
The Impact of Kefir Consumption on Inflammation, Oxidative Stress Status, and Metabolic-Syndrome-Related Parameters in Animal Models: A Systematic Review and Meta-Analysis

 ,
,  , , , ,
, , , ,  , ,
, , 
Abstract

1. Introduction
2. Methods
2.1. Data Sources and Searches
2.2. Study Selection and Eligibility
2.3. Data Collection
2.4. Quality Assessment of the Included Articles
2.5. Statistical Analysis
3. Results
3.1. Study Inclusion
3.2. Characteristics of the Included Studies
3.3. Experimental Models
3.4. Interventions
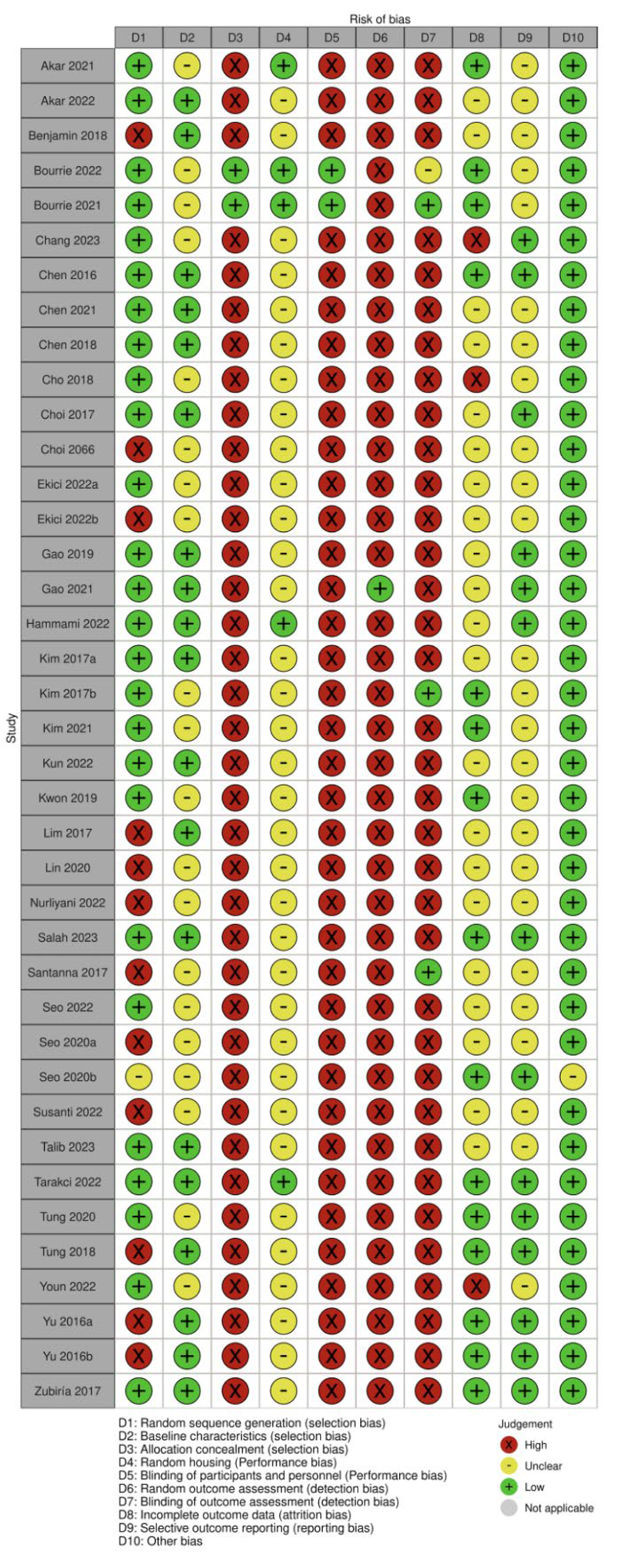
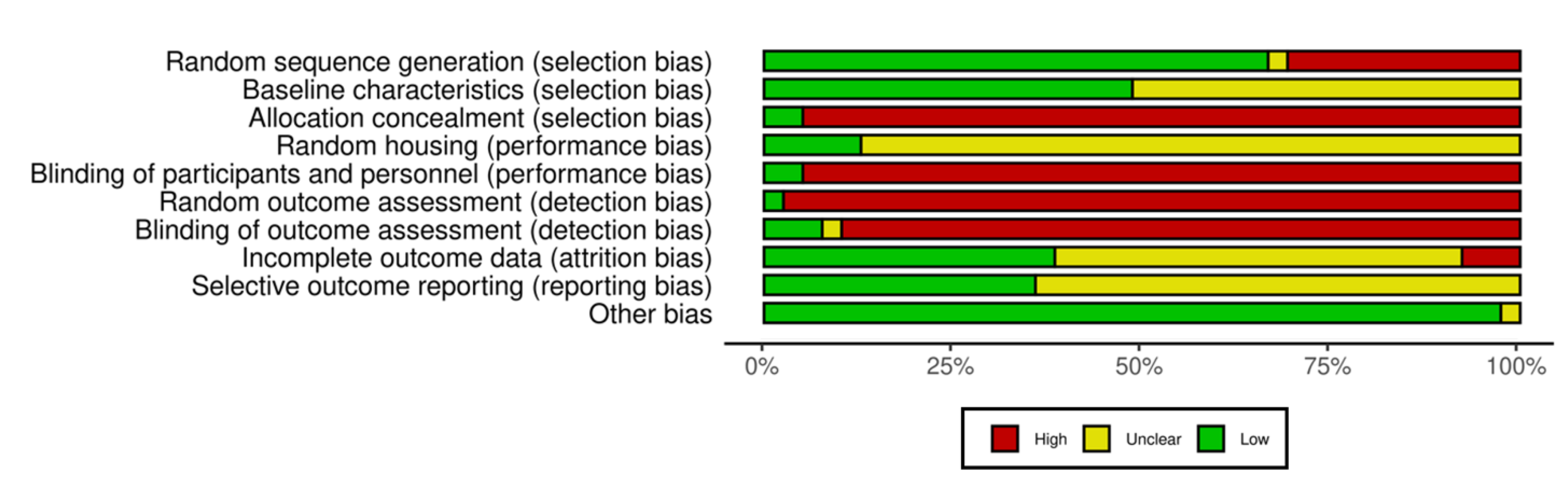
3.5. Quality Assessment
3.6. Qualitative Synthesis
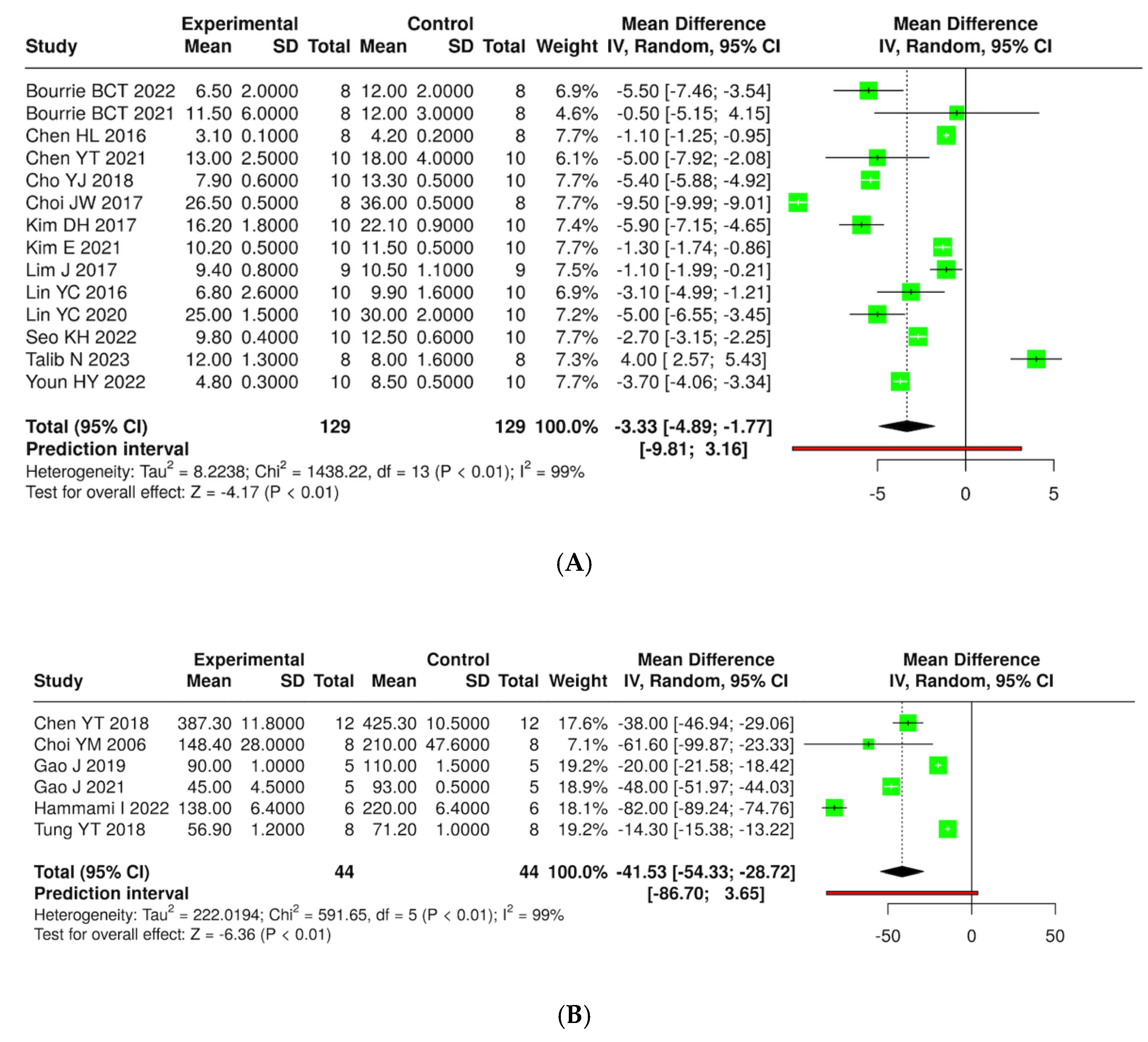
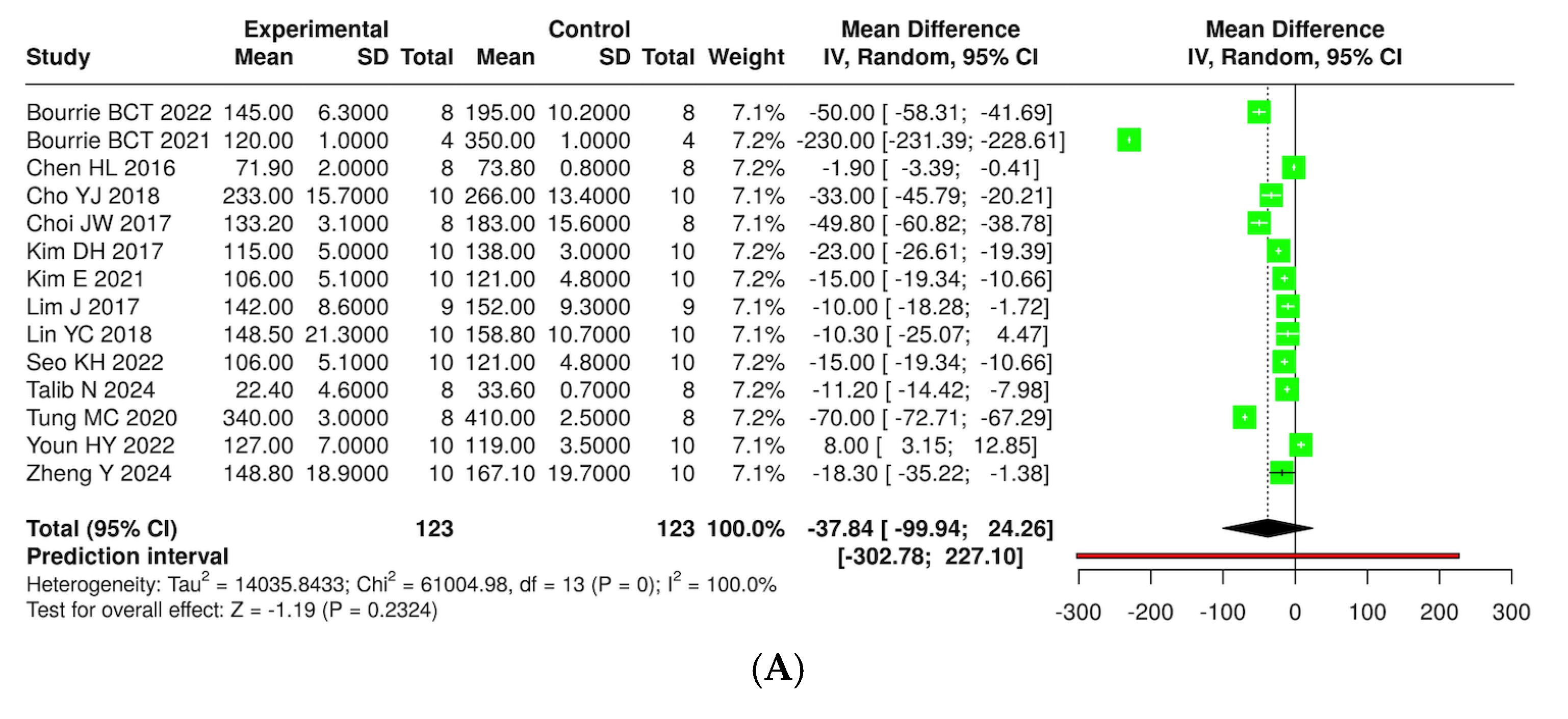
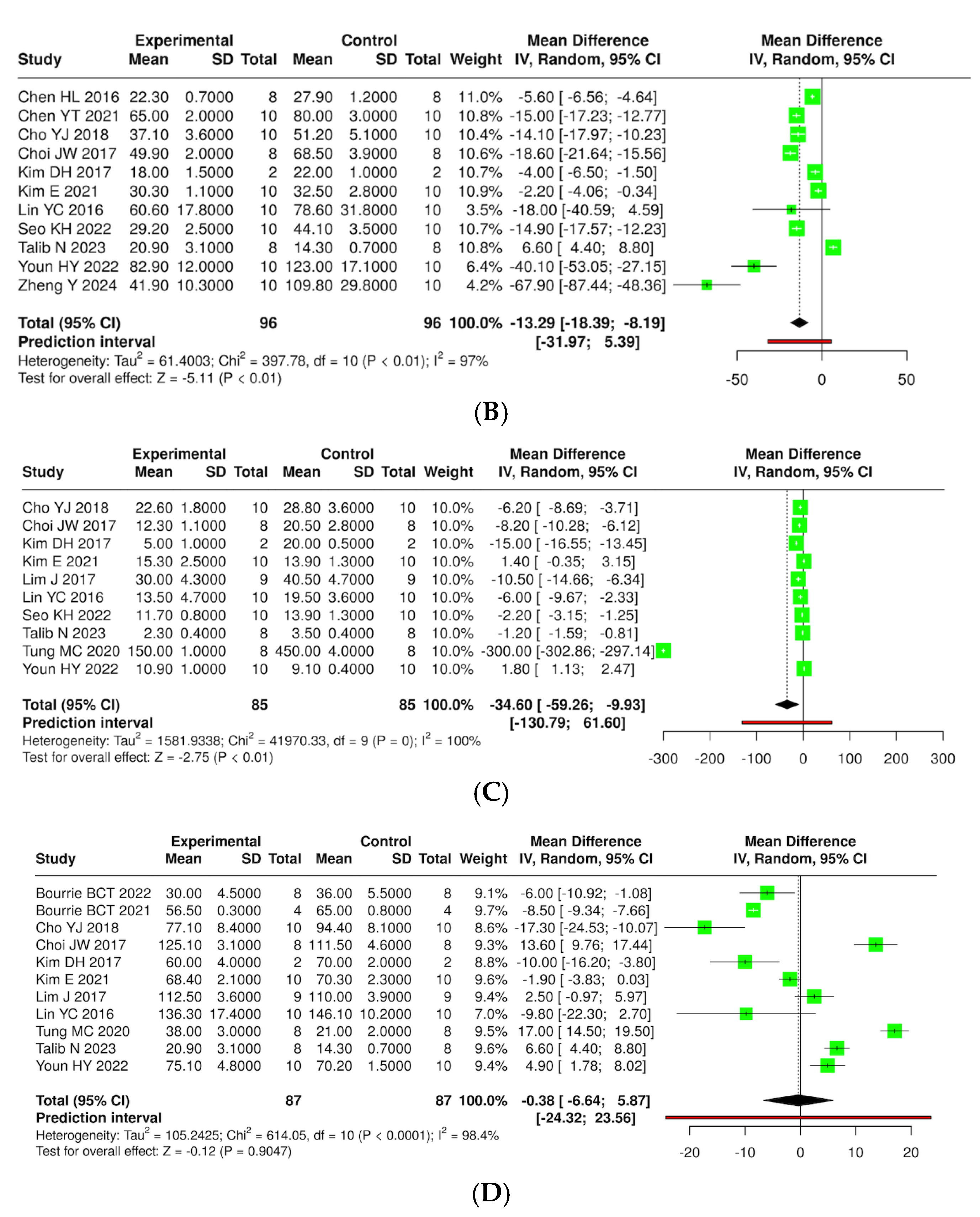
3.7. Quantitative Synthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, H.; Zhou, X.D.; Shapiro, M.D.; Lip, G.Y.H.; Tilg, H.; Valenti, L.; Somers, V.K.; Byrne, C.D.; Targher, G.; Yang, W.; et al. Global burden of metabolic diseases, 1990–2021. Metabolism 2024, 160, 155999. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ma, L.; Han, Z.; Xiong, P. The global burden of disease attributable to high body mass index in 204 countries and territories: Findings from 1990 to 2019 and predictions to 2035. Diabetes Obes. Metab. 2024, 26, 3998–4010. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Zuo, P.; Ma, G. Association of weight-adjusted waist index with cardiovascular disease and mortality among metabolic syndrome population. Sci. Rep. 2024, 14, 18684. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; He, Y.; Wang, J.; Yu, L.; Gong, Q.; Chen, Y.; An, Y.; He, S.; Li, G.; Zhang, B. Influence of impaired glucose tolerance alone and combined with metabolic syndrome on long-term risk of cardiovascular events and mortality. J. Diabetes 2024, 16, e13598. [Google Scholar] [CrossRef]
- Pammer, L.M.; Lamina, C.; Schultheiss, U.T.; Kotsis, F.; Kollerits, B.; Stockmann, H.; Lipovsek, J.; Meiselbach, H.; Busch, M.; Eckardt, K.U.; et al. Association of the metabolic syndrome with mortality and major adverse cardiac events: A large chronic kidney disease cohort. J. Intern. Med. 2021, 290, 1219–1232. [Google Scholar] [CrossRef]
- Sahoo, J.P.; Mukherjee, J.J.; Lee, K.O.; Khoo, C.M. Chapter 7—Epidemiology of metabolic syndrome in South and South-East Asia. In Metabolic Syndrome; Mukhopadhyay, S., Mondal, S., Eds.; Academic Press: Cambridge, MA, USA, 2024; pp. 73–83. [Google Scholar] [CrossRef]
- Noubiap, J.J.; Nansseu, J.R.; Lontchi-Yimagou, E.; Nkeck, J.R.; Nyaga, U.F.; Ngouo, A.T.; Tounouga, D.N.; Tianyi, F.L.; Foka, A.J.; Ndoadoumgue, A.L.; et al. Global, regional, and country estimates of metabolic syndrome burden in children and adolescents in 2020: A systematic review and modelling analysis. Lancet Child Adolesc. Health 2022, 6, 158–170. [Google Scholar] [CrossRef]
- Wu, Z.; Xia, F.; Wang, W.; Zhang, K.; Fan, M.; Lin, R. The global burden of disease attributable to high body mass index in 204 countries and territories from 1990 to 2021 with projections to 2050: An analysis of the Global Burden of Disease Study 2021. Eur. J. Heart Fail. 2025, 27, 354–365. [Google Scholar] [CrossRef]
- Monserrat-Mesquida, M.; Quetglas-Llabrés, M.; Capó, X.; Bouzas, C.; Mateos, D.; Pons, A.; Tur, J.A.; Sureda, A. Metabolic syndrome is associated with oxidative stress and proinflammatory state. Antioxidants 2020, 9, 236. [Google Scholar] [CrossRef]
- Savulescu-Fiedler, I.; Mihalcea, R.; Dragosloveanu, S.; Scheau, C.; Baz, R.O.; Caruntu, A.; Scheau, A.E.; Caruntu, C.; Benea, S.N. The Interplay between obesity and inflammation. Life 2024, 14, 856. [Google Scholar] [CrossRef]
- Li, A.; Zheng, N.; Ding, X. Mitochondrial abnormalities: A hub in metabolic syndrome-related cardiac dysfunction caused by oxidative stress. Heart Fail. Rev. 2022, 27, 1387–1394. [Google Scholar] [CrossRef]
