
Prebiotic Effect of Oxidized Hydroxypropyl Starch via In Vitro and In Vivo


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Methods
2.2.1. Preparation of Hydroxypropyl Starch (HPS)
2.2.2. Preparation of Oxidized Hydroxypropyl Starch (OHS)
2.2.3. Crystalline Structure Determination
2.2.4. FT-IR Spectra Determination
2.2.5. Measurement of Changes in Thermodynamic Properties
2.2.6. Determination of Carboxyl Group Content
2.2.7. In Vitro Digestion Characterization
2.2.8. In Vitro Fecal Fermentation Experiments
2.2.9. In Vivo Enzymolysis of Starch
2.2.10. Analysis of Metabolites by GC-MS and LC-MS Measurements
2.2.11. 16S rRNA Gene and Bioinformatics Analysis
2.2.12. Data Processing and Analysis
3. Results and Discussion
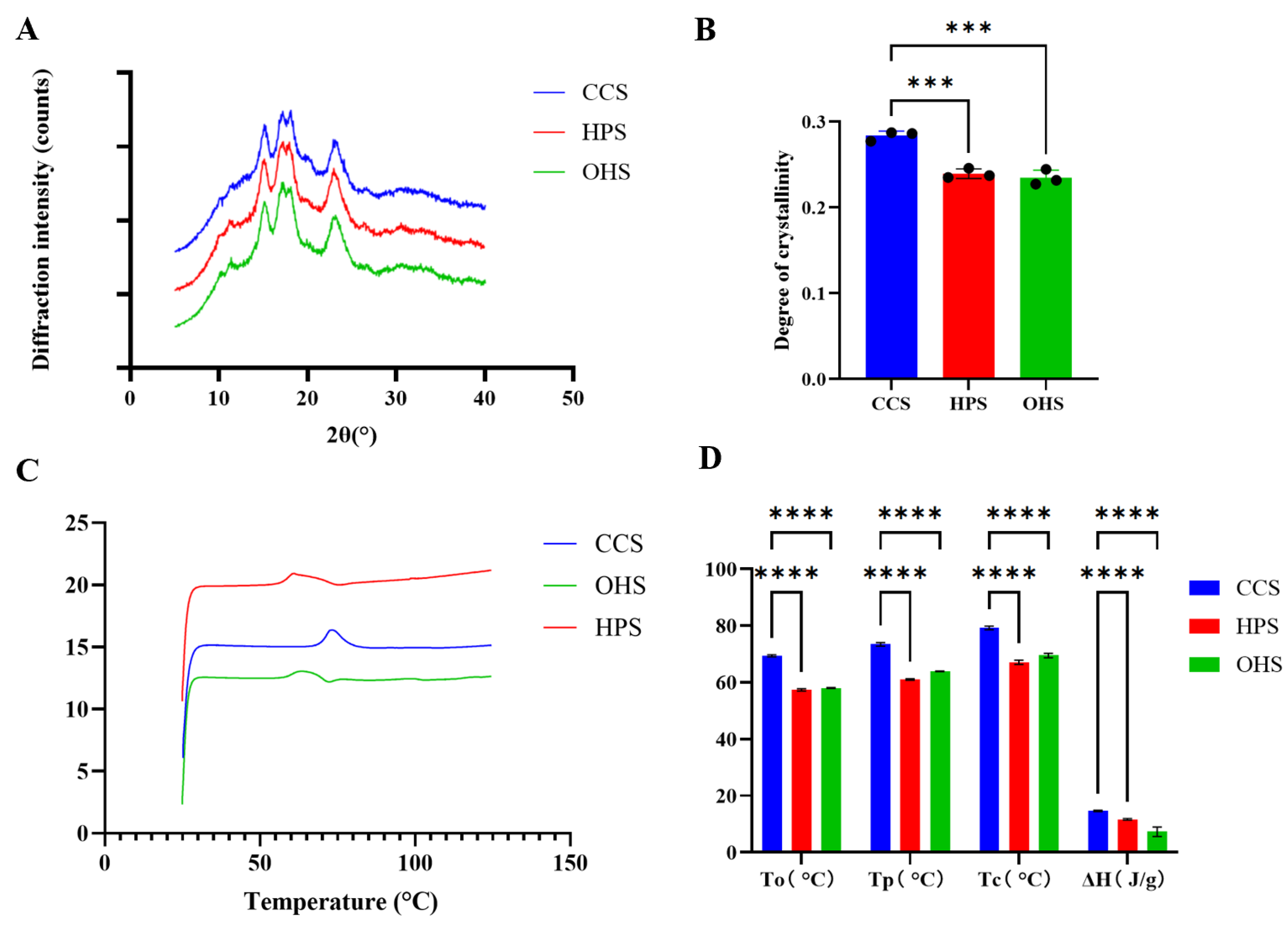
3.1. Preparation of Resistant Starch and Characterization of Its Structural Properties
3.1.1. Crystal Structures and Pasting Properties
3.1.2. FT-IR Spectra and In Vitro Digestion Results Analysis
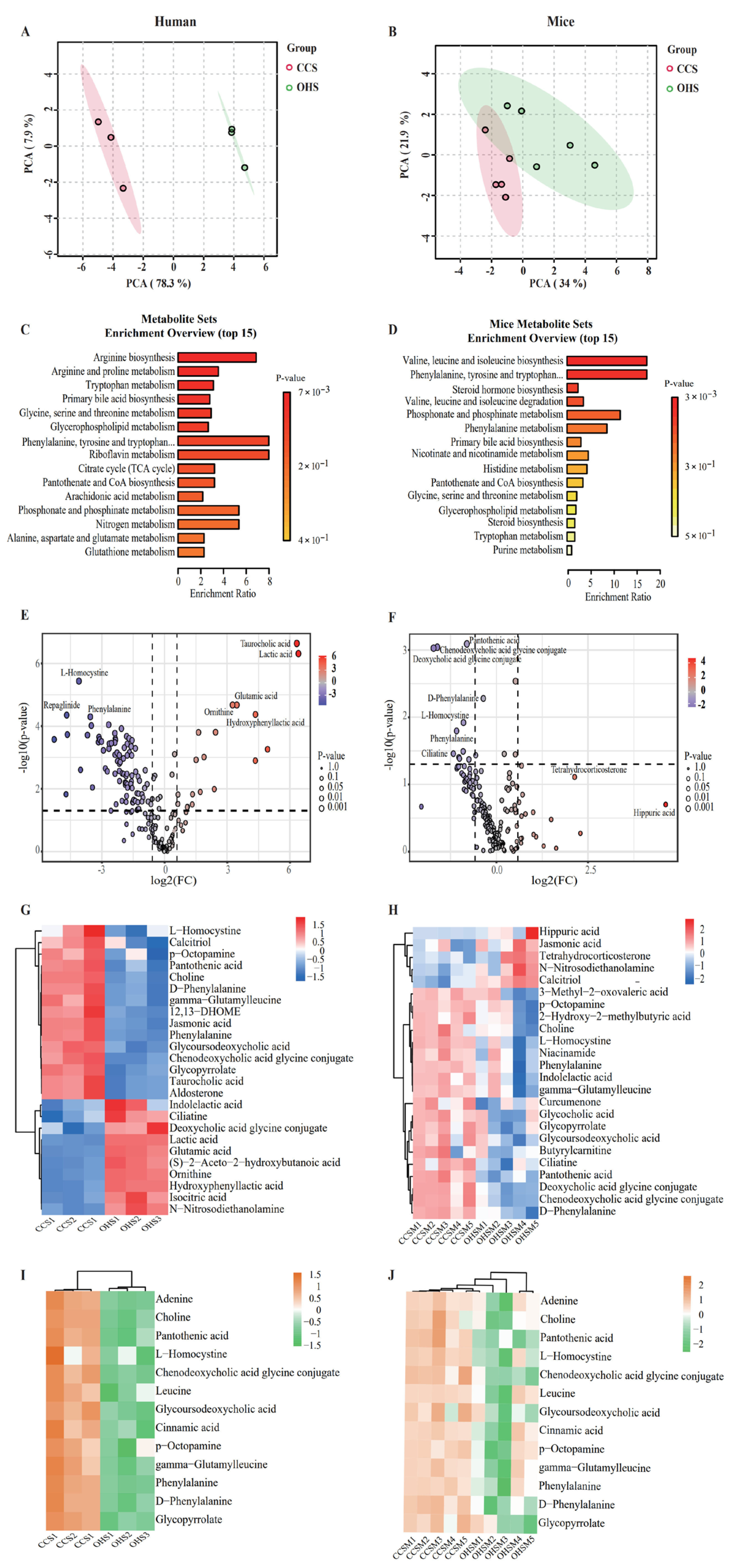
3.2. Analysis of Intestinal Microbiota Results
3.3. Predicting Functional Changes in the Microbiota
3.4. Validation of Microbial Metabolites
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Milanezzi, G.C.; Silva, E.K. Pulsed electric field-induced starch modification for food industry applications: A review of native to modified starches. Carbohydr. Polym. 2024, 348, 122793. [Google Scholar] [CrossRef] [PubMed]
- Compart, J.; Singh, A.; Fettke, J.; Apriyanto, A. Customizing starch properties: A review of starch modifications and their applications. Polymers 2023, 15, 3491. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Wang, W.; Fan, Z.; Li, B.; Wei, Y.; Yu, R.; Pan, T.; Wang, N.; Lu, W.; Fang, Z. Gut microbes mediate prebiotic-like effects of resistant starch. Food Biosci. 2024, 61, 104627. [Google Scholar] [CrossRef]
- Tekin, T.; Dincer, E. Effect of resistant starch types as a prebiotic. Appl. Microbiol. Biotechnol. 2023, 107, 491–515. [Google Scholar] [CrossRef]
- Zhu, Y.; Dong, L.; Huang, L.; Shi, Z.; Dong, J.; Yao, Y.; Shen, R. Effects of oat β-glucan, oat resistant starch, and the whole oat flour on insulin resistance, inflammation, and gut microbiota in high-fat-diet-induced type 2 diabetic rats. J. Funct. Foods 2020, 69, 103939. [Google Scholar] [CrossRef]
