
The Phase-Dependent Regulation of Lux-Type Genes on the Spoilage Characteristics of Hafnia alvei



Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Reagents
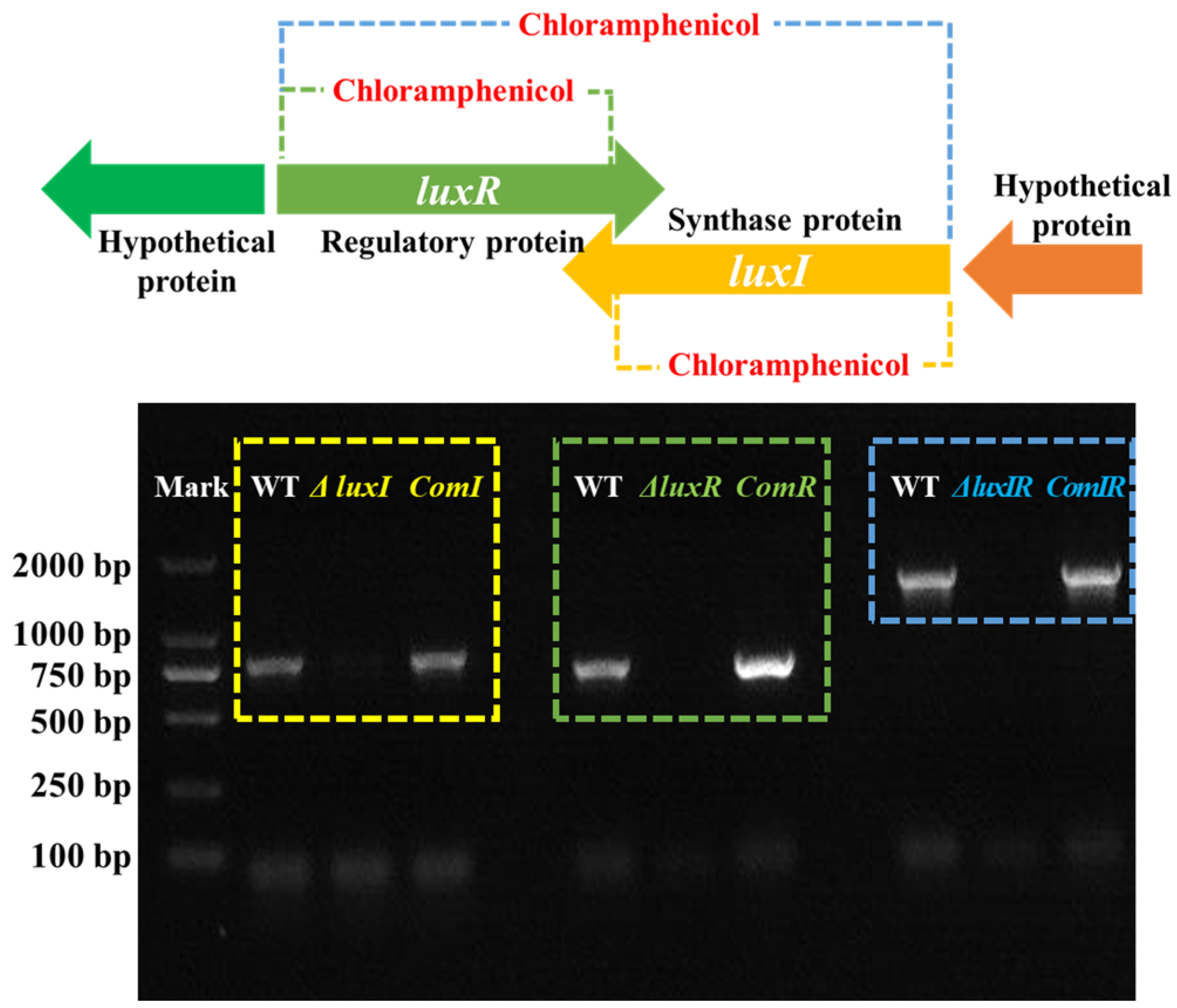
2.2. Construction of luxI and luxR Mutants and Complemented Strains
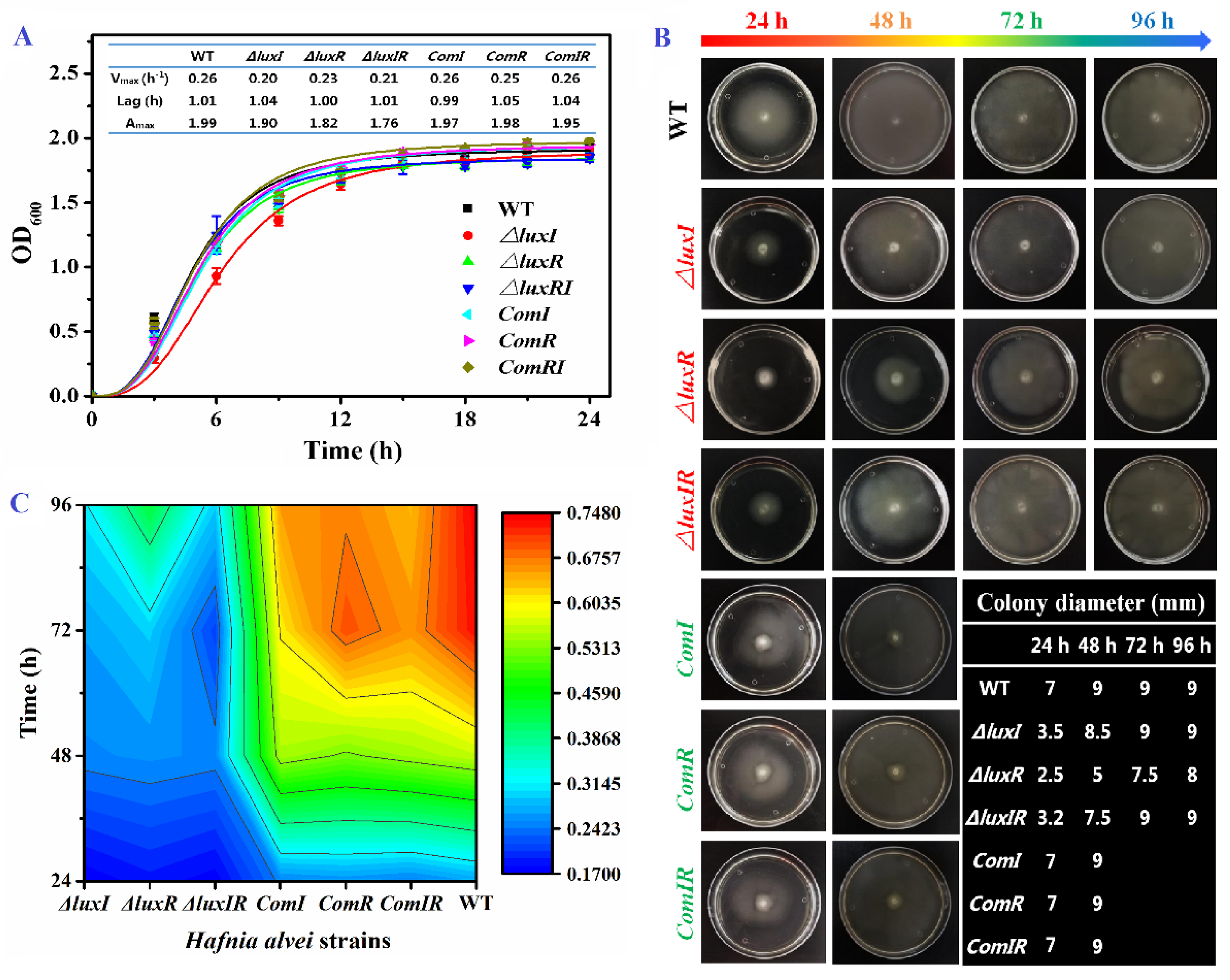
2.3. Growth Curve
2.4. Swimming Motility Assay
2.5. Biofilm Formation Assay
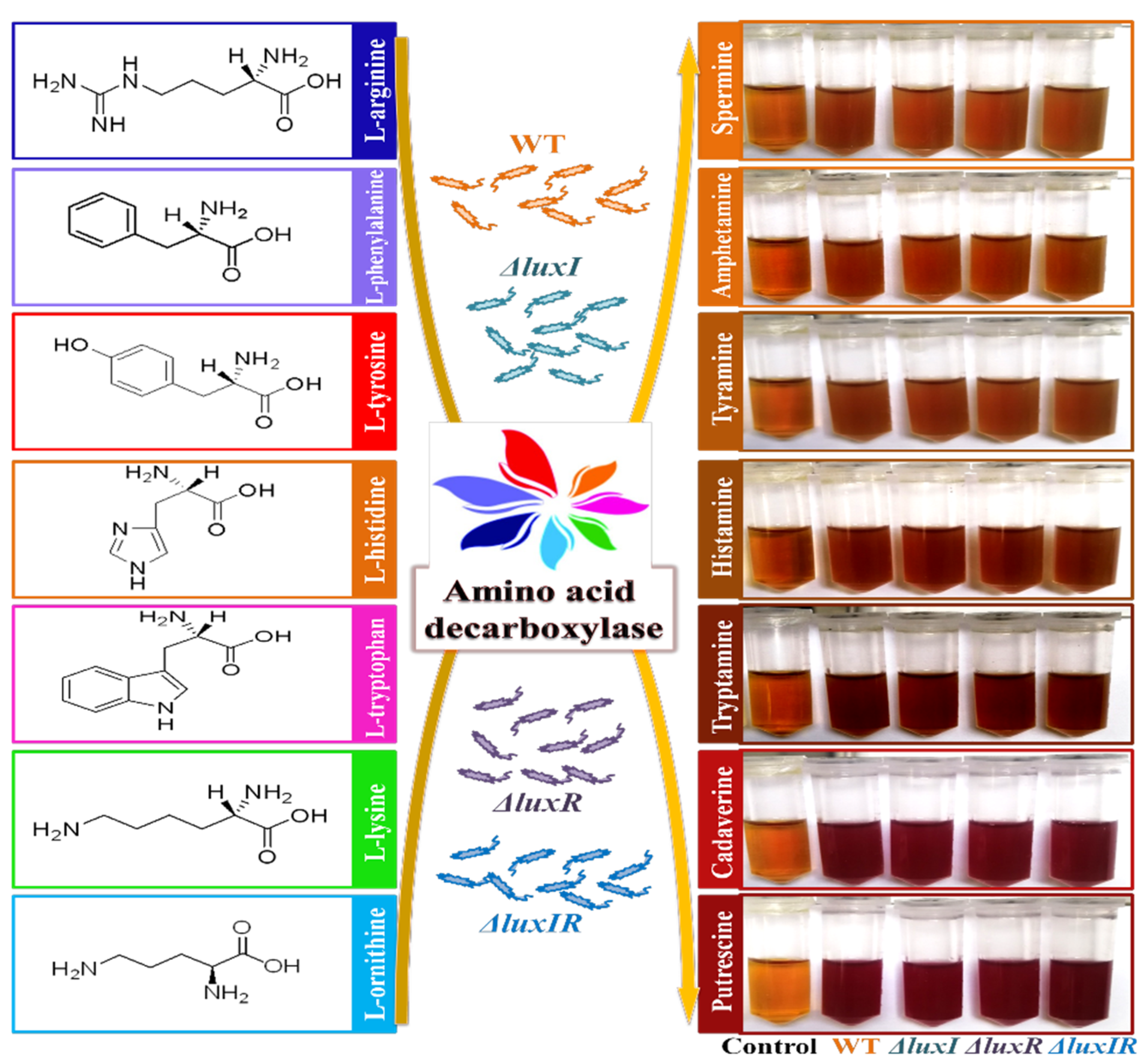
2.6. Decarboxylase Detection
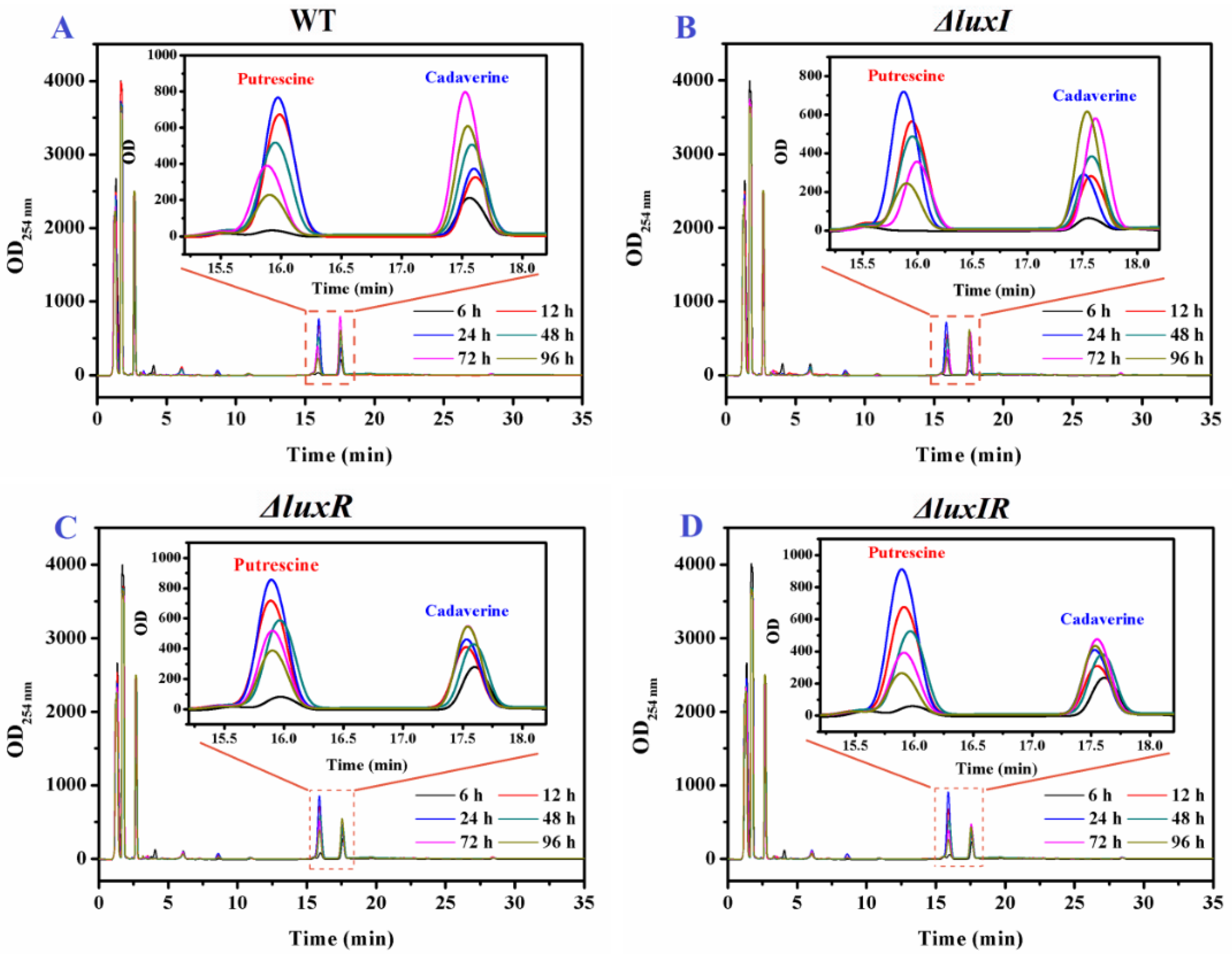
2.7. HPLC Analysis of BA Production
2.8. Statistical Analysis
3. Results
3.1. Mutant and Complementation Strains Construction
3.2. Growth Ability, Swimming Motility and Biofilm Formation
3.3. Decarboxylase Detection
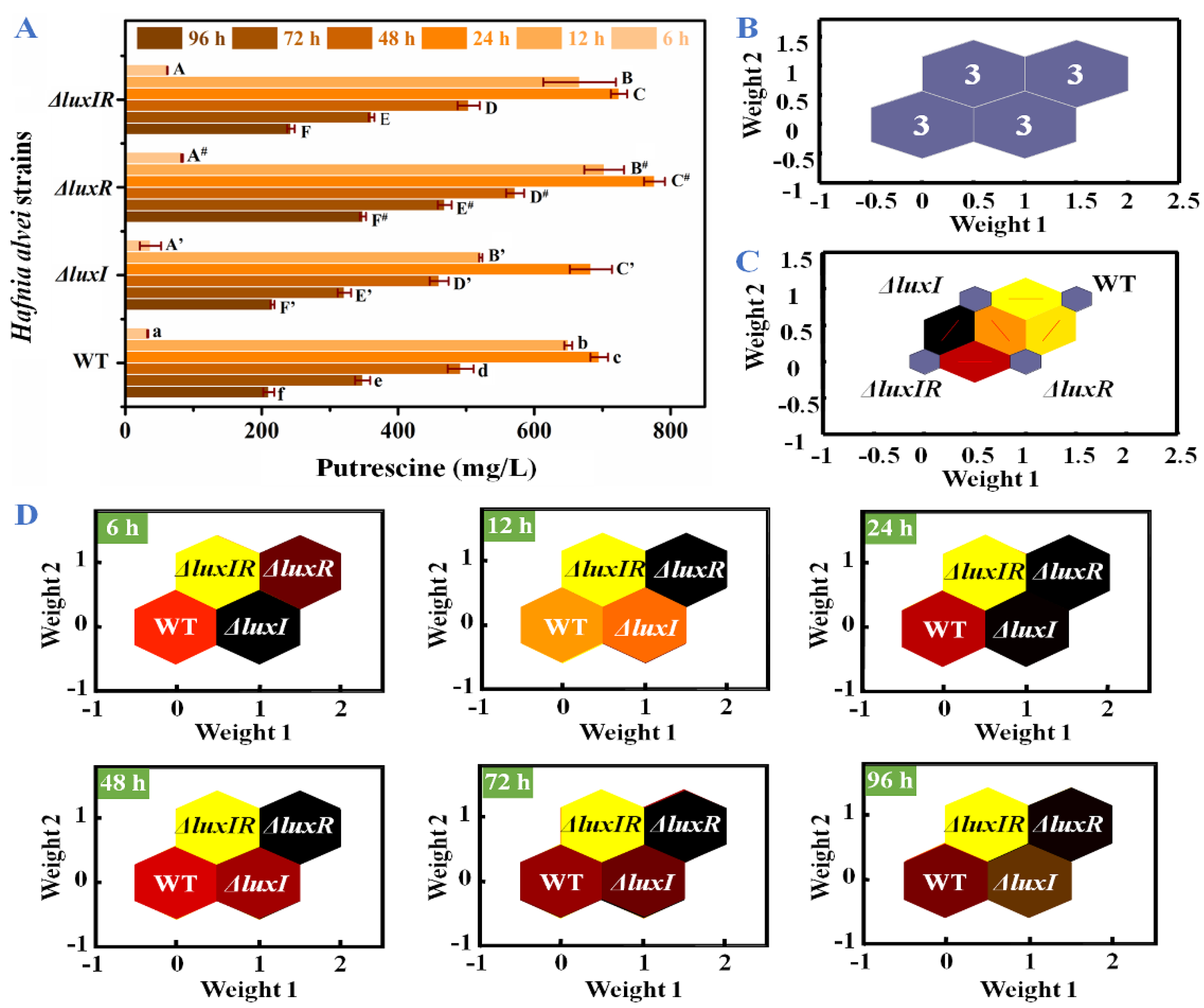
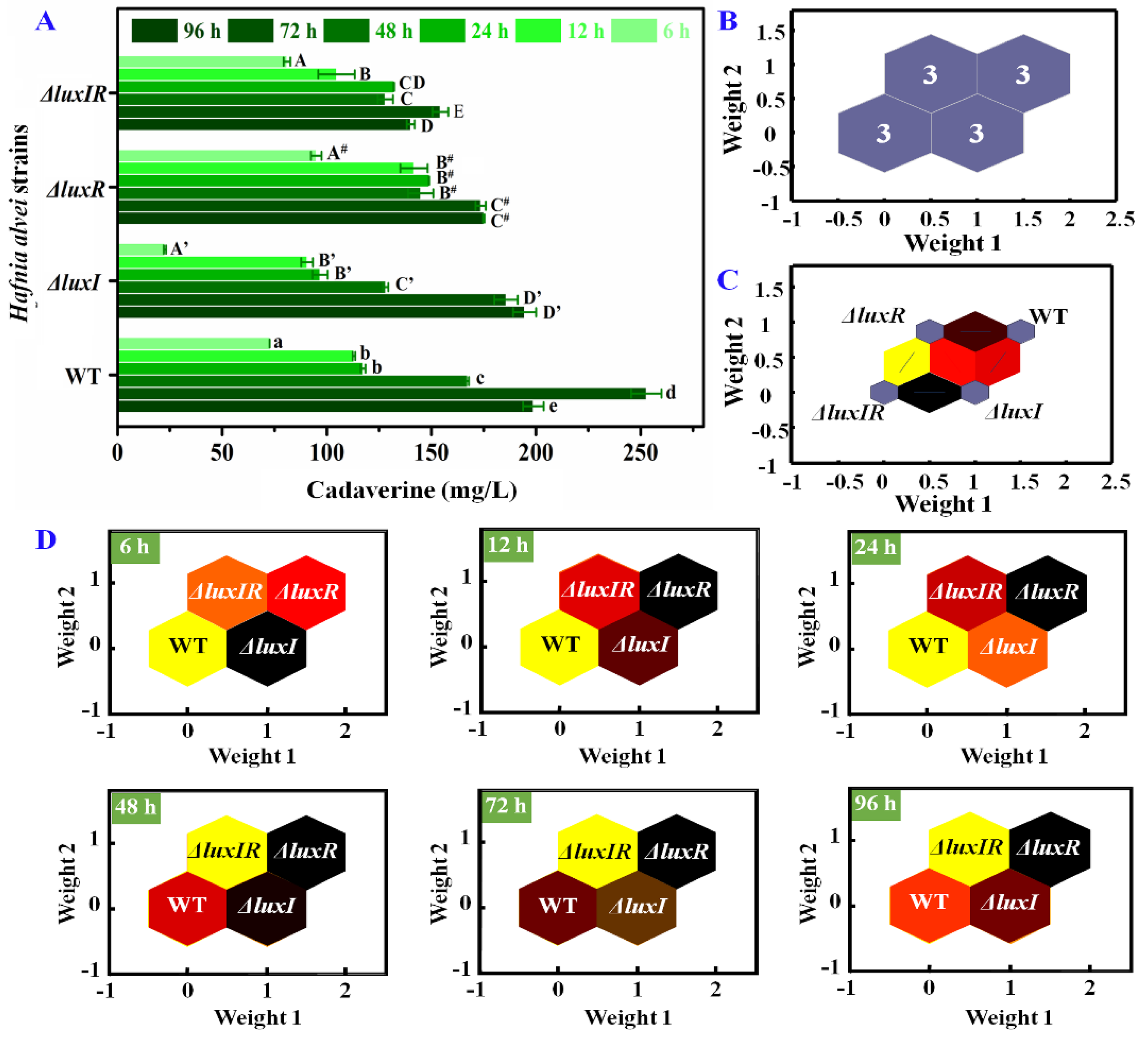
3.4. Putrescine and Cadaverine Production
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davares, A.K.L.; Arsene, M.M.J.; Viktorovna, P.I.; Vyacheslavovna, Y.N.; Vladimirovna, Z.A.; Aleksandrovna, V.E.; Nikolayevich, S.A.; Nadezhda, S.; Anatolievna, G.O.; Nikolaevna, S.I.; et al. Quorum-Sensing Inhibitors from Probiotics as a Strategy to Combat Bacterial Cell-to-Cell Communication Involved in Food Spoilage and Food Safety. Fermentation 2022, 8, 711. [Google Scholar] [CrossRef]
- Ashaolu, T.J.; Khalifa, I.; Mesak, M.A.; Lorenzo, J.M.; Farag, M.A. A comprehensive review of the role of microorganisms on texture change, flavor and biogenic amines formation in fermented meat with their action mechanisms and safety. Crit. Rev. Food Sci. Nutr. 2023, 63, 3538–3555. [Google Scholar] [CrossRef]