- Mafe, A.N.; Edo, G.I.; Majeed, O.S.; Gaaz, T.S.; Akpoghelie, P.O.; Isoje, E.F.; Igbuku, U.A.; Owheruo, J.O.; Opiti, R.A.; Garba, Y.; et al. A review on probiotics and dietary bioactives: Insights on metabolic well-being, gut microbiota, and inflammatory responses. Food Chem. Adv. 2025, 6, 100919. [Google Scholar] [CrossRef]
- Peng, X.; Xian, H.; Ge, N.; Hou, L.; Tang, T.; Xie, D.; Gao, L.; Yue, J. Effect of probiotics on glycemic control and lipid profiles in patients with type 2 diabetes mellitus: A randomized, double blind, controlled trial. Front. Endocrinol. 2024, 15, 1440286. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Ma, C.; Yang, Y.; Liu, X.; Wang, B.; Wang, Y.; Zhang, G.; Bian, X.; Zhang, N. The role and mechanism of probiotics supplementation in blood glucose regulation: A Review. Foods 2024, 13, 2719. [Google Scholar] [CrossRef]
- Yang, L.; Xie, X.; Li, Y.; Wu, L.; Fan, C.; Liang, T.; Xi, Y.; Yang, S.; Li, H.; Zhang, J.; et al. Evaluation of the cholesterol-lowering mechanism of Enterococcus faecium strain 132 and Lactobacillus paracasei strain 201 in hypercholesterolemia rats. Nutrients 2021, 13, 1982. [Google Scholar] [CrossRef]
- Bernini, L.J.; Simão, A.N.C.; de Souza, C.H.B.; Alfieri, D.F.; Segura, L.G.; Costa, G.N.; Dichi, I. Effect of Bifidobacterium lactis HN019 on inflammatory markers and oxidative stress in subjects with and without the metabolic syndrome. Br. J. Nutr. 2018, 120, 645–652. [Google Scholar] [CrossRef]
- Culpepper, T. The Effects of kefir and kefir components on immune and metabolic physiology in pre-clinical studies: A narrative review. Cureus 2022, 14, e27768. [Google Scholar] [CrossRef]
- Júnior, J.; Meireles Mafaldo, Í.; de Lima Brito, I.; Tribuzy de Magalhães Cordeiro, A.M. Kombucha: Formulation, chemical composition, and therapeutic potentialities. Curr. Res. Food Sci. 2022, 5, 360–365. [Google Scholar] [CrossRef]
- Mousavi, S.N.; Saboori, S.; Asbaghi, O. Effect of daily probiotic yogurt consumption on inflammation: A systematic review and meta-analysis of randomized Controlled Clinical trials. Obes. Med. 2020, 18, 100221. [Google Scholar] [CrossRef]
- Khanturgaev, A.G.; Khamagaeva, I.S.; Shiretorova, V.G. Production of probiotic kvass beverages enriched with pine nut shell extract and propionic acid bacteria. Food Sci. Technol. 2023, 43. [Google Scholar] [CrossRef]
- Apalowo, O.E.; Adegoye, G.A.; Mbogori, T.; Kandiah, J.; Obuotor, T.M. Nutritional characteristics, health impact, and applications of kefir. Foods 2024, 13, 1026. [Google Scholar] [CrossRef]
- Bourrie, B.C.T.; Cotter, P.D.; Willing, B.P. Traditional kefir reduces weight gain and improves plasma and liver lipid profiles more successfully than a commercial equivalent in a mouse model of obesity. J. Funct. Foods 2018, 46, 29–37. [Google Scholar] [CrossRef]
- Saleem, K.; Ali, I.; Farhan, S.; Muhammad, A.; Huda, A.; Muzammal, H.; Awais, R.; Amara, R.; Aasma, A.; and Asif Shah, M. Nutritional and functional properties of kefir: Review. Int. J. Food Prop. 2023, 26, 3261–3274. [Google Scholar] [CrossRef]
- Chen, H.L.; Tsai, T.C.; Tsai, Y.C.; Liao, J.W.; Yen, C.C.; Chen, C.M. Kefir peptides prevent high-fructose corn syrup-induced non-alcoholic fatty liver disease in a murine model by modulation of inflammation and the JAK2 signaling pathway. Nutr. Diabetes 2016, 6, e237. [Google Scholar] [CrossRef]
- Kim, D.-H.; Kim, H.; Jeong, D.; Kang, I.-B.; Chon, J.-W.; Kim, H.-S.; Song, K.-Y.; Seo, K.-H. Kefir alleviates obesity and hepatic steatosis in high-fat diet-fed mice by modulation of gut microbiota and mycobiota: Targeted and untargeted community analysis with correlation of biomarkers. J. Nutr. Biochem. 2017, 44, 35–43. [Google Scholar] [CrossRef]
- Akar, F.; Sumlu, E.; Alçığır, M.E.; Bostancı, A.; Sadi, G. Potential mechanistic pathways underlying intestinal and hepatic effects of kefir in high-fructose-fed rats. Food Res. Int. 2021, 143, 110287. [Google Scholar] [CrossRef]
