Influence of Microbial Transglutaminase on the Formation of Physico-Chemical Properties of Meat Analogs

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
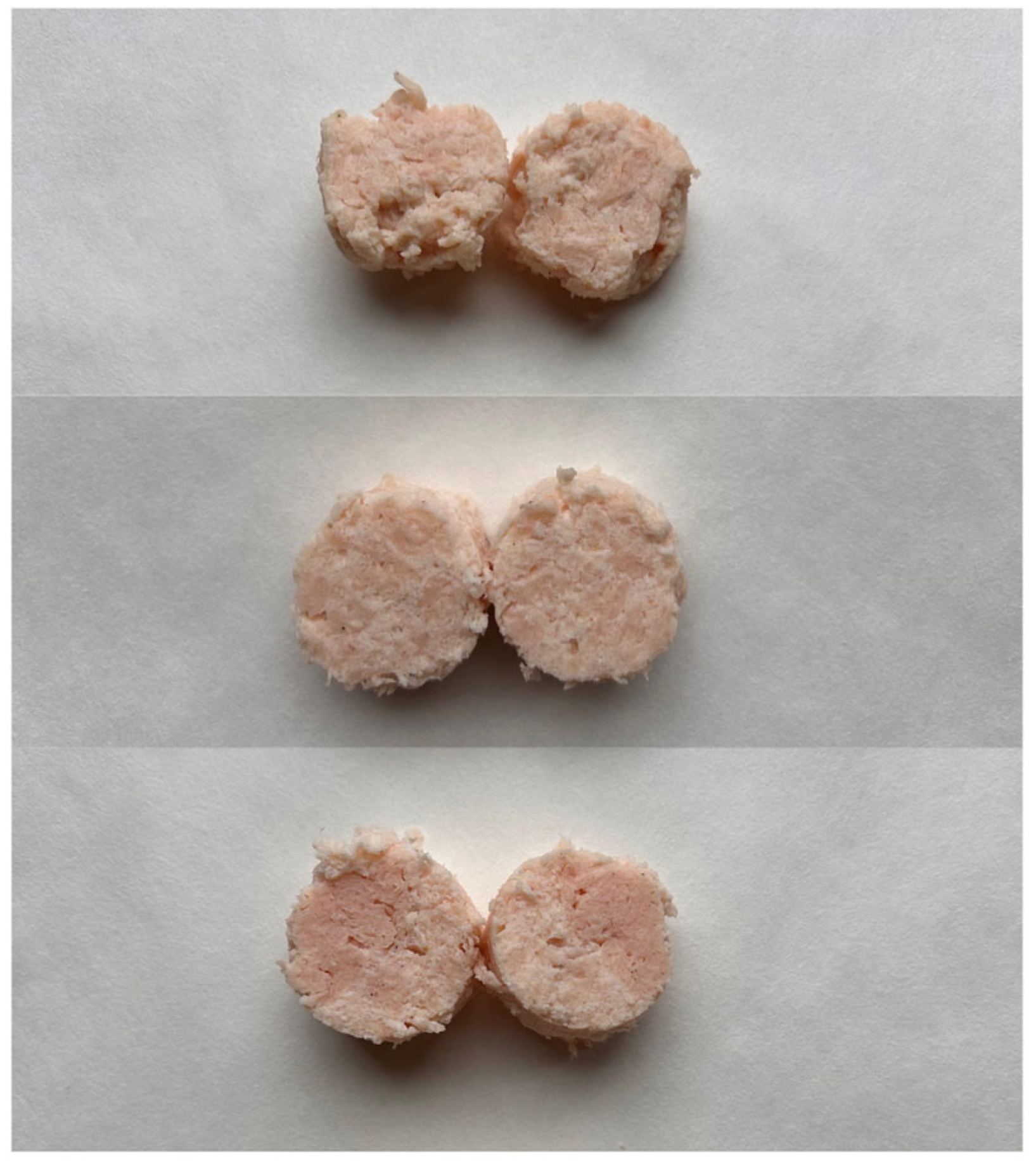
2.2.1. Samples Preparation
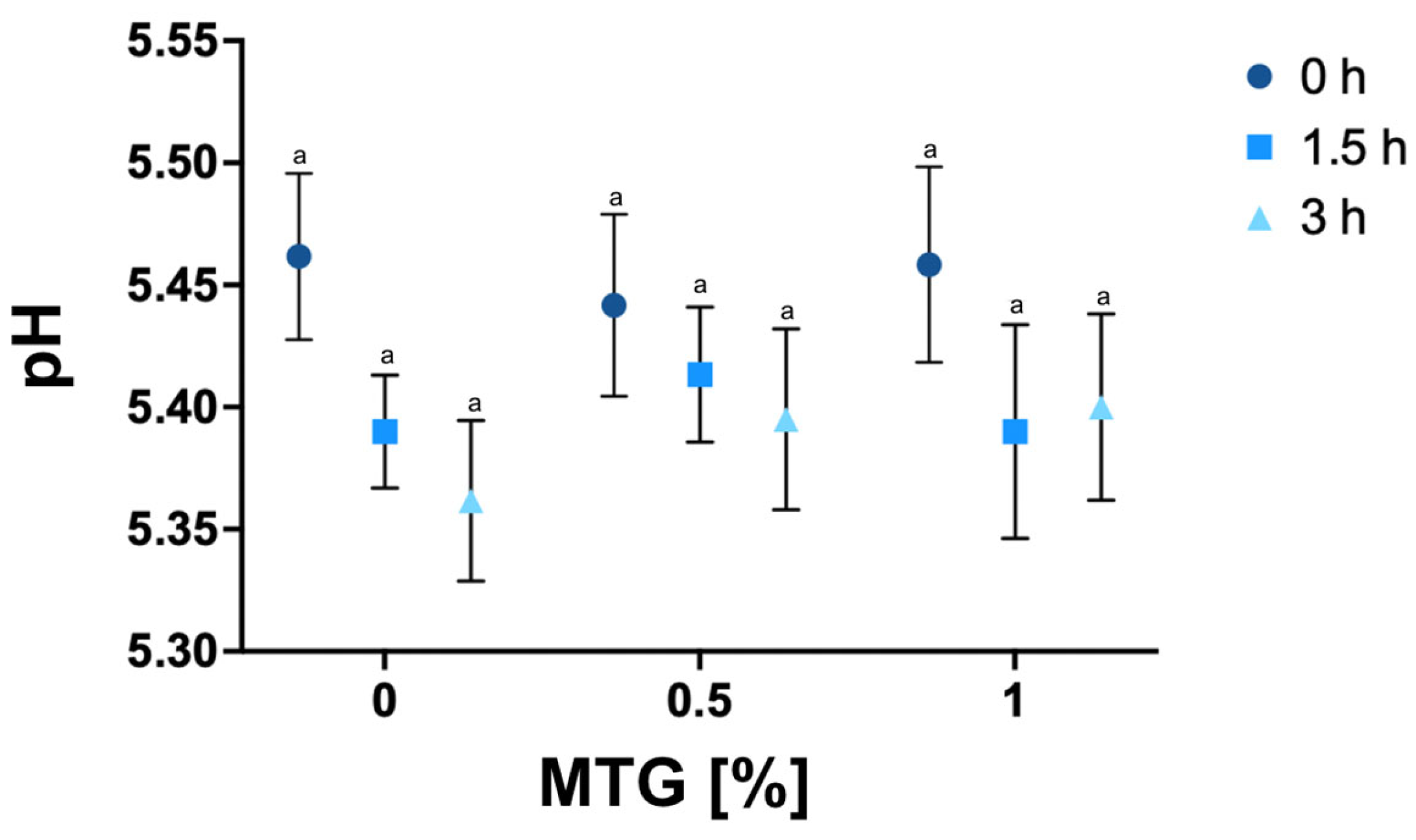
2.2.2. pH Determination
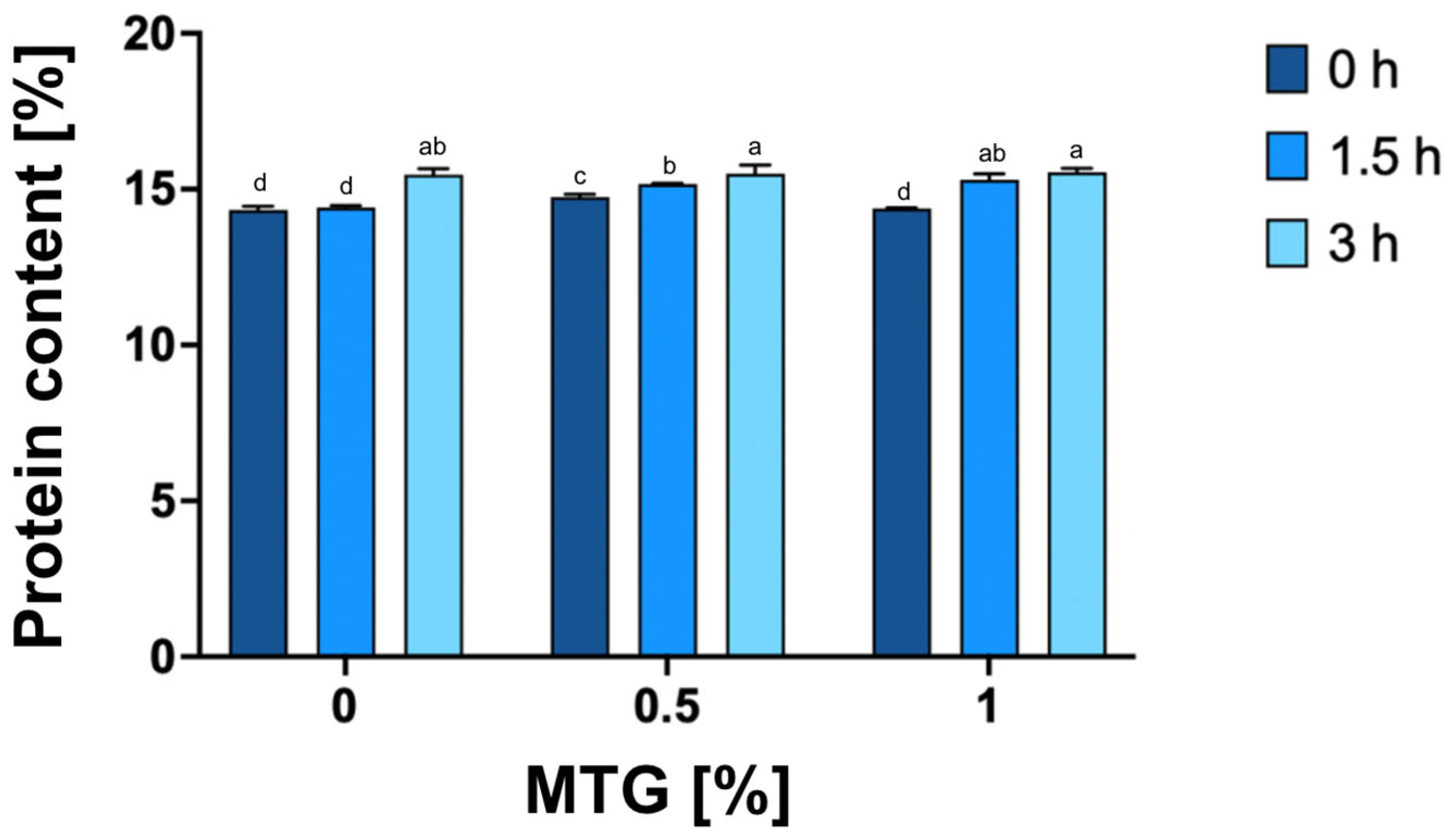
2.2.3. Protein Content Determination
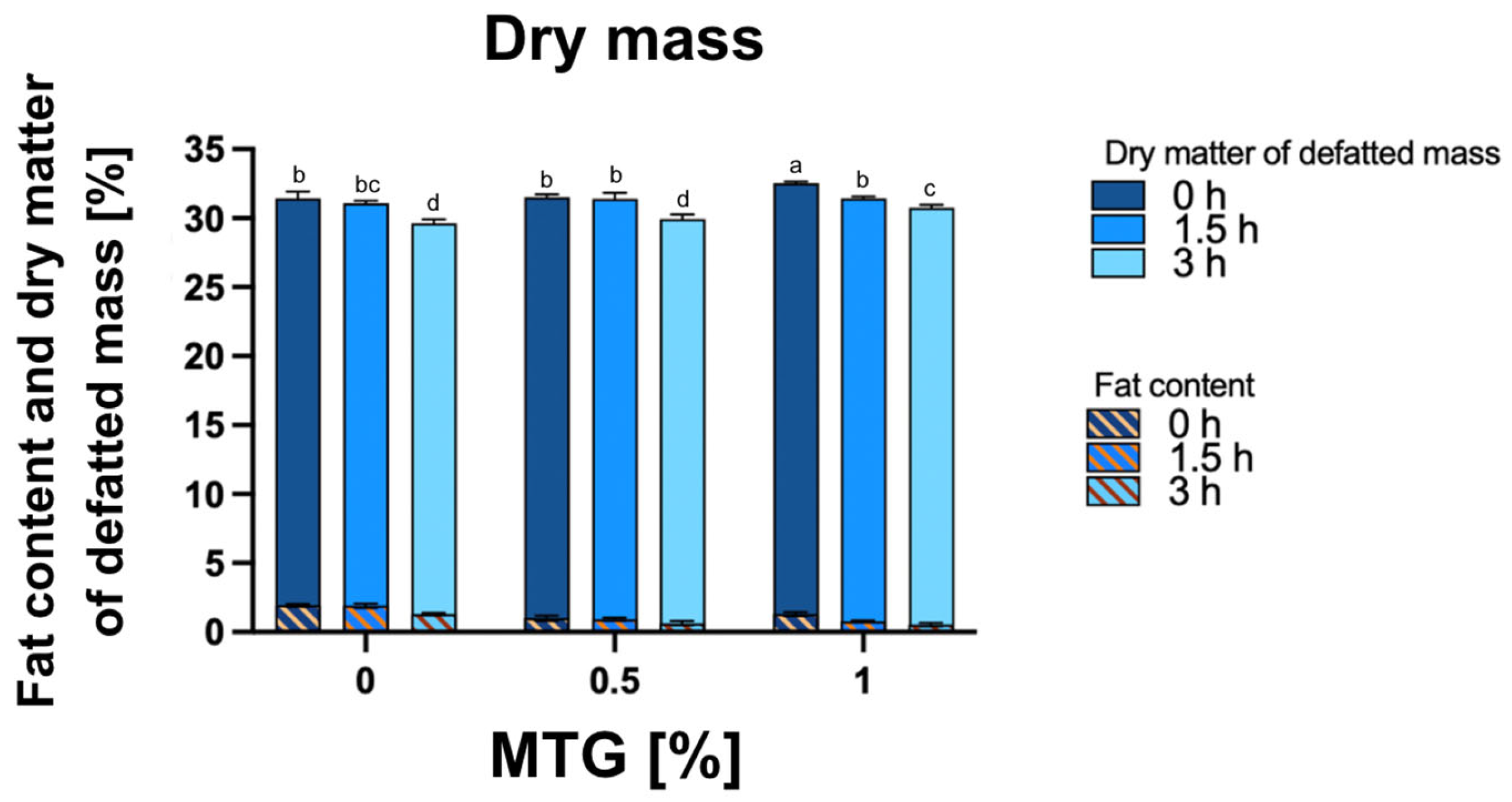
2.2.4. Determination of Dry Matter
2.2.5. Determination of Fat Content and Dry Matter of Defatted Mass
2.2.6. Identification of the Total Water and Fat Leakage, the Water and the Fat Content in the Total Leakage
2.2.7. Thermal Loss Determination
2.2.8. Determination of Selected Textural Parameters
2.3. Statistical Analysis
3. Results and Discussion
3.1. pH
3.2. Protein Content
3.3. Dry Matter, Fat Content, and Dry Matter of Defatted Mass
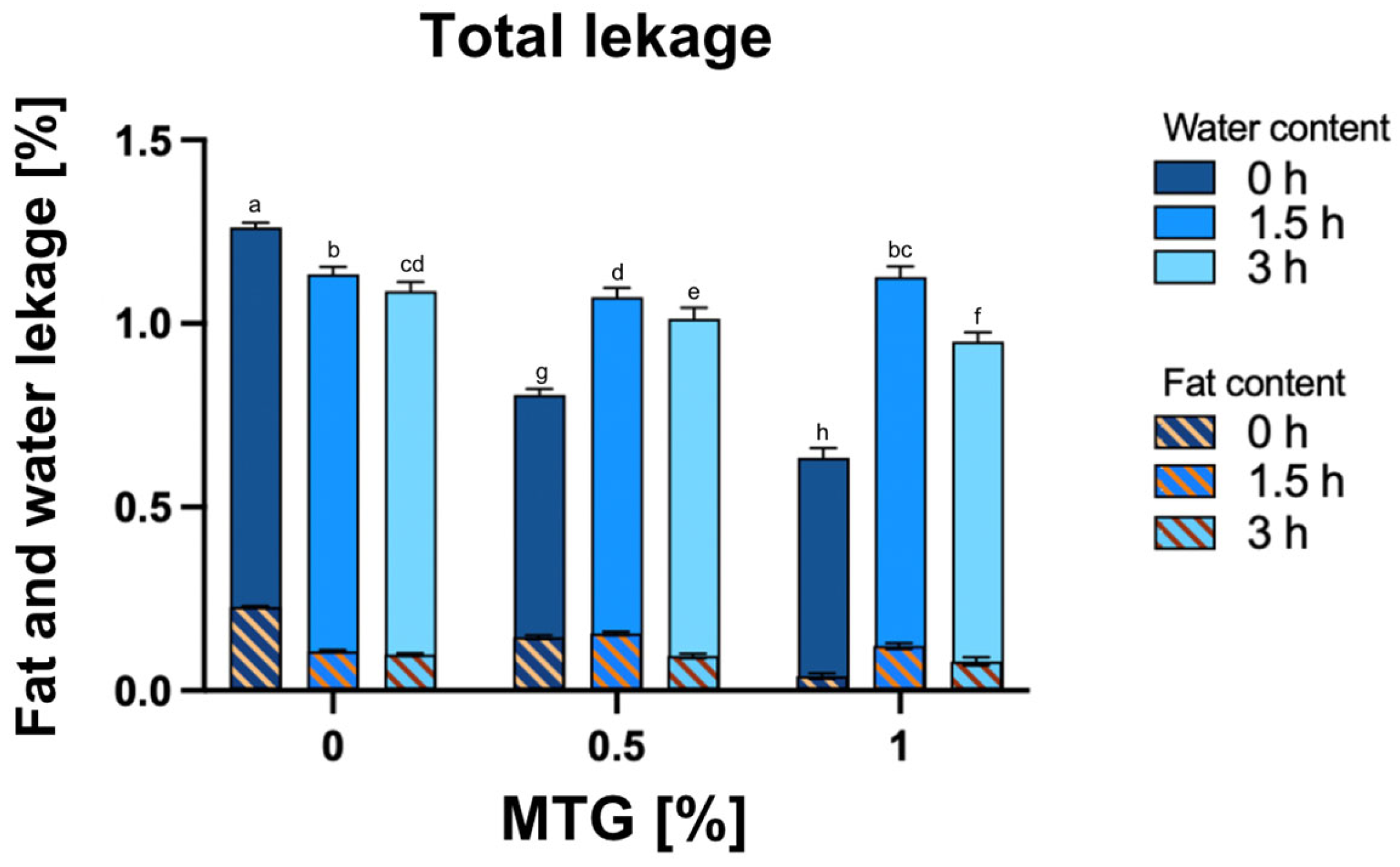
3.4. Total Water and Fat Leakage, the Water and the Fat Content in the Total Leakage
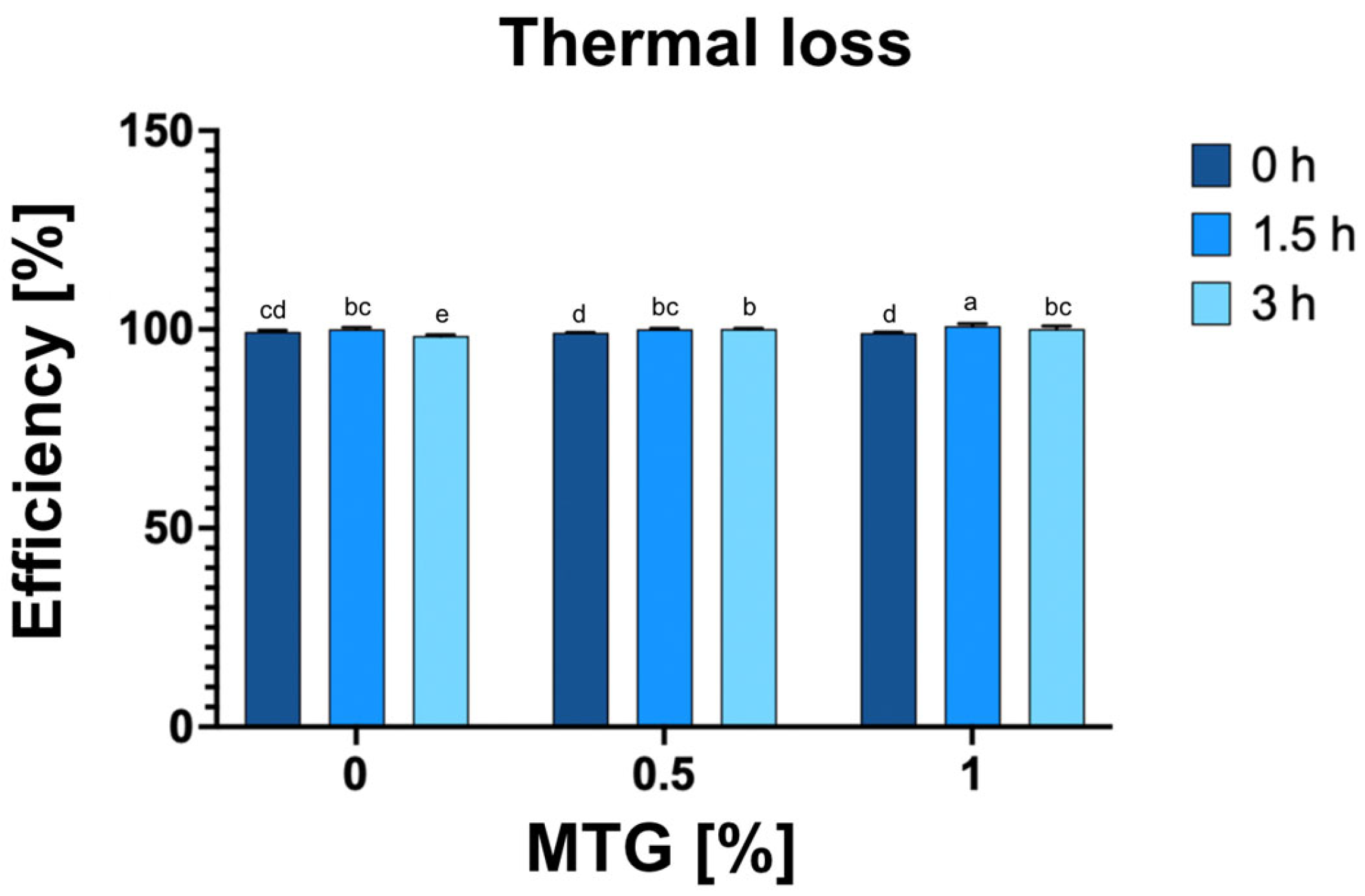
3.5. Thermal Loss
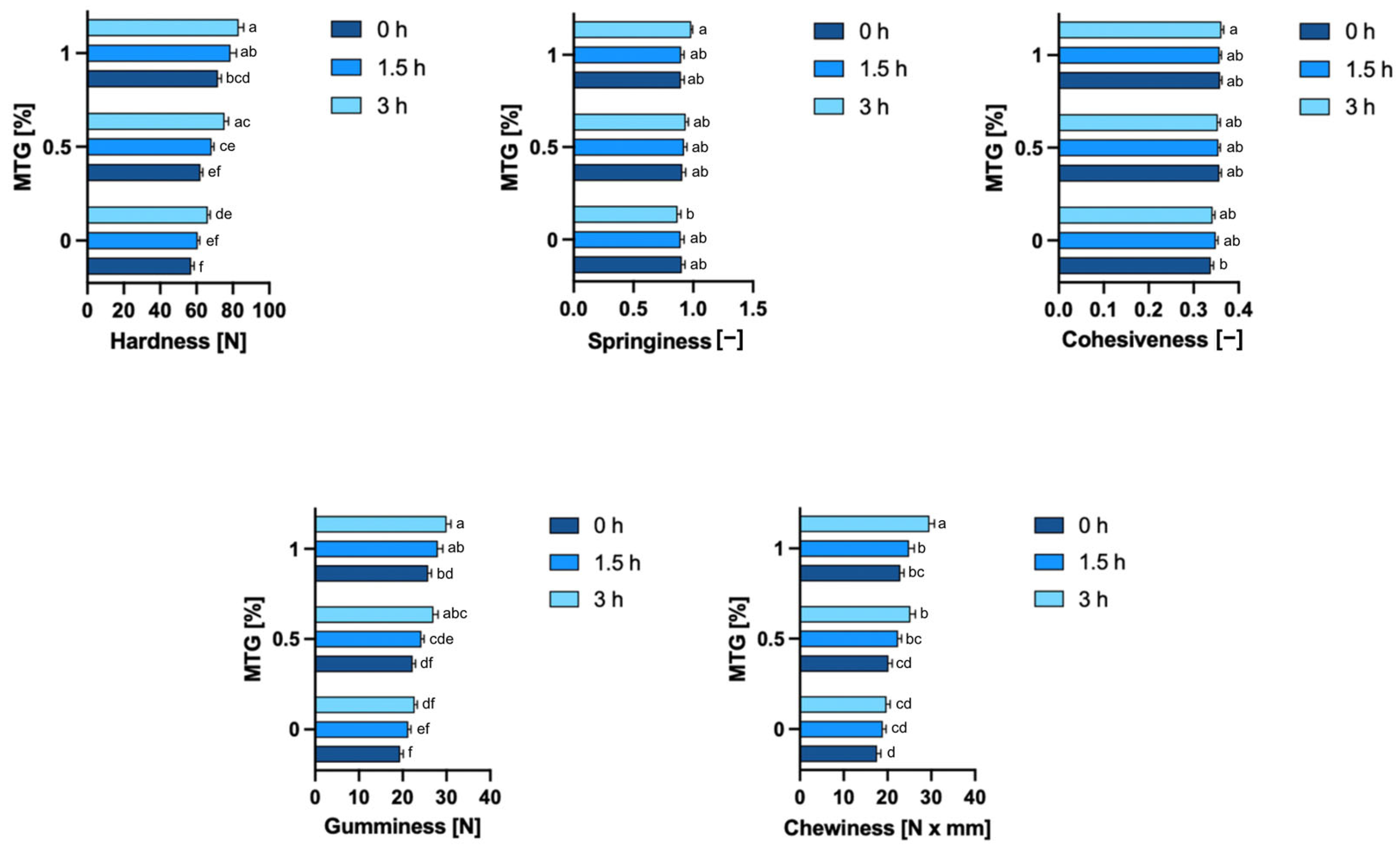
3.6. Textural Properties of Samples
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lima, M.; Costa, R.; Rodrigues, I.; Lameiras, J.; Botelho, G. A Narrative Review of Alternative Protein Sources: Highlights on Meat, Fish, Egg and Dairy Analogues. Foods 2022, 11, 2053. [Google Scholar] [CrossRef] [PubMed]
