
Microalgae Bioactives for Functional Food Innovation and Health Promotion



Abstract

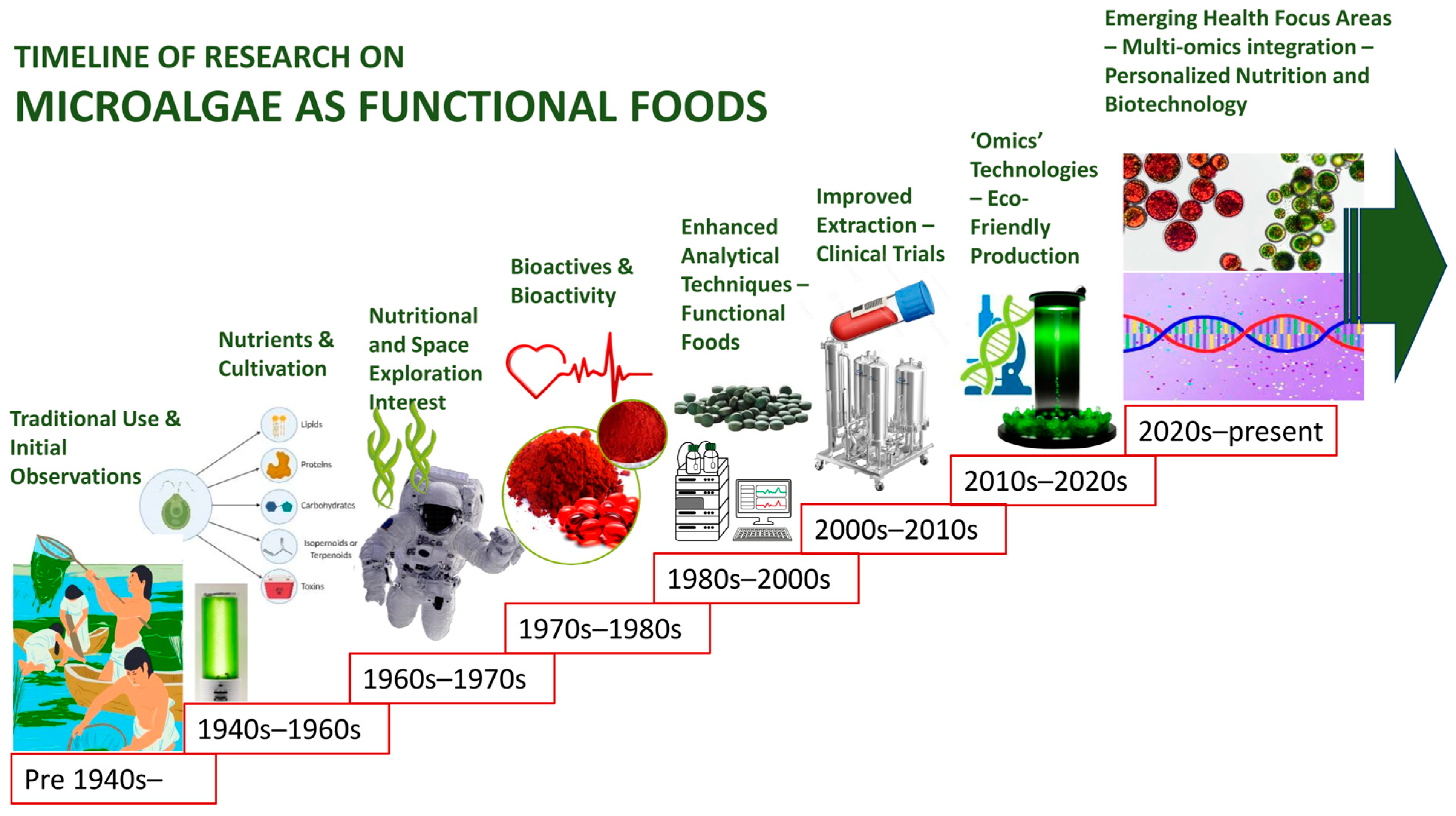
1. Introduction
2. Materials and Methods
3. Nutritional and Functional Composition of Microalgae
3.1. Major Edible Microalgae Species
3.2. Key Bioactive Compounds
3.3. Nutritional Variability and Influence of Cultivation
3.4. Analytical Determination
4. Health-Promoting Properties of Microalgae Bioactives
4.1. Antioxidant Activity and Reduction of Oxidative Stress
4.2. Anti-Inflammatory Effects and Immune Modulation
4.3. Metabolic Regulation: Impacts on Lipid and Glucose Metabolism
4.4. Gut Microbiota Modulation and Digestive Health
5. Evidence from Preclinical and Clinical Studies
5.1. In Vitro and Animal Model Findings
5.2. Key Human Studies and Clinical Trial Outcomes
5.3. Limitations and Gaps in the Existing Research
6. Application in Functional Foods and Nutraceuticals
6.1. Integration of Microalgae into Food Systems
6.2. Bioavailability Considerations and Enhancement Strategies
6.3. Synergistic Effects of Whole Biomass vs. Isolated Compounds
7. Sustainability and Innovation Potential
7.1. Environmental Advantages of Microalgae Cultivation
7.2. Role in Future Food Systems and Dietary Recommendations
8. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ortega, A.; Campos, M. Bioactive Compounds as Therapeutic Alternatives. In Bioactive Compounds; Springer Nature: Cham, Switzerland, 2019; pp. 247–264. [Google Scholar]
- Badhwar, R.; Kaur, G.; Popli, H.; Yadav, D.; Buttar, H. Pathophysiology of Obesity-Related Non-communicable Chronic Diseases and Advancements in Preventive Strategies. In Pathophysiology of Obesity-Induced Health Complications. Advances in Biochemistry in Health and Disease; Springer: Cham, Switzerland, 2020; pp. 317–340. [Google Scholar]
- Rahman, M.; Islam, R.; Rabbi, F.; Islam, M.T.; Sultana, S.; Ahmed, M.; Sehgal, A.; Singh, S.; Sharma, N.; Behl, T. Bioactive Compounds and Diabetes Mellitus: Prospects and Future Challenges. Curr. Pharm. Des. 2022, 28, 1304–1320. [Google Scholar] [CrossRef] [PubMed]
- Ampofo, J.; Abbey, L. Microalgae: Bioactive Composition, Health Benefits, Safety and Prospects as Potential High-Value Ingredients for the Functional Food Industry. Foods 2022, 11, 1744. [Google Scholar] [CrossRef]
- Baldemir, P.; Cakli, S. Sustainable Food Ingredients: Micro-Algae as Source Bioactive Compounds. Food Bull. 2024, 3, 34–40. [Google Scholar] [CrossRef]
- Naik, B.; Mishra, R.; Kumar, V.; Mishra, S.; Gupta, U.; Rustagi, S.; Gupta, A.; Preet, M.S.; Bhatt, S.; Rizwanuddin, S. Micro-algae: Revolutionizing food production for a healthy and sustainable future. J. Agric. Food Res. 2023, 15, 100939. [Google Scholar] [CrossRef]
- Zhu, J.; Xiao, X.; Du, W.; Cai, Y.; Yang, Z.; Yin, Y.; Wakisaka, M.; Wang, J.; Zhou, Z.; Liu, D.; et al. Leveraging microalgae as a sustainable ingredient for meat analogues. Food Chem. 2024, 450, 139360. [Google Scholar] [CrossRef]
- Sandgruber, F.; Gielsdorf, A.; Baur, A.; Schenz, B.; Müller, S.; Schwerdtle, T.; Stangl, G.; Griehl, C.; Lorkowski, S.; Dawczynski, C. Variability in Macro- and Micronutrients of 15 Commercially Available Microalgae Powders. Mar. Drugs 2021, 19, 310. [Google Scholar] [CrossRef] [PubMed]
- Davani, L.; Terenzi, C.; Tumiatti, V.; De Simone, A.; Andrisano, V.; Montanari, S. Integrated analytical approaches for the characterization of Spirulina and Chlorella microalgae. J. Pharm. Biomed. Anal. 2022, 219, 114943. [Google Scholar] [CrossRef]
- Dimopoulou, M.; Kolonas, A.; Stagos, D.; Gortzi, O. A Review of the Sustainability, Chemical Composition, Bioactive Compounds, Antioxidant and Antidiabetic Activity, Neuroprotective Properties, and Health Benefits of Microalgae. Biomass 2025, 5, 11. [Google Scholar] [CrossRef]
- Mendes, A.R.; Spínola, M.P.; Lordelo, M.; Prates, J.A.M. Chemical Compounds, Bioactivities, and Applications of Chlorella vulgaris in Food, Feed and Medicine. Appl. Sci. 2024, 14, 10810. [Google Scholar] [CrossRef]
- Spínola, M.P.; Mendes, A.R.; Prates, J.A.M. Chemical Composition, Bioactivities, and Applications of Spirulina (Limnospira platensis) in Food, Feed, and Medicine. Foods 2024, 13, 3656. [Google Scholar] [CrossRef] [PubMed]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Panaite, T.; Cornescu, G.; Predescu, N.; Cișmileanu, A.; Turcu, R.; Sărăcilă, M.; Șoica, C. Microalgae (Chlorella vulgaris and Spirulina platensis) as a Protein Alternative and Their Effects on Productive Performances, Blood Parameters, Protein Digestibility, and Nutritional Value of Laying Hens’ Egg. Appl. Sci. 2023, 13, 10451. [Google Scholar] [CrossRef]
- Prates, J.A.M. The Role of Meat Lipids in Nutrition and Health: Balancing Benefits and Risks. Nutrients 2025, 17, 350. [Google Scholar] [CrossRef]