- Vieira, C.P.; Rosario, A.; Lelis, C.A.; Rekowsky, B.S.S.; Carvalho, A.P.A.; Rosário, D.K.A.; Elias, T.A.; Costa, M.P.; Foguel, D.; Conte-Junior, C.A. Bioactive compounds from kefir and their potential benefits on health: A systematic review and meta-analysis. Oxid. Med. Cell Longev. 2021, 2021, 9081738. [Google Scholar] [CrossRef]
- Albuquerque Pereira, M.d.F.; Matias Albuini, F.; Gouveia Peluzio, M.d.C. Anti-inflammatory pathways of kefir in murine model: A systematic review. Nutr. Rev. 2023, 82, 210–227. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Macleod, M.R.; O’Collins, T.; Howells, D.W.; Donnan, G.A. Pooling of animal experimental data reveals influence of study design and publication bias. Stroke 2004, 35, 1203–1208. [Google Scholar] [CrossRef]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef]
- Fekete, J.T.; Győrffy, B. MetaAnalysisOnline.com: Web-Based tool for the rapid meta-analysis of clinical and epidemiological studies. J. Med. Internet Res. 2025, 27, e64016. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.M.; Bae, S.H.; Kang, D.H.; Suh, H.J. Hypolipidemic effect of lactobacillus ferment as a functional food supplement. Phytother. Res. 2006, 20, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Lee, Y.; Kweon, M.; Kim, S.; Kang, I.-J. Evaluate and compare the anti-obesity effects of two probiotic preparations in high-fat diet-induced obese mice. J. Funct. Foods 2024, 116, 106199. [Google Scholar] [CrossRef]
- Cho, Y.J.; Lee, H.G.; Seo, K.H.; Yokoyama, W.; Kim, H. Antiobesity effect of prebiotic polyphenol-rich grape seed flour supplemented with probiotic kefir-derived lactic acid bacteria. J. Agric. Food Chem. 2018, 66, 12498–12511. [Google Scholar] [CrossRef]
- Choi, J.W.; Kang, H.W.; Lim, W.C.; Kim, M.K.; Lee, I.Y.; Cho, H.Y. Kefir prevented excess fat accumulation in diet-induced obese mice. Biosci. Biotechnol. Biochem. 2017, 81, 958–965. [Google Scholar] [CrossRef]
- Kim, D.H.; Jeong, D.; Kang, I.B.; Kim, H.; Song, K.Y.; Seo, K.H. Dual function of Lactobacillus kefiri DH5 in preventing high-fat-diet-induced obesity: Direct reduction of cholesterol and upregulation of PPAR-α in adipose tissue. Mol. Nutr. Food Res. 2017, 61, 1700252. [Google Scholar] [CrossRef]
- Kim, E.; Lee, H.G.; Han, S.; Seo, K.H.; Kim, H. Effect of surface layer proteins derived from paraprobiotic kefir lactic acid bacteria on inflammation and high-fat diet-induced obesity. J. Agric. Food Chem. 2021, 69, 15157–15164. [Google Scholar] [CrossRef]
- Kwon, J.H.; Lee, H.G.; Seo, K.H.; Kim, H. Combination of whole grapeseed flour and newly isolated kefir lactic acid bacteria reduces high-fat-induced hepatic steatosis. Mol. Nutr. Food Res. 2019, 63, e1801040. [Google Scholar] [CrossRef]
- Lim, J.; Kale, M.; Kim, D.H.; Kim, H.S.; Chon, J.W.; Seo, K.H.; Lee, H.G.; Yokoyama, W.; Kim, H. Antiobesity effect of exopolysaccharides isolated from kefir grains. J. Agric. Food Chem. 2017, 65, 10011–10019. [Google Scholar] [CrossRef]
- Seo, K.-H.; Gyu Lee, H.; Young Eor, J.; Jin Jeon, H.; Yokoyama, W.; Kim, H. Effects of kefir lactic acid bacteria-derived postbiotic components on high fat diet-induced gut microbiota and obesity. Food Res. Int. 2022, 157, 111445. [Google Scholar] [CrossRef]
- Youn, H.-Y.; Seo, K.-H.; Kim, H.-J.; Kim, Y.-S.; Kim, H. Effect of postbiotics derived from kefir lactic acid bacteria-mediated bioconversion of citrus pomace extract and whey on high-fat diet-induced obesity and gut dysbiosis. Food Res. Int. 2022, 162, 111930. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Hsu, A.H.; Chiou, S.Y.; Lin, Y.C.; Lin, J.S. AB-Kefir reduced body weight and ameliorated inflammation in adipose tissue of obese mice fed a high-fat diet, but not a high-sucrose diet. Nutrients 2021, 13, 2182. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Yang, N.S.; Lin, Y.C.; Ho, S.T.; Li, K.Y.; Lin, J.S.; Liu, J.R.; Chen, M.J. A combination of Lactobacillus mali APS1 and dieting improved the efficacy of obesity treatment via manipulating gut microbiome in mice. Sci. Rep. 2018, 8, 6153. [Google Scholar] [CrossRef]