- Geiker, N.R.W.; Bertram, H.C.; Mejborn, H.; Dragsted, L.O.; Kristensen, L.; Carrascal, J.R.; Bügel, S.; Astrup, A. Meat and Human Health—Current Knowledge and Research Gaps. Foods 2021, 10, 1556. [Google Scholar] [CrossRef] [PubMed]
- FAO. Food Outlook–Biannual Report on Global Food Markets; FAO: Rome, Italy, 2023; ISBN 978-92-5-137230-2. [Google Scholar]
- Sans, P.; Combris, P. World Meat Consumption Patterns: An Overview of the Last Fifty Years (1961–2011). Meat Sci. 2015, 109, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Steenson, S.; Buttriss, J.L. Healthier and More Sustainable Diets: What Changes Are Needed in High-income Countries? Nutr. Bull. 2021, 46, 279–309. [Google Scholar] [CrossRef]
- Stoll-Kleemann, S.; Schmidt, U.J. Reducing Meat Consumption in Developed and Transition Countries to Counter Climate Change and Biodiversity Loss: A Review of Influence Factors. Reg. Environ. Chang. 2017, 17, 1261–1277. [Google Scholar] [CrossRef]
- Giromini, C.; Givens, D.I. Benefits and Risks Associated with Meat Consumption during Key Life Processes and in Relation to the Risk of Chronic Diseases. Foods 2022, 11, 2063. [Google Scholar] [CrossRef]
