

Screening, Identification, and Fermentation Condition Optimization of a High-Yield 3-Methylthiopropanol Yeast and Its Aroma-Producing Characteristics
 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials, Reagents, and Media
2.2. Screening for Strain with High Yield of 3-Met
2.3. Identification and Biochemical Characteristics of S. fibuligera Y1402
2.3.1. Morphological Observation
2.3.2. Physiological and Biochemical Identification
2.3.3. Molecular Biology Identification
2.3.4. Determination of S. fibuligera Performance
2.3.5. Production of Aromatic Compounds
2.4. Optimization of Fermentation Conditions for 3-Met Production
2.4.1. Single-Factor Experiment
2.4.2. Plackett–Burman Design
2.4.3. Steepest Ascent Test
2.4.4. Response Surface Analysis
2.5. Scale-Up Fermentation in Fermenter
2.6. Analytical Methods
2.7. Statistical Analysis
3. Results and Discussion
3.1. Screening of High-Yield 3-Met Strains
3.2. Identification of Yeast Y1402
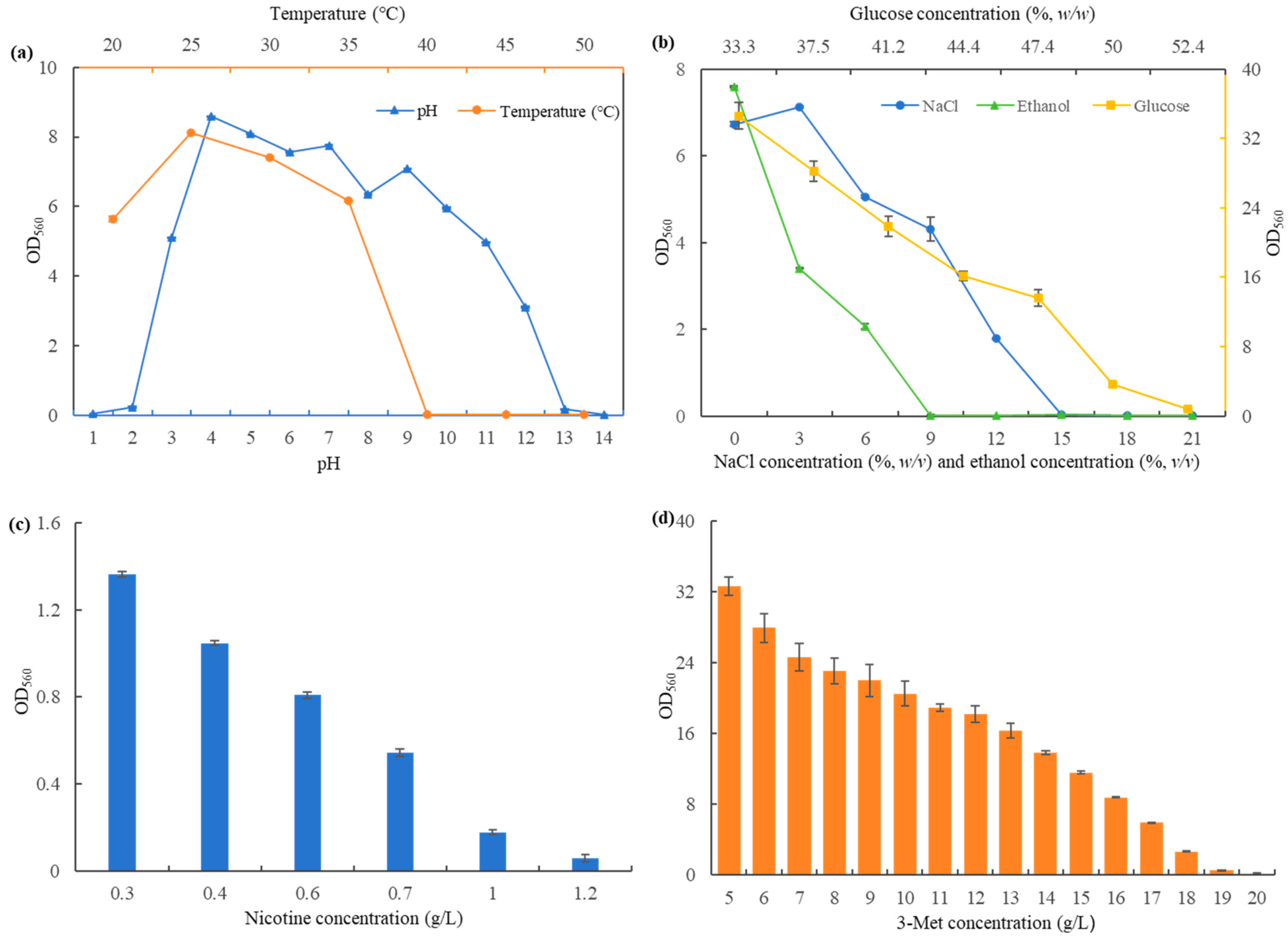
3.3. Determination of S. fibuligera Y1402 Performance
3.4. Optimization of S. fibuligera Y1402 Fermentation Conditions for 3-Met Production
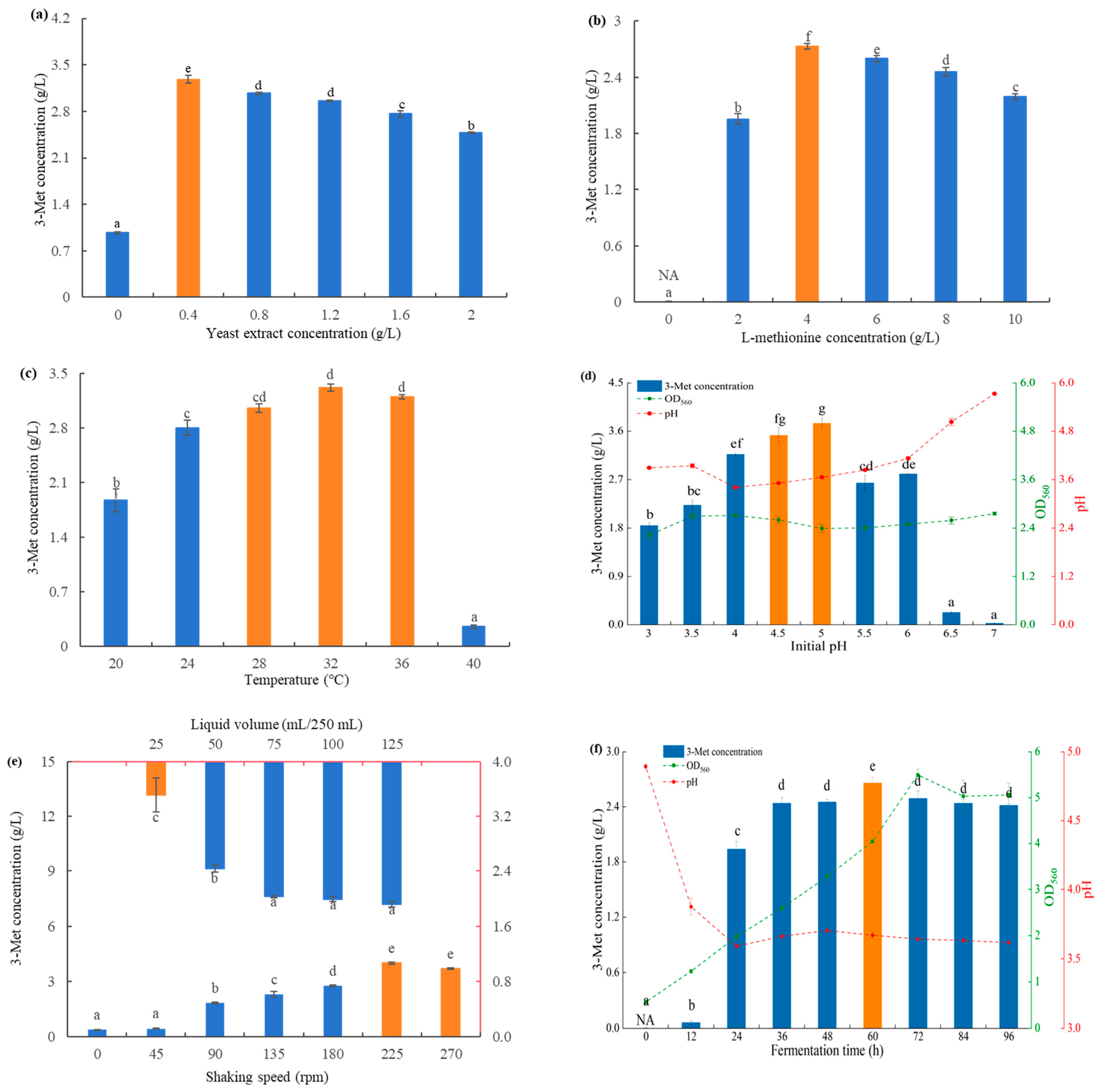
3.4.1. Single-Factor Experiment
3.4.2. Plackett–Burman Design
3.4.3. Steepest Ascent Test
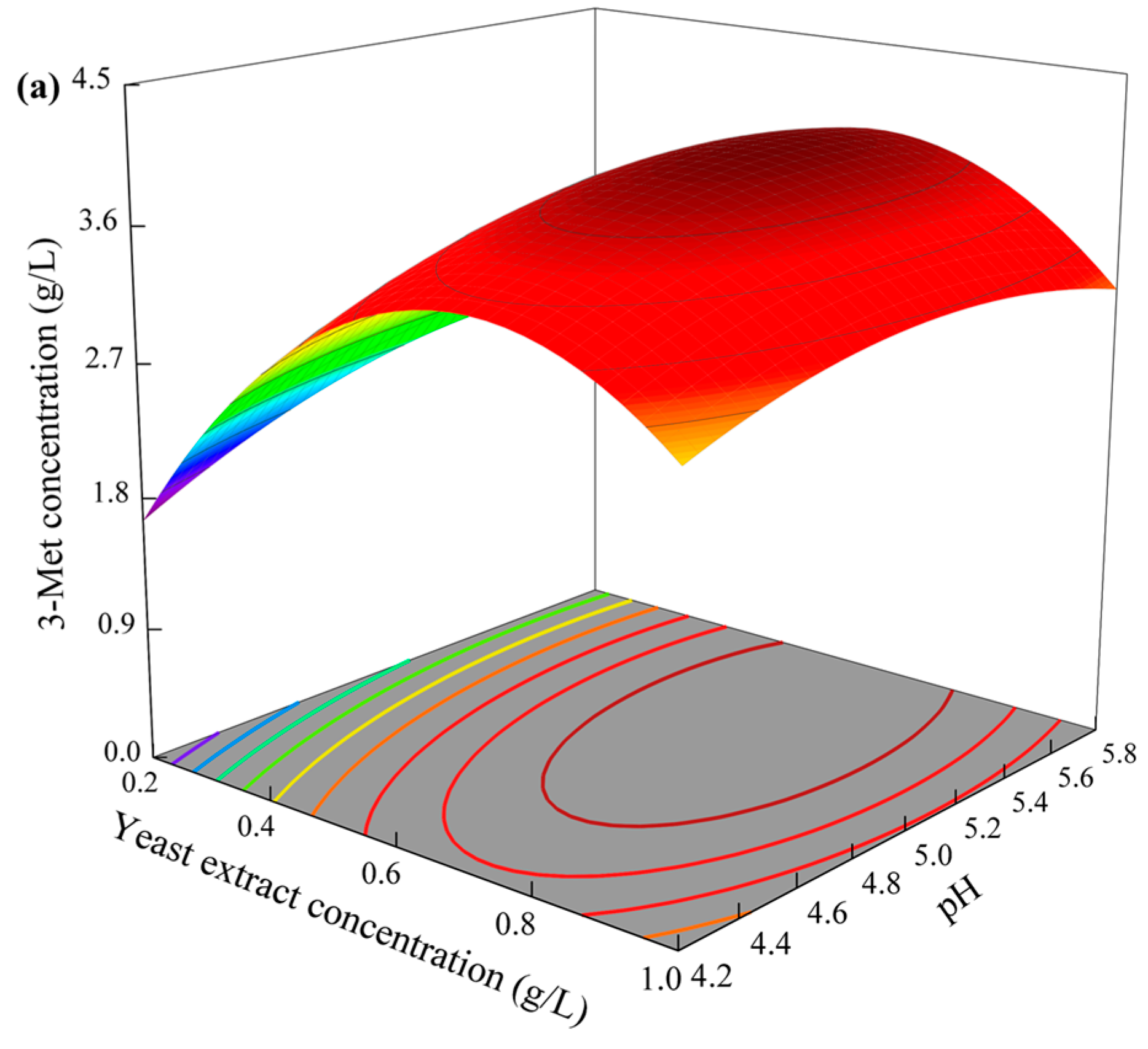
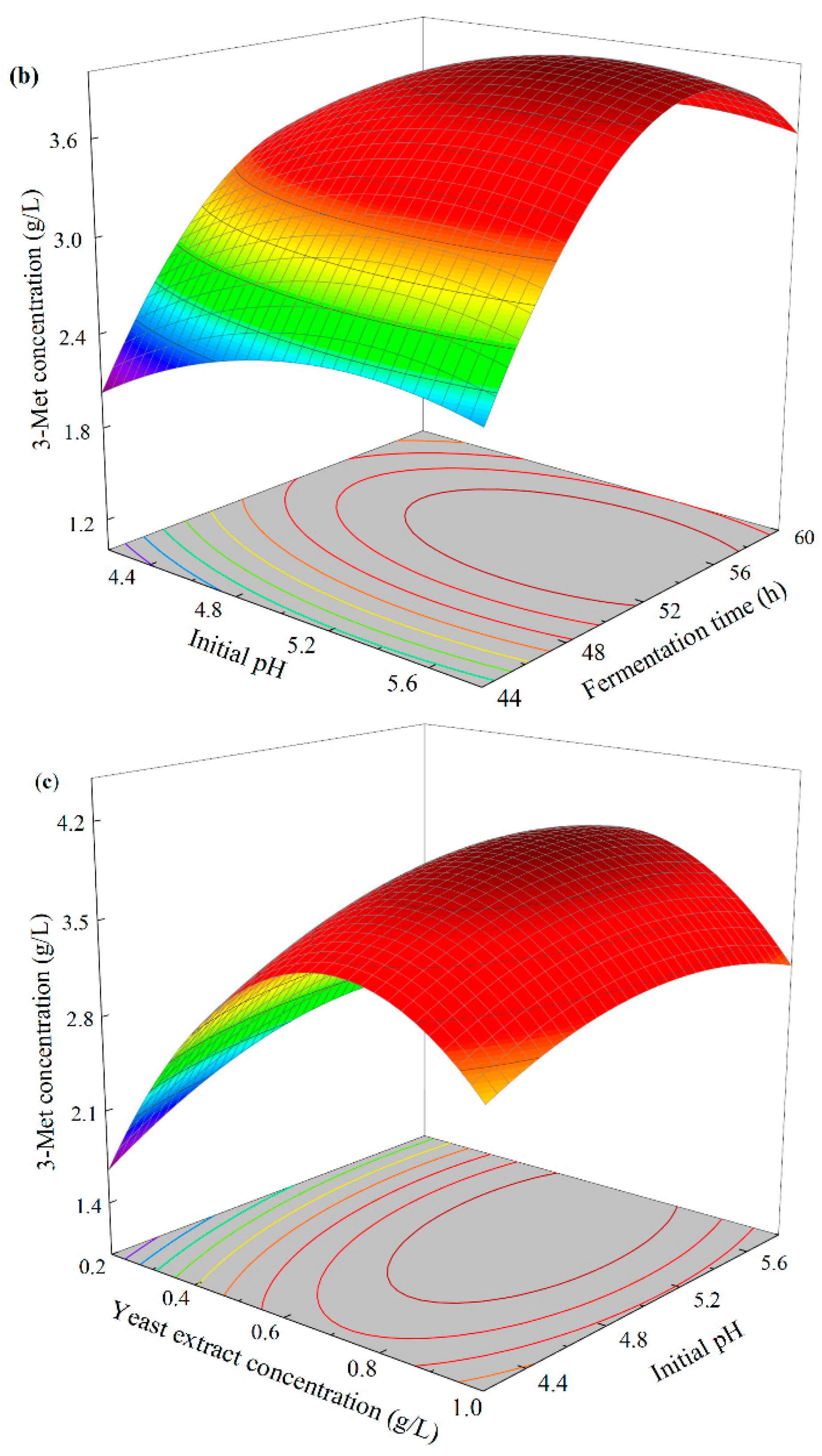
3.4.4. Response Surface Analysis
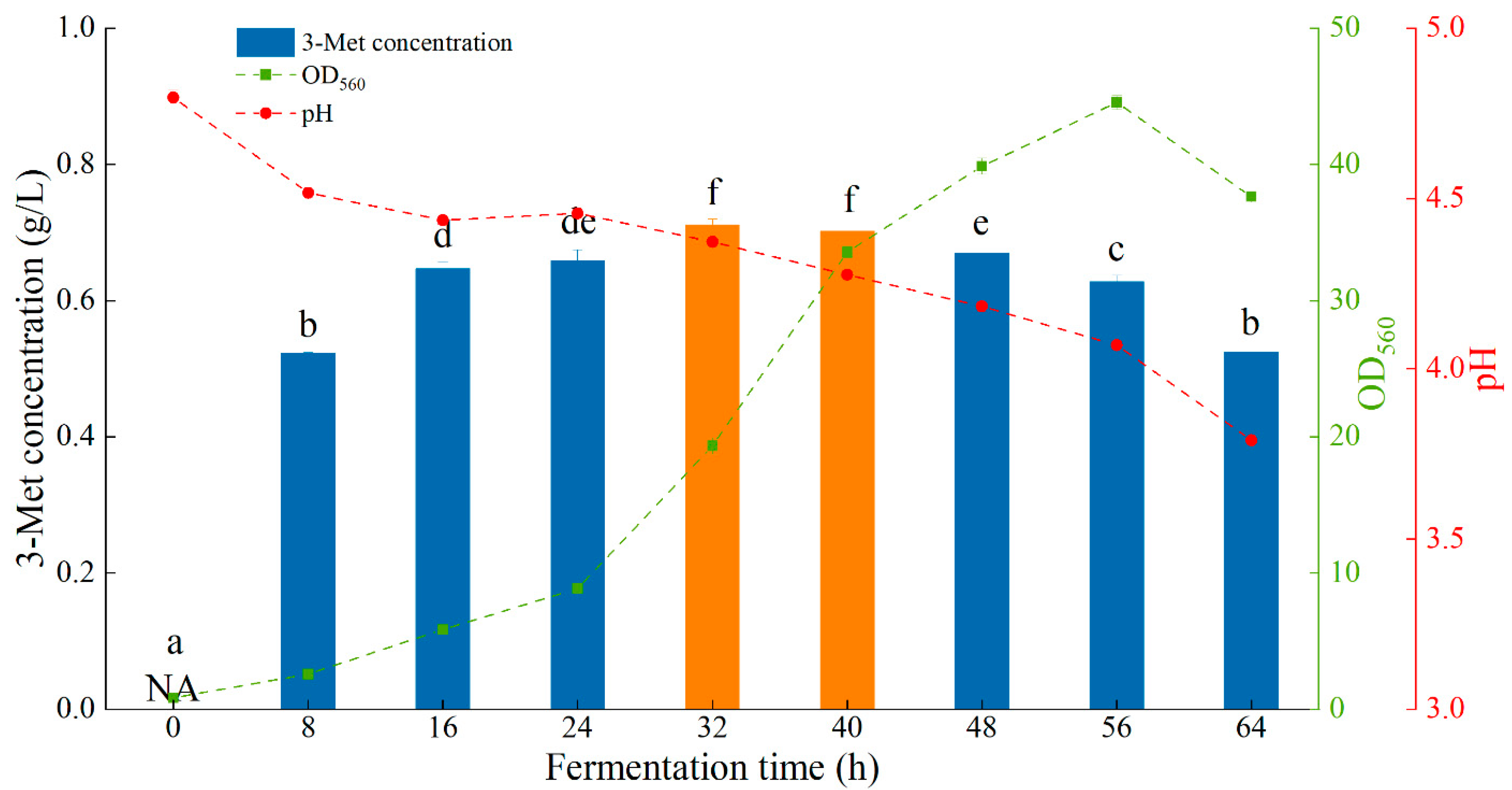
3.5. Scale-Up Fermentation in a 3 L Fermenter
3.6. Production of Aromatic Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seow, Y.X.; Ong, P.; Liu, S.Q. Production of flavour-active methionol from methionine metabolism by yeasts in coconut cream. Int. J. Food Microbiol. 2010, 143, 235–240. [Google Scholar] [CrossRef]
- Che, Y.X.; Yin, S.; Wang, H.; Yang, H.Q.; Xu, R.X.; Wang, Q.; Wu, Y.P.; Boutet, J.; Huet, R.; Wang, C.T. Production of methionol from 3-methylthiopropionaldehyde by catalysis of the yeast alcohol dehydrogenase Adh4p. J. Agric. Food Chem. 2020, 68, 4650–4656. [Google Scholar] [CrossRef]
- Lwa, H.Q.; Sun, J.C.; Liu, S.Q. Optimization of methionol bioproduction by Saccharomyces cerevisiae using response surface methodology. Ann. Microbiol. 2015, 65, 197–205. [Google Scholar] [CrossRef]
- Ma, J.H.; Wang, Y.; Zhang, Y.J.; Cheng, L.J.; Teng, C.; Li, X.T.; Fan, G.S. Research progress of microbial synthesis of 3-methylthiopropanol. China Brew. 2022, 41, 1–7. [Google Scholar] [CrossRef]
- Etschmann, M.; Kotter, P.; Hauf, J.; Bluemke, W.; Entian, K.D.; Schrader, J. Production of the aroma chemicals 3-(methylthio)-1-propanol and 3-(methylthio)-propylacetate with yeasts. Appl. Microbiol. Biotechnol. 2008, 80, 579–587. [Google Scholar] [CrossRef]
- Yang, X.L.; Zhang, K.L.; Li, J.Y.; Zhang, Q.C.; Wang, C.T. Optimization of fermentation conditions for producing 3-methylthiopropanol in Saccharomyces cerevisiae by response surface methodology. J. Univ. Hebei 2014, 34, 398–404. [Google Scholar] [CrossRef]
- Ma, J.H.; Cheng, L.J.; Zhang, Y.J.; Liu, Y.C.; Sun, Q.; Zhang, J.; Liu, X.Y.; Fan, G.S. Screening of yeasts isolated from Baijiu environments for producing 3-methylthio-1-propanol and optimizing production conditions. Foods. 2022, 11, 3616. [Google Scholar] [CrossRef]
