow’s milk protein allergy: Clinical relevance, allergen features, and influences of food processing. Trends Food Sci. Technol. 2025, 156, 104830. [Google Scholar] [CrossRef]
- Ahmed, I.; Lin, H.; Xu, L.; Li, S.; Costa, J.; Mafra, I.; Chen, G.; Gao, X.; Li, Z. Immunomodulatory effect of laccase/caffeic acid and transglutaminase in alleviating shrimp tropomyosin (Met e 1) allergenicity. J. Agric. Food Chem. 2020, 68, 7765–7778. [Google Scholar] [CrossRef] [PubMed]
- Fei, D.X.; Liu, Q.M.; Chen, F.; Yang, Y.; Chen, Z.-W.; Cao, M.-J.; Liu, G.-M. Assessment of the sensitizing capacity and allergenicity of enzymatic cross-linked arginine kinase, the crab allergen. Mol. Nutr. Food Res. 2016, 60, 1707–1718. [Google Scholar] [CrossRef]
- Liu, G.Y.; Hu, M.J.; Sun, L.C.; Han, X.; Liu, Q.; Alcocer, M.; Fei, D.; Cao, M.-J.; Liu, G.-M. Allergenicity and oral tolerance of enzymatic cross-linked tropomyosin evaluated using cell and mouse models. J. Agric. Food Chem. 2017, 65, 2205–2213. [Google Scholar] [CrossRef]
- Ahmed, I.; Lin, H.; Li, Z.; Xu, L.; Qazi, I.M.; Luo, C.; Gao, X.; Khan, M.U.; Iqbal, A.; Guo, Y.; et al. Tyrosinase/caffeic acid cross-linking alleviated shrimp (Metapenaeus ensis) tropomyosin-induced allergic responses by modulating the Th1/Th2 immunobalance. Food Chem. 2021, 340, 127948. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.L.; Ma, J.J.; Ahmed, I.; Lv, L.; Li, Z.; Lin, H. Effect of tyrosinase-catalyzed crosslinking on the structure and allergenicity of turbot parvalbumin mediated by caffeic acid. J. Sci. Food Agric. 2019, 99, 3501–3508. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; Lu, F.P.; Liu, Y.H. A Review of the mechanism, properties, and applications of hydrogels prepared by enzymatic cross-linking. J. Agric. Food Chem. 2023, 71, 10238–10249. [Google Scholar] [CrossRef]
- Vasic, K.; Knez, Z.; Leitgeb, M. Transglutaminase in foods and biotechnology. Int. J. Mol. Sci. 2023, 24, 12402. [Google Scholar] [CrossRef]
- Xu, Y.; Ahmed, I.; Zhao, Z.; Lv, L. A comprehensive review on glycation and its potential application to reduce food allergenicity. Crit. Rev. Food Sci. Nutr. 2024, 64, 12184–12206. [Google Scholar] [CrossRef]
- Han, X.Y.; Yang, H.; Rao, S.T.; Liu, G.Y.; Hu, M.J.; Zeng, B.C.; Cao, M.J.; Liu, G.M. The Maillard reaction reduced the sensitization of tropomyosin and arginine kinase from Scylla paramamosain, Simultaneously. J. Agric. Food Chem. 2018, 66, 2934–2943. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Xiao, H.; Zhang, X.F.; Zhou, P. Insight into the effects of deglycosylation and glycation of shrimp tropomyosin on in vivo allergenicity and mast cell function. Food Funct. 2019, 10, 3934–3941. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.T.; Ahmed, I.; Qu, X.; Ju, G.; Yang, N.; Guo, Y.; Li, Z. Effect of the structure and potential allergenicity of glycated tropomyosin, the shrimp allergen. Int. J. Food Sci. Technol. 2022, 57, 1782–1790. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, J.; Xu, L.; Wang, H.; Zhang, Z.; Lin, H.; Li, Z. Insights into the mechanism underlying the influence of glycation with different saccharides and temperatures on the IgG/IgE binding ability, immunodetection, in vitro digestibility of shrimp (Litopenaeus vannamei) tropomyosin. Foods 2023, 12, 3049. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.-W.; Liu, Y.-Y.; Chen, F.; Hu, J.-W.; Cao, M.-J.; Su, W.-J.; Liu, G.-M. Purification, characterization and immunoreactivity of tropomyosin, the allergen in Octopus fangsiao. Process Biochem. 2014, 49, 1747–1756. [Google Scholar] [CrossRef]
- Bai, T.L.; Han, X.Y.; Li, M.S.; Yang, Y.; Liu, M.; Ji, N.-R.; Yu, C.-C.; Lai, D.; Cao, M.-J.; Liu, G.-M. Effects of the Maillard reaction on the epitopes and immunoreactivity of tropomyosin, a major allergen in Chlamys nobilis. Food Funct. 2021, 12, 5096–5108. [Google Scholar] [CrossRef]
- Ji, N.R.; Yu, C.C.; Han, X.; He, X.; Kang, S.; Bai, T.; Liu, H.; Chen, G.; Cao, M.; Liu, G. Effects of three processing technologies on the structure and immunoreactivity of α-tropomyosin from Haliotis discus hannai. Food Chem. 2023, 405, 134947. [Google Scholar] [CrossRef]
- Wu, Y.T.; Lu, Y.Y.; Huang, Y.; Lin, H.; Chen, G.; Chen, Y.; Li, Z. Glycosylation reduces the allergenicity of turbot (Scophthalmus maximus) parvalbumin by regulating digestibility, cellular mediators release and Th1/Th2 immunobalance. Food Chem. 2022, 382, 132574. [Google Scholar] [CrossRef]
- Ahmed, I.; Chen, H.; Li, J.L.; Wang, B.; Li, Z.; Huang, G. Enzymatic crosslinking and food allergenicity: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5856–5879. [Google Scholar] [CrossRef]
- Yuan, F.Z.; Lv, L.T.; Li, Z.X.; Mi, N.; Chen, H.; Lin, H. Effect of transglutaminase-catalyzed glycosylation on the allergenicity and conformational structure of shrimp (Metapenaeus ensis) tropomyosin. Food Chem. 2017, 219, 215–222. [Google Scholar] [CrossRef]
- Li, D.X.; He, X.R.; Li, F.; Yang, Y.; Liu, M.; Liu, Q.; Luo, L.; Chen, G.; Liu, G. Effect of transglutaminase-catalyzed glycosylation on the allergenicity of tropomyosin in the Perna viridis food matrix. Food Funct. 2024, 15, 9136–9148. [Google Scholar] [CrossRef]
- Gunal-Koroglu, D.; Karabulut, G.; Ozkan, G.; Yilmaz, H.; Gultekin-Subasi, B.; Capanoglu, E. Allergenicity of alternative proteins: Reduction mechanisms and processing strategies. J. Agric. Food Chem. 2025, 73, 7522–7546. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Salam, M.H.; El-Shibiny, S. Reduction of Milk protein antigenicity by enzymatic hydrolysis and fermentation. A review. Food Rev. Int. 2021, 37, 276–295. [Google Scholar] [CrossRef]
- Untersmayr, E.; Poulsen, L.K.; Platzer, M.H.; Pedersen, M.H.; Boltz-Nitulescu, G.; Skov, P.S.; Jensen-Jarolim, E. The effects of gastric digestion on codfish allergenicity. J. Allergy Clin. Immunol. 2005, 115, 377–382. [Google Scholar] [CrossRef]
- Liu, G.M.; Huang, Y.Y.; Cai, Q.-F.; Weng, W.-Y.; Su, W.-J.; Cao, M.-J. Comparative study of in vitro digestibility of major allergen, tropomyosin and other proteins between Grass prawn (Penaeus monodon) and Pacific white shrimp (Litopenaeus vannamei). J. Sci. Food Agric. 2011, 91, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Gamez, C.; Paz Zafra, M.; Sanz, V.; Mazzeo, C.; Dolores Ibanez, M.; Sastre, J.; del Pozo, V. Simulated gastrointestinal digestion reduces the allergic reactivity of shrimp extract proteins and tropomyosin. Food Chem. 2015, 173, 475–481. [Google Scholar] [CrossRef]