- Kennedy, J.; Alexander, P.; Taillie, L.S.; Jaacks, L.M. Estimated Effects of Reductions in Processed Meat Consumption and Unprocessed Red Meat Consumption on Occurrences of Type 2 Diabetes, Cardiovascular Disease, Colorectal Cancer, and Mortality in the USA: A Microsimulation Study. Lancet Planet. Health 2024, 8, e441–e451. [Google Scholar] [CrossRef]
- González, N.; Marquès, M.; Nadal, M.; Domingo, J.L. Meat Consumption: Which Are the Current Global Risks? A Review of Recent (2010–2020) Evidences. Food Res. Int. 2020, 137, 109341. [Google Scholar] [CrossRef]
- Farvid, M.S.; Sidahmed, E.; Spence, N.D.; Mante Angua, K.; Rosner, B.A.; Barnett, J.B. Consumption of Red Meat and Processed Meat and Cancer Incidence: A Systematic Review and Meta-Analysis of Prospective Studies. Eur. J. Epidemiol. 2021, 36, 937–951. [Google Scholar] [CrossRef]
- Rust, N.A.; Ridding, L.; Ward, C.; Clark, B.; Kehoe, L.; Dora, M.; Whittingham, M.J.; McGowan, P.; Chaudhary, A.; Reynolds, C.J.; et al. How to Transition to Reduced-Meat Diets That Benefit People and the Planet. Sci. Total Environ. 2020, 718, 137208. [Google Scholar] [CrossRef]
- Turnes, A.; Pereira, P.; Cid, H.; Valente, A. Meat Consumption and Availability for Its Reduction by Health and Environmental Concerns: A Pilot Study. Nutrients 2023, 15, 3080. [Google Scholar] [CrossRef] [PubMed]