- Lopez Del Castillo-Lozano, M.; Delile, A.; Spinnler, H.E.; Bonnarme, P.; Landaud, S. Comparison of volatile sulphur compound production by cheese-ripening yeasts from methionine and methionine-cysteine mixtures. Appl. Microbiol. Biotechnol. 2007, 75, 1447–1454. [Google Scholar] [CrossRef]
- Williams, A.G.; Noble, J.; Banks, J.M. Catabolism of amino acids by lactic acid bacteria isolated from Cheddar cheese. Int. Dairy J. 2001, 11, 203–215. [Google Scholar] [CrossRef]
- Vallet, A.; Lucas, P.; Lonvaud-Funel, A.; de Revel, G. Pathways that produce volatile sulphur compounds from methionine in Oenococcus oeni. J. Appl. Microbiol. 2008, 104, 1833–1840. [Google Scholar] [CrossRef]
- Hou, S.; Zhao, L.; Yang, X.L.; Wang, C.T. Study on conditions of bio-transformation of 3-(methylthio)-1-propanol by Saccharomyces Cerevisiae SC408. J. Food Sci. Technol. 2011, 29, 18–22. [Google Scholar] [CrossRef]
- Wang, C.T.; Hou, S.; Zhao, L.; Yang, X.L.; Wen, M.X.; Sun, B.G. In Application of coupling of reaction and separation technique (CRST) to biocatalytic processes of natural 3-methylthiopropanol. In Proceedings of the 2010 First International Conference on Cellular, Molecular Biology, Biophysics and Bioengineering, Qiqihar, China, 21–24 December 2010. [Google Scholar]
- Matthew, K.; Jingcan, S.; Shao, L. Optimization of L-methionine bioconversion to aroma-active methionol by Kluyveromyces lactis using the Taguchi method. J. Food Res. 2013, 2, 90. [Google Scholar] [CrossRef]
- Liu, L.; Yin, S.; Liang, J.R.; Wang, C.T.; Zhao, J.X. Cloning of the aminotransferase gene ARO8 and its influence on the production of methionol in Saccharomyces cerevisiae. Sci. Technol. Food Ind. 2014, 35, 132–135. [Google Scholar] [CrossRef]
- Deed, R.C.; Hou, R.Y.; Kinzurik, M.I.; Gardner, R.C.; Fedrizzi, B. The role of yeast ARO8, ARO9 and ARO1O genes in the biosynthesis of 3-(methylthio)-1-propanol from L-methionine during fermentation in synthetic grape medium. FEMS Yeast Res. 2019, 19, foy109. [Google Scholar] [CrossRef]
- Wen, M.X.; Wang, C.T.; Yang, X.L.; Zhao, L.; Liu, G.R.; Tong, J.M. Cloning and expression of ARO10 gene in Saccharomyces cerevisiae and it’s influence on 3-(methylthio)-1-propanol biosynthetic metabolism. Food Ferment. Ind. 2012, 38, 1–5. [Google Scholar] [CrossRef]
- Yin, S.; Lang, T.D.; Xiao, X.; Liu, L.; Sun, B.G.; Wang, C.T. Significant enhancement of methionol production by co-expression of the aminotransferase gene ARO8 and the decarboxylase gene ARO10 in Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2015, 362, fnu043. [Google Scholar] [CrossRef]
- Yang, X.L.; Zhang, K.L.; Li, J.Y.; Zhang, Q.C.; Luo, X.; Wang, C.T. Optimization of fermentation conditions for 3-methylthiopropanol production by recombinant Saccharomyces cerevisiae strain. J. Hebei Agric. Univ. 2014, 37, 94–101. [Google Scholar] [CrossRef]
- Du, R.B.; Liu, J.; Jiang, J.; Wang, Y.Q.; Ji, X.A.; Yang, N.; Wu, Q.; Xu, Y. Construction of a synthetic microbial community for the biosynthesis of volatile sulfur compound by multi-module division of labor. Food Chem. 2021, 347, 129036. [Google Scholar] [CrossRef]