- Keshavarz, B.; Rao, Q.C.; Jiang, X.; Hsieh, Y.-H.P. Immunochemical analysis of pepsin-digested fish tropomyosin. Food Control 2020, 118, 107427. [Google Scholar] [CrossRef]
- Garcia-Moreno, P.J.; Perez-Galvez, R.; Javier Espejo-Carpio, F.; Ruiz-Quesada, C.; Perez-Morilla, A.I.; Martinez-Agustin, O.; Guadix, A.; Guadix, E.M. Functional, bioactive and antigenicity properties of blue whiting protein hydrolysates: Effect of enzymatic treatment and degree of hydrolysis. J. Sci. Food Agric. 2017, 97, 299–308. [Google Scholar] [CrossRef]
- Wu, M.-X.; Huan, F.; Gao, S.; Ni, L.-N.; Gu, Y.; Han, T.-J.; Liu, M.; Lai, D.; Liu, Q.-M.; Liu, G.-M. Preparation of the hypoallergenic enzymatic hydrolyzate of Cra a 4 with the potential to induce immune tolerance. J. Agric. Food Chem. 2025, 73, 4299–4308. [Google Scholar] [CrossRef]
- Pi, X.W.; Yang, Y.L.; Sun, Y.X.; Cui, Q.; Wan, Y.; Fu, G.; Chen, H.; Cheng, J. Recent advances in alleviating food allergenicity through fermentation. Crit. Rev. Food Sci. Nutr. 2022, 62, 7255–7268. [Google Scholar] [CrossRef]
- Mejrhit, N.; Azdad, O.; Aarab, L. Effect of industrial processing on the IgE reactivity of three commonly consumed Moroccan fish species in Fez region. Eur. Ann. Allergy Clin. Immunol. 2018, 50, 202–210. [Google Scholar] [CrossRef]
- Zou, H.; Zhao, J.; Qin, Z.; Li, Z.; Zhang, Z.; Lin, H.; Wang, H. Influence of fermentation by Lactobacillus helveticus on the immunoreactivity of Atlantic Cod Allergens. J. Agric. Food Chem. 2023, 71, 10144–10154. [Google Scholar] [CrossRef]
- Zhu, Y.D.; Gao, L.; Xie, G.J.; Yang, F.; Gao, P.; Yu, D.; Yu, P.; Jiang, Q.; Xu, Y.; Xia, W. Effect of fermentation on immunological properties of allergens from black carp (Mylopharyngodon piceus) sausages. Int. J. Food Sci. Technol. 2020, 55, 3162–3172. [Google Scholar] [CrossRef]
- Amalia, U.; Shimizu, Y.; Saeki, H. Variation in shrimp tropomyosin allergenicity during the production of Terasi, an Indonesian fermented shrimp paste. Food Chem. 2023, 398, 133876. [Google Scholar] [CrossRef] [PubMed]
- Long, F.Y.; Yang, X.; Wang, R.R.; Hu, X.; Chen, F. Effects of combined high pressure and thermal treatments on the allergenic potential of shrimp (Litopenaeus vannamei) tropomyosin in a mouse model of allergy. Innov. Food Sci. Emerg. Technol. 2015, 29, 119–124. [Google Scholar] [CrossRef]
- Liu, M.; Han, T.-J.; Huan, F.; Li, M.-S.; Xia, F.; Yang, Y.; Wu, Y.-H.; Chen, G.-X.; Cao, M.-J.; Liu, G.-M. Effects of thermal processing on the allergenicity, structure, and critical epitope amino acids of crab tropomyosin. Food Funct. 2021, 12, 2032–2043. [Google Scholar] [CrossRef]
- Tai, J.J.; Qiao, D.; Huang, X.; Hu, H.; Li, W.; Liang, X.; Zhang, F.; Lu, Y.; Zhang, H. Structural property, immunoreactivity and gastric digestion characteristics of glycated parvalbumin from mandarin fish (Siniperca chuaisi) during microwave-assisted Maillard reaction. Foods 2023, 12, 52. [Google Scholar] [CrossRef]