- Fassarella, M.; Blaak, E.E.; Penders, J.; Nauta, A.; Smidt, H.; Zoetendal, E.G. Gut microbiome stability and resilience: Elucidating the response to perturbations in order to modulate gut health. Gut 2021, 70, 595–605. [Google Scholar] [CrossRef]
- Westerbeke, F.H.M.; Attaye, I.; Rios-Morales, M.; Nieuwdorp, M. Glycaemic sugar metabolism and the gut microbiota: Past, present and future. FEBS J. 2025, 292, 1421–1436. [Google Scholar] [CrossRef]
- Nishida, A.; Ando, Y.; Kimura, I.; Miyamoto, J. Involvement of gut microbial metabolites derived from diet on host energy homeostasis. Int. J. Mol. Sci. 2022, 23, 5562. [Google Scholar] [CrossRef]
- Wen, J.-J.; Li, M.-Z.; Hu, J.-L.; Tan, H.-Z.; Nie, S.-P. Resistant starches and gut microbiota. Food Chem. 2022, 387, 132895. [Google Scholar] [CrossRef]
- Park, M.H.; Kim, M. Physicochemical properties of hydroxypropylated apios starches. Prev. Nutr. Food Sci. 2020, 25, 286. [Google Scholar] [CrossRef]
- Zhang, Y.; Lv, J.; Qiu, Z.; Chen, L. Influence of carboxyl content on the rheological properties and printability of oxidized starch for 3D printing applications. Int. J. Biol. Macromol. 2025, 289, 138794. [Google Scholar] [CrossRef] [PubMed]
- Sares, L.A.C.; Año, A.C.; Pérez, M.A.G.; Peña, N.D.J.; Román, J.V.M. Coating of oxidized banana starch and olive oil for the preservation of cherry tomatoes (Solanum lycopersicum cv. Cerasiforme). Rev. Fac. Nac. Agron. Medellín 2024, 77, 10853–10864. [Google Scholar] [CrossRef]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally im-portant starch fractions. Eur. J. Clin. Nutr. 1992, 46, S33–S50. [Google Scholar] [PubMed]
- Liu, C.; Du, P.; Cheng, Y.; Guo, Y.; Hu, B.; Yao, W.; Zhu, X.; Qian, H. Study on fecal fermentation characteristics of aloe polysaccharides in vitro and their predictive modeling. Carbohydr. Polym. 2021, 256, 117571. [Google Scholar] [CrossRef]
- Shi, J.; Wang, Y.; Jiang, F.; Liu, Y.; Xu, Y.-J. The effect of krill oil on longevity and locomotion: A pilot study. Mol. Omics 2022, 18, 206–213. [Google Scholar] [CrossRef]
- Liu, C.; Guo, Y.; Cheng, Y.; Qian, H. A colon-targeted delivery system of torularhodin encapsulated in electrospinning micro-spheres, and its co-metabolic regulation mechanism of gut microbiota. Food Hydrocoll. 2023, 135, 108189. [Google Scholar] [CrossRef]
