
Lactobacilli-Mediated Regulation of the Microbial–Immune Axis: A Review of Key Mechanisms, Influencing Factors, and Application Prospects



Abstract

{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Immunoregulatory Mechanisms of Lactobacilli
2.1. Immunoregulatory Mechanisms of Lactobacilli Immunogen
2.1.1. Peptidoglycan
2.1.2. Lipoteichoic Acid
2.1.3. Additional Immunomodulatory Components
2.2. Immunomodulatory Mechanisms of Lactobacilli Metabolites
2.2.1. Immunomodulatory Effects of Short-Chain Fatty Acids
2.2.2. Immunomodulatory Effects of Bacteriocins
2.2.3. Immunomodulatory Effects of Exopolysaccharides
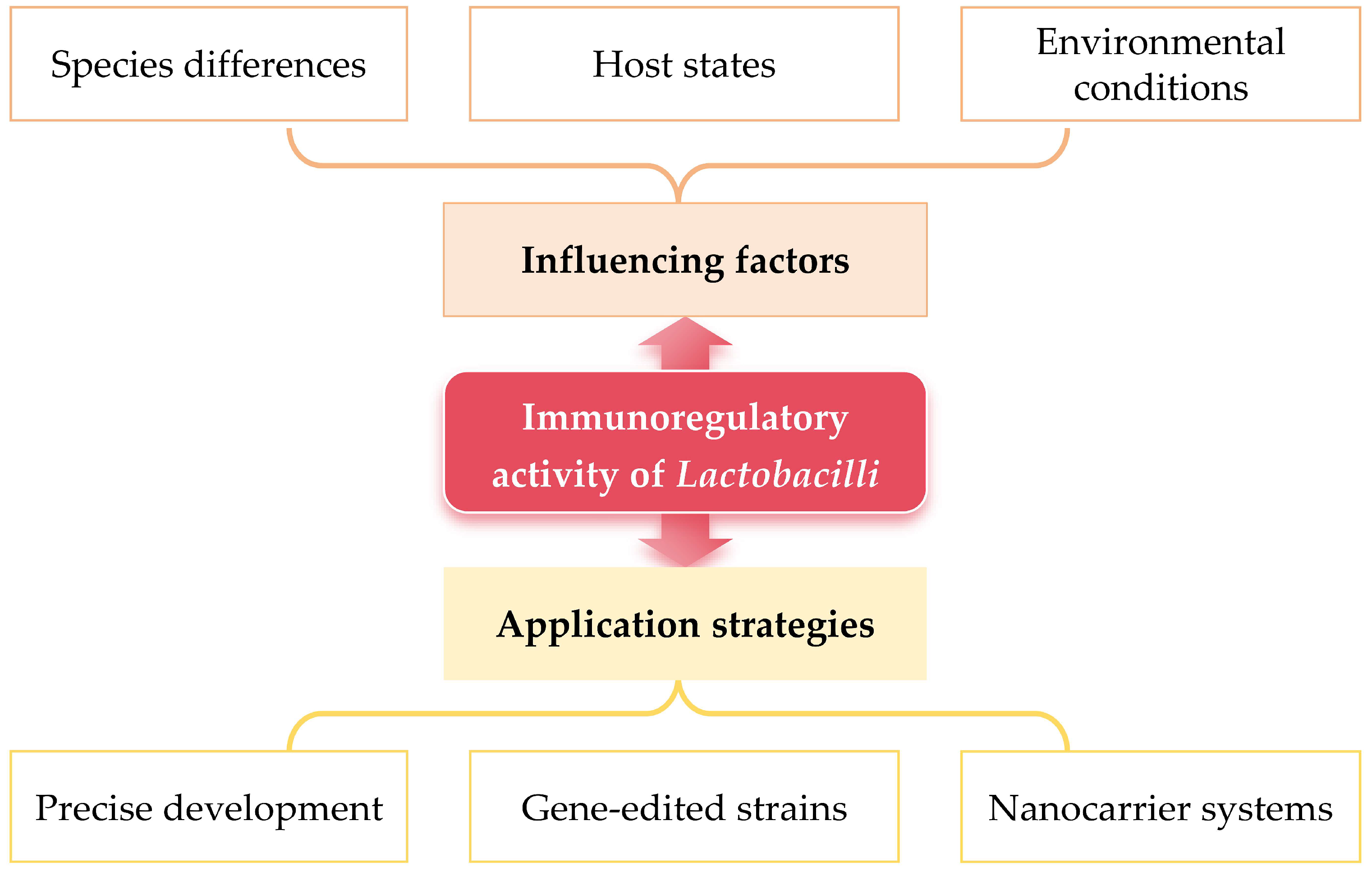
3. Factors Influencing Immunomodulatory Activity of Lactobacilli
3.1. Lactobacilli Species Differences
3.2. Host Factors
3.3. Environmental Factors
4. Application Strategies of Lactobacilli
4.1. Precision Development of Probiotics
4.2. Gene-Editing-Driven Engineering of Lactobacilli
4.3. Nanocarrier Systems
5. Summary and Prospects
5.1. Summary
5.2. Prospects in the Food Industry
5.3. Prospects in the Pharmaceutical Industry
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, Prebiotics and Immunomodulation of Gut Mucosal Defences: Homeostasis and Immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef] [PubMed]
- Mehany, T.; Khalifa, I.; Barakat, H.; Althwab, S.A.; Alharbi, Y.M.; El-Sohaimy, S. Polyphenols as promising biologically active substances for preventing SARS-CoV-2: A review with research evidence and underlying mechanisms. Food Biosci. 2021, 40, 100891. [Google Scholar] [CrossRef]
- Ying, Y.; Hao, W. Immunomodulatory function and anti-tumor mechanism of natural polysaccharides: A review. Front. Immunol. 2023, 14, 1147641. [Google Scholar] [CrossRef]
- Meng, L.; Chen, H.M.; Zhang, J.S.; Wu, Y.R.; Xu, Y.Z. Matricellular proteins: From cardiac homeostasis to immune regulation. Biomed. Pharmacother. 2024, 180, 117463. [Google Scholar] [CrossRef]
- Xiong, H.T.; Han, X.P.; Cai, L.; Zheng, H.G. Natural polysaccharides exert anti-tumor effects as dendritic cell immune enhancers. Front. Oncol. 2023, 13, 1274048. [Google Scholar] [CrossRef]
- Li, C.X.; Liu, Y.; Zhang, Y.Z.; Li, J.C.; Lai, J. Astragalus polysaccharide: A review of its immunomodulatory effect. Arch. Pharmacal Res. 2022, 45, 367–389. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Yang, L.; Liang, Q.; Sun, H.; Sun, Y.; Yan, G.; Zhang, X.; Han, Y.; Wang, X.; Wang, X. Natural resourced polysaccharides: Preparation, purification, structural elucidation, structure-activity relationships and regulating intestinal flora, a system review. Int. J. Biol. Macromol. 2024, 280, 135956. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, X.; Gao, W.; Luo, X.; Lv, J.; Wang, Y.; Liu, D. The role of intestinal flora on tumor immunotherapy: Recent progress and treatment implications. Heliyon 2024, 10, e23919. [Google Scholar] [CrossRef] [PubMed]
- Saha, U.B.; Saroj, S.D. Lactic acid bacteria: Prominent player in the fight against human pathogens. Expert Rev. Anti-Infect. Ther. 2022, 20, 1435–1453. [Google Scholar] [CrossRef]
- Feng, J.; Cen, Q.Y.; Cui, Y.R.; Hu, X.M.; Li, M.; Wang, L.J.; Wei, J.F.; Sun, N.Y.; Wang, J.Y.; Zhang, A.R. Lactobacillus rhamnosus: An emerging probiotic with therapeutic potential for depression. Pharmacol. Res. 2025, 211, 107541. [Google Scholar] [CrossRef]
- Shi, L.; Li, M.; Miyazawa, K.; Li, Y.; Hiramatsu, M.; Xu, J.Y.; Gong, C.; Jing, X.F.; He, F.; Huang, C.Y. Effects of heat-inactivated Lactobacillus gasseri TMC0356 on metabolic characteristics and immunity of rats with the metabolic syndrome. Br. J. Nutr. 2013, 109, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zheng, F.L.; Stanton, C.; Ross, R.P.; Zhao, J.X.; Zhang, H.; Chen, W. Lactobacillus reuteri FYNLJ109L1 Attenuating Metabolic Syndrome in Mice via Gut Microbiota Modulation and Alleviating Inflammation. Foods 2021, 10, 2081. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Liu, Z.J.; Liu, F.; Chen, L.J.; Wang, W.; Ma, J.G.; Xu, C.; Jiang, Z.M.; Hou, J.C. Study of the immunoregulatory effect of Lactobacillus rhamnosus 1.0320 in immunosuppressed mice. J. Funct. Foods 2021, 79, 104423. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, Y.S.; Kwok, L.Y.; Cai, T.Q.; Zhang, W.Y. The immune regulatory role of Lactobacillus acidophilus: An updated meta-analysis of randomized controlled trials. Food Biosci. 2020, 36, 100656. [Google Scholar] [CrossRef]