- Christaki, E.; Bonos, E.; Florou-Paneri, P. Innovative microalgae pigments as functional ingredients in animal nutrition. Food Res. Int. 2015, 77, 572–577. [Google Scholar]
- Sañé, E.; del Mondo, A.; Ambrosino, L.; Smerilli, A.; Sansone, C.; Brunet, C. The recent advances in microalgal phytosterols: Bioactive ingredients with human health driven potential applications. Food Rev. Int. 2021, 39, 1859–1878. [Google Scholar] [CrossRef]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for high-value products: A way towards green nutraceutical and pharmaceutical applications. Renew. Sustain. Energy Rev. 2019, 112, 574–584. [Google Scholar]
- Morais, M.G.d.; Morais, E.G.d.; Cardias, B.B.; Vaz, B.d.S.; Moreira, J.B.; Mitchell, B.G.; Costa, J.A.V. Chapter 13—Microalgae as a source of sustainable biofuels. In Recent Developments in Bioenergy Research; Gupta, V.K., Treichel, H., Kuhad, R.C., Rodriguez-Cout, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 253–271. [Google Scholar]
- Marques, L.; Domingues, M.R.; da Costa, E.; Abreu, M.H.; Lillebø, A.I.; Calado, R. Screening for Health-Promoting Fatty Acids in Ascidians and Seaweeds Grown under the Influence of Fish Farming Activities. Mar. Drugs 2021, 19, 469. [Google Scholar] [CrossRef]
- Pagels, F.; Amaro, H.M.; Tavares, T.G.; Amil, B.F.; Guedes, A.C. Potential of Microalgae Extracts for Food and Feed Supplementation—A Promising Source of Antioxidant and Anti-Inflammatory Compounds. Life 2022, 12, 1901. [Google Scholar] [CrossRef]
- Chien, Y.-H.; Yu, Y.-H.; Chen, Y.-W. Taiwanese green propolis ameliorates metabolic syndrome via remodeling of white adipose tissue and modulation of gut microbiota in diet-induced obese mice. Biomed. Pharmacother. 2023, 160, 114386. [Google Scholar] [CrossRef]
- Alves, J.L.B.; Costa, P.C.T.D.; Sales, L.C.S.; Silva Luis, C.C.; Bezerra, T.P.T.; Souza, M.L.A.; Costa, B.A.; de Souza, E.L. Shedding light on the impacts of Spirulina platensis on gut microbiota and related health benefits. Crit. Rev. Food Sci. 2025, 65, 2062–2075. [Google Scholar] [CrossRef] [PubMed]
- Ang, W.S.; Kee, P.E.; Lan, J.C.W.; Chen, W.H.; Chang, J.S.; Khoo, K.S. Unveiling the rise of microalgae-based foods in the global market: Perspective views and way forward. Food Biosci. 2025, 66, 105390. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Hayes, M. Red and Green Macroalgae for Fish and Animal Feed and Human Functional Food Development. In Sustainable Seaweed Technologies: Cultivation, Biorefinery, and Applications; Torres, M.D., Kraan, S., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 399–420. [Google Scholar]
- Vieira, M.V.; Pastrana, L.M.; Fuciños, P. Chapter 30—Encapsulation of microalgae-based products for food and feed applications. In Handbook of Food and Feed from Microalgae; Jacob-Lopes, E., Queiroz, M.I., Maroneze, M.M., Zepka, L.Q., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 371–393. [Google Scholar]
- Bernaerts, T.M.M.; Verstreken, H.; Dejonghe, C.; Gheysen, L.; Foubert, I.; Grauwet, T.; Van Loey, A.M. Cell disruption of Nannochloropsis sp. improves in vitro bioaccessibility of carotenoids and ω3-LC-PUFA. J. Funct. Foods 2020, 65, 103770. [Google Scholar] [CrossRef]
- Çelekli, A.; Özbal, B.; Bozkurt, H. Challenges in functional food products with the incorporation of some microalgae. Foods 2024, 13, 725. [Google Scholar] [CrossRef]
- Nethravathy, M.U.; Mehar, J.G.; Mudliar, S.N.; Shekh, A.Y. Recent advances in microalgal bioactives for food, feed, and healthcare products: Commercial potential, market space, and sustainability. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1882–1897. [Google Scholar]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Mehariya, S.; Goswami, R.; Karthikeysan, O.P.; Verma, P. Microalgae for high-value products: A way towards green nutraceutical and pharmaceutical compounds. Chemosphere 2021, 280, 130553. [Google Scholar] [CrossRef]
- Ampofo, J.O.; Abbey, L.D. Microalgae: Bioactive composition, health benefits, safety, and prospects as food and feed. Heliyon 2022, 8, e09531. [Google Scholar]
- Ramos-Romero, S.; Torrella, J.R.; Roura, E. Edible microalgae and their bioactive compounds in the prevention and treatment of metabolic alterations. Int. J. Mol. Sci. 2021, 22, 4415. [Google Scholar] [CrossRef]
- Yang, Y.; Du, L.; Hosokawa, M.; Miyashita, K. Total Lipids Content, Lipid Class and Fatty Acid Composition of Ten Species of Microalgae. J. Oleo Sci. 2020, 69, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Molino, A.; Iovine, A.; Casella, P.; Musmarra, D. Microalgae characterization for consolidated and new application in human food, animal feed and nutraceuticals. Int. J. Environ. Res. Public Health 2018, 15, 2436. [Google Scholar] [CrossRef]