- Xu, Q.; Ma, R.; Zhan, J.; Lu, X.; Liu, C.; Tian, Y. Acylated resistant starches: Changes in structural properties during digestion and their fermentation characteristics. Food Hydrocoll. 2023, 139, 108578. [Google Scholar] [CrossRef]
- Kuakpetoon, D.; Wang, Y.J. Characterization of different starches oxidized by hypochlorite. Starch-Stärke 2001, 53, 211–218. [Google Scholar] [CrossRef]
- Cooke, D.; Gidley, M.J. Loss of crystalline and molecular order during starch gelatinisation: Origin of the enthalpic transition. Carbohydr. Res. 1992, 227, 103–112. [Google Scholar] [CrossRef]
- Zhou, Y.-L.; Cui, L.-H.; You, X.-Y.; Jiang, Z.-H.; Qu, W.-H.; Liu, P.-D.; Ma, D.-Y.; Cui, Y.-Y. Effects of repeated and continuous dry heat treatments on the physicochemical and struc-tural properties of quinoa starch. Food Hydrocoll. 2021, 113, 106532. [Google Scholar] [CrossRef]
- Perera, C.; Hoover, R. Influence of hydroxypropylation on retrogradation properties of native, defatted and heat-moisture treated potato starches. Food Chem. 1999, 64, 361–375. [Google Scholar] [CrossRef]
- Woggum, T.; Sirivongpaisal, P.; Wittaya, T. Characteristics and properties of hydroxypropylated rice starch based biodegradable films. Food Hydrocoll. 2015, 50, 54–64. [Google Scholar] [CrossRef]
- Sánchez-Rivera, M.; García-Suárez, F.; del Valle, M.V.; Gutierrez-Meraz, F.; Bello-Pérez, L. Partial characterization of banana starches oxidized by different levels of so-dium hypochlorite. Carbohydr. Polym. 2005, 62, 50–56. [Google Scholar] [CrossRef]
- Sangseethong, K.; Termvejsayanon, N.; Sriroth, K. Characterization of physicochemical properties of hypochlorite-and peroxide-oxidized cassava starches. Carbohydr. Polym. 2010, 82, 446–453. [Google Scholar] [CrossRef]
- Bakouri, H.; Guemra, K. Etherification and cross-linking effect on physicochemical properties of Zea mays starch executed at different sequences in 1-butyl-3-methylimidazolium chloride [BMIM] Cl ionic liquid media. Int. J. Biol. Macromol. 2019, 125, 1118–1127. [Google Scholar] [CrossRef]
- Rutkaitė, R.; Baranauskienė, R.; Pečiulytė, L.; Pukalskienė, M.; Venskutonis, P.R. Preparation and properties of propylene oxide and octenylsuccinic anhydride modified potato starches. J. Food Sci. Technol. 2016, 53, 4187–4196. [Google Scholar] [CrossRef]