- Frank, D.C.; Owen, C.M.; Patterson, J. Solid phase microextraction (SPME) combined with gas-chromatography and olfactometry-mass spectrometry for characterization of cheese aroma compounds. LWT Food Sci. Technol. 2004, 37, 139–154. [Google Scholar] [CrossRef]
- Yang, R.; Liu, P.X.; Chang, X.; Xu, J.Q.; Yin, H.; Fan, G.S.; Teng, C.; Li, X.T.; Gong, Y. Optimization of fermentation conditions for production of ethyl caproate in Baijiu using a selected isolate of Saccharomyces cerevisiae. Emir. J. Food Agric. 2021, 34, 59–69. [Google Scholar] [CrossRef]
- Wu, C.L.; Li, F.; Ma, C.; Yuan, Y.; Fan, G.S.; Li, X.T. Optimizing preparation conditions of cross-linked acid-stable xylanase aggregates. J. Food Sci. Technol. 2016, 34, 48–54. [Google Scholar]
- Fan, G.S.; Sun, B.G.; Xu, D.; Teng, C.; Fu, Z.L.; Du, Y.H.; Li, X.T. Isolation and identification of high-yield ethyl acetate-producing yeast from Gujinggong Daqu and its fermentation characteristics. J. Am. Soc. Brew. Chem. 2018, 76, 117–124. [Google Scholar] [CrossRef]
- Buchana, R.E.; Gibbons, N.E. Bergey’s Manual of Determinative Bacteriology, 8th ed.; Science Press: Beijing, China, 1984; pp. 382–533. [Google Scholar]
- Dong, X.Z.; Cai, M.Y. Ommon Bacteria Manual System Identification; Science Press: Beijing, China, 2001; pp. 348–392. [Google Scholar]
- Fan, G.S.; Liu, P.X.; Chang, X.; Yin, H.; Cheng, L.J.; Teng, C.; Gong, Y.; Li, X.T. Isolation and identification of a high-yield ethyl caproate-producing yeast from Daqu and optimization of its fermentation. Front. Microbiol. 2021, 12, 663744. [Google Scholar] [CrossRef]
- Wang, Z.N.; Yang, J.; Li, X.S.; Geng, J.Z.; Wu, D.P. Study on biological characteristics and physiological and biochemical characteristics of Saccharomyces cerevisiae. J. Shaanxi Univ. Technol. 2022, 38, 39–46. [Google Scholar] [CrossRef]
- Valachova, K.; Horvathova, V. Starch degradation by glucoamylase Glm from Saccharomycopsis fibuligera IFO 0111 in the presence and absence of a commercial pullulanase. Chem. Biodivers. 2007, 4, 874–880. [Google Scholar] [CrossRef]
- Fan, G.S.; Xu, D.; Fu, Z.L.; Xu, C.Y.; Teng, C.; Sun, B.G.; Li, X.T. Screen of aroma-producing yeast strains from Gujinggong Daqu and analysis of volatile flavor compounds produced by them. J. Chin. Inst. Food Sci. Technol. 2018, 18, 220–229. [Google Scholar] [CrossRef]
- Chi, Z.M.; Chi, Z.; Liu, G.L.; Wang, F.; Ju, L.; Zhang, T. Saccharomycopsis fibuligera and its applications in biotechnology. Biotechnol. Adv. 2009, 27, 423–431. [Google Scholar] [CrossRef]
- Xie, Z.B.; Zhang, K.Z.; Kang, Z.H.; Yang, J.G. Saccharomycopsis fibuligera in liquor production: A review. Eur. Food Res. Technol. 2021, 247, 1569–1577. [Google Scholar] [CrossRef]
- Wang, X.S.; Wang, B.W.; Sun, Z.G.; Tan, W.; Zheng, J.; Zhu, W.Y. Effects of modernized fermentation on the microbial community succession and ethyl lactate metabolism in Chinese baijiu fermentation. Food Res. Int. 2022, 159, 111566. [Google Scholar] [CrossRef]