- Zou, S.; Wang, F.Q.; Cheng, J.H.; Wong, S.H.D. Mechanism of cold plasma combined with glycation in altering IgE-binding capacity and digestion stability of tropomyosin from shrimp. J. Agric. Food Chem. 2023, 71, 15796–15808. [Google Scholar] [CrossRef]
- Crespo, J.F.; Cabanillas, B. Recent advances in cellular and molecular mechanisms of IgE-mediated food allergy. Food Chem. 2023, 411, 135500. [Google Scholar] [CrossRef]
- Yang, Y.; He, X.; Li, F.; He, S.; Liu, M.; Li, M.; Xia, F.; Su, W.; Liu, G. Animal-derived food allergen: A review on the available crystal structure and new insights into structural epitope. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13340. [Google Scholar] [CrossRef]
- Gou, J.; Liang, R.; Huang, H.; Ma, X. Maillard reaction induced changes in allergenicity of food. Foods 2022, 11, 530. [Google Scholar] [CrossRef]
- Liu, Q.Z.; Lin, S.Y.; Sun, N. How does food matrix components affect food allergies, food allergens and the detection of food allergens? A systematic review. Trends Food Sci. Technol. 2022, 127, 280–290. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Fish Species | Allergen Names in IUIS | MW (kDa) |
|---|---|---|---|
| β-Parvalbumin | Gadus callarias | Gad c 1 | 12 |
| Gadus morhua | Gad m 1 | 12 | |
| Crocodylus porosus | Cro p 1 | 11.6 | |
| Clupea harengus | Clu h 1 | 12 | |
| Ctenopharyngodon idella | Cten i 1 | 9 | |
| Cyprinus carpio | Cyp c 1 | 12 | |
| Lates calcarifer | Lat c 1 | 11.5 | |
| Lateolabrax maculatus | Late m 1 | 12 and 13 | |
| Lepidorhombus whiffiagonis | Lep w 1 | 11.5 | |
| Oncorhynchus mykiss | Onc m 1 | 12 | |
| Pangasianodon hypophthalmus | Pan h 1 | 11 | |
| Rastrelliger kanagurta | Ras k 1 | 11.3 | |
| Salmo salar | Sal s 1 | 12 | |
| Sardinops sagax | Sar sa 1 | 12 | |
| Scomber scombrus | Sco s 1 | 12 | |
| Sebastes marinus | Seb m 1 | 11 | |
| Solea solea | Sole s 1 | 11~14 | |
| Thunnus albacares | Thu a 1 | 11 | |
| Trichiurus lepturus | Ttic l 1 | 11 | |
| Xiphias gladius | Xip g 1 | 11.5 | |
| α-Parvalbumin | Crocodylus porosus | Cro p 2 | 13 |
| β-Enolase | Gadus morhua | Gad m 2 | 47.3 |
| Cyprinus carpio | Cyp c 2 | 47 | |
| Pangasianodon hypophthalmus | Pan h 2 | 50 | |
| Salmo salar | Sal s 2 | 47.3 | |
| Thunnus albacares | Thu a 2 | 50 | |
| Aldolase A | Gadus morhua | Gad m 3 | 40 |
| Pangasianodon hypophthalmus | Pan h 3 | 40 | |
| Salmo salar | Sal s 3 | 40 | |
| Thunnus albacares | Thu a 3 | 40 | |
| Tropomyosin | Salmo salar | Sal s 4 | 37 |
| Oreochromis mossambicus | Ore m 4 | 33 | |
| Pangasianodon hypophthalmus | Pan h 4 | 35 | |
| β-Prime-component of vitellogenin | Oncorhynchus keta | Onc k 5 | 18 |
| α-Collagen | Lates calcarifer | Lat c 6 | 130~1140 |
| Salmo salar | Sal s 6 | 130~140 | |
| Creatine kinase | Pangasianodon hypophthalmus | Pan h 7 | 43 |
| Salmo salar | Sal s 7 | 43 | |
| Triosephosphate isomerase | Pangasianodon hypophthalmus | Pan h 8 | 25 |
| Salmo salar | Sal s 8 | 25 | |
