
Identification of Characteristic Flavor Compounds and Quality Evaluation of Butyriboletus roseoflavus from Different Regions in Yunnan

 ,
, 
Abstract
1. Introduction
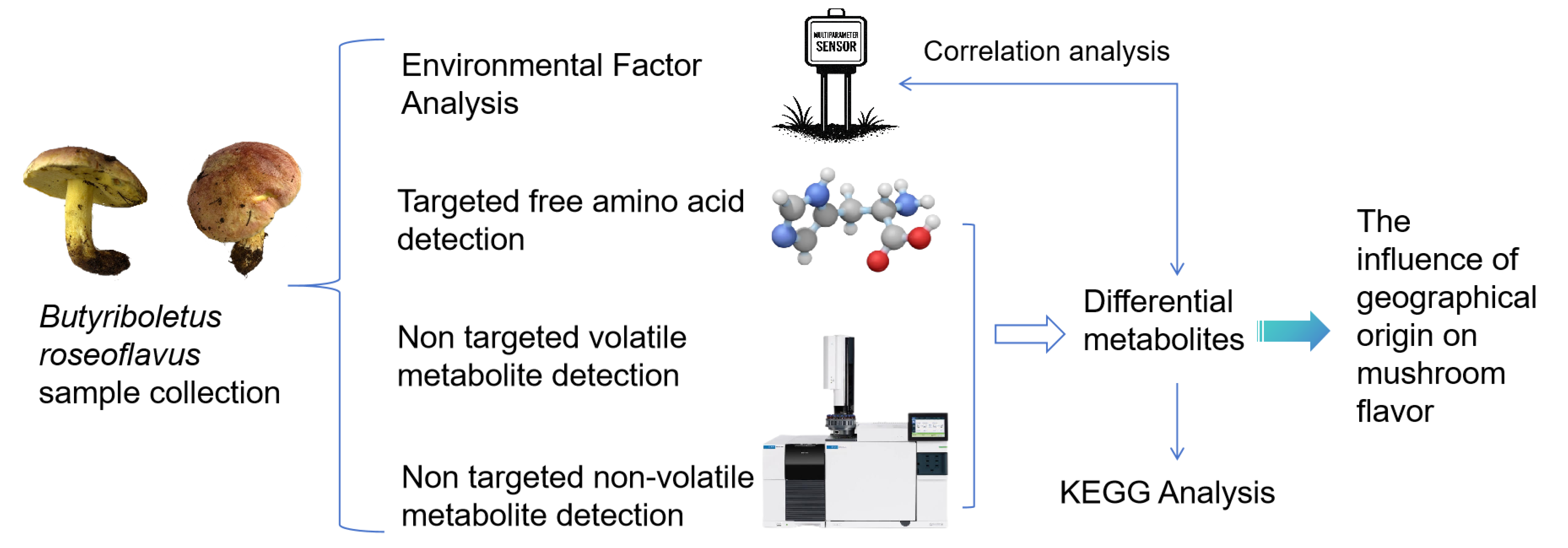
2. Materials and Methods
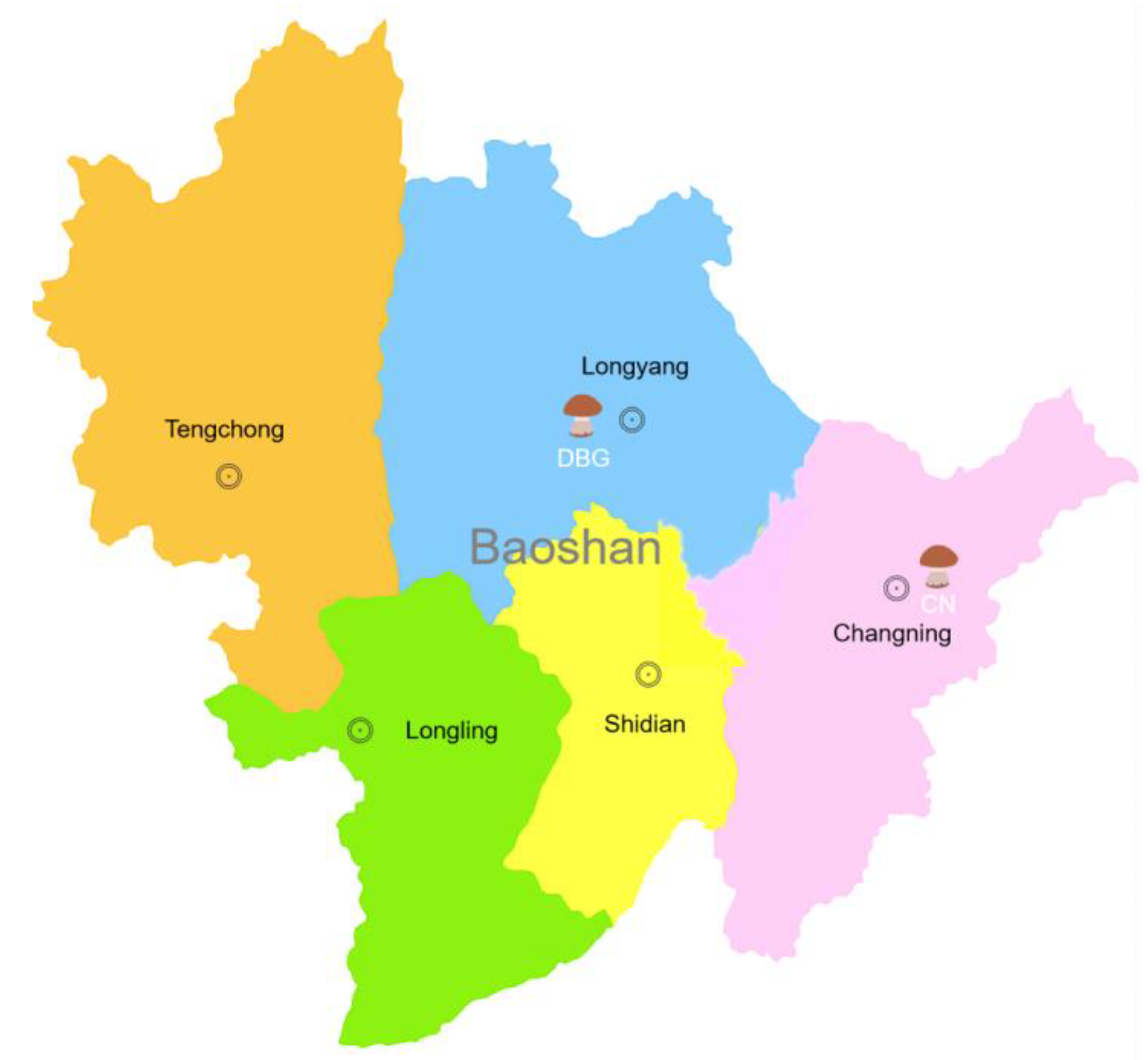
2.1. Sample Preparation
2.2. Determination of Environmental Factors
2.3. Molecular Identification
2.4. HS-SPME-GC–MS Analysis
2.4.1. HS-SPME Extraction Conditions
2.4.2. Chromatographic Conditions (Continued)
2.4.3. Volatile Metabolite Qualitative and Quantitative Analysis
2.5. Free Amino Acid Content Detection
2.6. UPLC-MS/MS Analysis
2.6.1. Sample Extraction
2.6.2. Chromatographic Conditions
2.6.3. Mass Spectrometry Conditions
2.6.4. Metabolite Qualitative and Quantitative Analysis
2.7. Taste Active Value (TAV) Analysis
2.8. Relative Odor Active Value (rOAV) Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. Molecular Characteristics of B. roseoflavis
3.2. Environmental Factor Analysis
3.3. Free Amino Acids (FAAs) Determination
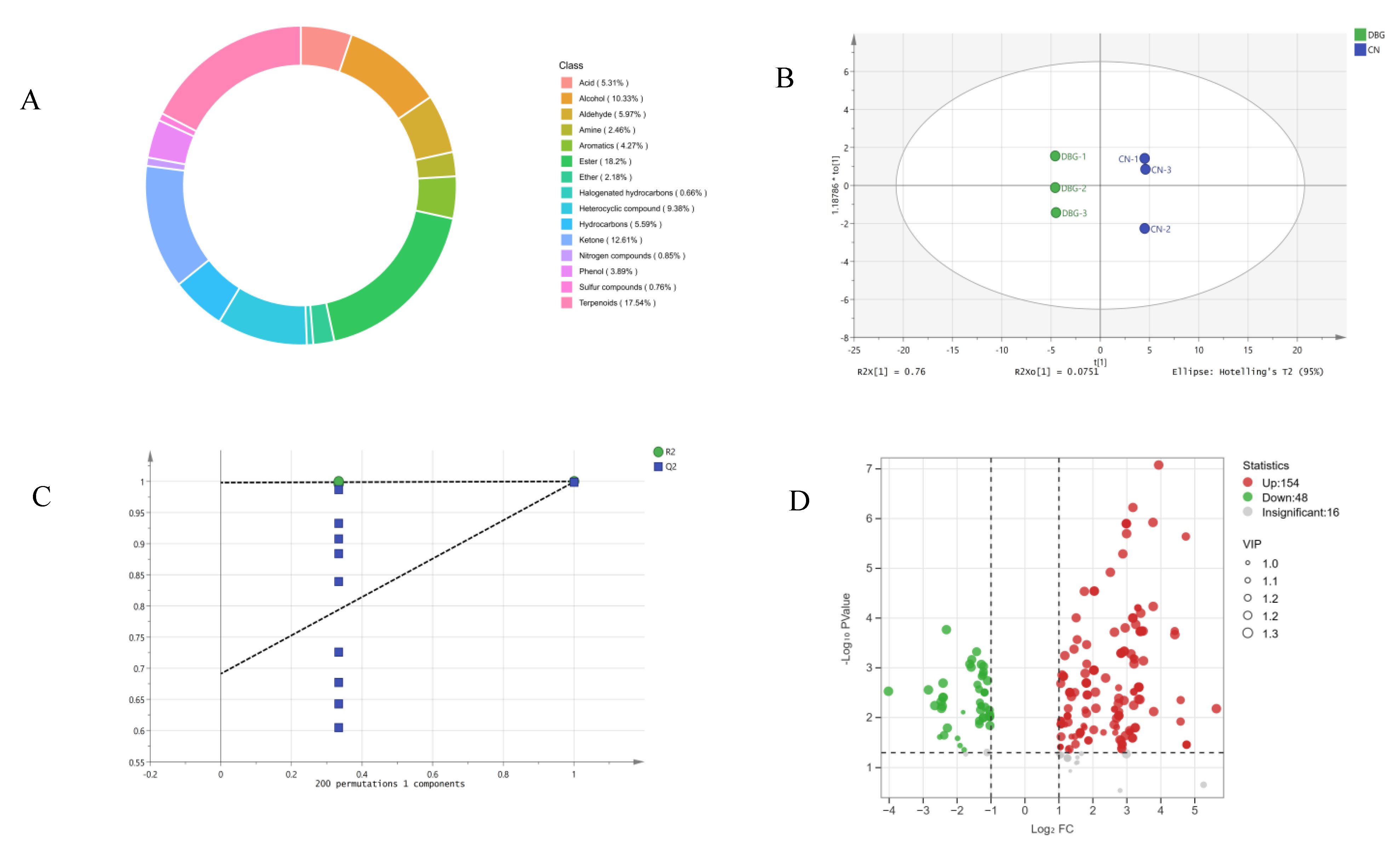
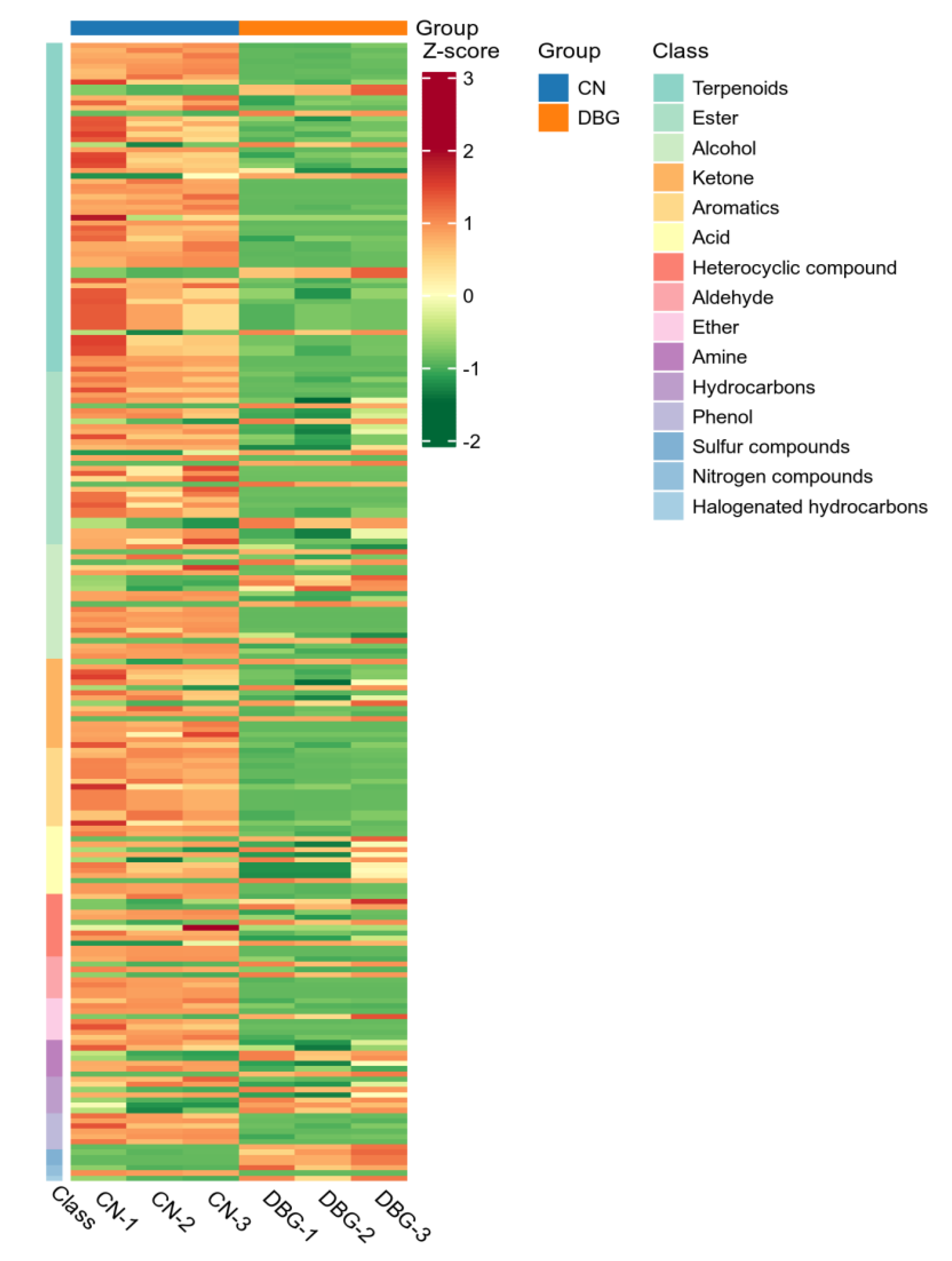
3.4. Aroma Concentration and Composition Analysis of B. roseoflavus Samples
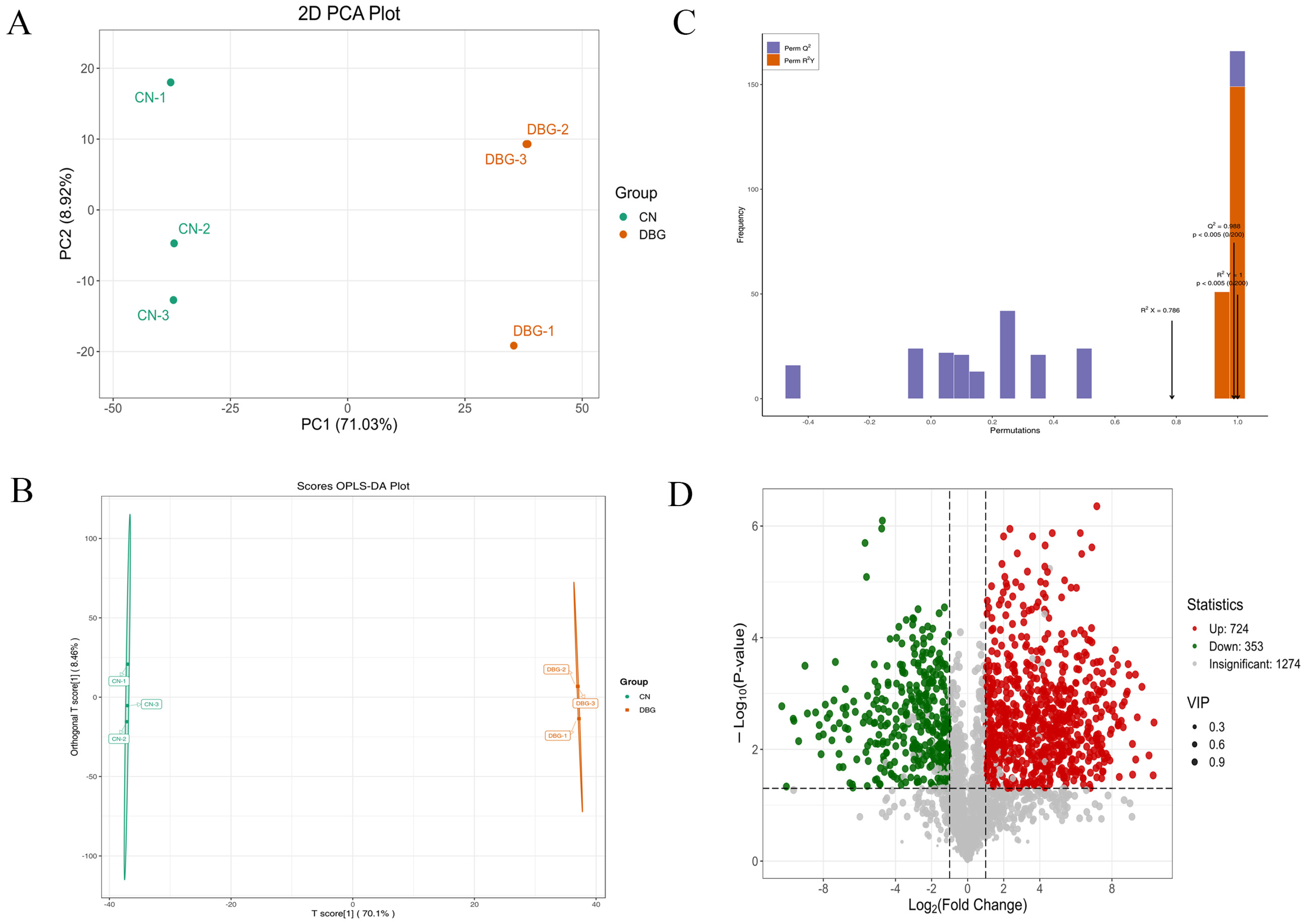
3.5. UPLC-MS/MS Analysis of B. roseoflavus Samples
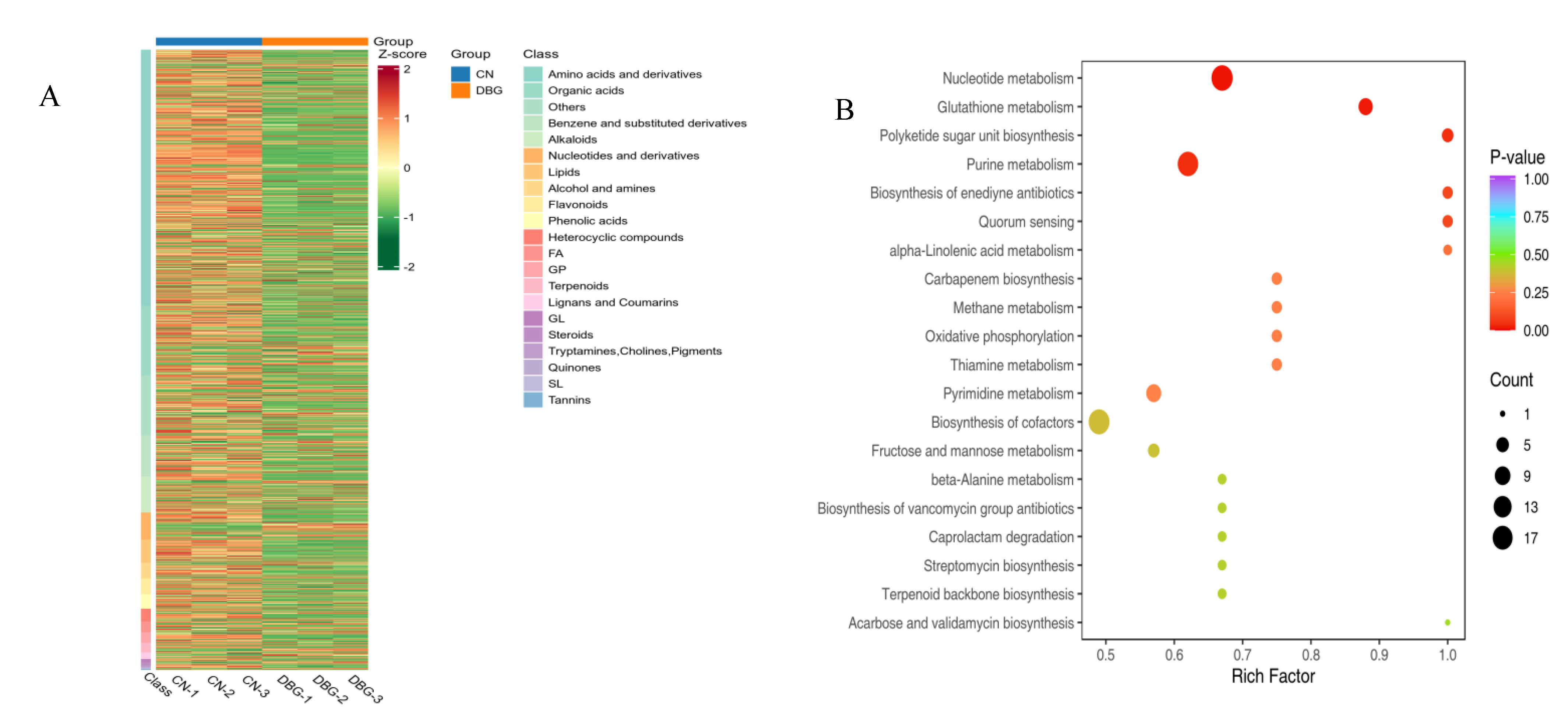
3.6. Differential Metabolite Screening