- Remize, F.; Planchon, F.; Lohézic-Le Dévéhat, F.; Falaise, C.; Guihéneuf, F. Bioactive compounds from marine microalgae: Potential applications and challenges. Mar. Drugs 2021, 19, 445. [Google Scholar]
- Strauch, S.M.; Coutinho, F. Bioactive pigments from microalgae and cyanobacteria: A review on their chemistry, biological activities and industrial applications. Mar. Drugs 2021, 19, 469. [Google Scholar]
- Zhou, J.; Li, Y.; Wang, H.; Chen, X.; Liu, F.; Zhang, L.; Yang, Q.; Xu, R.; Zhao, S.; Huang, D. Bioactive compounds in microalgae and their potential in functional foods. Food Chem. 2022, 370, 131118. [Google Scholar]
- Hachicha, R.; Machhout, M.; Karoud, W.; Ammar, E. Enzymatic hydrolysis of Spirulina proteins: Bioactive peptides and functional properties. Appl. Sci. 2022, 12, 9660. [Google Scholar]
- Hashem, H.; El Lahoty, S.; Abd El-Baky, H.H. Evaluation of the phytochemicals and nutritional properties of some microalgal species. Saudi J. Biol. Sci. 2021, 28, 5721–5729. [Google Scholar]
- Khemiri, I.; Bouchech, R. Effects of growth medium variation on the nutrifunctional properties of Tetraselmis sp. microalgae. J. Biotechnol. Biomater. 2021, 11. [Google Scholar]
- Maghembe, R.; Damian, D.; Makaranga, A.; Nyandoro, S.; Lyantagaye, S.; Kusari, S.; Hatti-Kaul, R. Omics for Bioprospecting and Drug Discovery from Bacteria and Microalgae. Antibiotics 2020, 9, 229. [Google Scholar] [CrossRef]
- Kuo, E.; Yang, R.; Chin, Y.Y.; Chien, Y.-L.; Chen, Y.C.; Wei, C.-Y.; Kao, L.-J.; Chang, Y.-H.; Li, Y.-J.; Chen, T.-Y.; et al. Multiomics approaches and genetic engineering of metabolism for improved biorefinery and wastewater treatment in microalgae. Biotechnol. J. 2022, 17, e2100603. [Google Scholar] [CrossRef]
- Herráiz-Gil, E.; Gil-Arriba, M.J.; Herrero, M. Multiomic data integration in food science and analysis: Current advances and future perspectives. TrAC Trends Anal. Chem. 2023, 158, 116915. [Google Scholar]
- Pereira, L.; Cotas, J.; Valado, A. Antioxidants from microalgae and their potential impact on human well-being. Explor. Drug Sci. 2024, 2, 292–321. [Google Scholar] [CrossRef]
- Raposo, M.; De Morais, A.M.M.B.; De Morais, R.M.S.C. Carotenoids from Marine Microalgae: A Valuable Natural Source for the Prevention of Chronic Diseases. Mar. Drugs 2015, 13, 5128–5155. [Google Scholar] [CrossRef]
- Vignaud, J.; Loiseau, C.; Hérault, J.; Mayer, C.; Côme, M.; Martin, I.; Ulmann, L. Microalgae Produce Antioxidant Molecules with Potential Preventive Effects on Mitochondrial Functions and Skeletal Muscular Oxidative Stress. Antioxidants 2023, 12, 1050. [Google Scholar] [CrossRef]
- Baechle, J.; Chen, N.; Makhijani, P.; Winer, S.; Furman, D.; Winer, D. Chronic inflammation and the hallmarks of aging. Mol. Metab. 2023, 74, 101755. [Google Scholar] [CrossRef]
- Cifuentes Verdejo, C.; Rodriguez-Garcia, C.; Verdejo, C. Low-grade chronic inflammation: A shared mechanism for the pathogenesis of chronic diseases. Curr. Pharm. Des. 2024, 40:1, 4–25. [Google Scholar] [CrossRef]
- Oslan, S.; Tan, J.; Oslan, S.N.; Matanjun, P.; Mokhtar, R.; Shapawi, R.; Huda, N. Haematococcus pluvialis as a Potential Source of Astaxanthin with Diverse Applications in Industrial Sectors: Current Research and Future Directions. Molecules 2021, 26, 6470. [Google Scholar] [CrossRef]
- Baky, H.; El-Baroty, G. Healthy Benefit of Microalgal Bioactive Substances. J. Aquat. Sci. 2013, 1, 11–22. [Google Scholar]
- Goff, L.; Ferrec, E.L.; Mayer, C.; Mimouni, V.; Lagadic-Gossmann, D.; Schoefs, B.; Ulmann, L. Microalgal carotenoids and phytosterols regulate biochemical mechanisms involved in human health and disease prevention. Biochimie 2019, 167, 106–118. [Google Scholar] [CrossRef]
- Dhandayuthapani, K.; Malathy, S.; Mulla, S.; Gupta, S. An Insight into the Potential Application of Microalgae in Pharmaceutical and Nutraceutical Production. In Algae; Springer: Singapore, 2020; pp. 135–179. [Google Scholar]
- Zhang, H.; Wang, M.; Zhou, Y.; Bao, S.; Wang, F.; Li, C. Protective Effects of Astaxanthin against Oxidative Stress: Attenuation of TNF-α-Induced Oxidative Damage in SW480 Cells and Azoxymethane/Dextran Sulfate Sodium-Induced Colitis-Associated Cancer in C57BL/6 Mice. Mar. Drugs 2024, 22, 469. [Google Scholar] [CrossRef]
- Tian, S.; Zhao, H.; Song, H. Shared signaling pathways and targeted therapy by natural bioactive compounds for obesity and type 2 diabetes. Crit. Rev. Food Sci. 2022, 64, 5039–5056. [Google Scholar] [CrossRef]
- Gao, L.; Qin, Y.; Zhou, X.; Jin, W.; He, Z.; Li, X.; Wang, Q. Microalgae as future food: Rich nutrients, safety, production costs and environmental effects. Sci. Total Environ. 2024, 927, 172167. [Google Scholar] [CrossRef]