- Hizay, A.; Dag, K.; Oz, N.; Comak-Gocer, E.M.; Ozbey-Unlu, O.; Ucak, M.; Keles-Celik, N. Lactobacillus acidophilus regulates abnormal serotonin availability in experimental ulcerative colitis. Anaerobe 2023, 80, 102710. [Google Scholar] [CrossRef]
- Zamberlin, S.; Spehar, I.D.; Kelava, N.; Samarzija, D. Probiotic bacterium Lactobacillus rhamnosus: Beneficial and a dverse effects on human health. Milchwiss.-Milk Sci. Int. 2012, 67, 30–33. [Google Scholar]
- Sun, X.H.; Pei, Z.M.; Wang, H.C.; Zhao, J.X.; Chen, W.; Lu, W.W. Bridging dietary polysaccharides and gut microbiome: How to achieve precision modulation for gut health promotion. Microbiol. Res. 2025, 292, 128046. [Google Scholar] [CrossRef]
- Khan, R.; Shah, M.D.; Shah, L.Q.; Lee, P.C.; Khan, I. Bacterial polysaccharides-A big source for prebiotics and therapeutics. Front. Nutr. 2022, 9, 1031935. [Google Scholar] [CrossRef]
- Tsai, Y.T.; Cheng, P.C.; Pan, T.M. The immunomodulatory effects of lactic acid bacteria for improving immune functions and benefits. Appl. Microbiol. Biotechnol. 2012, 96, 853–862. [Google Scholar] [CrossRef]
- Wu, Z.; Pan, D.D.; Guo, Y.X.; Sun, Y.Y.; Zeng, X.Q. Peptidoglycan diversity and anti-inflammatory capacity in Lactobacillus strains. Carbohydr. Polym. 2015, 128, 130–137. [Google Scholar] [CrossRef]
- Liu, M.H.; Peng, R.; Tian, C.F.; Shi, J.P.; Ma, J.N.; Shi, R.W.; Qi, X.; Zhao, R.W.; Guan, H.B. Effects of the gut microbiota and its metabolite short-chain fatty acids on endometriosis. Front. Cell. Infect. Microbiol. 2024, 14, 1373004. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, S.T.; Ross, R.P.; Stanton, C. The Progress of Multi-Omics Technologies: Determining Function in Lactic Acid Bacteria Using a Systems Level Approach. Front. Microbiol. 2020, 10, 3084. [Google Scholar] [CrossRef] [PubMed]
- Kwoji, I.D.; Aiyegoro, O.A.; Okpeku, M.; Adeleke, M.A. ‘Multi-omics’ data integration: Applications in probiotics studies. Npj Sci. Food 2023, 7, 25. [Google Scholar] [CrossRef]
- Laiño, J.; Villena, J.; Kanmani, P.; Kitazawa, H. Immunoregulatory Effects Triggered by Lactic Acid Bacteria Exopolysaccharides: New Insights into Molecular Interactions with Host Cells. Microorganisms 2016, 4, 27. [Google Scholar] [CrossRef]
- Wells, J.M. Immunomodulatory mechanisms of lactobacilli. Microb. Cell Factories 2011, 10, S17. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, R.; Shah, N.P. Immune System Stimulation by Probiotic Microorganisms. Crit. Rev. Food Sci. Nutr. 2014, 54, 938–956. [Google Scholar] [CrossRef]
- Andre, G.; Kulakauskas, S.; Chapot-Chartier, M.P.; Navet, B.; Deghorain, M.; Bernard, E.; Hols, P.; Dufrêne, Y.F. Imaging the nanoscale organization of peptidoglycan in living Lactococcus lactis cells. Nat. Commun. 2010, 1, 27. [Google Scholar] [CrossRef]
- Yao, X.; Yi, Z.K.; Xu, M.; Han, Y. A Review on the Extraction, Structural Characterization, Function, and Applications of Peptidoglycan. Macromol. Rapid Commun. 2025, 46, e2400654. [Google Scholar] [CrossRef]
- Zhang, F.; Jiang, Q.; Cai, J.; Meng, F.; Tang, W.; Liu, Z.; Lin, X.; Liu, W.; Zhou, Y.; Shen, X.; et al. Activation of NOD1 on tumor-associated macrophages augments CD8+ T cell-mediated antitumor immunity in hepatocellular carcinoma. Sci. Adv. 2024, 10, eadp8266. [Google Scholar] [CrossRef]
- Stafford, C.A.; Gassauer, A.-M.; Mann, C.C.d.O.; Tanzer, M.C.; Fessler, E.; Wefers, B.; Nagl, D.; Kuut, G.; Sulek, K.; Vasilopoulou, C.; et al. Phosphorylation of muramyl peptides by NAGK is required for NOD2 activation. Nature 2022, 609, 590–596. [Google Scholar] [CrossRef]