- Ye, S.; Qiu-Hua, W.; Xue-Chun, X.; Wen-Yong, J.; Shu-Cai, G.; Hai-Feng, Z. Oxidation of cornstarch using oxygen as oxidant without catalyst. LWT-Food Sci. Technol. 2011, 44, 139–144. [Google Scholar] [CrossRef]
- Sumardiono, S.; Jos, B.; Pudjihastuti, I.; Yafiz, A.M.; Rachmasari, M.; Cahyono, H. Physicochemical properties of sago ozone oxidation: The effect of reaction time, acidity, and concentration of starch. Foods 2021, 10, 1309. [Google Scholar] [CrossRef]
- Jiang, H.; Feng, Y.; Jane, J.-L.; Yang, Y. Progress in understanding resistant-starch formation in hydroxypropyl starch: A minireview. Food Hydrocoll. 2024, 149, 109628. [Google Scholar] [CrossRef]
- Liu, C.; Jiang, W.; Yang, F.; Cheng, Y.; Guo, Y.; Yao, W.; Zhao, Y.; Qian, H. The combination of microbiome and metabolome to analyze the cross-cooperation mechanism of Echinacea purpurea polysaccharide with the gut microbiota in vitro and in vivo. Food Funct. 2022, 13, 10069–10082. [Google Scholar] [CrossRef]
- Ladewig, L.; Tufail, M.A.; Kinfu, B.M.; Fokt, H.; Baines, J.F.; Schmitz, R.A. Exploring new Bacteroidota strains: Functional diversity and probiotic characteristics. bioRxiv 2025. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The firmicutes/bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Molinas-Vera, M.; Ferreira-Sanabria, G.; Peña, P.; Sandoval-Espinola, W.J. The Paraguayan gut microbiome contains high abundance of the phylum Actino-bacteriota and reveals the influence of health and lifestyle factors. Gut Microbes Rep. 2024, 1, 1–16. [Google Scholar]
- Zhang, S.; Nie, Q.; Sun, Y.; Zuo, S.; Chen, C.; Li, S.; Yang, J.; Hu, J.; Zhou, X.; Yu, Y.; et al. Bacteroides uniformis degrades β-glucan to promote Lactobacillus johnsonii improving in-dole-3-lactic acid levels in alleviating colitis. Microbiome 2024, 12, 177. [Google Scholar] [CrossRef]
- Morita, H.; Kano, C.; Ishii, C.; Kagata, N.; Ishikawa, T.; Hirayama, A.; Uchiyama, Y.; Hara, S.; Nakamura, T.; Fukuda, S. Bacteroides uniformis and its preferred substrate, α-cyclodextrin, enhance endurance exer-cise performance in mice and human males. Sci. Adv. 2023, 9, eadd2120. [Google Scholar] [CrossRef]