- Turna, N.S.; Chung, R.; McIntyre, L. A review of biogenic amines in fermented foods: Occurrence and health effects. Heliyon 2024, 10, e24501. [Google Scholar] [CrossRef]
- Wójcik, W.; Łukasiewicz, M.; Puppel, K. Biogenic amines: Formation, action and toxicity—A review. J. Sci. Food Agric. 2021, 101, 2634–2640. [Google Scholar] [CrossRef]
- Ramos-Vivas, J.; Tapia, O.; Elexpuru-Zabaleta, M.; Pifarre, K.T.; Armas Diaz, Y.; Battino, M.; Giampieri, F. The Molecular Weaponry Produced by the Bacterium Hafnia alvei in Foods. Molecules 2022, 27, 5585. [Google Scholar] [CrossRef]
- Hernandez-Galan, L.; Cattenoz, T.; Le Feunteun, S.; Canette, A.; Briandet, R.; Le-Guin, S.; Guedon, E.; Castellote, J.; Delettre, J.; Dugat Bony, E.; et al. Effect of dairy matrices on the survival of Streptococcus thermophilus, Brevibacterium aurantiacum and Hafnia alvei during digestion. Food Res. Int. (Ott. Ont.) 2017, 100 Pt 1, 477–488. [Google Scholar] [CrossRef]
- Orozova, P.; Sirakov, I.; Chikova, V.; Popova, R.; Al-Harbi, A.H.; Crumlish, M.; Austin, B. Recovery of Hafnia alvei from diseased brown trout, Salmo trutta L., and healthy noble crayfish, Astacus astacus (L.), in Bulgaria. J. Fish Dis. 2014, 37, 891–898. [Google Scholar] [CrossRef]
- Guo, Q.-Y.; Shan, K.; Yang, X.; Jiang, C.-J.; Zhu, L. Inhibitory effects of pH, salinity, and tea polyphenols concentration on the specific spoilage organisms isolated from lightly-salted large yellow croaker (Pseudosciaena crocea). Food Sci. Nutr. 2022, 10, 3062–3071. [Google Scholar] [CrossRef]