- Yang, X.C.; Chen, Z.J.; Zhao, J.; Ding, P.Z.; Jia, M.F.; Gao, X.; Hu, J.X. Breeding of acid-resistant yeast strain by UV mutation and its characteristics. China Brew. 2017, 36, 109–113. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, C.Y.; Xiang, X.Q.; Xu, H.L.; Han, G.Q. Analysis of microbial diversity and succession during Xiaoqu Baijiu fermentation using high-throughput sequencing technology. Eng. Life Sci. 2022, 22, 495–504. [Google Scholar] [CrossRef]
- Fan, G.S.; Cheng, L.J.; Fu, Z.L.; Sun, B.G.; Teng, C.; Jiang, X.Y.; Li, X.T. Screening of yeasts isolated from Baijiu environments for 2-phenylethanol production and optimization of production conditions. 3 Biotech. 2020, 10, 275. [Google Scholar] [CrossRef]
- Hong, L.X.; Fu, G.M.; Liu, T.; Chen, Y.R.; Wu, S.W.; Cai, W.Q.; Xie, Z.J.; Wan, Y. Functional microbial agents enhance ethanol contents and regulate the volatile compounds in Chinese Baijiu. Food Biosci. 2021, 44, 101411. [Google Scholar] [CrossRef]
- Xu, Z.W.; Wang, M.Y.; Tan, S.Y.; Wang, Q.; Lu, G.Y.; Ma, J. Study on reducing nicotine content in tobacco industry waste. Tianjin Agric. Sci. 2022, 28, 81–86. [Google Scholar] [CrossRef]
- Landaud, S.; Helinck, S.; Bonnarme, P. Formation of volatile sulfur compounds and metabolism of methionine and other sulfur compounds in fermented food. Appl. Microbiol. Biotechnol. 2008, 77, 1191–1205. [Google Scholar] [CrossRef]
- Eshkol, N.; Sendovski, M.; Bahalul, M.; Katz-Ezov, T.; Kashi, Y.; Fishman, A. Production of 2-phenylethanol from L-phenylalanine by a stress tolerant Saccharomyces cerevisiae strain. J. Appl. Microbiol. 2009, 106, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Quek, J.; Seow, Y.X.; Ong, P.; Liu, S.Q. Formation of volatile sulfur-containing compounds by Saccharomyces cerevisiae in soymilk supplemented with L-methionine. Food Biotechnol. 2011, 25, 292–304. [Google Scholar] [CrossRef]
- Peng, A.A.; Liu, H.C.; Nie, Z.Y.; Xia, J.L. Effect of surfactant Tween-80 on sulfur oxidation and expression of sulfur metabolism relevant genes of Acidithiobacillus ferrooxidans. Trans. Nonferrous Met. Soc. China. 2012, 22, 3147–3155. [Google Scholar] [CrossRef]
- Liang, Y.; Zhu, L.; Gao, M.J.; Zheng, Z.Y.; Wu, J.R.; Zhan, X.B. Influence of Tween-80 on the production and structure of water-insoluble curdlan from Agrobacterium sp. Int. J. Biol. Macromol. 2018, 106, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Vinche, M.H.; Khanahmadi, M.; Ataei, S.A.; Danafar, F. Optimization of process variables for production of beta-glucanase by Aspergillus niger CCUG33991 in solid-state fermentation using wheat bran. Waste Biomass Valorization 2021, 12, 3233–3243. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.M.; van Maris, A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Mouret, J.R.; Perez, M.; Angenieux, M.; Nicolle, P.; Farines, V.; Sablayrolles, J.M. Online-based kinetic analysis of higher alcohol and ester synthesis during winemaking fermentations. Food Bioprocess Technol. 2014, 7, 1235–1245. [Google Scholar] [CrossRef]