3.7. KEGG Analysis of Differential Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, X.M.; Zhang, J.; Wu, L.H.; Zhao, Y.L.; Li, T.; Li, J.Q.; Wang, Y.Z.; Liu, H.-G. A Mini-Review of Chemical Composition and Nutritional Value of Edible Wild-Grown Mushroom from China. Food Chem. 2014, 151, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Li, Y.C.; Zhu, X.T.; Zhao, K.; Han, L.H.; Cui, Y.Y.; Li, F.; Xu, J.P.; Yang, Z.L. One Hundred Noteworthy Boletes from China. Fungal Divers. 2016, 81, 25–188. [Google Scholar] [CrossRef]
- Ding, X.; Hou, Y.; Hou, W. Structure Elucidation and Antioxidant Activity of a Novel Polysaccharide Isolated from Boletus Speciosus Forst. Int. J. Biol. Macromol. 2012, 50, 613–618. [Google Scholar] [CrossRef]
- Wang, D.; Sun, S.Q.; Wu, Z.W.; Yang, S.L.; Tan, J.M. Characterization of a Water-Soluble Polysaccharide from Boletus Edulis and Its Antitumor and Immunomodulatory Activities on Renal Cancer in Mice. Carbohydr. Polym. 2014, 105, 127–134. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, R.; Liu, F.; Ng, T.B. Purification and Characterization of a Novel Protein with Activity against Non-Small-Cell Lung Cancer in Vitro and in Vivo from the Edible Mushroom Boletus Edulis. Int. J. Biol. Macromol. 2021, 174, 77–88. [Google Scholar] [CrossRef]
- Su, L.H.; Geng, C.A.; Li, T.Z.; Huang, X.Y.; Ma, Y.B.; Zhang, X.M.; Wu, G.; Yang, Z.L.; Chen, J.J. Spiroseoflosterol, a Rearranged Ergostane-Steroid from the Fruiting Bodies of Butyriboletus Roseof Lavus. J. Nat. Prod. 2020, 83, 1706–1710. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Che, T.; Li, Z.; Zhang, Q.; Hu, X. Identification, Taste Properties and Molecular Docking of Novel Umami Peptides from Butyriboletus Roseoflavus. Eur. Food Res. Technol. 2022, 248, 2665–2677. [Google Scholar] [CrossRef]