- Geeraerts, W.; De Vuyst, L.; Leroy, F.; Van Kerrebroeck, S. Monitoring of volatile production in cooked poultry products using selected ion flow tube-mass spectrometry. Food Res. Int. (Ott. Ont.) 2019, 119, 196–206. [Google Scholar] [CrossRef]
- Merchán, A.V.; Ruiz-Moyano, S.; Hernández, M.V.; Martín, A.; Lorenzo, M.J.; Benito, M.J. Characterization of autochthonal Hafnia spp. strains isolated from Spanish soft raw ewe’s milk PDO cheeses to be used as adjunct culture. Int. J. Food Microbiol. 2022, 373, 109703. [Google Scholar] [CrossRef]
- Zhou, J.-W.; Ruan, L.-Y.; Chen, H.-J.; Luo, H.-Z.; Jiang, H.; Wang, J.-S.; Jia, A.-Q. Inhibition of Quorum Sensing and Virulence in Serratia marcescens by Hordenine. J. Agric. Food Chem. 2019, 67, 784–795. [Google Scholar] [CrossRef]
- Cao, Z.; Liu, Z.; Mao, X. Application of Quorum Sensing in Metabolic Engineering. J. Agric. Food Chem. 2023, 71, 5062–5074. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, B.; Ding, X.; Bin, P.; Yang, Y.; Zhu, G. Regulatory Mechanisms between Quorum Sensing and Virulence in Salmonella. Microorganisms 2022, 10, 2211. [Google Scholar] [CrossRef]
- Wang, Y.; Bian, Z.; Wang, Y. Biofilm formation and inhibition mediated by bacterial quorum sensing. Appl. Microbiol. Biotechnol. 2022, 106, 6365–6381. [Google Scholar] [CrossRef]
- Prazdnova, E.V.; Gorovtsov, A.V.; Vasilchenko, N.G.; Kulikov, M.P.; Statsenko, V.N.; Bogdanova, A.A.; Refeld, A.G.; Brislavskiy, Y.A.; Chistyakov, V.A.; Chikindas, M.L. Quorum-Sensing Inhibition by Gram-Positive Bacteria. Microorganisms 2022, 10, 350. [Google Scholar] [CrossRef]