- Ivanov, S.; Ostrovskaya, R. Neuroprotective substances: Are they able to protect the pancreatic beta-cells too? Endocr. Metab. Immune Disord. Drug Targets 2022, 22, 834–841. [Google Scholar]
- Paterson, S.; Majchrzak, M.; Gómez-Garre, D.; Ortega-Hernández, A.; Sánchez-González, S.; De La Fuente, M.Á.; Gómez-Cortés, P.; Hernández-Ledesma, B. Role of Simulated Nannochloropsis gaditana Digests in Shaping Gut Microbiota and Short-Chain Fatty Acid Levels. Nutrients 2024, 17, 99. [Google Scholar] [CrossRef]
- Zhou, J.-Q.; Wang, M.; Grimi, N.; Dar, B.; Calvo-Lerma, J.; Barba, F. Research progress in microalgae nutrients: Emerging extraction and purification technologies, digestive behavior, and potential effects on human gut. Crit. Rev. Food Sci. 2023, 64, 11375–11395. [Google Scholar] [CrossRef]
- Oleskin, A.V.; Boyang, C. Microalgae in Terms of Biomedical Technology: Probiotics, Prebiotics, and Metabiotics. Appl. Biochem. Microbiol. 2022, 58, 813–825. [Google Scholar] [CrossRef]
- Patel, A.K.; Singhania, R.R.; Awasthi, M.K.; Varjani, S.; Bhatia, S.K.; Tsai, M.-L.; Hsieh, S.-L.; Chen, C.-W.; Dong, C.-D. Emerging prospects of macro- and microalgae as prebiotic. Microb. Cell Factories 2021, 20, 112. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Wang, M.F.; Wang, Y.T. Sulfated polysaccharides from marine microalgae improve intestinal barrier function and reduce endotoxemia. Biomolecules 2022, 12, 1871. [Google Scholar]
- Sahoo, D.; Soren, A. Microalgal polyphenols and their impact on gut health. In Algal Biotechnology and Environmental Monitoring; Sahoo, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 219–230. [Google Scholar]
- Benameur, T.; Soares, F.A.A.; Iffland, L.; Benameur, N.; Panaro, M.A.; Filannino, F.M.; Hasan, A. Effects of microalgae on immune response and brain health: Insights into anti-inflammatory mechanisms. Brain Sci. 2023, 13, 1226. [Google Scholar] [CrossRef]
- Savvidou, M.G.; Georgiopoulou, I.; Antoniou, N.; Tzima, S.; Kontou, M.; Louli, V.; Fatouros, C.; Magoulas, K.; Kolisis, F.N. Extracts from Chlorella vulgaris Protect Mesenchymal Stromal Cells from Oxidative Stress Induced by Hydrogen Peroxide. Plants 2023, 12, 361. [Google Scholar] [CrossRef]
- Neumann, U.; Derwenskus, F.; Flaiz Flister, V.; Schmid-Staiger, U.; Hirth, T.; Bischoff, S.C. Fucoxanthin, A Carotenoid Derived from Phaeodactylum tricornutum Exerts Antiproliferative and Antioxidant Activities In Vitro. Antioxidants 2019, 8, 183. [Google Scholar] [CrossRef]
- Conde, T.; Neves, B.; Couto, D.; Melo, T.; Lopes, D.; Pais, R.; Batista, J.; Cardoso, H.; Silva, J.L.; Domingues, P.; et al. Polar Lipids of Marine Microalgae Nannochloropsis oceanica and Chlorococcum amblystomatis Mitigate the LPS-Induced Pro-Inflammatory Response in Macrophages. Mar. Drugs 2023, 21, 629. [Google Scholar] [CrossRef]
- De Andrade, A.; Hort, M.A.; Schimith, L.E.; Peraza, G.G.; Maria Lda, S.; Soares, M.C.F.; Giroldo, D.; Muccillo-Baisch, A.L. Antinociceptive and anti-inflammatory effects of cellular and extracellular extracts from microalga Chlamydomonas pumilioniformis on mice. Acta Scientiarum. Biol. Sci. 2021, 43, e52889. [Google Scholar] [CrossRef]
- Silva, M.; Kamberovic, F.; Uota, S.T.; Kovan, I.-M.; Viegas, C.S.B.; Simes, D.C.; Gangadhar, K.N.; Varela, J.; Barreira, L. Microalgae as Potential Sources of Bioactive Compounds for Functional Foods and Pharmaceuticals. Appl. Sci. 2022, 12, 5877. [Google Scholar] [CrossRef]
- Severo, I.A.; Dias, R.R.; do Nascimento, T.C.; Deprá, M.C.; Maroneze, M.M.; Zepka, L.Q.; Jacob-Lopes, E. Microalgae-derived polysaccharides: Potential building blocks for biomedical applications. World J. Microbiol. Biotechnol. 2022, 38, 150. [Google Scholar] [CrossRef]
- Tamel Selvan, K.; Goon, J.A.; Makpol, S.; Tan, J.K. Therapeutic Potentials of Microalgae and Their Bioactive Compounds on Diabetes Mellitus. Mar. Drugs 2023, 21, 462. [Google Scholar] [CrossRef]
- Nowruz, B. Investigating the effect of microalgae bioactive compounds on sleep and mood. J. Shahid Sadoughi Univ. Med. Sci. 2024, 32. [Google Scholar] [CrossRef]
- Camacho, F.; Macedo, A.; Malcata, F. Potential Industrial Applications and Commercialization of Microalgae in the Functional Food and Feed Industries: A Short Review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef]
- Niazi, S. Advice to the FDA to Improve Its Proposed Guidelines to Rationalize Clinical Trials by Restricting Placebo Control, Preventing Low-Powered Studies, and Disallowing Studies Where Bioavailability Is Not Proven. Pharmaceuticals 2024, 17, 1424. [Google Scholar] [CrossRef]