- Zhang, J.; Barałkiewicz, D.; Hanć, A.; Falandysz, J.; Wang, Y. Contents and Health Risk Assessment of Elements in Three Edible Ectomycorrhizal Fungi (Boletaceae) from Polymetallic Soils in Yunnan Province, SW China. Biol. Trace Elem. Res. 2020, 195, 250–259. [Google Scholar] [CrossRef]
- Donglu, F.; Wenjian, Y.; Kimatu, B.M.; Liyan, Z.; Xinxin, A.; Qiuhui, H. Comparison of Flavour Qualities of Mushrooms (Flammulina velutipes) Packed with Different Packaging Materials. Food Chem. 2017, 232, 1–9. [Google Scholar] [CrossRef]
- Wang, L.M.; Zhou, Y.F.; Wang, Y.Y.; Bu, H.Y.; Dong, T. Changes in Cell Wall Metabolism and Flavor Qualities of Mushrooms (Agaricus bernardii) under EMAP Treatments during Storage. Food Packag. Shelf Life 2021, 29, 100732. [Google Scholar] [CrossRef]
- Zhuang, J.; Xiao, Q.; Feng, T.; Huang, Q.; Ho, C.-T.; Song, S. Comparative Flavor Profile Analysis of Four Different Varieties of Boletus Mushrooms by Instrumental and Sensory Techniques. Food Res. Int. 2020, 136, 109485. [Google Scholar] [CrossRef]
- Lola, D.; Miliordos, D.E.; Goulioti, E.; Kontoudakis, N.; Myrtsi, E.D.; Haroutounian, S.A.; Kotseridis, Y. Assessment of the Volatile and Non-Volatile Profile of Savatiano PGI Wines as Affected by Various Terroirs in Attica, Greece. Food Res. Int. 2023, 174, 113649. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.F.; Wu, F.R.; Wu, W.; Yu, W.J.; Zhang, G.W.; Huang, X.Y.; Hao, Y.B.; Luo, L.P. Identification and Quality Evaluation of Lushan Yunwu Tea from Different Geographical Origins Based on Metabolomics. Food Res. Int. 2024, 186, 114379. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, M.; Zhen Wu, Z.; Qin, N.; Fu, Y.; Guo, S. Nutrient Composition and Functional Constituents of Daylily from Different Producing Areas Based on Widely Targeted Metabolomics. Food Chem. X 2024, 21, 101239. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Peng, Z.; Jiang, X.; Zhu, Q.; Chen, R.; Wang, W.; Liu, A.; Wu, C.; Ma, C.; Zhang, J. Metabolomics Analysis of Flavor Differences in Shuixian (Camellia sinensis) Tea from Different Production Regions and Their Microbial Associations. Food Chem. 2024, 443, 138542. [Google Scholar] [CrossRef]
- Cai, Y.; Deng, P.; Liu, J.; Luo, Y.; Sangzhu, T.; Li, H.; Zhao, Y.; Tang, C.; Yang, M. Metabolomics-Based Discrimination of Geographical Variability in Quality and Antioxidant Activity of Golden Mushroom (Floccularia luteovirens) from the Qinghai-Tibet Plateau. Food Biosci. 2025, 68, 106536. [Google Scholar] [CrossRef]
- Harun, M.U.; Palma, M.; Setyaningsih, W. Development and Validation of Microwave-Assisted Extraction for Phenolic Compound Profiling in Diverse Oyster Mushrooms (Pleurotus spp.) Sourced from Various Geographical Regions. J. Agric. Food Res. 2025, 20, 101754. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhao, Q.; Deng, H.; Li, Y.; Gong, D.; Huang, X.; Long, D.; Zhang, Y. The Nutrients and Volatile Compounds in Stropharia Rugoso-Annulata by Three Drying Treatments. Foods 2023, 12, 2077. [Google Scholar] [CrossRef]