- Yan, C.; Li, X.; Zhang, G.; Bi, J.; Hao, H.; Hou, H. Quorum Sensing (QS)-regulated target predictions of Hafnia alvei H4 based on the joint application of genome and STRING database. Food Res. Int. (Ott. Ont.) 2022, 157, 111356. [Google Scholar] [CrossRef]
- Li, S.; Wu, S.; Ren, Y.; Meng, Q.; Yin, J.; Yu, Z. Characterization of differentiated autoregulation of LuxI/LuxR-type quorum sensing system in Pseudoalteromonas. Biochem. Biophys. Res. Commun. 2022, 590, 177–183. [Google Scholar] [CrossRef]
- Ridgway, W.; Ward, M.J.; Wetton, B.T. Quorum-sensing induced transitions between bistable steady-states for a cell-bulk ODE-PDE model with lux intracellular kinetics. J. Math. Biol. 2021, 84, 5. [Google Scholar] [CrossRef]
- Hou, H.M.; Zhu, Y.L.; Wang, J.Y.; Jiang, F.; Qu, W.Y.; Zhang, G.L.; Hao, H.S. Characteristics of N-Acylhomoserine Lactones Produced by Hafnia alvei H4 Isolated from Spoiled Instant Sea Cucumber. Sensors 2017, 17, 772. [Google Scholar] [CrossRef]
- Dehio, C.; Meyer, M. Maintenance of broad-host-range incompatibility group P and group Q plasmids and transposition of Tn5 in Bartonella henselae following conjugal plasmid transfer from Escherichia coli. J. Bacteriol. 1997, 179, 538–540. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Hou, H.M.; Zhang, G.L.; Wang, Y.F.; Hao, H.S. AHLs Regulate Biofilm Formation and Swimming Motility of Hafnia alvei H4. Front. Microbiol. 2019, 10, 1330. [Google Scholar] [CrossRef]