- Lee, E.; Park, J.; Choi, Y.; Huh, K.; Kim, W. A randomized study to establish the effects of spirulina in type 2 diabetes mellitus patients. Nutr. Res. Pract. 2008, 2, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Szulińska, M.; Gibas-Dorna, M.; Miller-Kasprzak, E.; Suliburska, J.; Miczke, A.; Walczak-Gałęzewska, M.; Stelmach-Mardas, M.; Walkowiak, J.; Bogdański, P. Spirulina maxima improves insulin sensitivity, lipid profile, and total antioxidant status in obese patients with well-treated hypertension: A randomized double-blind placebo-controlled study. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2473–2481. [Google Scholar]
- Yousefi, R.; Mottaghi, A.; Saidpour, A. Spirulina platensis effectively ameliorates anthropometric measurements and obesity-related metabolic disorders in obese or overweight healthy individuals: A randomized controlled trial. Complement. Ther. Med. 2018, 40, 106–112. [Google Scholar] [CrossRef]
- Wu, J.; Gu, X.; Yang, D.; Xu, S.; Wang, S.; Chen, X.; Wang, Z. Bioactive substances and potentiality of marine microalgae. Food Sci. Nutr. 2021, 9, 5279–5292. [Google Scholar] [CrossRef]
- Montero-Lobato, Z.; Vázquez, M.; Navarro, F.; Fuentes, J.; Bermejo, E.; Garbayo, I.; Vílchez, C.; Cuaresma, M. Chemically-induced production of anti-inflammatory molecules in microalgae. Mar. Drugs 2018, 16, 478. [Google Scholar] [CrossRef]
- da Silva Matos, J.P. Bioprospection, Study and Application of Bioactive Compounds From Microalgae and Seaweeds. Doctoral Thesis, Universidade de Lisboa, 2021. Available online: https://www.proquest.com/openview/f7ee1c83fe983c548b432b6ae6b77af0/1?pq-origsite=gscholar&cbl=2026366&diss=y (accessed on 28 April 2025).
- Benavente-Valdés, J.R.; Méndez-Zavala, A.; Hernández-López, I.; Carreón-González, B.A.; Velázquez-Arellano, M.E.; Morales-Oyervides, L.; Montañez-Saénz, J.C. Unconventional microalgae species and potential for their use in the food industry. In Cultured Microalgae for the Food Industry; Academic Press: Cambridge, MA, USA, 2021. [Google Scholar]
- Ferreira de Oliveira, A.P.; Bragotto, A.P.A. Microalgae-based products: Food and public health. Future Foods 2022, 6, 100157. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M. Food applications. In Microalgae: Cultivation, Recovery of Compounds and Applications; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Berde, C.V.; Berde, V.B.; Bramhachari, P.V. Bioprospection of marine microalgae for novel antioxidants in human health and medicine. In Marine Antioxidants; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Nourmohammadi, N.; Soleimanian-Zad, S.; Shekarchizadeh, H. Effect of Spirulina (Arthrospira platensis) microencapsulated in alginate and whey protein concentrate addition on physicochemical and organoleptic properties of functional stirred yogurt. J. Sci. Food Agric. 2020, 100, 5260–5268. [Google Scholar] [CrossRef]
- Soma, S.; Jayanth, C.C. Taste masking using microencapsulation in food and pharmaceutical application. World J. Adv. Res. Rev. 2024, 24, 1228–1240. [Google Scholar] [CrossRef]
- Quintero-Borregales, L.; Goyanes, S.; Famá, L. Containers for Encapsulation of Aroma/Flavour for Food Applications. In Micro- and Nano-Containers for Smart Applications; Springer: Singapore, 2022. [Google Scholar]
- Nuzzo, D.; Contardi, M.; Kossyvaki, D.; Picone, P.; Cristaldi, L.; Galizzi, G.; Bosco, G.; Scoglio, S.; Athanassiou, A.; Di Carlo, M. Heat-Resistant Aphanizomenon flos-aquae (AFA) Extract (Klamin®) as a Functional Ingredient in Food Strategy for Prevention of Oxidative Stress. Oxidative Med. Cell. Longev. 2019, 2019, 9481390. [Google Scholar] [CrossRef]
- Borowitzka, M. Microalgae in Medicine and Human Health: A Historical Perspective. In Microalgae in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2018; pp. 195–210. [Google Scholar]
- Martínez-Ruiz, F.E.; Andrade-Bustamante, G.; Holguín-Peña, R.J.; Renganathan, P.; Gaysina, L.A.; Sukhanova, N.V.; Puente, E.O.R. Microalgae as Functional Food Ingredients: Nutritional Benefits, Challenges, and Regulatory Considerations for Safe Consumption. Biomass 2025, 5, 25. [Google Scholar] [CrossRef]
- Mostafa, H.; Hashem, M.M. Microalgae as a source of carotenoids in foods, obstacles and solutions. Phytochem. Rev. 2024. [Google Scholar] [CrossRef]
- Spínola, M.P.; Costa, M.M.; Simões, R.S.; Fernandes, V.O.; Cardoso, V.; Pires, V.M.R.; Afonso, C.; Cardoso, C.; Bandarra, N.M.; Fontes, C.M.G.A.; et al. Improving protein hydrolysis and digestibility in Arthrospira platensis biomass through recombinant peptidases (EC 3.4): Opportunities for monogastric animal diets. Heliyon 2025, 11, e41460. [Google Scholar] [CrossRef]
- Spínola, M.P.; Costa, M.M.; Prates, J.A.M. Enhancing Digestibility of Chlorella vulgaris Biomass in Monogastric Diets: Strategies and Insights. Animals 2023, 13, 1017. [Google Scholar] [CrossRef]