- Liu, Q.; Hu, S.; Song, Z.; Cui, X.; Kong, W.; Song, K.; Zhang, Y. Relationship between Flavor and Energy Status in Shiitake Mushroom (Lentinula edodes) Harvested at Different Developmental Stages. J. Food Sci. 2021, 86, 4288–4302. [Google Scholar] [CrossRef]
- Xie, L.; Guo, S.; Rao, H.; Lan, B.; Zheng, B.; Zhang, N. Characterization of Volatile Flavor Compounds and Aroma Active Components in Button Mushroom (Agaricus bisporus) across Various Cooking Methods. Foods 2024, 13, 685. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.A.; Gelfand, D.H.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Li, W.; Zi, L.; Xu, N.; Yang, H.; Dong, S.; Qin, F.; Guo, L. Identification of Characteristic Flavor Compounds of Boletus Edulis from Different Regions Based on by E-Nose, HS-GC-IMS and HS-SPME-GC–MS. Food Chem. 2024, 23, 101601. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Cao, G.; Hou, X.; Huang, M.; Du, P.; Tan, T.; Zhang, Y.; Zhou, H.; Liu, X.; Liu, L.; et al. Development of a Widely Targeted Volatilomics Method for Profiling Volatilomes in Plants. Mol. Plant 2022, 15, 189–202. [Google Scholar] [CrossRef]
- National Standard of the People’s Republic of China. National Food Safety Standard—Determination of Amino Acid in Foods. Available online: https://www.chinesestandard.net/PDF.aspx/GB5009.124-2016?PREVIEW (accessed on 11 November 2024).