- Xia, F.; Zhang, C.; Jiang, Q.; Wu, Z.; Cao, S.; Wu, P.; Gao, Y.; Cheng, X. Microbiome analysis and growth behaviors prediction of potential spoilage bacteria inhabiting harvested edible mushrooms. J. Plant Dis. Prot. 2024, 131, 77–90. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, Z.; Chen, S.; Huang, J.; Li, T.; Duan, C.; Zhang, L.H.; Xu, Z. CzcR Is Essential for Swimming Motility in Pseudomonas aeruginosa during Zinc Stress. Microbiol. Spectr. 2022, 10, e0284622. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, Y.; Dong, P.; Ni, L.; Luo, X.; Zhang, Y.; Zhu, L. Inhibitory effects of clove and oregano essential oils on biofilm formation of Salmonella Derby isolated from beef processing plant. LWT 2022, 162, 113486. [Google Scholar] [CrossRef]
- Chang, M.; Chang, H.C. Development of a screening method for biogenic amine producing Bacillus spp. Int. J. Food Microbiol. 2012, 153, 269–274. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Zhang, G.; Bi, J.; Hou, H. Transcriptome Reveals Regulation of Quorum Sensing of Hafnia alvei H4 on the Coculture System of Hafnia alvei H4 and Pseudomonas fluorescens ATCC13525. Foods 2024, 13, 336. [Google Scholar] [CrossRef]
- Hongman, H.; Yifang, W.; Gongliang, Z.; Yaolei, Z.; Longquan, X.; Hongshun, H.; Yue, W.; Meishan, L.J. Effects of Sulfide Flavors on AHL-Mediated Quorum Sensing and Biofilm Formation of Hafnia alvei. J. Food Sci. 2018, 83, 2550–2559. [Google Scholar] [CrossRef]
- Yan, H.; Liu, C.; Yu, W.; Zhu, X.; Chen, B. The aggregate distribution of Pseudomonas aeruginosa on biochar facilitates quorum sensing and biofilm formation. Sci. Total Environ. 2023, 856, 159034. [Google Scholar] [CrossRef]