- Chang, G.R.; Cheng, W.Y.; Fan, H.C.; Chen, H.L.; Lan, Y.W.; Chen, M.S.; Yen, C.C.; Chen, C.M. Kefir peptides attenuate atherosclerotic vascular calcification and osteoporosis in atherogenic diet-fed ApoE (−/−) knockout mice. Front. Cell Dev. Biol. 2023, 11, 1158812. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Chen, Y.-T.; Hsieh, H.-H.; Chen, M.-J. Effect of Lactobacillus mali APS1 and L. kefiranofaciens M1 on obesity and glucose homeostasis in diet-induced obese mice. J. Funct. Foods 2016, 23, 580–589. [Google Scholar] [CrossRef]
- Lin, Y.C.; Chen, Y.T.; Li, K.Y.; Chen, M.J. Investigating the Mechanistic Differences of obesity-inducing Lactobacillus kefiranofaciens M1 and anti-obesity Lactobacillus mali APS1 by microbolomics and metabolomics. Front. Microbiol. 2020, 11, 1454. [Google Scholar] [CrossRef]
- Tung, Y.T.; Chen, H.L.; Wu, H.S.; Ho, M.H.; Chong, K.Y.; Chen, C.M. Kefir peptides prevent hyperlipidemia and obesity in high-fat-diet-induced obese rats via lipid metabolism modulation. Mol. Nutr. Food Res. 2018, 62, 1700505. [Google Scholar] [CrossRef]
- Tung, M.-C.; Lan, Y.-W.; Li, H.-H.; Chen, H.-L.; Chen, S.-Y.; Chen, Y.-H.; Lin, C.-C.; Tu, M.-Y.; Chen, C.-M. Kefir peptides alleviate high-fat diet-induced atherosclerosis by attenuating macrophage accumulation and oxidative stress in ApoE knockout mice. Sci. Rep. 2020, 10, 8802. [Google Scholar] [CrossRef]
- Akar, F.; Yildirim, O.G.; Yucel Tenekeci, G.; Tunc, A.S.; Demirel, M.A.; Sadi, G. Dietary high-fructose reduces barrier proteins and activates mitogenic signalling in the testis of a rat model: Regulatory effects of kefir supplementation. Andrologia 2022, 54, e14342. [Google Scholar] [CrossRef]
- Ekici, O.; Aslan, E.; Guzel, H.; Korkmaz, O.A.; Sadi, G.; Gurol, A.M.; Boyaci, M.G.; Pektas, M.B. Kefir alters craniomandibular bone development in rats fed excess dose of high fructose corn syrup. J. Bone Miner. Metab. 2022, 40, 56–65. [Google Scholar] [CrossRef]
- Ekici, Ö.; Aslan, E.; Aladağ, T.; Güzel, H.; Korkmaz, Ö.A.; Bostancı, A.; Sadi, G.; Pektaş, M.B. Masseter muscle and gingival tissue inflammatory response following treatment with high-fructose corn syrup in rats: Anti-inflammatory and antioxidant effects of kefir. J. Food Biochem. 2022, 46, e13732. [Google Scholar] [CrossRef] [PubMed]
- Tarakci, N.G.; Erdem, N.Z.; Dumen, E. Probiotic Foods Are Effective on Weight Loss, Biochemical Parameters, and Intestinal Microbiota in Wistar Albino Rats with Obese Microbiota. Int. J. Clin. Pract. 2022, 2022, 4569100. [Google Scholar] [CrossRef] [PubMed]
- Bourrie, B.C.T.; Forgie, A.J.; Ju, T.; Richard, C.; Cotter, P.D.; Willing, B.P. Consumption of the cell-free or heat-treated fractions of a pitched kefir confers some but not all positive impacts of the corresponding whole kefir. Front. Microbiol. 2022, 13, 1056526. [Google Scholar] [CrossRef]
- Bourrie, B.C.T.; Ju, T.; Fouhse, J.M.; Forgie, A.J.; Sergi, C.; Cotter, P.D.; Willing, B.P. Kefir microbial composition is a deciding factor in the physiological impact of kefir in a mouse model of obesity. Br. J. Nutr. 2021, 125, 129–138. [Google Scholar] [CrossRef]
- Seo, K.H.; Jeong, J.; Kim, H. Synergistic Effects of Heat-Killed Kefir Paraprobiotics and Flavonoid-Rich Prebiotics on Western Diet-Induced Obesity. Nutrients 2020, 12, 2465. [Google Scholar] [CrossRef]
- Gao, J.; Mao, K.; Wang, X.; Mi, S.; Fu, M.; Li, X.; Xiao, J.; Simal-Gandara, J.; Sang, Y. Tibet Kefir Milk Regulated Metabolic Changes Induced by High-Fat Diet via Amino Acids, Bile Acids, and Equol Metabolism in Human-Microbiota-Associated Rats. J. Agric. Food Chem. 2021, 69, 6720–6732. [Google Scholar] [CrossRef]