- Caruso, R.; Warner, N.; Inohara, N.; Núñez, G. NOD1 and NOD2: Signaling, Host Defense, and Inflammatory Disease. Immunity 2014, 41, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, T.; Hovingh, E.S.; Foerster, E.G.; Abdel-Nour, M.; Philpott, D.J.; Girardin, S.E. NOD1 and NOD2 in inflammation, immunity and disease. Arch. Biochem. Biophys. 2019, 670, 69–81. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, E.A.; Drake, W.R.; Mashayekhd, S.; Ukaegbu, O.; Brown, A.R.; Grimes, C.L. Modulation of the NOD-like receptors NOD1 and NOD2: A chemist’s perspective. Bioorg. Med. Chem. Lett. 2019, 29, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.D.; Ye, J.J.; Pei, Y.M.; Wang, C.T.; Yang, H.Z.; Tian, J.Y.; Si, G.X.; Ma, Y.; Wang, K.; Liu, G. Extracellular vesicles from colorectal cancer cells promote metastasis via the NOD1 signalling pathway. J. Extracell. Vesicles 2022, 11, e12264. [Google Scholar] [CrossRef]
- Yin, R.P.; Wang, T.; Sun, J.Z.; Dai, H.Q.; Zhang, Y.T.; Liu, N.N.; Liu, H.W. Postbiotics From Lactobacillus Johnsonii Activates Gut Innate Immunity to Mitigate Alcohol-Associated Liver Disease. Adv. Sci. 2025, 12, e2405781. [Google Scholar] [CrossRef]
- Song, X.; Li, F.J.; Zhang, M.S.; Xia, Y.J.; Ai, L.Z.; Wang, G.Q. Effect of D-Ala-Ended Peptidoglycan Precursors on the Immune Regulation of Lactobacillus plantarum Strains. Front. Immunol. 2022, 12, 825825. [Google Scholar] [CrossRef]
- Kolling, Y.; Salva, S.; Villena, J.; Alvarez, S. Are the immunomodulatory properties of Lactobacillus rhamnosus CRL1505 peptidoglycan common for all Lactobacilli during respiratory infection in malnourished mice? PLoS ONE 2018, 13, e0194034. [Google Scholar] [CrossRef]
- Tian, P.J.; Li, B.L.; Shan, Y.J.; Zhang, J.N.; Chen, J.Y.; Yu, M.; Zhang, L.W. Extraction of Peptidoglycan from L. paracasei subp. Paracasei X12 and Its Preliminary Mechanisms of Inducing Immunogenic Cell Death in HT-29 Cells. Int. J. Mol. Sci. 2015, 16, 20033–20049. [Google Scholar] [CrossRef]
- Lu, Q.Q.; Guo, Y.Q.; Yang, G.; Cui, L.; Wu, Z.; Zeng, X.Q.; Pan, D.D.; Cai, Z.D. Structure and Anti-Inflammation Potential of Lipoteichoic Acids Isolated from Lactobacillus Strains. Foods 2022, 11, 1610. [Google Scholar] [CrossRef]
- Shiraishi, T.; Yokota, S.; Fukiya, S.; Yokota, A. Structural diversity and biological significance of lipoteichoic acid in Gram-positive bacteria: Focusing on beneficial probiotic lactic acid bacteria. Biosci. Microbiota Food Health 2016, 35, 147–161. [Google Scholar] [CrossRef]
- Zhou, W.; Li, X.; Zhou, H.; Hu, Y.; Chen, Y.; Guo, D. TNF-α/IL-1β/IL-1α/IL-12 inflammatory cytokine axes coupled with TLR1/TLR3/TLR5/MYD88 immune signaling pathway over-activation contribute to simultaneous carotid and coronary artery and occlusion in elderly patients. Cytokine 2025, 185, 156808. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Sim, J.R.; Yun, C.H.; Han, S.H. Lipoteichoic acids as a major virulence factor causing inflammatory responses via Toll-like receptor 2. Arch. Pharmacal Res. 2016, 39, 1519–1529. [Google Scholar] [CrossRef]
- Volz, T.; Kaesler, S.; Draing, C.; Hartung, T.; Röcken, M.; Skabytska, Y.; Biedermann, T. Induction of IL-10-balanced immune profiles following exposure to LTA from Staphylococcusepidermidis. Exp. Dermatol. 2018, 27, 318–326. [Google Scholar] [CrossRef]
- Mizuno, H.; Arce, L.; Tomotsune, K.; Albarracin, L.; Funabashi, R.; Vera, D.; Islam, A.; Vizoso-Pinto, M.G.; Takahashi, H.; Sasaki, Y.; et al. Lipoteichoic Acid Is Involved in the Ability of the Immunobiotic Strain Lactobacillus plantarum CRL1506 to Modulate the Intestinal Antiviral Innate Immunity Triggered by TLR3 Activation. Front. Immunol. 2020, 11, 571. [Google Scholar] [CrossRef]