- Law, S.K.K.; Tan, H.S. The role of quorum sensing, biofilm formation, and iron acquisition as key virulence mechanisms in Acinetobacter baumannii and the corresponding anti-virulence strategies. Microbiol. Res. 2022, 260, 127032. [Google Scholar] [CrossRef]
- Li, T.; Mei, Y.; He, B.; Sun, X.; Li, J. Reducing Quorum Sensing-Mediated Virulence Factor Expression and Biofilm Formation in Hafnia alvei by Using the Potential Quorum Sensing Inhibitor L-Carvone. Front. Microbiol. 2018, 9, 3324. [Google Scholar] [CrossRef]
- Yang, Q.; Sun, D.W.; Cheng, W. Development of simplified models for nondestructive hyperspectral imaging monitoring of TVB-N contents in cured meat during drying process. J. Food Eng. 2017, 192, 53–60. [Google Scholar] [CrossRef]
- Zhang, T.; Yang, M.; Pan, H.; Li, S.; Ren, B.; Ren, Z.; Xing, N.; Qi, L.; Ren, Q.; Xu, S.; et al. Does time difference of the acetylcholinesterase (AChE) inhibition in different tissues exist? A case study of zebra fish (Danio rerio) exposed to cadmium chloride and deltamethrin. Chemosphere 2017, 168, 908–916. [Google Scholar] [CrossRef]
- Yan, C.; Li, X.; Zhang, G.; Zhu, Y.; Bi, J.; Hao, H.; Hou, H. Quorum Sensing-Mediated and Growth Phase-Dependent Regulation of Metabolic Pathways in Hafnia alvei H4. Front. Microbiol. 2021, 12, 567942. [Google Scholar] [CrossRef]
- Choi, Y.; Park, H.-Y.; Park, S.J.; Park, S.-J.; Kim, S.-K.; Ha, C.; Im, S.-J.; Lee, J.-H. Growth phase-differential quorum sensing regulation of anthranilate metabolism in Pseudomonas aeruginosa. Mol. Cells 2011, 32, 57–65. [Google Scholar] [CrossRef]