- Gao, J.; Ding, G.; Li, Q.; Gong, L.; Huang, J.; Sang, Y. Tibet kefir milk decreases fat deposition by regulating the gut microbiota and gene expression of Lpl and Angptl4 in high fat diet-fed rats. Food Res. Int. 2019, 121, 278–287. [Google Scholar] [CrossRef]
- Nurliyani, N.; Harmayani, E.; Sunarti, S. Synbiotic goat milk kefir improves health status in rats fed a high-fat and high-fructose diet. Vet. World 2022, 15, 173–181. [Google Scholar] [CrossRef]
- Susanti, S.; Nurwantoro, N.; Elto, J.; Nugroho, T.; Erma Suryani, A.; Rizqiati, H. Preclinical study of goat milk kefir as an antihyperglycemic food. Funct. Food Health Dis. 2022, 12, 705. [Google Scholar] [CrossRef]
- Angelis-Pereira, M.C.; Barcelos Mde, F.; Sousa, M.S.; Pereira Jde, A. Effects of the kefir and banana pulp and skin flours on hypercholesterolemic rats. Acta Cir. Bras. 2013, 28, 481–486. [Google Scholar] [CrossRef]
- Santanna, A.F.; Filete, P.F.; Lima, E.M.; Porto, M.L.; Meyrelles, S.S.; Vasquez, E.C.; Endringer, D.C.; Lenz, D.; Abdalla, D.S.P.; Pereira, T.M.C.; et al. Chronic administration of the soluble, nonbacterial fraction of kefir attenuates lipid deposition in LDLr(−/−) mice. Nutrition 2017, 35, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Talib, N.; Mohamad, N.E.; Yeap, S.K.; Ho, C.L.; Masarudin, M.J.; Abd-Aziz, S.; Izham, M.N.M.; Kumar, M.R.; Hussin, Y.; Alitheen, N.B. Anti-diabetic effect of Lactobacillus paracasei isolated from Malaysian water kefir grains. Probiotics Antimicrob. Proteins 2024, 16, 2161–2180. [Google Scholar] [CrossRef] [PubMed]
- Salah, N.; Eissa, S.; Mansour, A.; El Magd, N.M.A.; Hasanin, A.H.; El Mahdy, M.M.; Hassan, M.K.; Matboli, M. Evaluation of the role of kefir in management of non-alcoholic steatohepatitis rat model via modulation of NASH linked mRNA-miRNA panel. Sci. Rep. 2023, 13, 236. [Google Scholar] [CrossRef] [PubMed]
- Zubiría, M.G.; Gambaro, S.E.; Rey, M.A.; Carasi, P.; Serradell, M.; Giovambattista, A. Deleterious metabolic effects of high fructose intake: The preventive effect of Lactobacillus kefiri administration. Nutrients 2017, 9, 470. [Google Scholar] [CrossRef]
- Hammami, I.; Ben Ali, R.; Nahdi, A.; Boussada, M.; Mahjoub, R.; Bibi, A.; El May, M.V. Kefir milk consumption decreases sperm alterations due to the high-fat diet in adult male rats. Andrologia 2022, 54, 1631–1642. [Google Scholar] [CrossRef]
- Jalili, M.; Nazari, M.; Magkos, F. Fermented foods in the management of obesity: Mechanisms of action and future challenges. Int. J. Mol. Sci. 2023, 24, 2665. [Google Scholar] [CrossRef]
- Tian, Y.; Li, F.; Du, L.; Peng, D.; Yang, Z.; Li, J.; Zhang, J. Fermented fruits ameliorate obesity by controlling food intake and regulating lipid metabolism in high-fat dietary mice. J. Funct. Foods 2024, 114, 106072. [Google Scholar] [CrossRef]
- Xiao, X.; Li, S.; Zhou, X.; Li, M.; Zhang, Y.; Ye, H. The anti-obesogenic effects and underpinning mechanisms of fermented plant-based foods: A review. Trends Food Sci. Technol. 2023, 136, 1–10. [Google Scholar] [CrossRef]
- Markowiak-Kopeć, P.; Śliżewska, K. The Effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef]
- Yang, M.; Wang, J.H.; Shin, J.H.; Lee, D.; Lee, S.N.; Seo, J.G.; Shin, J.H.; Nam, Y.D.; Kim, H.; Sun, X. Pharmaceutical efficacy of novel human-origin Faecalibacterium prausnitzii strains on high-fat-diet-induced obesity and associated metabolic disorders in mice. Front. Endocrinol. 2023, 14, 1220044. [Google Scholar] [CrossRef]
- Alex, S.; Lange, K.; Amolo, T.; Grinstead, J.S.; Haakonsson, A.K.; Szalowska, E.; Koppen, A.; Mudde, K.; Haenen, D.; Al-Lahham, S.; et al. Short-chain fatty acids stimulate angiopoietin-like 4 synthesis in human colon adenocarcinoma cells by activating peroxisome proliferator-activated receptor γ. Mol. Cell. Biol. 2013, 33, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Elamin Elhaseen, E.; Masclee Ad, A.; Jan, D.; Harm-Jan, P.; Jonkers Daisy, M. Short-chain fatty acids activate amp-activated protein kinase and ameliorate ethanol-induced intestinal barrier dysfunction in Caco-2 cell monolayers. J. Nutr. 2013, 143, 1872–1881. [Google Scholar] [CrossRef]