- Yang, H.; Li, W.; Zi, L.; Xu, N.; Guo, Z.; Chen, B.; Hua, Y.; Guo, L. Comprehensive Analysis of the Dynamic Changes of Volatile and Non-Volatile Metabolites in Boletus Edulis during Processing by HS-SPME-GC–MS and UPLC-MS/MS Analysis. Food Chem. X 2024, 22, 101487. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, M.; Gao, C.; Fu, F.-F.; Wang, T.; El-Kassaby, Y.A.; Wang, G. Amino Acid Metabolism Reprogramming in Response to Changing Growth Environment in Ginkgo Biloba Leaves. LWT 2021, 144, 111276. [Google Scholar] [CrossRef]
- Manninen, H.; Rotola-Pukkila, M.; Aisala, H.; Hopia, A.; Laaksonen, T. Free Amino Acids and 5′-Nucleotides in Finnish Forest Mushrooms. Food Chem. 2018, 247, 23–28. [Google Scholar] [CrossRef]
- Du, X.; Muniz, A.; Davila, M.; Juma, S. Egg White Partially Substituted with Mushroom: Taste Impartment with Mushroom Amino Acids, 5′-Nucleotides, Soluble Sugars, and Organic Acids, and Impact Factors. ACS Food Sci. Technol. 2021, 1, 1333–1348. [Google Scholar] [CrossRef]
- Gonda, I.; Davidovich-Rikanati, R.; Bar, E.; Lev, S.; Jhirad, P.; Meshulam, Y.; Wissotsky, G.; Portnoy, V.; Burger, J.; Schaffer, A.A.; et al. Differential Metabolism of L–Phenylalanine in the Formation of Aromatic Volatiles in Melon (Cucumis melo L.) Fruit. Phytochemistry 2018, 148, 122–131. [Google Scholar] [CrossRef]
- Kochevenko, A.; Araújo, W.L.; Maloney, G.S.; Tieman, D.M.; Do, P.T.; Taylor, M.G.; Klee, H.J.; Fernie, A.R. Catabolism of Branched Chain Amino Acids Supports Respiration but Not Volatile Synthesis in Tomato Fruits. Mol. Plant 2012, 5, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Elazari, Y.; Ovadia, R.; Bar, E.; Nissim-Levi, A.; Carmi, N.; Lewinsohn, E.; Oren-Shamir, M. Phenylalanine Treatment Generates Scent in Flowers by Increased Production of Phenylpropanoid-Benzenoid Volatiles. Postharvest Biol. Technol. 2021, 181, 111657. [Google Scholar] [CrossRef]
- Maoz, I.; Lewinsohn, E.; Gonda, I. Amino Acids Metabolism as a Source for Aroma Volatiles Biosynthesis. Curr. Opin. Plant Biol. 2022, 67, 102221. [Google Scholar] [CrossRef]
- Davila, M.; Muniz, A.; Du, X. The Impact of Roasting and Steaming on Savory Flavors Contributed by Amino Acids, 5′-Nucleotides, and Volatiles in Agaricus Bisporus Mushrooms. Int. J. Gastron. Food Sci. 2022, 30, 100590. [Google Scholar] [CrossRef]
- Kfoury, N.; Morimoto, J.; Kern, A.; Scott, E.R.; Orians, C.M.; Ahmed, S.; Griffin, T.; Cash, S.B.; Stepp, J.R.; Xue, D.; et al. Striking Changes in Tea Metabolites Due to Elevational Effects. Food Chem. 2018, 264, 334–341. [Google Scholar] [CrossRef]
- Lee, J.E.; Lee, B.J.; Chung, J.O.; Hwang, J.A.; Lee, S.J.; Lee, C.H.; Hong, Y.-S. Geographical and Climatic Dependencies of Green Tea (Camellia sinensis) Metabolites: A 1H NMR-Based Metabolomics Study. J. Agric. Food Chem. 2010, 58, 10582–10589. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zang, E.; Sun, S.; Li, M. Main Flavor Compounds and Molecular Regulation Mechanisms in Fruits and Vegetables. Crit. Rev. Food Sci. Nutr. 2023, 63, 11859–11879. [Google Scholar] [CrossRef]
- Wang, M.-Q.; Ma, W.-J.; Shi, J.; Zhu, Y.; Lin, Z.; Lv, H.-P. Characterization of the Key Aroma Compounds in Longjing Tea Using Stir Bar Sorptive Extraction (SBSE) Combined with Gas Chromatography-Mass Spectrometry (GC–MS), Gas Chromatography-Olfactometry (GC-O), Odor Activity Value (OAV), and Aroma Recombination. Food Res. Int. 2020, 130, 108908. [Google Scholar] [CrossRef]
- Yun, J.; Cui, C.; Zhang, S.; Zhu, J.; Peng, C.; Cai, H.; Yang, X.; Hou, R. Use of Headspace GC/MS Combined with Chemometric Analysis to Identify the Geographic Origins of Black Tea. Food Chem. 2021, 360, 130033. [Google Scholar] [CrossRef]