- Bruckmaier, R.M.; Arnold, E.T.; Wellnitz, O. Pathogen-specific and dose-dependent response of the bovine mammary gland to lipopolysaccharide from E. coli and lipoteichoic acid from S. aureus. J. Dairy Sci. 2010, 93, 647–648. [Google Scholar]
- Algorri, M.; Wong-Beringer, A. Antibiotics Differentially Modulate Lipoteichoic Acid-Mediated Host Immune Response. Antibiotics 2020, 9, 573. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Im, J.; Lee, D.; Ko, K.H.; Yun, C.-H.; Han, S.H. Lipoproteins are key immunostimulatory components of Bacillus species for dendritic cell maturation and activation. Mol. Immunol. 2024, 165, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Macri, C.; Pang, E.S.; Patton, T.; O’Keeffe, M. Dendritic cell subsets. Semin. Cell Dev. Biol. 2018, 84, 11–21. [Google Scholar] [CrossRef]
- Friedrich, A.D.; Leoni, J.; Paz, M.L.; Maglio, D.G.H. Lipoteichoic Acid from Lacticaseibacillus rhamnosus GG Modulates Dendritic Cells and T Cells in the Gut. Nutrients 2022, 14, 723. [Google Scholar] [CrossRef]
- Yang, S.; Xu, X.Q.; Peng, Q.; Ma, L.; Qiao, Y.; Shi, B. Exopolysaccharides from lactic acid bacteria, as an alternative to antibiotics, on regulation of intestinal health and the immune system. Anim. Nutr. 2023, 13, 78–89. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Wu, Y.; Mehwish, H.M.; Bansal, M.; Zhao, L. Lactobacillus exopolysaccharides: New perspectives on engineering strategies, physiochemical functions, and immunomodulatory effects on host health. Trends Food Sci. Technol. 2020, 103, 36–48. [Google Scholar] [CrossRef]
- Górska, S.; Hermanova, P.; Ciekot, J.; Schwarzer, M.; Srutkova, D.; Brzozowska, E.; Kozakova, H.; Gamian, A. Chemical characterization and immunomodulatory properties of polysaccharides isolated from probiotic Lactobacillus casei LOCK 0919. Glycobiology 2016, 26, 1014–1024. [Google Scholar] [CrossRef]
- Liu, C.F.; Tseng, K.C.; Chiang, S.S.; Lee, B.H.; Hsua, W.H.; Pan, T.M. Immunomodulatory and antioxidant potential of Lactobacillus exopolysaccharides. J. Sci. Food Agric. 2011, 91, 2284–2291. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, J.; Nie, X.; Chitrakar, B.; Gao, J.; Sang, Y. Mutual adhesion of Lactobacillus spp. to intestinal cells: A review of perspectives on surface layer proteins and cell surface receptors. Int. J. Biol. Macromol. 2024, 282, 137031. [Google Scholar] [CrossRef]
- Lightfoot, Y.L.; Selle, K.; Yang, T.; Goh, Y.J.; Sahay, B.; Zadeh, M.; Owen, J.L.; Colliou, N.; Li, E.; Johannssen, T.; et al. SIGNR3-dependent immune regulation by Lactobacillus acidophilus surface layer protein A in colitis. Embo J. 2015, 34, 881–895. [Google Scholar] [CrossRef]
- Zhang, D.D.; Wu, M.T.; Guo, Y.X.; Xun, M.Y.; Wang, W.W.; Wu, Z.; Pan, D.D. Purification of Lactobacillus acidophilus surface-layer protein and its immunomodulatory effects on RAW264.7 cells. J. Sci. Food Agric. 2017, 97, 4204–4209. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Dong, P.P.; Xin, X.G.; Meng, Y.; Ding, C.H.; Xi, J. Antibacterial capacity and mechanism of the metabolites from two vaginal Lactobacillus strains. Food Biosci. 2025, 63, 105838. [Google Scholar] [CrossRef]
- Garbacz, K. Anticancer activity of lactic acid bacteria. Semin. Cancer Biol. 2022, 86, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Kim, Y.; Park, S.; Lee, D.; Lee, J.; Hlaing, S.P.; Yoo, J.W.; Rhee, S.H.; Im, E. Lactobacillus plantarum Metabolites Elicit Anticancer Effects by Inhibiting Autophagy-Related Responses. Molecules 2023, 28, 1890. [Google Scholar] [CrossRef]
- Hotchkiss, A.T.; Renye, J.A.; White, A.K.; Nunez, A.; Guron, G.K.P.; Chau, H.; Simon, S.; Poveda, C.; Walton, G.; Rastall, R.; et al. Cranberry Arabino-Xyloglucan and Pectic Oligosaccharides Induce Lactobacillus Growth and Short-Chain Fatty Acid Production. Microorganisms 2022, 10, 1346. [Google Scholar] [CrossRef]
- Hadinia, N.; Dovom, M.R.E.; Yavarmanesh, M. The effect of fermentation conditions (temperature, salt concentration, and pH) with lactobacillus strains for producing Short Chain Fatty Acids. Lwt-Food Sci. Technol. 2022, 165, 113709. [Google Scholar] [CrossRef]