- Ezeji, J.C.; Sarikonda, D.K.; Hopperton, A.; Erkkila, H.L.; Cohen, D.E.; Martinez, S.P.; Cominelli, F.; Kuwahara, T.; Dichosa, A.E.K.; Good, C.E.; et al. Parabacteroides distasonis: Intriguing aerotolerant gut anaerobe with emerging antimicrobial resistance and pathogenic and probiotic roles in human health. Gut Microbes 2021, 13, 1922241. [Google Scholar] [CrossRef]
- Lv, X.; Zhan, L.; Ye, T.; Xie, H.; Chen, Z.; Lin, Y.; Cai, X.; Yang, W.; Liao, X.; Liu, J.; et al. Gut commensal Agathobacter rectalis alleviates microglia-mediated neuroinflammation against pathogenesis of Alzheimer disease. iScience 2024, 27, 111116. [Google Scholar] [CrossRef]
- Huber-Ruano, I.; Calvo, E.; Mayneris-Perxachs, J.; Rodríguez-Peña, M.-M.; Ceperuelo-Mallafré, V.; Cedó, L.; Núñez-Roa, C.; Miro-Blanch, J.; Arnoriaga-Rodríguez, M.; Balvay, A.; et al. Orally administered Odoribacter laneus improves glucose control and inflammatory profile in obese mice by depleting circulating succinate. Microbiome 2022, 10, 135. [Google Scholar] [CrossRef]
- Weng, H.; Deng, L.; Wang, T.; Xu, H.; Wu, J.; Zhou, Q.; Yu, L.; Chen, B.; Huang, L.; Qu, Y.; et al. Humid heat environment causes anxiety-like disorder via impairing gut microbiota and bile acid metabolism in mice. Nat. Commun. 2024, 15, 5697. [Google Scholar] [CrossRef]
- Ndeh, D.A.; Nakjang, S.; Kwiatkowski, K.J.; Sawyers, C.; Koropatkin, N.M.; Hirt, R.P.; Bolam, D.N. A Bacteroides thetaiotaomicron genetic locus encodes activities consistent with mucin O-glycoprotein processing and N-acetylgalactosamine metabolism. Nat. Commun. 2025, 16, 3485. [Google Scholar] [CrossRef]
- Porter, N.T.; Larsbrink, J. Investigation and alteration of organic acid synthesis pathways in the mammalian gut symbiont Bacteroides thetaiotaomicron. Microbiol. Spectr. 2022, 10, e02312–e02321. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xu, W.; Dong, W.; Chen, G.; Sun, Y.; Zeng, X. Anti-diabetic effect of dicaffeoylquinic acids is associated with the modulation of gut microbiota and bile acid metabolism. J. Adv. Res. 2024, 72, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hu, B.; Cheng, Y.; Guo, Y.; Yao, W.; Qian, H. In-depth analysis of the mechanisms of aloe polysaccharides on mitigating subacute colitis in mice via microbiota informatics. Carbohydr. Polym. 2021, 265, 118041. [Google Scholar] [CrossRef]