- Hou, Z.S.; Xia, R.R.; Li, Y.T.; Xu, H.R. Key Components, Formation Pathways, Affecting Factors, and Emerging Analytical Strategies for Edible Mushrooms Aroma: A Review. Food Chem. 2024, 438, 137993. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, W.S.; Chen, Q.; Edleman, D.; Annor, G.A.; Dias, F.F.G. Unraveling the Impacts of Germination on the Volatile and Fatty Acid Profile of Intermediate Wheatgrass (Thinopyrum intermedium) Seeds. Molecules 2024, 29, 4268. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, J.B.; Chen, W.C.; Yang, Y.; Zhang, J.S. Analysis of Volatile Compounds of Lentinula Edodes Grown in Different Culture Substrate Formulations. Food Res. Int. 2019, 125, 108517. [Google Scholar] [CrossRef]
- Asproudi, A.; Bonello, F.; Ragkousi, V.; Gianotti, S.; Petrozziello, M. Aroma Precursors of Grignolino Grapes (Vitis vinifera L.) and Their Modulation by Vintage in a Climate Change Scenario. Front. Plant Sci. 2023, 14, 1179111. [Google Scholar] [CrossRef]
- Sirilertpanich, P.; Ekkaphan, P.; Andriyas, T.; Leksungnoen, N.; Ruengphayak, S.; Vanavichit, A.; De-Eknamkul, W.; Tansawat, R. Metabolomics Study on the Main Volatile Components of Thai Colored Rice Cultivars from Different Agricultural Locations. Food Chem. 2024, 434, 137424. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Watanabe, N.; Yang, Z. Understanding the Biosyntheses and Stress Response Mechanisms of Aroma Compounds in Tea (Camellia sinensis) to Safely and Effectively Improve Tea Aroma. Crit. Rev. Food Sci. Nutr. 2018, 59, 2321–2334. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Masood, H.A.; Muhammad, S. Unveiling the Aesthetic Secrets: Exploring Connections between Genetic Makeup, Chemical, and Environmental Factors for Enhancing/Improving the Color and Fragrance/Aroma of Chimonanthus Praecox. PeerJ 2024, 12, e17238. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.J.; Chen, X.; Tan, R.R.; Wang, H.J.; Jiao, L.; Tang, H.Y.; Zong, Q.B.; Mao, Y.X. Comprehensive Metabolomics Analysis of Volatile and Non-Volatile Compounds in Matcha Processed from Different Tea Varieties. Food Chem. 2024, 21, 101234. [Google Scholar] [CrossRef]
- Noctor, G.; Cohen, M.; Trémulot, L.; Châtel-Innocenti, G.; Van Breusegem, F.; Mhamdi, A. Glutathione: A Key Modulator of Plant Defence and Metabolism through Multiple Mechanisms. J. Exp. Bot. 2024, 75, 4549–4572. [Google Scholar] [CrossRef]
- Witte, C.-P.; Herde, M. Nucleotides and Nucleotide Derivatives as Signal Molecules in Plants. J. Exp. Bot. 2024, 75, 6918–6938. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Taste Attribute | FAAs Content (mg/100 g) | |
|---|---|---|---|
| CN | DBG | ||
| Glu | Umami | 766.538 ± 12.28 b | 525.525 ± 2.928 a |
| Asp | Umami | 14.163 ± 0.336 b | 16.154 ± 0.05 a |
| Umami FAAs | 780.702 ± 12.475 b | 541.679 ± 2.896 a | |
| Ala | Sweet | 224.178 ± 4.561 b | 202.069 ± 1.594 a |
| Gly | Sweet | 168.067 ± 5.437 b | 56.984 ± 0.412 a |
| Thr | Sweet | 81.371 ± 1.814 b | 60.639 ± 0.578 a |
| Ser | Sweet | 180.823 ± 7.561 b | 69.907 ± 0.679 a |
| Sweet FAAs | 654.439 ± 18.449 b | 389.6 ± 2.237 a | |
| Arg | Bitter | 44.081 ± 1.168 b | 58.113 ± 0.818 a |
| Ile | Bitter | 46.034 ± 0.623 b | 23.775 ± 0.267 a |
| His | Bitter | 32.294 ± 0.854 b | 25.718 ± 0.291 a |
| Leu | Bitter | 32.327 ± 0.353 b | 28.311 ± 0.514 a |
| Phe | Bitter | 4.866 ± 0.085 a | 4.805 ± 0.095 a |
| Tyr | Bitter | 30.758 ± 0.691 b | 35.116 ± 1.109 a |
| Trp | Bitter | 7.902 ± 0.033 a | 7.941 ± 0.237 a |
| Bitter FAAs | 198.261 ± 3.649 b | 183.778 ± 2.373 a | |
| Lys | Bittersweet | 21.443 ± 0.923 a | 19.845 ± 0.095 a |
| Met | Bittersweet | 6.985 ± 0.206 b | 17.721 ± 0.139 a |
| Pro | Bittersweet | 40.432 ± 0.757 b | 34.826 ± 0.628 a |
| Val | Bittersweet | 19.643 ± 0.418 b | 16.418 ± 0.131 a |
| Bittersweet FAAs | 88.503 ± 2.227 a | 88.809 ± 0.989 a | |
| Asn | Other | 50.736 ± 0.758 b | 44.752 ± 0.499 a |
| Gln | Other | 652.776 ± 7.797 b | 624.854 ± 2.544 a |