- Tan, A.; Lee, P.R.; Seow, Y.X.; Ong, P.; Liu, S.Q. Volatile sulphur compounds and pathways of L-methionine catabolism in Williopsis yeasts. Appl. Microbiol. Biotechnol. 2012, 95, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Chursov, A.; Kopetzky, S.J.; Bocharov, G.; Frishman, D.; Shneider, A. RNAtips: Analysis of temperature-induced changes of RNA secondary structure. Nucleic Acids Res. 2013, 41, W486–W491. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.S.; Liu, P.X.; Wu, Q.H.; Fu, Z.L.; Cheng, L.J.; Zhu, Y.T.; Zhu, Y.P.; Yang, R.; Li, X.T. Optimization of cultural conditions for ethyl alcohol production by Saccharomyces cerevisiae YF1914 in aerobic conditions and its aroma-producing characteristics. Sci. Technol. Food Ind. 2019, 40, 52–58. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, C.; Liang, J.R.; Wang, C.T.; Zhao, J.X. Effects of knockout of gene CYS3 on metionol anabolism in Saccharomy cescerevisiae. Food Sci. 2014, 35, 139–143. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, H.; Xi, J.; Jin, Y.; Chen, Y.; Guo, L.; Jin, Z.; Xu, X. Improving bread aroma using low-temperature sourdough fermentation. Food Biosci. 2020, 37, 100704. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Li, J.Z.; Xu, Y. New development direction of Baijiu enterprises-rejucenation of Baijiu consumption. China Brew. 2021, 40, 227–230. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Factors | Y: 3-Met Concentration (g/L) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| A: Glucose Concentration (g/L) | B: Yeast Extract Concentration (g/L) | C: L-Methionine Concentration (g/L) | D: Temperature (°C) | E: Initial pH | F: Fermentation Time (h) | G: Inoculum Size (%) | H: Liquid Volume (mL/250 mL) | I: Shaking Speed (rpm) | ||
| 1 | 50 (+1) | 0.4 (−1) | 6 (+1) | 36 (+1) | 4.5 (−1) | 60 (+1) | 0.4 (−1) | 12.5 (−1) | 180 (−1) | 2.34 |
| 2 | 30 (−1) | 0.4 | 2 (−1) | 28 (−1) | 4.5 | 48 (−1) | 0.4 | 12.5 | 180 | 0.50 |
| 3 | 40 (0) | 0.6 (0) | 4 (0) | 32 (0) | 5 (0) | 54 (0) | 0.8 (0) | 25 (0) | 225 (0) | 3.65 |
| 4 | 30 | 0.4 | 6 | 36 | 5.5 (+1) | 48 | 1.2 (+1) | 37.5 (+1) | 180 | 2.45 |
| 5 | 50 | 0.4 | 2 | 28 | 5.5 | 60 | 1.2 | 12.5 | 270 (+1) | 2.74 |
| 6 | 30 | 0.4 | 2 | 36 | 5.5 | 60 | 0.4 | 37.5 | 270 | 2.75 |
| 7 | 40 | 0.6 | 4 | 32 | 5 | 54 | 0.8 | 25 | 225 | 3.81 |
| 8 | 50 | 0.4 | 6 | 28 | 4.5 | 48 | 1.2 | 37.5 | 270 | 2.13 |
| 9 | 50 | 0.4 | 2 | 36 | 4.5 | 48 | 0.4 | 37.5 | 270 | 1.90 |
| 10 | 40 | 0.4 | 4 | 32 | 5 | 54 | 0.8 | 25 | 225 | 3.83 |
| 11 | 50 | 0.8 (+1) | 6 | 28 | 5.5 | 60 | 0.4 | 37.5 | 180 | 2.73 |
| 12 | 30 | 0.8 | 6 | 28 | 5.5 | 48 | 0.4 | 12.5 | 270 | 2.89 |
| 13 | 30 | 0.8 | 2 | 28 | 4.5 | 60 | 1.2 | 37.5 | 180 | 2.32 |
| 14 | 50 | 0.8 | 2 | 36 | 5.5 | 48 | 1.2 | 12.5 | 180 | 2.96 |
| 15 | 30 | 0.8 | 6 | 36 | 4.5 | 60 | 1.2 | 12.5 (−1) | 270 | 3.54 |
| Effect (Exi) | 0.00291 | 0.712 | 0.121 | 0.0548 | 0.631 | 0.02493 | 0.632 | −0.00459 | 0.00494 | |
| F values | 0.19 | 18.54 | 13.38 | 10.96 | 22.7 | 20.43 | 14.59 | 0.75 | 11.29 | |
| p values | 0.683 | 0.013 | 0.022 | 0.03 | 0.009 | 0.011 | 0.019 | 0.435 | 0.028 | |
| Rank | 9 | 3 | 5 | 7 | 1 | 2 | 4 | 8 | 6 | |
| Significance | - | * | * | * | ** | * | * | - | * | |