- Boo, A.; Ledesma Amaro, R.; Stan, G.-B. Quorum sensing in synthetic biology: A review. Curr. Opin. Syst. Biol. 2021, 28, 100378. [Google Scholar] [CrossRef]
- Zhu, Y.; Sang, X.; Li, X.; Zhang, Y.; Hao, H.; Bi, J.; Zhang, G.; Hou, H. Effect of quorum sensing and quorum sensing inhibitors on the expression of serine protease gene in Hafnia alvei H4. Appl. Microbiol. Biotechnol. 2020, 104, 7457–7465. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain or Plasmid | Relevant Genotype or Description | Reference or Source |
|---|---|---|
| H. alvei H4 | Genbank ID: GCA_008362885.1, wild-type strain | Isolated from spoiled instant sea cucumber by our lab [19] |
| ΔluxI | H. alvei H4 derivative, luxI mutant strain | This study |
| ΔluxR | H. alvei H4 derivative, luxR mutant strain | This study |
| ΔluxIR | H. alvei H4 derivative, luxIR mutant strain | This study |
| comI | luxI complementation strain, luxI- containing pUC19 | This study |
| comR | luxR complementation strain, luxR-containing pUC19 | This study |
| comIR | luxIR complementation strain, luxIR-containing pUC19 | This study |
| E.coli TOP10 | DH10b derivative of MG1655 (Genbank ID: GCA_000005845.2), recipient strain | Purchased from Takara |
| E. coli β2155 | thrB1004 pro thi strA hsdS lacZ1M15 (F′ lacZ1M15 laclq traD36 proA+ proB+)1dap:: erm (Ermr))recA:: RPA-2-tet(Tcr)::Mu-km (Kmr) λpi [20] chloramphenicol resistance gene cassette (CmR) | Purchased from Takara |