| Other FAAs | 703.512 ± 8.525 b | 669.605 ± 2.821 a | |
| Total amount | 2425.416 ± 44.729 b | 1873.471 ± 7.596 a | |
| Taste Compound | Taste Threshold (mg/100 g) | TAV | |
|---|---|---|---|
| CN | DBG | ||
| Glu | 30 | 25.551 ± 0.409 b | 17.518 ± 0.098 a |
| Asp | 53 | 0.267 ± 0.006 b | 0.305 ± 0.001 a |
| Ala | 60 | 25.819 ± 0.413 b | 17.822 ± 0.097 a |
| Gly | 130 | 3.736 ± 0.076 b | 3.368 ± 0.027 a |
| Thr | 260 | 1.293 ± 0.042 b | 0.438 ± 0.003 a |
| Ser | 150 | 0.313 ± 0.007 b | 0.233 ± 0.002 a |
| Arg | 50 | 1.206 ± 0.05 b | 0.466 ± 0.005 a |
| Ile | 90 | 6.548 ± 0.167 b | 4.505 ± 0.029 a |
| His | 20 | 0.882 ± 0.023 b | 1.162 ± 0.016 a |
| Leu | 190 | 0.512 ± 0.007 b | 0.264 ± 0.003 a |
| Phe | 90 | 1.615 ± 0.043 b | 1.286 ± 0.015 a |
| Tyr | 96.6 | 0.17 ± 0.002 b | 0.149 ± 0.003 a |
| Trp | 90 | 0.054 ± 0.001 a | 0.053 ± 0.001 a |
| Lys | 50 | 0.318 ± 0.007 b | 0.364 ± 0.011 a |
| Met | 30 | 0.088 ± 0 a | 0.088 ± 0.003 a |
| Pro | 300 | 3.638 ± 0.082 b | 3.367 ± 0.028 a |
| Val | 40 | 0.429 ± 0.018 a | 0.397 ± 0.002 a |
| Asn | 100 | 0.233 ± 0.007 b | 0.591 ± 0.005 a |
| Gln | 730 | 0.135 ± 0.003 b | 0.116 ± 0.002 a |
| No. | Compounds | Threshold | OAVs | |
|---|---|---|---|---|
| μg/g | DBG | CN | ||
| 1 | Propyl sulfide | 0.003 | 1.302 ± 0.063 b | 0.349 ± 0.031 a |
| 2 | Naphthalene, 2,7-dimethyl- | 0.042 | 1.369 ± 0.088 b | 0.128 ± 0.003 a |
| 3 | Naphthalene, 1,3-dimethyl- | 0.042 | 1.369 ± 0.088 b | 0.128 ± 0.003 a |
| 4 | Pentanoic acid, 4-methyl-, ethyl ester | 0.006 | 1.517 ± 0.043 b | 0.279 ± 0.001 a |
| 5 | 3-Hexen-1-ol, acetate, (Z)- | 0.031 | 3.198 ± 0.538 b | 6.925 ± 0.336 a |
| 6 | Phenol, 2-methoxy-4-propyl- | 0.0044 | 3.561 ± 0.041 b | 0.375 ± 0.039 a |
| 7 | Butane, 1-(ethylthio)- | 0.001 | 3.906 ± 0.19 b | 1.047 ± 0.092 a |
| 8 | 3-mercapto-2-pentanone | 0.0007 | 4.412 ± 0.492 b | 9.761 ± 0.23 a |
| 9 | 5,9-Undecadien-2-one, 6,10-dimethyl-, (E)- | 0.01 | 4.422 ± 0.435 b | 2.016 ± 0.119 a |
| 10 | 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | 0.014 | 5.269 ± 0.324 b | 1.465 ± 0.465 a |
| 11 | (1R)-2,6,6-Trimethylbicyclo [3.1.1]hept-2-ene | 0.0053 | 5.664 ± 0.331 b | 11.086 ± 0.675 a |
| 12 | Naphthalene, 2,6-dimethyl- | 0.01 | 5.752 ± 0.37 b | 0.536 ± 0.014 a |
| 13 | 2,4-Undecadienal, (E,E)- | 0.001 | 6.493 ± 0.165 b | 0.684 ± 0.002 a |
| 14 | Styrene | 0.0036 | 40.645 ± 0.737 b | 11.353 ± 0.405 a |
| 15 | Octanal | 0.0007 | 41.244 ± 4.394 b | 120.834 ± 6.005 a |
| 16 | 1,3,5-Trithiane | 0.00004 | 359.051 ± 37.124 b | 14.2 ± 8.097 a |
| 17 | 2,4-Undecadienal | 0.00001 | 649.305 ± 16.517 b | 68.398 ± 0.17 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, L.; Zhao, Q.; Shao, X.; Lv, X.; Lu, J.; Luo, R.; Liu, Y.; Zhou, X.; Li, Q.; Gui, M. Identification of Characteristic Flavor Compounds and Quality Evaluation of Butyriboletus roseoflavus from Different Regions in Yunnan. Foods 2025, 14, 1676. https://doi.org/10.3390/foods14101676
Song L, Zhao Q, Shao X, Lv X, Lu J, Luo R, Liu Y, Zhou X, Li Q, Gui M. Identification of Characteristic Flavor Compounds and Quality Evaluation of Butyriboletus roseoflavus from Different Regions in Yunnan. Foods. 2025; 14(10):1676. https://doi.org/10.3390/foods14101676
Chicago/Turabian StyleSong, Ling, Qiming Zhao, Xuebin Shao, Xiangqian Lv, Juan Lu, Ruiping Luo, Yurong Liu, Xing Zhou, Qiang Li, and Mingying Gui. 2025. "Identification of Characteristic Flavor Compounds and Quality Evaluation of Butyriboletus roseoflavus from Different Regions in Yunnan" Foods 14, no. 10: 1676. https://doi.org/10.3390/foods14101676
APA StyleSong, L., Zhao, Q., Shao, X., Lv, X., Lu, J., Luo, R., Liu, Y., Zhou, X., Li, Q., & Gui, M. (2025). Identification of Characteristic Flavor Compounds and Quality Evaluation of Butyriboletus roseoflavus from Different Regions in Yunnan. Foods, 14(10), 1676. https://doi.org/10.3390/foods14101676