- Ratajczak, W.; Ryl, A.; Mizerski, A.; Walczakiewicz, K.; Sipak, O.; Laszczynska, M. Immunomodulatory potential of gut microbiome-derived short-chain fatty acids (SCFAs). Acta Biochim. Pol. 2019, 66, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Keshelava, A.; Solis, G.P.; Hersch, M.; Koval, A.; Kryuchkov, M.; Bergmann, S.; Katanaev, V.L. High capacity in G protein-coupled receptor signaling. Nat. Commun. 2018, 9, 876. [Google Scholar] [CrossRef]
- Kaya, B.; Melhem, H.; Niess, J.H. GPR35 in Intestinal Diseases: From Risk Gene to Function. Front. Immunol. 2021, 12, 717392. [Google Scholar] [CrossRef]
- Priyadarshini, M.; Kotlo, K.U.; Dudeja, P.K.; Layden, B.T. Role of Short Chain Fatty Acid Receptors in Intestinal Physiology and Pathophysiology. Compr. Physiol. 2018, 8, 1091–1115. [Google Scholar] [CrossRef]
- Yang, N.; Ma, T.; Xie, Y.; Li, Q.; Li, Y.; Zheng, L.; Li, Y.; Xiao, Q.; Sun, Z.; Zuo, K.; et al. Lactiplantibacillus plantarum P9 for chronic diarrhea in young adults: A large double-blind, randomized, placebo-controlled trial. Nat. Commun. 2024, 15, 6823. [Google Scholar] [CrossRef]
- Natarajan, N.; Pluznick, J.L. From microbe to man: The role of microbial short chain fatty acid metabolites in host cell biology. Am. J. Physiol.-Cell Physiol. 2014, 307, C979–C985. [Google Scholar] [CrossRef]
- Gu, M.; Yin, W.; Zhang, J.; Yin, J.; Tang, X.; Ling, J.; Tang, Z.; Yin, W.; Wang, X.; Ni, Q.; et al. Role of gut microbiota and bacterial metabolites in mucins of colorectal cancer. Front. Cell. Infect. Microbiol. 2023, 13, 1119992. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, L.; Wang, N.; Li, Q.; Zhang, L.; Han, N.; Yan, T.; Ren, D.; Zhang, B.; Zhao, Y.; et al. Gut Bacterial Indole-3-acetic Acid Induced Immune Promotion Mediates Preventive Effects of Fu Brick Tea Polyphenols on Experimental Colitis. J. Agric. Food Chem. 2023, 71, 1201–1213. [Google Scholar] [CrossRef]
- Niu, J.; Cui, M.; Yang, X.; Li, J.; Yao, Y.; Guo, Q.; Lu, A.; Qi, X.; Zhou, D.; Zhang, C.; et al. Microbiota-derived acetate enhances host antiviral response via NLRP3. Nat. Commun. 2023, 14, 642. [Google Scholar] [CrossRef]
- Lorefice, L.; Zoledziewska, M. Propionic Acid Impact on Multiple Sclerosis: Evidence and Challenges. Nutrients 2024, 16, 3887. [Google Scholar] [CrossRef] [PubMed]
- Gerunova, L.K.; Gerunov, T.V.; P’Yanova, L.G.; Lavrenov, A.V.; Sedanova, A.V.; Delyagina, M.S.; Fedorov, Y.N.; Kornienko, N.V.; Kryuchek, Y.O.; Tarasenko, A.A. Butyric acid and prospects for creation of new medicines based on its derivatives: A literature review. J. Vet. Sci. 2024, 25, e23. [Google Scholar] [CrossRef] [PubMed]
- Banasiewicz, T.; Domagalska, D.; Borycka-Kiciak, K.; Rydzewska, G. Determination of butyric acid dosage based on clinical and experimental studies—A literature review. Prz. Gastroenterol. 2020, 15, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Seo, S.U.; Kweon, M.N. Gut microbiota-derived metabolites tune host homeostasis fate. Semin. Immunopathol. 2024, 46, 2. [Google Scholar] [CrossRef]
- Hao, F.; Tian, M.; Zhang, X.; Jin, X.; Jiang, Y.; Sun, X.; Wang, Y.; Peng, P.; Liu, J.; Xia, C.; et al. Butyrate enhances CPT1A activity to promote fatty acid oxidation and iTreg differentiation. Proc. Natl. Acad. Sci. USA 2021, 118, e2014681118. [Google Scholar] [CrossRef]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR-S6K pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef]
- Deliorman Orhan, D. Chapter 18—Bacteriocins Produced by Probiotic Microorganisms. In Advances in Probiotics; Dhanasekaran, D., Sankaranarayanan, A., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 277–291. [Google Scholar]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M.T. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Moein, M.; Imani Fooladi, A.A.; Mahmoodzadeh Hosseini, H. Determining the effects of green chemistry synthesized Ag-nisin nanoparticle on macrophage cells. Microb. Pathog. 2018, 114, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Guryanova, S.V. Immunomodulation, Bioavailability and Safety of Bacteriocins. Life 2023, 13, 1521. [Google Scholar] [CrossRef]