| Pyruvate kinase PKM-like | Pangasianodon hypophthalmus | Pan h 9 | 65 |
| L-lactate dehydrogenase | Pangasianodon hypophthalmus | Pan h 10 | 34 |
| Glucose 6-phosphate isomerase | Pangasianodon hypophthalmus | Pan h 11 | 60 |
| Glyceraldehyde-3-phosphate dehydrogenase | Pangasianodon hypophthalmus | Pan h 13 | 36 |
| Protein | MW (kDa) | Species | Biological Function | Remark |
|---|---|---|---|---|
| β-Parvalbumin | 11~14 | Fish | Calcium-binding protein |
|
| α-Parvalbumin | ~13 | Fish | Calcium-binding protein |
|
| β-Enolase | 47~50 | Fish | Glycolytic enzyme |
|
| Aldolase A | 40 | Fish | Glycolytic enzyme |
|
| Tropomyosin | 33~40 | Fish Crustacean Mollusk | Actin-binding protein |
|
| Collagen | 130~140 | Fish | Structural protein |
|
| Pyruvate kinase PKM-like | 65 | Fish Crustacean | A regulation enzyme involved in glycolysis |
|
| Triosephosphate isomerase | 25~28 | Fish Crustacean Mollusk | A key enzyme involved in glycolysis |
|
| Creatine kinase | 43 | Fish | Phosphoryl transfer enzymes |
|
| L-lactate dehydrogenase | 34 | Fish | An enzyme involved in glycolysis | - |
| Glucose 6-phosphate isomerase | 60 | Fish | A housekeeping enzyme of glycolysis and gluconeogenesis | - |
| Glyceraldehyde-3-phosphate dehydrogenase | 36 | Fish | - | - |
| β-Prime-component of vitellogenin | 18 | Fish | - | - |
| Arginine kinase | 38~45 | Crustacean Mollusk | Phosphoryl transfer enzymes |
|
| Paramyosin | 99 | Mollusk | Actin-binding protein |
|
| Myosin light chain 2 | 18~23 | Crustacean | Actin-binding protein | - |
| Myosin light chain 1 | 17.5~18 | Crustacean | Actin-binding protein |
|
| Sarcoplasmic calcium-binding protein | 20~25 | Fish Crustacean Mollusk | Calcium-binding protein |
|
| Filamin C | ~90 | Crustacean | Actin-binding protein |
|
| Troponin C | 16.8~21 | Crustacean | Actin-binding protein |
|
| Troponin I | ~30 | Crustacean | Actin-binding protein | - |
| Hemocyanin | 76 | Crustacean | - |
|
| Ovary development-related protein | 28.2 | Crustacean | - | - |
| Cytoplasmic fatty acid-binding protein | 15~20 | Crustacean | Lipid binding protein superfamily |
|
| Glycogen phosphorylase-like protein | 95 | Crustacean | - | - |
| Protein | Shellfish Species | Allergen Names in IUIS | MW (kDa) |
|---|---|---|---|
| Tropomyosin | Charybdis feriatus | Cha f 1 | 34 |
| Crangon crangon | Cra c 1 | ~38 | |
| Crassostrea angulata | Cra a 1 | 38 | |
| Crassostrea gigas | Cra g 1 | 38 | |
| Exopalaemon modestus | Exo m 1 | 38 | |
| Haliotis laevigata x Haliotis rubra | Hal l 1 | 33.4 | |
| Helix aspersa | Hel as 1 | 36 | |
| Homarus americanus | Hom a 1 | 34 | |
| Litopenaeus vannamei | Lit v 1 | 36 | |
| Macrobrachium rosenbergii | Mac r 1 | 37 | |
| Melicertus latisulcatus | Mel l 1 | 38 | |
| Metapenaeus ensis | Met e 1 | 34 | |
| Pandalus borealis | Pan b 1 | 37 | |
| Panulirus stimpsoni | Pan s 1 | 34 | |
| Penaeus aztecus | Pen a 1 | 36 | |
| Penaeus indicus | Pen i 1 | 34 | |
| Penaeus monodon | Pen m 1 | 38 | |
| Portunus pelagicus | Por p 1 | 36 | |
| Procambarus clarkii | Pro c 1 | 36 | |
| Saccostrea glomerata | Sac g 1 | 38 | |