- Viroli, G.; Kalmpourtzidou, A.; Cena, H. Exploring Benefits and Barriers of Plant-Based Diets: Health, Environmental Impact, Food Accessibility and Acceptability. Nutrients 2023, 15, 4723. [Google Scholar] [CrossRef] [PubMed]
- Alcorta, A.; Porta, A.; Tárrega, A.; Alvarez, M.D.; Vaquero, M.P. Foods for Plant-Based Diets: Challenges and Innovations. Foods 2021, 10, 293. [Google Scholar] [CrossRef] [PubMed]
- Ford, H.; Zhang, Y.; Gould, J.; Danner, L.; Bastian, S.E.P.; Yang, Q. Comparing Motivations and Barriers to Reduce Meat and Adopt Protein Alternatives amongst Meat-Eaters in Australia, China and the UK. Food Qual. Prefer. 2024, 118, 105208. [Google Scholar] [CrossRef]
- Fiorentini, M.; Kinchla, A.J.; Nolden, A.A. Role of Sensory Evaluation in Consumer Acceptance of Plant-Based Meat Analogs and Meat Extenders: A Scoping Review. Foods 2020, 9, 1334. [Google Scholar] [CrossRef]
- Ishaq, A.; Irfan, S.; Sameen, A.; Khalid, N. Plant-Based Meat Analogs: A Review with Reference to Formulation and Gastrointestinal Fate. Curr. Res. Food Sci. 2022, 5, 973–983. [Google Scholar] [CrossRef]
- Mazumder, M.A.R.; Panpipat, W.; Chaijan, M.; Shetty, K.; Rawdkuen, S. Role of Plant Protein on the Quality and Structure of Meat Analogs: A New Perspective for Vegetarian Foods. Future Foods 2023, 8, 100280. [Google Scholar] [CrossRef]