- Ahmadi, S.; Ghollasi, M.; Hosseini, H.M. The apoptotic impact of nisin as a potent bacteriocin on the colon cancer cells. Microb. Pathog. 2017, 111, 193–197. [Google Scholar] [CrossRef]
- Ismael, M.; Qayyum, N.; Gu, Y.; Zhezhe, Y.; Cui, Y.; Zhang, Y.; Lü, X. Protective effect of plantaricin bio-LP1 bacteriocin on multidrug-resistance Escherichia Coli infection by alleviate the inflammation and modulate of gut-microbiota in BALB/c mice model. Int. J. Biol. Macromol. 2023, 246, 125700. [Google Scholar] [CrossRef] [PubMed]
- Umair, M.; Jabbar, S.; Zhaoxin, L.; Jianhao, Z.; Abid, M.; Khan, K.-U.R.; Korma, S.A.; Alghamdi, M.A.; El-Saadony, M.T.; El-Hack, M.E.A.; et al. Probiotic-Based Bacteriocin: Immunity Supplementation Against Viruses. An Updated Review. Front. Microbiol. 2022, 13, 876058. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, R.; Xiao, Y.; Wang, H.; Chen, W.; Lu, W. Improvement effects of Lactobacillus-derived mannose-containing exopolysaccharides on ulcerative colitis. Food Biosci. 2024, 61, 104585. [Google Scholar] [CrossRef]
- Gotoh, Y.; Suzuki, S.; Amako, M.; Kitamura, S.; Toda, T. Effect of orally administered exopolysaccharides produced by Lactococcus lactis subsp. cremoris FC on a mouse model of dermatitis induced by repeated exposure to 2,4,6-trinitro-1-chlorobenzene. J. Funct. Foods 2017, 35, 43–50. [Google Scholar] [CrossRef]
- Surayot, U.; Wang, J.; Seesuriyachan, P.; Kuntiya, A.; Tabarsa, M.; Lee, Y.; Kim, J.-K.; Park, W.; You, S. Exopolysaccharides from lactic acid bacteria: Structural analysis, molecular weight effect on immunomodulation. Int. J. Biol. Macromol. 2014, 68, 233–240. [Google Scholar] [CrossRef]
- Ranji, P.; Agah, S.; Heydari, Z.; Rahmati-Yamchi, M.; Alizadeh, A.M. Effects of Lactobacillus acidophilus and Bifidobacterium bifidum probiotics on the serum biochemical parameters, and the vitamin D and leptin receptor genes on mice colon cancer. Iran. J. Basic Med. Sci. 2019, 22, 631–636. [Google Scholar] [CrossRef]
- Khavari-Daneshvar, H.; Mosavi, M.; Khodayari, H.; Rahimi, E.; Ranji, P.; Mohseni, A.H.; Mahmudian, R.; Shidfar, F.; Agah, S.; Alizadeh, A.M. Modifications of mice gut microflora following oral consumption of Lactobacillus acidophilus and Bifidobacterium bifidum probiotics. Turk. J. Med. Sci. 2017, 47, 689–694. [Google Scholar] [CrossRef]
- Tada, I.; Tanizawa, Y.; Endo, A.; Tohno, M.; Arita, M. Revealing the genomic differences between two subgroups in Lactobacillus gasseri. Biosci. Microbiota Food Health 2017, 36, 155–159. [Google Scholar] [CrossRef]
- Douillard, F.P.; Ribbera, A.; Kant, R.; Pietilä, T.E.; Järvinen, H.M.; Messing, M.; Randazzo, C.L.; Paulin, L.; Laine, P.; Ritari, J.; et al. Comparative Genomic and Functional Analysis of 100 Lactobacillus rhamnosus Strains and Their Comparison with Strain GG. PLoS Genet. 2013, 9, e1003683. [Google Scholar] [CrossRef]
- Zhang, Q.X.; Zhang, L.L.; Ross, P.; Zhao, J.X.; Zhang, H.; Chen, W. Comparative Genomics of Lactobacillus crispatus from the Gut and Vagina Reveals Genetic Diversity and Lifestyle Adaptation. Genes 2020, 11, 360. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.X.B.; Zheng, J.S.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, S.; Lucquin, I.; Najjari, A.; Zagorec, M.; Champomier-Vergès, M.C. Population Genetics of Lactobacillus sakei Reveals Three Lineages with Distinct Evolutionary Histories. PLoS ONE 2013, 8, e73253. [Google Scholar] [CrossRef] [PubMed]
- Moossavi, S.; Miliku, K.; Sepehri, S.; Pour, E.K.F.; Azad, M.B. The Prebiotic and Probiotic Properties of Human Milk: Implications for Infant Immune Development and Pediatric Asthma. Front. Pediatr. 2018, 6, 197. [Google Scholar] [CrossRef] [PubMed]