| Scylla paramamosain | Scy p 1 | 38 | |
| Todarodes pacificus | Tod p 1 | 15 | |
| Arginine kinase | Callinectes bellicosus | Cal b 2 | 40 |
| Crassostrea angulata | Cra a 2 | 38~41 | |
| Crangon crangon | Cra c 2 | ~45 | |
| Litopenaeus vannamei | Lit v 2 | 40 | |
| Macrobrachium rosenbergii | Mac r 2 | 40 | |
| Penaeus monodon | Pen m 2 | 40 | |
| Procambarus clarkii | Pro c 2 | 40 | |
| Scylla paramamosain | Scy p 2 | 40 | |
| Ovary development-related protein | Eriocheir sinensis | Eri s 2 | 28.2 |
| Paramyosin | Rapana venosa | Rap v 2 | 99 |
| Myosin light chain 2 | Homarus americanus | Hom a 3 | ~23 |
| Litopenaeus vannamei | Lit v 3 | 20 | |
| Penaeus monodon | Pen m 3 | 20 | |
| Scylla paramamosain | Scy p 3 | 18 | |
| Myosin light chain 1 | Artemia franciscana | Art fr 5 | ~17.5 |
| Crangon crangon | Cra c 5 | ~17.5 | |
| Procambarus clarkii | Pro c 5 | 18 | |
| Sarcoplasmic calcium-binding protein | Crangon crangon | Cra c 4 | ~25 |
| Crassostrea angulata | Cra a 4 | 20 | |
| Litopenaeus vannamei | Lit v 4 | 20 | |
| Penaeus monodon | Pen m 4 | 20 | |
| Pontastacus leptodactylus | Pon l 4 | ~24 | |
| Portunus trituberculatus | Por t 4 | 22 | |
| Scylla paramamosain | Scy p 4 | 20 | |
| Troponin C | Crangon crangon | Cra c 6 | ~21 |
| Homarus americanus | Hom a 6 | ~20 | |
| Penaeus monodon | Pen m 6 | 16.8 | |
| Troponin I | Pontastacus leptodactylus | Pon l 7 | ~30 |
| Hemocyanin | Penaeus monodon | Pen m 7 | 76 |
| Triosephosphate isomerase | Archaeopotamobius sibiriensis | Arc s 8 | ~28 |
| Crangon crangon | Cra c 8 | ~28 | |
| Penaeus monodon | Pen m 8 | 27 | |
| Procambarus clarkii | Pro c 8 | 28 | |
| Scylla paramamosain | Scy p 8 | 28 | |
| Filamin C | Scylla paramamosain | Scy p 9 | 90 |
| Cytoplasmic fatty acid-binding protein | Litopenaeus vannamei | Lit v 13 | 15 |
| Penaeus monodon | Pen m 13 | 20 | |
| Glycogen phosphorylase-like protein | Penaeus monodon | Pen m 14 | 95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zhang, Y.; He, X.; Huan, F.; Chen, J.; Liu, M.; He, S.; Gu, S.; Liu, G. An Overview of Seafood Allergens: Structure–Allergenicity Relationship and Allergenicity Elimination Processing Techniques. Foods 2025, 14, 2241. https://doi.org/10.3390/foods14132241
Yang Y, Zhang Y, He X, Huan F, Chen J, Liu M, He S, Gu S, Liu G. An Overview of Seafood Allergens: Structure–Allergenicity Relationship and Allergenicity Elimination Processing Techniques. Foods. 2025; 14(13):2241. https://doi.org/10.3390/foods14132241
Chicago/Turabian StyleYang, Yang, Yehao Zhang, Xinrong He, Fei Huan, Jinli Chen, Meng Liu, Siyang He, Shinong Gu, and Guangming Liu. 2025. "An Overview of Seafood Allergens: Structure–Allergenicity Relationship and Allergenicity Elimination Processing Techniques" Foods 14, no. 13: 2241. https://doi.org/10.3390/foods14132241
APA StyleYang, Y., Zhang, Y., He, X., Huan, F., Chen, J., Liu, M., He, S., Gu, S., & Liu, G. (2025). An Overview of Seafood Allergens: Structure–Allergenicity Relationship and Allergenicity Elimination Processing Techniques. Foods, 14(13), 2241. https://doi.org/10.3390/foods14132241