- Lee, W.H.; Kim, S.G. AMPK-Dependent metabolic regulation by PPAR Agonists. PPAR Res. 2010, 2010, 549101. [Google Scholar] [CrossRef]
- Bourrie, B.C.T.; Forgie, A.J.; Makarowski, A.; Cotter, P.D.; Richard, C.; Willing, B.P. Consumption of kefir made with traditional microorganisms resulted in greater improvements in LDL cholesterol and plasma markers of inflammation in males when compared to a commercial kefir: A randomized pilot study. Appl. Physiol. Nutr. Metab. 2023, 48, 668–677. [Google Scholar] [CrossRef]
- Arslan, B.; Yilmaz, I. The effect of kefir consumption on the lipid profile for individuals with normal and dyslipidemic properties: A randomized controlled trial. Rev. Nutr. 2022, 35, e210098. [Google Scholar] [CrossRef]
- Huang, Y.; Wu, F.; Wang, X.; Sui, Y.; Yang, L.; Wang, J. Characterization of Lactobacillus plantarum Lp27 isolated from Tibetan kefir grains: A potential probiotic bacterium with cholesterol-lowering effects. J. Dairy Sci. 2013, 96, 2816–2825. [Google Scholar] [CrossRef]
- Liu, H.; Xie, Y.; Xiong, L.; Dong, R.; Pan, C.; Teng, G.; Zhang, H. Effect and Mechanism of cholesterol-lowering by Kluyveromyces from Tibetan kefir. Adv. Mater. Res. 2011, 343–344, 1290–1298. [Google Scholar] [CrossRef]
- Salari, A.; Ghodrat, S.; Gheflati, A.; Jarahi, L.; Hashemi, M.; Afshari, A. Effect of kefir beverage consumption on glycemic control: A systematic review and meta-analysis of randomized controlled clinical trials. Complement. Ther. Clin. Pract. 2021, 44, 101443. [Google Scholar] [CrossRef]
- Hadisaputro, S.; Ks, I.; Cahyono, B.; Suzery, M.; Widiastuti, Y.; Purnawan, A. Effects of clear kefir on biomolecular aspects of glycemic status of Type 2 diabetes mellitus (T2DM) patients in bandung, West Java [Study on Human Blood Glucose, c Peptide, and Insulin]. Funct. Foods Health Dis. 2014, 4, 340–348. [Google Scholar] [CrossRef]
- Yahyapoor, F.; Haghighat, N.; Sohrabi, Z.; Asbaghi, O.; Bagherniya, M.; Jamialahmadi, T.; Sahebkar, A. Effects of kefir consumption on cardiometabolic risk factors: A systematic review and meta-analysis of randomized controlled trials. Curr. Drug Targets 2023, 24, 599–612. [Google Scholar] [CrossRef]
- Peluzio, M.; Dias, M.M.E.; Martinez, J.A.; Milagro, F.I. Kefir and intestinal microbiota modulation: Implications in human health. Front. Nutr. 2021, 8, 638740. [Google Scholar] [CrossRef]
- Teruya, K.; Yamashita, M.; Tominaga, R.; Nagira, T.; Shim, S.Y.; Katakura, Y.; Tokumaru, S.; Tokumaru, K.; Barnes, D.; Shirahata, S. Fermented milk, Kefram-kefir enhances glucose uptake into insulin-responsive muscle cells. Cytotechnology 2002, 40, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, D.; Chrysikopoulou, V.; Rampaouni, A.; Tsoupras, A. Antioxidant and anti-inflammatory properties of water kefir microbiota and its bioactive metabolites for health promoting bio-functional products and applications. AIMS Microbiol. 2024, 10, 756–811. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; Sharma, H.; Melekoglu, E.; Ozogul, F. Recent developments in dairy kefir-derived lactic acid bacteria and their health benefits. Food Biosci. 2022, 46, 101592. [Google Scholar] [CrossRef]
- Aligita, W.; Singgih, M.; Sutrisno, E.; Adnyana, I.K. Hepatoprotective study of Indonesian water kefir against CCl4-induced liver injury in rats. J. Pharm. Pharmacogn. Res. 2023, 11, 1002–1016. [Google Scholar] [CrossRef]
- Kumar, M.R.; Yeap, S.K.; Lee, H.C.; Mohamad, N.E.; Nazirul Mubin Aziz, M.; Khalid, M.; Masarudin, M.J.; Leow, A.T.C.; Abdullah, J.O.; Alitheen, N.B. Selected kefir water from Malaysia attenuates hydrogen peroxide-induced oxidative stress by upregulating endogenous antioxidant levels in SH-SY5Y neuroblastoma cells. Antioxidants 2021, 10, 940. [Google Scholar] [CrossRef]