| Groups | Initial pH | Fermentation Time (h) | Yeast Extract Concentration (g/L) | Inoculum Size (%) | L-Methionine Concentration (g/L) | Shaking Speed (rpm) | Temperature (°C) | 3-Met Concentration (g/L) |
|---|---|---|---|---|---|---|---|---|
| 1 | 4.5 | 36 | 0.2 | 0.4 | 3 | 180 | 28 | 2.28 |
| 2 | 4.75 | 44 | 0.4 | 0.6 | 4 | 195 | 30 | 3.13 |
| 3 | 5 | 52 | 0.6 | 0.8 | 5 | 210 | 32 | 3.73 |
| 4 | 5.25 | 60 | 0.8 | 1.0 | 6 | 225 | 34 | 3.62 |
| 5 | 5.5 | 68 | 1.0 | 1.2 | 7 | 240 | 36 | 2.66 |
| NO. | Fermentation Time (h) | Yeast Extract Concentration (g/L) | Initial pH | 3-Met Concentration (g/L) | |||
|---|---|---|---|---|---|---|---|
| A | Code A | B | Code B | C | Code C | Y | |
| 1 | 60 | 1 | 1.0 | 1 | 5.8 | 1 | 1.83 |
| 2 | 44 | −1 | 1.0 | 1 | 4.2 | −1 | 2.51 |
| 3 | 60 | 1 | 0.6 | 0 | 4.2 | −1 | 3.15 |
| 4 | 44 | −1 | 0.6 | 0 | 5 | 0 | 2.61 |
| 5 | 60 | 1 | 1.0 | 1 | 5 | 0 | 2.09 |
| 6 | 52 | 0 | 0.2 | −1 | 4.2 | −1 | 1.66 |
| 7 | 44 | −1 | 0.6 | 0 | 5.8 | 1 | 2.41 |
| 8 | 52 | 0 | 0.6 | 0 | 5 | 0 | 3.83 |
| 9 | 52 | 0 | 0.2 | −1 | 5.8 | 1 | 2.53 |
| 10 | 60 | 1 | 0.2 | −1 | 5 | 0 | 2.89 |
| 11 | 52 | 0 | 1.0 | 1 | 5.8 | 1 | 3.19 |
| 12 | 52 | 0 | 0.6 | 0 | 5 | 0 | 3.91 |
| 13 | 52 | 0 | 0.6 | 0 | 5 | 0 | 4.09 |
| 14 | 52 | 0 | 1.0 | 1 | 4.2 | −1 | 2.83 |
| 15 | 44 | −1 | 1.0 | 1 | 5 | 1 | 2.83 |
| Strain Number | 3-Met Production (mg/L) | Strain Number | 3-Met Production (mg/L) | Strain Number | 3-Met Production (mg/L) | Strain Number | 3-Met Production (mg/L) |
|---|---|---|---|---|---|---|---|
| B1 | 0 ± 0 | B25 | 0 ± 0 | B49 | 24.57 ± 1.03 | Y12 | 217.73 ± 3.26 |
| B2 | 0 ± 0 | B26 | 0 ± 0 | B50 | 0 ± 0 | Y13 | 0 ± 0 |
| B3 | 0 ± 0 | B27 | 0 ± 0 | B51 | 0 ± 0 | Y14 | 895.29 ± 5.09 |
| B4 | 2.94 ± 0.22 | B28 | 0 ± 0 | B52 | 0 ± 0 | Y1402 | 2001 ± 10.37 |
| B5 | 0 ± 0 | B29 | 45.78 ± 2.12 | B53 | 0 ± 0 | Y15 | 0 ± 0 |
| B6 | 0 ± 0 | B30 | 0 ± 0 | B54 | 0 ± 0 | Y16 | 0 ± 0 |
| B7 | 53.85 ± 4.01 | B31 | 0 ± 0 | B55 | 0 ± 0 | Y17 | 1131 ± 9.27 |
| B8 | 5.52 ± 1.38 | B32 | 0 ± 0 | B56 | 0 ± 0 | M1 | 39.88 ± 2.03 |
| B9 | 30.57 ± 4.91 | B33 | 6.33 ± 0.24 | B57 | 34.65 ± 6.36 | M2 | 0 ± 0 |
| B10 | 0 ± 0 | B34 | 0 ± 0 | B58 | 0 ± 0 | M3 | 0 ± 0 |
| B11 | 0 ± 0 | B35 | 0 ± 0 | B59 | 0 ± 0 | M4 | 0 ± 0 |
| B12 | 0 ± 0 | B36 | 0 ± 0 | B60 | 52.49 ± 7.18 | M5 | 0 ± 0 |
| B13 | 0 ± 0 | B37 | 0 ± 0 | B61 | 39.09 ± 0.95 | M6 | 0 ± 0 |
| B14 | 0 ± 0 | B38 | 42.61 ± 2.00 | Y1 | 0 ± 0 | M7 | 0 ± 0 |
| B15 | 0 ± 0 | B39 | 0 ± 0 | Y2 | 1411.63 ± 17.39 | M8 | 0 ± 0 |
| B16 | 0 ± 0 | B40 | 0 ± 0 | Y3 | 1464.34 ± 8.46 | M9 | 0 ± 0 |
| B17 | 0 ± 0 | B41 | 41.20 ± 1.73 | Y4 | 1021.86 ± 5.19 | M10 | 43.63 ± 2.28 |
| B18 | 0 ± 0 | B42 | 0 ± 0 | Y5 | 1208.04 ± 17.30 | M11 | 54.58 ± 3.03 |
| B19 | 0 ± 0 | B43 | 0 ± 0 | Y6 | 1690.45 ± 14.22 | M12 | 30.39 ± 8.26 |
| B20 | 0 ± 0 | B44 | 0 ± 0 | Y7 | 0 ± 0 | M13 | 0 ± 0 |
| B21 | 52.73 ± 6.02 | B45 | 0 ± 0 | Y8 | 115.09 ± 2.97 | M14 | 34.83 ± 6.17 |
| B22 | 14.93 ± 3.17 | B46 | 0.73 ± 0.16 | Y9 | 559.45 ± 7.13 | M15 | 52.69 ± 2.04 |
| B23 | 49.32 ± 0.74 | B47 | 49.16 ± 3.36 | Y10 | 1488.61 ± 12.05 | ||
| B24 | 0 ± 0 | B48 | 0 ± 0 | Y11 | 0 ± 0 |