- Mikelsaar, M.; Stsepetova, J.; Hütt, P.; Kolk, H.; Sepp, E.; Lõivukene, K.; Zilmer, K.; Zilmer, M. Intestinal Lactobacillus sp. is associated with some cellular and metabolic characteristics of blood in elderly people. Anaerobe 2010, 16, 240–246. [Google Scholar] [CrossRef]
- Zeyneb, H.; Song, Y.; Wang, L.; Zheng, J.; Wang, W.; Pei, H.; Cao, X. Preventive effect of quinoa polysaccharides on lipopolysaccharide-induced inflammation in mice through gut microbiota regulation. Int. J. Biol. Macromol. 2025, 307, 141899. [Google Scholar] [CrossRef]
- Yinadsawaphan, T.; Kulthamrongsri, N.; Malone, R.M.; Surapongpairat, S.; Puchongmart, C.; Thiravetyan, B.; Imwattana, K.; Witayanan, J.N. A Rare Case of Native Mitral Valve Infective Endocarditis by Lactobacillus jensenii in a Healthy Young Patient. IJID Reg. 2025, 15, 100648. [Google Scholar] [CrossRef]
- Triggiano, F.; Diella, G.; Lorusso, M.; Veneziani, P.; D’Ambrosio, M.; Nesta, D.; Santacroce, L. Assessing the Relationship between Lactobacilli and HPV: A Decade of Research. Biocell 2025, 49, 199–220. [Google Scholar] [CrossRef]
- Tomasello, G.; Mazzola, M.; Leone, A.; Sinagra, E.; Zummo, G.; Farina, F.; Damiani, P.; Cappello, F.; Geagea, A.G.; Jurjus, A.; et al. Nutrition, oxidative stress and intestinal dysbiosis: Influence of diet on gut microbiota in inflammatory bowel diseases. Biomed. Pap.-Olomouc 2016, 160, 461–466. [Google Scholar] [CrossRef]
- Yang, Z.; Wen, A.; Qin, L.; Zhu, Y. Effect of Coix Seed Extracts on Growth and Metabolism of Limosilactobacillus reuteri. Foods 2022, 11, 187. [Google Scholar] [CrossRef]
- Batistela, M.E.; Bergamini, C.V.; Ale, E.C.; Sánchez, R.; Wolf, I.V.; Peralta, G.H. Culture medium based on whey permeate for biomass production of lactobacilli with technological aptitudes. Food Biosci. 2024, 62, 105445. [Google Scholar] [CrossRef]
- Shen, Y.; Miao, C.; Ma, M.; Zhen, Z.Z.; He, J.; Pei, X.Y.; Zhang, Y.; Man, C.X.; Zhao, Q.Y.; Jiang, Y.J. Mechanistic insights into the changes of biological activity and physicochemical characteristics in Lacticaseibacillus paracasei fortified milk powder during storage. Food Chem. 2024, 452, 139501. [Google Scholar] [CrossRef] [PubMed]
- Shahrajabian, M.H.; Sun, W.L. Survey on Multi-omics, and Multi-omics Data Analysis, Integration and Application. Curr. Pharm. Anal. 2023, 19, 267–281. [Google Scholar] [CrossRef]
- Zivkovic, M.; Hidalgo-Cantabrana, C.; Kojic, M.; Gueimonde, M.; Golic, N.; Ruas-Madiedo, P. Capability of exopolysaccharide-producing Lactobacillus paraplantarum BGCG11 and its non-producing isogenic strain NB1, to counteract the effect of enteropathogens upon the epithelial cell line HT29-MTX. Food Res. Int. 2015, 74, 199–207. [Google Scholar] [CrossRef]
- You, J.L.; Dong, H.L.; Mann, E.R.; Knight, S.C.; Yaqoob, P. Probiotic modulation of dendritic cell function is influenced by ageing. Immunobiology 2014, 219, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Harris, H.M.B.; McCann, A.; Guo, C.; Argimón, S.; Zhang, W.; Yang, X.; Jeffery, I.B.; Cooney, J.C.; Kagawa, T.F.; et al. Expanding the biotechnology potential of lactobacilli through comparative genomics of 213 strains and associated genera. Nat. Commun. 2015, 6, 8322. [Google Scholar] [CrossRef]
- Sola-Oladokun, B.; Culligan, E.P.; Sleator, R.D. Engineered Probiotics: Applications and Biological Containment. Annu. Rev. Food Sci. Technol. 2017, 8, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, N.; Breedon, A.M.E.; Davis, C.M.; Hwang, I.Y.; Chang, M.W. Engineering probiotics for therapeutic applications: Recent examples and translational outlook. Curr. Opin. Biotechnol. 2020, 65, 171–179. [Google Scholar] [CrossRef]
- Jiang, X.X.; Pan, D.D.; Tao, M.X.; Zhang, T.; Zeng, X.Q.; Wu, Z.; Guo, Y.X. New Nanocarrier System for Liposomes Coated with Lactobacillus acidophilus S-Layer Protein to Improve Leu-Gln-Pro-Glu Absorption through the Intestinal Epithelium. J. Agric. Food Chem. 2021, 69, 7593–7602. [Google Scholar] [CrossRef]