- Pugliero, S.; Lima, D.Y.; Rodrigues, A.M.; Bogsan, C.S.B.; Rogero, M.M.; Punaro, G.R.; Higa, E.M.S. Kefir reduces nitrosative stress and upregulates Nrf2 in the kidney of diabetic rats. Int. Dairy J. 2021, 114, 104909. [Google Scholar] [CrossRef]
- Liu, T.; Sun, Z.; Yang, Z.; Qiao, X. Microbiota-derived short-chain fatty acids and modulation of host-derived peptides formation: Focused on host defense peptides. Biomed. Pharmacother. 2023, 162, 114586. [Google Scholar] [CrossRef]
- Rodrigues, K.L.; Carvalho, J.C.; Schneedorf, J.M. Anti-inflammatory properties of kefir and its polysaccharide extract. Inflammopharmacology 2005, 13, 485–492. [Google Scholar] [CrossRef]
- Aires, R.; Amorim, F.; Côco, L.; Conceição, A.; Zanardo, T.; Taufner, G.; Nogueira, B.; Vasquez, E.; Pereira, T.; Campagnaro, B.; et al. Kefir peptide (Kef-1) as an emerging approach for the treatment of oxidative stress and inflammation in 2K1C mice. Food Funct 2022, 13, 1965–1974. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author, Year [Reference] | Types of Intervention * | Dose | Control Group # | Testing Duration (Weeks) |
|---|---|---|---|---|
| 1. Akar et al., 2021 [26] | Milk-based kefir grains | 0.001 mL/g bw, daily | Water | 6 |
| 2. Akar et al., 2022 [50] | Milk-based kefir grains | 0.01 mL/g bw, daily | Water | 6 |
| 3. Angelis-Pereira et al., 2013 [61] | Fermented kefir (kefir grains with distilled water and 5% brown sugar) | 8.6 mg/g bw, daily | Water | 3 |
| 4. Bourrie et al., 2018 [22] | Commercial kefir (grain) | 100 mL, daily | Milk | 12 |
| 5. Bourrie et al., 2022 [54] | Pitched kefir (fermented kefir grains and 2% fat milk with a mixture of microbes) | 3.1 g/mouse, daily | LFD with milk | 8 |
| 6. Bourrie et al., 2021 [55] | Pitched kefir (fermented kefir grains and 2% fat milk with a mixture of microbes) | 2 mL kefir/20 g food, daily | SCD | 8 |
| 7. Chang et al., 2023 [45] | Kefir peptides powder (KEFPEP®) containing 23.1 g of peptides per 100 g powder | 0.328 mg/g bw (for low dose) 0.655 mg/g bw (for high dose) | SCD | 13 |
| 8. Chen et al., 2016 [24] | Kefir peptides powder | 0.05, 0.10, 0.15 mg/g bw | Water | 8 |
| 9. Chen et al., 2021 [43] | AB-kefir (commercial) | 109 CFU/mouse//day | Saline | 10 |
| 10. Chen et al., 2018 [44] | Lactobacillus mali APS1 | 5 × 107, 5 × 108, and 5 × 109 CFU/mouse//day (for low, medium, and high doses) | Saline | 12 |
| 11. Cho et al., 2018 [35] | Kefir-derived lactic acid bacteria | 10 mL/kg bw | Saline | 5 |
| 12. Choi et al., 2017 [36] | Commercial kefir powder | 0.1% (w/w) kefir powder in HFD 0.2% (w/w) kefir powder in HFD | SCD | 8 |
| 13. Choi et al., 2006 [33] | Lactic-F (The ferment of kefir gain) | 10% (w/w) of the ferment in HFD | SCD | 4 |
| 14. Ekici et al., 2022 [51] | Commercial kefir | 10 mL/kg bw | SCD | 8 |
| 15. Ekici et al., 2022 [52] | Commercial kefir | 10 mL/kg bw | Saline | 8 |
| 16. Gao et al., 2019 [58] | Tibet kefir milk (TKM) | 18 mL/kg bw | Water | 8 |
| 17. Gao et al., 2021 [57] | Tibet kefir milk (TKM) | 18 mL/kg bw | Water | 4 (TKM2) and 8 (TKM1) |
| 18. Hammami et al., 2022 [66] | Kefir milk | 10 mL/kg bw | Semi-skimmed cow milk | 8 (+4 days) |
| 19. Kim et al., 2017 [37] | Kefir isolates (Leuconostoc mesenteroides (DH4) and Lactobacillus kefiri (DH5 and DH7)) | 0.2 mL of 2 × 108 CFU/mouse//day | Saline | 6 |
| 20. Kim et al., 2017 [25] | Kefir milk | 0.2 mL/mouse//day | Sterilized milk | 12 |
| 21. Kim et al., 2021 [38] | Cell surface layer proteins from the kefir probiotic lactic acid bacteria | 120 mg/kg bw | Saline | 6 |
| 22. Kwon et al., 2019 [39] | Leuconostoc mesenteroides and Lactobacillus kefiri isolated from kefir fermented milk | 120 mg/kg bw | Microcrystalline cellulose | 5 |