- Guyony, V.; Fayolle, F.; Jury, V. High Moisture Extrusion of Vegetable Proteins for Making Fibrous Meat Analogs: A Review. Food Rev. Int. 2023, 39, 4262–4287. [Google Scholar] [CrossRef]
- Dinali, M.; Liyanage, R.; Silva, M.; Newman, L.; Adhikari, B.; Wijesekara, I.; Chandrapala, J. Fibrous Structure in Plant-Based Meat: High-Moisture Extrusion Factors and Sensory Attributes in Production and Storage. Food Rev. Int. 2024, 40, 2940–2968. [Google Scholar] [CrossRef]
- Jang, J.; Lee, D.-W. Advancements in Plant Based Meat Analogs Enhancing Sensory and Nutritional Attributes. NPJ Sci. Food 2024, 8, 50. [Google Scholar] [CrossRef]
- Calton, A.; Lille, M.; Sozer, N. 3-D Printed Meat Alternatives Based on Pea and Single Cell Proteins and Hydrocolloids: Effect of Paste Formulation on Process-Induced Fibre Alignment and Structural and Textural Properties. Food Res. Int. 2023, 174, 113633. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, R.; Liang, H. Food Hydrocolloids: Structure, Properties, and Applications. Foods 2024, 13, 1077. [Google Scholar] [CrossRef] [PubMed]
- Mession, J.-L.; Blanchard, C.; Mint-Dah, F.-V.; Lafarge, C.; Assifaoui, A.; Saurel, R. The Effects of Sodium Alginate and Calcium Levels on Pea Proteins Cold-Set Gelation. Food Hydrocoll. 2013, 31, 446–457. [Google Scholar] [CrossRef]