| Source | Sum of Squares | DF | Mean Square | F Values | p Values | Significance |
|---|---|---|---|---|---|---|
| Model | 7.22573 | 9 | 0.80286 | 49.06 | 0.000 | *** |
| A—Fermentation time | 1.23168 | 1 | 1.23168 | 75.26 | 0.000 | *** |
| B—Yeast extract concentration | 1.27371 | 1 | 1.27371 | 77.83 | 0.000 | *** |
| C—Initial pH | 0.37036 | 1 | 0.37036 | 22.63 | 0.005 | ** |
| A2 | 0.22626 | 1 | 0.22626 | 13.83 | 0.014 | * |
| B2 | 2.41522 | 1 | 2.41522 | 147.58 | 0.000 | *** |
| C2 | 2.21996 | 1 | 2.21996 | 135.65 | 0.000 | *** |
| AB | 0.00101 | 1 | 0.00101 | 0.06 | 0.814 | - |
| AC | 0.10892 | 1 | 0.10892 | 6.66 | 0.049 | * |
| BC | 1.92710 | 1 | 1.92710 | 117.75 | 0.000 | *** |
| Residual | 0.08183 | 5 | 0.01637 | |||
| Lack of Fit | 0.04553 | 3 | 0.01518 | 0.84 | 0.585 | - |
| Pure Error | 0.03630 | 2 | 0.01815 | |||
| Cor Total | 7.30756 | 14 | ||||
| R2 = 0.9891 | R2Adj = 0.9694 |
| Strain Classify | Strain | Culture Institution | L-Met Concentration (g/L) | 3-Met Production (mg/L) | Reference |
|---|---|---|---|---|---|
| Natural strain | S. cerevisiae S288C | Beijing Technology and Business University, Beijing 100048, China | 10 | 590 | [14] |
| S. cerevisiae SC408 | Beijing Technology and Business University, Beijing 100048, China | 4 | 1600 | [11] | |
| H. burtonii YHM-G | Beijing Technology and Business University, Beijing 100048, China | 6 | 3160 | [7] | |
| K. lactis KL71 | National University of Singapore, Singapore 117543 | - | 990 | [13] | |
| S. cerevisiae SC57 | Beijing Technology and Business University (BTBU), Beijing 100048, China | 4 | 1600 (fed-batch fermentation without D101); 2260 (fed-batch fermentation with D101) | [12] | |
| S. fibuligera Y1402 | Beijing Technology and Business University (BTBU), Beijing 100048, China | 5 | 4030 | This study | |
| Engineered strain | S. cerevisiae S288C-CYS3 | Beijing Technology and Business University, Beijing 100048, China | 4 | 690 | [6] |
| S. cerevisiae s288c-ARO10 | Beijing Technology and Business University, Beijing 100048, China | 10 | 900 | [16] | |
| S. cerevisiae C3 | Beijing Technology and Business University, Beijing 100048, China | 10 | 600 | [49] | |
| S. cerevisiae S288C | Beijing Technology and Business University, Beijing 100048, China | 10 | 940 | [49] | |
| S. cerevisiae AR8 | Beijing Technology and Business University, Beijing 100048, China | 10 | 760 | [14] | |
| S. cerevisiae S288C-AR010 | Beijing Technology and Business University, Beijing 100048, China | 5 | 4380 | [18] | |
| S. cerevisiae CEN.PK113-7D | Biochemical Engineering Group, Theodor-Heuss-Allee 25, 60486 Frankfurt am Main, Germany | 2200 | [5] | ||
| S. cerevisiae (co-expression of ARO8 and ARO100 | School of Food and Chemical Engineering, Beijing Technology and Business University, Beijing 100048, China | 10 | 3240 | [17] |
| Volatile Compounds | SHM a | Y1402 b | Aroma Descriptors |
|---|---|---|---|
| (Z)-3,7-Dimethyl-3,6-octadien-1-ol | - c | 280 ± 35 | Floral |
| 2-Methyl-1-propanol | - | 33 ± 5 | Sweet, wine |
| 3-Methyl-3-buten-1-ol | - | 5 ± 1 | Slightly apple-like |
| (R)-2-Octanol | - | 300 ± 23 | Creamy, cucumber |