- Spínola, M.P.; Costa, M.M.; Prates, J.A.M. Digestive Constraints of Arthrospira platensis in Poultry and Swine Feeding. Foods 2022, 11, 2984. [Google Scholar] [CrossRef]
- Sun, H.; Shufang, Y.; Weiyang, Z.; Qing, K.; Changliang, Z.; Xiaodan, F.; Fang, Z.; Zhemin, L.; Yuming, Z.; Haijin, M.; et al. Fucoxanthin from marine microalgae: A promising bioactive compound for industrial production and food application. Crit. Rev. Food Sci. 2023, 63, 7996–8012. [Google Scholar] [CrossRef]
- Alishah Aratboni, H.; Rafiei, N.; Mehdizadeh Allaf, M.; Abedini, S.; Naseema Rasheed, R.; Seif, A.; Barati, B.; Wang, S.; Morones-Ramírez, J.R. Nanotechnology: An outstanding tool for increasing and better exploitation of microalgae valuable compounds. Algal Res. 2023, 71, 103019. [Google Scholar] [CrossRef]
- Markou, G.; Economou, C.Ν.; Chentir, I. Bioprocessing of microalgae for the production of value compounds. In Handbook of Algal Technologies and Phytochemicals; CRC Press: Boca Raton, FL, USA, 2019; pp. 203–210. [Google Scholar]
- Chiriví-Salomón, J.S.; García-Huérfano, S.; Giraldo, I.A. Bioprospecting Microalgae: A Systematic Review of Current Trends. Phycology 2024, 4, 508–532. [Google Scholar] [CrossRef]
- Kaur, M.; Bhatia, S.; Gupta, U.; Decker, E.; Tak, Y.; Bali, M.; Gupta, V.; Dar, R.; Bala, S. Microalgal bioactive metabolites as promising implements in nutraceuticals and pharmaceuticals: Inspiring therapy for health benefits. Phytochem. Rev. 2023, 22, 903–933. [Google Scholar] [CrossRef]
- Berde, C.V.; Berde, V.B.; Bramhachari, P.V. Functional foods: Benefits, challenges and future perspectives. Consensus 2022. [Google Scholar]
- Palmer, J.S.; Muñoz, D.C.; Edwards, C. Biological activities of natural products from microalgae. In Cultured Microalgae for the Food Industry; Academic Press: Cambridge, MA, USA, 2021; pp. 287–324. [Google Scholar]
- Saadaoui, I.; Rasheed, R.; Aguilar, A.; Cherif, M.; Al Jabri, H.; Sayadi, S.; Manning, S.R. Microalgal-based feed: Promising alternative feedstocks for livestock and poultry production. J. Anim. Sci. Biotechnol. 2021, 12, 76. [Google Scholar]
- Afzaal, M.; Imran, M.; Iqbal, M.W.; Mansoor, S. Potential of isolated microalgae bioactives for health applications. Mar. Drugs 2025, 23, 223. [Google Scholar]
- Siddhnath Surasani, V.K.R.; Singh, A.; Singh, S.M.; Hauzoukim Murthy, L.N.; Baraiya, K.G. Bioactive compounds from micro-algae and its application in foods: A review. Discov. Food 2024, 4, 27. [Google Scholar]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for High-Value Products Towards Human Health and Nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Z. Advances in the biological fixation of carbon dioxide by microalgae. J. Chem. Technol. Biotechnol. 2021, 96, 1475–1495. [Google Scholar]
- Prates, J.A.M. Microalgae in Sustainable Monogastric Systems: Bridging Nutritional Enhancement and Environmental Sustainability. In Smart Technologies for Sustainable Livestock Systems; CRC Press: Boca Raton, FL, USA, 2025; pp. 157–172. [Google Scholar]
- Costa, M.M.; Prates, J.A.M. Sustainable livestock production and poverty alleviation. In Smart Technologies for Sustainable Development Goals: No Poverty; CRC Press: Boca Raton, FL, USA, 2024; pp. 109–124. [Google Scholar]
- Spínola, M.P.; Mendes, A.R.; Prates, J.A.M. Innovations in Limnospira platensis Fermentation: From Process Enhancements to Biotechnological Applications. Fermentation 2024, 10, 633. [Google Scholar] [CrossRef]
- Mendes, A.R.; Spínola, M.P.; Lordelo, M.; Prates, J.A.M. Advances in Bioprocess Engineering for Optimising Chlorella vulgaris Fermentation: Biotechnological Innovations and Applications. Foods 2024, 13, 4154. [Google Scholar] [CrossRef]
- McGinn, P.J.; Dickinson, K.E.; Bhatti, S.; Frigon, J.C.; Guiot, S.R.; O’Leary, S.J. Integration of microalgae cultivation with in-dustrial waste remediation for biofuel and bioenergy production: Opportunities and limitations. Photosynth. Res. 2011, 109, 231–247. [Google Scholar]
- Rathinavel, L.; Singh, S.; Rai, P.K.; Chandra, N.; Jothinathan, D.; Gaffar, I.; Pandey, A.K.; Choure, K.; Waoo, A.A.; Joo, J.C.; et al. Sustainable Microalgal Biomass for Efficient and Scalable Green Energy Solutions: Fueling Tomorrow. Fuels 2024, 5, 868–894. [Google Scholar] [CrossRef]
- Acién, F.G.; Gómez-Serrano, C.; Morales-Amaral, M.D.M.; Fernández-Sevilla, J.M.; Molina-Grima, E. Wastewater treatment using microalgae: How realistic a contribution might it be to significant urban wastewater treatment? Appl. Microbiol. Biotechnol. 2016, 100, 9013–9022. [Google Scholar]
- Prasad, R.; Gupta, S.K.; Shabnam, N.; Oliveira, C.Y.B.; Nema, A.K.; Ansari, F.A.; Bux, F. Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective. Sustainability 2021, 13, 13061. [Google Scholar] [CrossRef]