| PCVD442 | Suicide plasmid, SacB, oriT, ampicillin resistance gene cassette (AmR) | Purchased from Songon |
| pUC 19 | GenBank ID: M77789, AmR | Purchased from Takara |
| pET28a(+)/FaGH17A | GenBank ID: CDF79584.1, kanamycin resistance gene cassette (KmR) | Purchased from Takara |
| Sequence (5′-3′, Restriction Enzyme Sites are Underlined) | Restriction Enzyme |
|---|---|
| ATAGAATTCGTCGACATCACATTGATGTCAGACCTCAAGATTTC | EcoRI-SalI |
| ATAGGATCCATATCTGAGTGAGGATGAGCGAATTTATC | BamHI |
| TATGAATTCGTCGACATCAACATGCTCCCAATATCGCAC | EcoRI-SalI |
| TATGGATCCTTGGGCTCCTAGACGTTCAATTTCC | BamHI |
| ATAGGATCCATATGAATATCCTCCTTAGTTCCTATTC | BamHI |
| ATAGGATCCGAGCTGCTTCGAAGTTCCTA | BamHI |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bi, J.; Yao, Q.; Zhang, G.; Hou, H. The Phase-Dependent Regulation of Lux-Type Genes on the Spoilage Characteristics of Hafnia alvei. Foods 2024, 13, 688. https://doi.org/10.3390/foods13050688
Bi J, Yao Q, Zhang G, Hou H. The Phase-Dependent Regulation of Lux-Type Genes on the Spoilage Characteristics of Hafnia alvei. Foods. 2024; 13(5):688. https://doi.org/10.3390/foods13050688
Chicago/Turabian StyleBi, Jingran, Qiaoli Yao, Gongliang Zhang, and Hongman Hou. 2024. "The Phase-Dependent Regulation of Lux-Type Genes on the Spoilage Characteristics of Hafnia alvei" Foods 13, no. 5: 688. https://doi.org/10.3390/foods13050688
APA StyleBi, J., Yao, Q., Zhang, G., & Hou, H. (2024). The Phase-Dependent Regulation of Lux-Type Genes on the Spoilage Characteristics of Hafnia alvei. Foods, 13(5), 688. https://doi.org/10.3390/foods13050688