- Wei, Z.; Zhang, W.; Du, M.; Zhong, H.; Fang, X. Widely targeted metabolomic and KEGG analyses of natural deep eutectic solvent-based saponins extraction from Camellia oleifera Abel.: Effects on composition. Food Chem. 2024, 450, 139333. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, Y.; Li, Q.; Li, J.; Zhang, D.; Zhang, Y.; Li, Y.; Li, X.; Xu, P.; Bai, L.; et al. Multi-omics analysis reveals phenylalanine enhance mitochondrial function and hypoxic en-durance via LKB1/AMPK activation. J. Transl. Med. 2024, 22, 920. [Google Scholar] [CrossRef]
- Kim, J.-H.; Jang, H.-J.; Cho, W.-Y.; Yeon, S.-J.; Lee, C.-H. In vitro antioxidant actions of sulfur-containing amino acids. Arab. J. Chem. 2020, 13, 1678–1684. [Google Scholar] [CrossRef]
- Ligi, S.; Ali, A.; Yang, G. Cystathionine gammalyase deficiency exaggerates diethylnitrosamine-induced liver damage in mice. Nitric Oxide 2024, 151, 1–9. [Google Scholar] [CrossRef]
- Fontana, M.; Akyildiz, A.G.; D’alonzo, C.; Giovannercole, F.; Zicchi, A.; Francioso, A.; Capuozzo, E.; De Biase, D. Synthesis and Biological Activity of Homohypotaurine Obtained by the Enzyme-Based Conversion of Homocysteine Sulfinic Acid Using Recombinant Escherichia coli Glutamate Decarboxylase. Molecules 2024, 29, 3985. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Carboxyl Contents (%) |
|---|---|
| HPS | 0.00 |
| OHS | 0.56 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.; Xu, Z.; Fan, Y.; Han, J.; Zhou, L.; Li, H.; Pan, X.; Ma, R.; Liu, C.; Tian, Y. Prebiotic Effect of Oxidized Hydroxypropyl Starch via In Vitro and In Vivo. Foods 2025, 14, 2217. https://doi.org/10.3390/foods14132217
Zheng H, Xu Z, Fan Y, Han J, Zhou L, Li H, Pan X, Ma R, Liu C, Tian Y. Prebiotic Effect of Oxidized Hydroxypropyl Starch via In Vitro and In Vivo. Foods. 2025; 14(13):2217. https://doi.org/10.3390/foods14132217
Chicago/Turabian StyleZheng, Huiwen, Zhipu Xu, Yiwen Fan, Jiazhi Han, Liyang Zhou, Han Li, Xiaohua Pan, Rongrong Ma, Chang Liu, and Yaoqi Tian. 2025. "Prebiotic Effect of Oxidized Hydroxypropyl Starch via In Vitro and In Vivo" Foods 14, no. 13: 2217. https://doi.org/10.3390/foods14132217
APA StyleZheng, H., Xu, Z., Fan, Y., Han, J., Zhou, L., Li, H., Pan, X., Ma, R., Liu, C., & Tian, Y. (2025). Prebiotic Effect of Oxidized Hydroxypropyl Starch via In Vitro and In Vivo. Foods, 14(13), 2217. https://doi.org/10.3390/foods14132217