- Tan, J.S.; Lee, S.Y.; Chew, K.W.; Lam, M.K.; Lim, J.W.; Ho, S.H.; Show, P.L. A review on microalgae cultivation and harvesting, and their biomass extraction processing using ionic liquids. Bioengineered 2020, 11, 116–129. [Google Scholar]
- Dragone, G. Challenges and opportunities to increase economic feasibility and sustainability of mixotrophic cultivation of green microalgae of the genus Chlorella. Renew. Sustain. Energy Rev. 2022, 160, 112284. [Google Scholar]
- Yadav, G.; Dubey, B.K.; Sen, R. A comparative life cycle assessment of microalgae production by CO2 sequestration from flue gas in outdoor raceway ponds under batch and semi-continuous regime. J. Clean. Prod. 2020, 258, 120703. [Google Scholar] [CrossRef]
- Abu-Ghosh, S.; Fixler, D.; Dubinsky, Z.; Iluz, D. Energy-input analysis of the life-cycle of microalgal cultivation systems and best scenario for oil-rich biomass production. Appl. Energy 2015, 154, 1082–1088. [Google Scholar] [CrossRef]
- Razzak, S.A.; Bahar, K.; Islam, K.O.; Haniffa, A.K.; Faruque, M.O.; Hossain, S.Z.; Hossain, M.M. Microalgae cultivation in photobioreactors: Sustainable solutions for a greener future. Green Chem. Eng. 2024, 5, 418–439. [Google Scholar]
- Yang, S.; Wang, Y.; Wang, J.; Cheng, K.; Liu, J.; He, Y.; Zhang, Y.; Mou, H.; Sun, H. Microalgal protein for sustainable and nutritious foods: A joint analysis of environmental impacts, health benefits and consumer’s acceptance. Trends Food Sci. Technol. 2023, 143, 104278. [Google Scholar] [CrossRef]
- Mosibo, O.K.; Ferrentino, G.; Udenigwe, C.C. Microalgae Proteins as Sustainable Ingredients in Novel Foods: Recent Developments and Challenges. Foods 2024, 13, 733. [Google Scholar]
- Jadhav, H.B. Omega-3 Fatty Acids from Microalgae: Emerging Trends and Applications. Front. Nutr. 2024. [Google Scholar]
- Ahmad Wani, S.; Elshikh, M.S.; Al-Wahaibi, M.S.; Naik, H.R. (Eds.) Functional Foods: Technological Challenges and Advancement in Health Promotion, 1st ed.; CRC Press: Boca Raton, FL, USA, 2023. [Google Scholar] [CrossRef]
- Dutra Molino, J.V.; Mayfield, S.P. Harnessing genetic engineering to drive economic bioproduct production in algae. Front. Bioeng. Biotechnol. 2024, 12, 1350722. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microalgae Species | Main Bioactive Compounds | Source Type (Extract/Isolated) | Primary Physiological Actions | Evidence (Preclinical/Clinical) | References |
|---|---|---|---|---|---|
| Chlorella vulgaris | Proteins, chlorophyll, B12, carotenoids, polysaccharides | Whole biomass | General health benefits, detoxifying, gut health | ↓ LDL, ↑ HDL, antioxidant improvement (clinical) | [11,65] |
| Extract | Antioxidant, hepatoprotective | ↑ GSH, ↓ ROS (in vitro) | [38] | ||
| Polysaccharides (isolated) | Immunomodulation | Cytokine regulation (preclinical) | [65] | ||
| Peptides (isolated) | ACE inhibition, antioxidant | ↓ BP markers (in vivo) | [39] | ||
| Lipid extract | Anti-inflammatory (skin) | ↓ IL-6 expression in NHDF cells | [28] | ||
| Dunaliella salina | β-carotene (up to 10%), lutein | Whole biomass/lipid extract | Antioxidant, skin and eye health | ROS scavenging, lipid membrane protection (preclinical) | [37] |
| Haematococcus pluvialis | Astaxanthin, carotenoids, PUFAs | Astaxanthin (isolated) | Antioxidant, anti-inflammatory, photoprotection | ↓ IL-6, TNF-α, NF-κB; ↑ mitochondrial function | [16] |
| Isochrysis galbana | DHA, fucoxanthin, phytosterols, proteins | Whole biomas/lipid extract | Antioxidant, lipid-lowering, thermogenic, neuroprotective | AMPK activation, PPAR modulation, UCP1 upregulation | [22] |
| Nannochloropsis spp. | EPA, chlorophyll, carotenoids, phytosterols | Whole biomass | General lipid support | ↑ lipid profile (animal) | [33] |
| Lipid extracts | Anti-inflammatory | ↓ cytokines (in vitro) | [67] | ||
| EPA (isolated) | Cardioprotective, lipid-lowering | Improved lipids (preclinical) | [33] | ||
| Phytosterols (isolated) | Anti-inflammatory | ↓ inflammatory cytokines | [67] | ||
| Lipid extract | Antioxidant, anti-inflammatory (skin) | ↓ ROS, ↓ IL-6 (in vitro) | [67] | ||
| Phaeodactylum tricornutum | Fucoxanthin, EPA, phenolics, fibres | Extract | Antioxidant | ↑ antioxidant capacity (in vitro) | [38] |
| Fucoxanthin (isolated) | Thermogenic, hypoglycemic | ↑ AMPK, ↓ glucose and weight (in vivo) | [22] | ||
| Phenolic acids (isolated) | Antioxidant | Protective in gut and metabolic models | [38] | ||
| Scenedesmus almeriensis | Lutein, carotenoids, proteins | Whole biomass/lutein-enriched extract | Antioxidant, eye health protection, anti-inflammatory | ROS scavenging, lipid membrane protection (preclinical) | [35] |
| Schizochytrium spp. | DHA, EPA | Whole biomass | Cardiovascular health, anti-inflammatory | PPAR-α activation, lipid metabolism modulation (preclinical) | [21,33] |