| 3-Methyl-1-butanol | - | 444 ± 51 | Floral |
| 1-Octen-3-ol | - | 4 ± 2 | Mushroom |
| 2-Propyl-1-pentanol | - | 3 ± 1 | Mushroom, cream |
| Phenylethyl alcohol | 19 ± 3 | 304 ± 57 | Rosy, honey |
| ∑Alcohols | 19 | 1373 | |
| Dibutyl phthalate | - | 30 ± 11 | Odorless |
| (E)-9-Octadecenoic acid ethyl ester | 3 ± 1 | 5 ± 1 | Lipid |
| Dimethyl phthalate | 4 ± 1 | 2 ± 0 | Odorless |
| Hexadecanoic acid ethyl ester | 6 ± 0 | - | Cream, herb |
| Ethyl linoleate | 2 ± 0 | - | Lipid |
| ∑Esters | 15 | 37 | |
| 2,4-Dimethyl-benzaldehyde | 14 ± 2 | 85 ± 13 | Semen armeniacae amarae |
| Furfural | - | 3 ± 0 | Roasted, almond |
| ∑Aldehydes | 14 | 88 | |
| 4-Ethyl-phenol | - | 15 ± 6 | Horsy, barnyard, smoky, medical aromatic |
| 2-Methoxy-phenol | - | 13 ± 3 | Castoreum, smoke, bacon, ham |
| 4-Ethyl-2-methoxy-phenol | - | 110 ± 33 | Spicy, smoky, bacon, clove |
| Phenol | - | 6 ± 2 | Rubber |
| 2-Methoxy-4-vinylphenol | - | 920 ± 49 | Fruity, clove |
| 2,4-Di-tert-butylphenol | - | 10 ± 1 | Phenolic |
| ∑Phenols | - | 1074 | |
| 3-Methyl-butanoic acid | - | 8 ± 2 | Sour, cheesy |
| 2-Methyl-propanoic acid | - | 5 ± 1 | Cheese, dairy, buttery |
| ∑Acids | - | 13 | |
| 2,3-Dihydro-benzofuran | - | 356 ± 52 | Roast, sweet |
| Dihydro-5-pentyl-2(3H)-furanone | - | 11 ± 2 | Coconut, creamy, sweet, buttery, oily |
| 5-Hexyldihydro-2(3H)-furanone | - | 11 ± 4 | Fruity, creamy, peach, coconut, buttery, sweet |
| ∑Furanone | - | 378 | |
| 1,2-Benzisothiazole | - | 2 ± 0 | N d |
| Benzothiazole | 83 ± 6 | - | Meaty, vegetative |
| ∑Azoles | 83 | 2 | |
| 7,9-Di-tert-butyl-1-oxaspiro(4,5)deca-6,9-diene-2,8-dione | - | 6 ± 0 | N |
| Pentadecane | - | 2 ± 0 | Waxy |
| (3Z,5E)-1,3,5-Undecatriene | - | 7 ± 3 | Oily |
| Aniline | 5 ± 0 | - | Sweet |
| Methyl-pyrazine | 2 ± 0 | - | Roast |
| ∑Others | 7 | 15 | |
| Sum | 138 | 2980 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Sun, Q.; Liu, X.; Basit, R.A.; Ma, J.; Fu, Z.; Cheng, L.; Fan, G.; Teng, C. Screening, Identification, and Fermentation Condition Optimization of a High-Yield 3-Methylthiopropanol Yeast and Its Aroma-Producing Characteristics. Foods 2024, 13, 418. https://doi.org/10.3390/foods13030418
Zhang Y, Sun Q, Liu X, Basit RA, Ma J, Fu Z, Cheng L, Fan G, Teng C. Screening, Identification, and Fermentation Condition Optimization of a High-Yield 3-Methylthiopropanol Yeast and Its Aroma-Producing Characteristics. Foods. 2024; 13(3):418. https://doi.org/10.3390/foods13030418
Chicago/Turabian StyleZhang, Yujiao, Qi Sun, Xiaoyan Liu, Rana Abdul Basit, Jinghao Ma, Zhilei Fu, Liujie Cheng, Guangsen Fan, and Chao Teng. 2024. "Screening, Identification, and Fermentation Condition Optimization of a High-Yield 3-Methylthiopropanol Yeast and Its Aroma-Producing Characteristics" Foods 13, no. 3: 418. https://doi.org/10.3390/foods13030418
APA StyleZhang, Y., Sun, Q., Liu, X., Basit, R. A., Ma, J., Fu, Z., Cheng, L., Fan, G., & Teng, C. (2024). Screening, Identification, and Fermentation Condition Optimization of a High-Yield 3-Methylthiopropanol Yeast and Its Aroma-Producing Characteristics. Foods, 13(3), 418. https://doi.org/10.3390/foods13030418