| 23. Lim et al., 2017 [40] | Water-soluble exopolysaccharides (EPS) from the probiotic kefir and kefir-gain residue (Res) | 5% (w/w) EPS in HFD 8% (w/w) Res in HFD | Microcrystalline cellulose | 4 |
| 24. Lin et al., 2016 [46] | Lactobacillus kefiranofaciens M1 and Lactobacillus mali APS1 | 1 × 108 CFU/mouse/day | PBS | 8 |
| 25. Lin et al., 2020 [47] | Lactobacillus kefiranofaciens M1 and Lactobacillus mali APS2 | 1 × 108 CFU/mouse/day | PBS | 8 |
| 26. Nurliyani et al., 2022 [59] | Symbiotic kefir and probiotic kefir | 18 mL/kg bw/day | NM | 4 |
| 27. Salah et al., 2023 [64] | Ready-made milk kefir, commercial | 1.8 mL/rat/day | NM | 12 (early probiotic treated) 3 (late probiotic treated) |
| 28. Santanna et al., 2017 [62] | Milk-based kefir grains | 22 mL/kg bw | Soluble fraction of milk | 4 |
| 29. Seo et al., 2022 [41] | Surface layer protein (SLP) and exopolysaccharides (EPS) from the probiotic kefir | 125 mg/kg bw (SLP) 250 mg/kg bw (EPS) | Saline | 6 |
| 30. Seo et al., 2020 [56] | Heat-killed lactic acid bacteria isolated from kefir grain | 10 mL/kg bw, daily | Microcrystalline cellulose | 8 |
| 31. Susanti et al., 2022 [60] | Goat’s milk kefir | 0.52 mL/mouse/day | water | 3 |
| 32. Talib et al., 2024 [63] | Lacticaseibacillus paracasei Isolated from Malaysian water kefir grains | 1 × 106 CFU/mL/day (a low dose) 1 × 1010 CFU/mL/day (a high dose) | SCD | 4 |
| 33. Tarakci et al., 2022 [53] | Commercial kefir | 6 mL/kg bw (3 days a week) | NM | 16 |
| 34. Tung et al., 2020 [49] | Kefir peptides powder containing 23.1 g of peptides per 100 g powder | 100 mg/kg bw (for low dose) 400 mg/kg bw (for high dose) | PBS | 12 |
| 35. Tung et al., 2018 [48] | Kefir peptides powder containing 23.1 g of peptides per 100 g powder | 164 mg/kg bw (for low dose) | Milk powder | 8 |
| 36. Youn et al., 2022 [42] | Lentilactobacillus kefiri DH5 Bioconversion media (postbiotics) | 1 × 108 CFU/kg bw/day 10 mL/kg bw/day | Saline | 5 |
| 37. Zheng et al., 2024 [34] | Manufactured fermented food kefir Lactiplantibacillus plantarum TWK10 | 5 × 108 CFU/day (a low dose) 1 × 109 CFU/day (a medium dose) 5 × 109 CFU/day (a high dose) | PBS | 8 |
| 38. Zubiría et al., 2017 [65] | Lactobacillus kefiri isolated from kefir fermented milk CIDCA 8348 | 1 × 108 CFU/mouse/day | Milk | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qaisrani, Z.N.; Lin, W.P.; Lay, B.B.; Phyo, K.Y.; San, M.M.; Awaeloh, N.; Aunsorn, S.; Pattanayaiying, R.; Na Ayudthaya, S.P.; Hongkulsup, C.; et al. The Impact of Kefir Consumption on Inflammation, Oxidative Stress Status, and Metabolic-Syndrome-Related Parameters in Animal Models: A Systematic Review and Meta-Analysis. Foods 2025, 14, 2077. https://doi.org/10.3390/foods14122077
Qaisrani ZN, Lin WP, Lay BB, Phyo KY, San MM, Awaeloh N, Aunsorn S, Pattanayaiying R, Na Ayudthaya SP, Hongkulsup C, et al. The Impact of Kefir Consumption on Inflammation, Oxidative Stress Status, and Metabolic-Syndrome-Related Parameters in Animal Models: A Systematic Review and Meta-Analysis. Foods. 2025; 14(12):2077. https://doi.org/10.3390/foods14122077
Chicago/Turabian StyleQaisrani, Zahid Naeem, Wai Phyo Lin, Bo Bo Lay, Khin Yadanar Phyo, Myat Mon San, Nurulhusna Awaeloh, Sasithon Aunsorn, Rinrada Pattanayaiying, Susakul Palakawong Na Ayudthaya, Choosit Hongkulsup, and et al. 2025. "The Impact of Kefir Consumption on Inflammation, Oxidative Stress Status, and Metabolic-Syndrome-Related Parameters in Animal Models: A Systematic Review and Meta-Analysis" Foods 14, no. 12: 2077. https://doi.org/10.3390/foods14122077
APA StyleQaisrani, Z. N., Lin, W. P., Lay, B. B., Phyo, K. Y., San, M. M., Awaeloh, N., Aunsorn, S., Pattanayaiying, R., Na Ayudthaya, S. P., Hongkulsup, C., Buntin, N., & Chusri, S. (2025). The Impact of Kefir Consumption on Inflammation, Oxidative Stress Status, and Metabolic-Syndrome-Related Parameters in Animal Models: A Systematic Review and Meta-Analysis. Foods, 14(12), 2077. https://doi.org/10.3390/foods14122077