- Poluri, K.M.; Gulati, K.; Sarkar, S. Structural and Functional Properties of Proteins. In Protein-Protein Interactions; Springer: Singapore, 2021; pp. 1–60. [Google Scholar]
- Akbari, M.; Razavi, S.H.; Kieliszek, M. Recent Advances in Microbial Transglutaminase Biosynthesis and Its Application in the Food Industry. Trends Food Sci. Technol. 2021, 110, 458–469. [Google Scholar] [CrossRef]
- Lerner, A.; Benzvi, C. Microbial Transglutaminase Is a Very Frequently Used Food Additive and Is a Potential Inducer of Autoimmune/Neurodegenerative Diseases. Toxics 2021, 9, 233. [Google Scholar] [CrossRef]
- Vasić, K.; Knez, Ž.; Leitgeb, M. Transglutaminase in Foods and Biotechnology. Int. J. Mol. Sci. 2023, 24, 12402. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Roohinejad, S.; George, S.; Barba, F.J.; Greiner, R.; Barbosa-Cánovas, G.V.; Mallikarjunan, K. Innovative Food Processing Technologies on the Transglutaminase Functionality in Protein-Based Food Products: Trends, Opportunities and Drawbacks. Trends Food Sci. Technol. 2018, 75, 194–205. [Google Scholar] [CrossRef]
- Demirkesen, I.; Mert, B.; Costa, R.; Szablewski, T. Rheological and Textural Properties of Food Analogues. In Food Analogues: Emerging Methods and Challenges; Springer Nature: Cham, Switzerland, 2024; pp. 291–328. [Google Scholar]