| Spirulina (Limnospira platensis) | Phycocyanin, GLA, proteins, polysaccharides, B vitamins | Whole biomass | Antioxidant, metabolic regulation, gut health | ↓ HbA1c, improved lipids (clinical) | [12,21] |
| Extract | Anti-inflammatory | ↓ IL-6, ↑ SOD/CAT (in vivo) | [12,21] | ||
| Phycocyanin (isolated) | Antioxidant, anti-inflammatory | ↓ oxidative stress markers | [21] | ||
| Peptides (isolated) | ACE inhibition | ↓ BP, improved vascular function | [39] | ||
| Lipid extract | Anti-inflammatory (skin) | ↓ IL-6 in fibroblast models | [12] | ||
| Tetraselmis suecica | Lutein, phytosterols, proteins, phenolics | Whole biomass | General cardiovascular support | ↓ cholesterol (in vivo), cytokine modulation | [17] |
| Extract | Antioxidant, cholesterol-lowering | Improved lipid and antioxidant profiles | [38] | ||
| Phytosterols (isolated) | Hypocholesterolemic | ↓ LDL, ↑ bile acid excretion | [17] | ||
| Lutein/Phenolics (isolated) | Antioxidant, antimicrobial | ↑ antioxidant enzymes, ↓ pathogens | [38] |
| Microalgae Species | Applications | Market Status | References |
|---|---|---|---|
| Aphanizomenon | Supplements, functional foods (traditional health applications) | Commercial | [88] |
| Chlorella vulgaris | Supplements, functional foods, beverages, snacks (rich in proteins, chlorophyll, and fibres) | Commercial | [24,28,82] |
| Dunaliella salina | Supplements, functional foods (rich in β-carotene for antioxidant support) | Commercial | [28,35] |
| Haematococcus pluvialis | Supplements, functional foods (rich in astaxanthin for antioxidant and anti-inflammatory support) | Commercial | [28,35] |
| Isochrysis galbana | Supplements, functional foods (rich in DHA, EPA, and fucoxanthin for cardiovascular and metabolic health) | Under development | [26,90] |
| Nannochloropsis gaditana | Supplements, functional foods (rich in omega-3 PUFA, especially EPA) | Commercial | [35] |
| Nostoc | Supplements, functional foods (traditional health applications) | Under development | [89] |
| Phaeodactylum tricornutum | Supplements, functional foods (rich in fucoxanthin and EPA for anti-obesity and antioxidant effects) | Under development | [21,22] |
| Porphyridium spp. | Supplements, functional foods (rich in sulphated polysaccharides for gut health and prebiotic effects) | Under development | [62,70] |
| Scenedesmus almeriensis | Supplements, functional foods (rich in lutein for eye health and antioxidant support) | Under development | [35] |
| Schizochytrium spp. | Supplements, functional foods (rich in DHA for cognitive and cardiovascular health) | Commercial | [26,90] |
| Spirulina (Limnospira platensis) | Supplements, functional foods, protein powders, and snacks (rich in proteins, phycocyanin, and GLA) | Commercial | [24,28,82] |
| Tetraselmis suecica | Supplements, functional foods (rich in phytosterols and polyphenols for heart health) | Under development | [17] |
| Strategy | Description | Benefits | Challenges | References |
|---|---|---|---|---|
| Cell Disruption | Breaking robust cell walls using high-pressure homogenisation or ultrasonication |
| High energy consumption and scalability issues | [27,93] |
| Encapsulation Technology | Wrapping bioactives (e.g., astaxanthin) in protective layers like liposomes or nanoemulsions |
| Costly production and potential regulatory hurdles | [26,95] |
| Nanotechnology | Using nanoparticles to improve light capture and nutrient delivery in cultivation (e.g., enhancing DHA production) |
| Safety concerns regarding nanoparticle ingestion | [96] |
| Nutrient Manipulation | Adjusting nutrients (e.g., nitrogen starvation) or light to stimulate bioactive production |
| May reduce overall biomass productivity | [97,98] |
| Thermal Processing | Applying mild heat to facilitate the extraction of proteins, lipids, and pigments |
| Potential degradation of heat-sensitive bioactives | [27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guil-Guerrero, J.L.; Prates, J.A.M. Microalgae Bioactives for Functional Food Innovation and Health Promotion. Foods 2025, 14, 2122. https://doi.org/10.3390/foods14122122
Guil-Guerrero JL, Prates JAM. Microalgae Bioactives for Functional Food Innovation and Health Promotion. Foods. 2025; 14(12):2122. https://doi.org/10.3390/foods14122122
Chicago/Turabian StyleGuil-Guerrero, José L., and José A. M. Prates. 2025. "Microalgae Bioactives for Functional Food Innovation and Health Promotion" Foods 14, no. 12: 2122. https://doi.org/10.3390/foods14122122
APA StyleGuil-Guerrero, J. L., & Prates, J. A. M. (2025). Microalgae Bioactives for Functional Food Innovation and Health Promotion. Foods, 14(12), 2122. https://doi.org/10.3390/foods14122122