- Jovaini, K.; Mortazavian Farsani, S.A.M.; Aghaee-Bakhtiari, S.H.; Baniyaghoob, S. Miracle drink supplemented with Lactobacillus bulgaricus loaded-chitosan/alginate nanoparticles as a medicinal food for control of MCF7 cancer cells. Electron. J. Biotechnol. 2024, 70, 1–13. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Smith, B. Legume proteins are smart carriers to encapsulate hydrophilic and hydrophobic bioactive compounds and probiotic bacteria: A review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1250–1279. [Google Scholar] [CrossRef]
- Wang, C.; Hu, X.; Tang, H.; Ge, W.; Di, L.; Zou, J.; Cui, Z.; Zhou, A. Multiple effects of dietary supplementation with Lactobacillus reuteri and Bacillus subtilis on the growth, immunity, and metabolism of largemouth bass (Micropterus salmoides). Dev. Comp. Immunol. 2024, 160, 105241. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Cho, Y.; Kim, T.S. Lactobacillus plantarum isolated from kimchi, a Korean fermented food, attenuates imiquimod-induced psoriasis in mice. Food Biosci. 2025, 64, 105836. [Google Scholar] [CrossRef]
- Divyashree, S.; Ramu, R.; Sreenivasa, M.Y. Evaluation of new candidate probiotic lactobacillus strains isolated from a traditional fermented food—multigrain-millet dosa batter. Food Biosci. 2024, 57, 103450. [Google Scholar] [CrossRef]
- de Jesus, L.C.L.; Santos, R.C.V.; Quaresma, L.S.; Américo, M.F.; Rodrigues, T.V.; Freitas, A.d.S.; Campos, G.M.; Dutra, J.d.C.F.; Barroso, F.A.L.; Laguna, J.G.; et al. Health-promoting effects and safety aspects of Lactobacillus delbrueckii: A food industry species. Trends Food Sci. Technol. 2024, 150, 104605. [Google Scholar] [CrossRef]
- Zhan, Y.; Li, Q.; Li, T.; Yu, Q.; Li, Y.; Cheng, X.; Sun, Y.; Wang, X.; Guo, F.; Zhang, H.; et al. Lactobacillus plantarum P9 ameliorates dysfunctional immune and metabolic homeostasis along the gut–liver axis in a nonalcoholic fatty liver disease with low-grade colitis mouse model. Food Biosci. 2024, 61, 104613. [Google Scholar] [CrossRef]
- Xin, Y.; Hu, C.; Li, Y.; Yang, Z.; Zhang, L.; Li, A.; Li, C.; Liu, L.; Du, P. Immunomodulatory potential of Lactobacillus helveticus KLDS 1.8701 postbiotics: By regulating the Th17/Treg balance. Food Biosci. 2024, 61, 104842. [Google Scholar] [CrossRef]
- Seguella, L.; Corpetti, C.; Lu, J.; Pesce, M.; Franzin, S.B.; Palenca, I.; Zilli, A.; Vincenzi, M.; Caprioli, D.; Paytuví-Gallart, A.; et al. Oleoylethanolamide-producing Lactobacillus paracasei F19 improves metabolic and behavioral disorders by restoring intestinal permeability and microbiota-gut-brain axis in high-fat diet-induced obese male mice. Brain Behav. Immun. 2025, 127, 25–44. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, H.-F.; Li, M.; Han, X.; Fan, Y.-T.; Yang, J.-J.; Long, Y.; Yu, J.; Ji, H.-Y. Lactobacilli-Mediated Regulation of the Microbial–Immune Axis: A Review of Key Mechanisms, Influencing Factors, and Application Prospects. Foods 2025, 14, 1763. https://doi.org/10.3390/foods14101763
Ji H-F, Li M, Han X, Fan Y-T, Yang J-J, Long Y, Yu J, Ji H-Y. Lactobacilli-Mediated Regulation of the Microbial–Immune Axis: A Review of Key Mechanisms, Influencing Factors, and Application Prospects. Foods. 2025; 14(10):1763. https://doi.org/10.3390/foods14101763
Chicago/Turabian StyleJi, Hong-Fei, Mei Li, Xiao Han, Yu-Ting Fan, Jia-Jing Yang, Yan Long, Juan Yu, and Hai-Yu Ji. 2025. "Lactobacilli-Mediated Regulation of the Microbial–Immune Axis: A Review of Key Mechanisms, Influencing Factors, and Application Prospects" Foods 14, no. 10: 1763. https://doi.org/10.3390/foods14101763
APA StyleJi, H.-F., Li, M., Han, X., Fan, Y.-T., Yang, J.-J., Long, Y., Yu, J., & Ji, H.-Y. (2025). Lactobacilli-Mediated Regulation of the Microbial–Immune Axis: A Review of Key Mechanisms, Influencing Factors, and Application Prospects. Foods, 14(10), 1763. https://doi.org/10.3390/foods14101763