- dos Santos, M.; Ribeiro, W.O.; Monteiro, J.d.S.; dos Santos, B.A.; Campagnol, P.C.B.; Pollonio, M.A.R. Effect of Transglutaminase Treatment on the Structure and Sensory Properties of Rice- or Soy-Based Hybrid Sausages. Foods 2023, 12, 4226. [Google Scholar] [CrossRef]
- PN-75/A-04018; Agricultural Food Products—Determination of Dry Matter Content. Polish Committee for Standardization: Warsaw, Poland, 1975.
- PN-ISO 1442:2000; Meat and Meat Products—Determination of Moisture Content. Polish Committee for Standardization: Warsaw, Poland, 2000.
- Lee, C.M.; Patel, K.M. Analysis of Juiciness of Commercial Frankfurters. J. Texture Stud. 1984, 15, 67–73. [Google Scholar] [CrossRef]
- PN-ISO 1444:2000; Meat and Meat Products—Determination of Fat Content. Polish Committee for Standardization: Warsaw, Poland, 2000.
- Bourne, M.C. Texture Profile Analysis. Food Technol. 1978, 32, 62–66. [Google Scholar]
- Forghani, Z.; Eskandari, M.H.; Aminlari, M.; Shekarforoush, S.S. Effects of Microbial Transglutaminase on Physicochemical Properties, Electrophoretic Patterns and Sensory Attributes of Veggie Burger. J. Food Sci. Technol. 2017, 54, 2203–2213. [Google Scholar] [CrossRef] [PubMed]
- Yuliana, I.; Mahendradatta, M.; Laga, A. Effects of Transglutaminase Enzyme on Physicochemical Properties of Milkfish (Chanos Chanos) Surimi Gel. Food Res. 2021, 5, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ge, K.; Jiang, L.; Guo, H.; Luo, J.; Wang, F.; Ren, F. Effect of Transglutaminase on Yield, Compositional and Functional Properties of Low-Fat Cheddar Cheese. Food Sci. Technol. Res. 2013, 19, 359–367. [Google Scholar] [CrossRef]
- Moon, S.-H.; Cho, S.-J. Effect of Microbial Transglutaminase Treatment on the Techno-Functional Properties of Mung Bean Protein Isolate. Foods 2023, 12, 1998. [Google Scholar] [CrossRef]
- Qin, X.-S.; Chen, S.-S.; Li, X.-J.; Luo, S.-Z.; Zhong, X.-Y.; Jiang, S.-T.; Zhao, Y.-Y.; Zheng, Z. Gelation Properties of Transglutaminase-Induced Soy Protein Isolate and Wheat Gluten Mixture with Ultrahigh Pressure Pretreatment. Food Bioprocess Technol. 2017, 10, 866–874. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, M.-H.; Bi, X.; Du, J. Physicochemical Properties, Structural Characteristics and in Vitro Digestion of Brown Rice–Pea Protein Isolate Blend Treated by Microbial Transglutaminase. Food Hydrocoll. 2023, 141, 108673. [Google Scholar] [CrossRef]
- Zhu, J.; Deng, H.; Yang, A.; Wu, Z.; Li, X.; Tong, P.; Chen, H. Effect of Microbial Transglutaminase Cross-Linking on the Quality Characteristics and Potential Allergenicity of Tofu. Food Funct. 2019, 10, 5485–5497. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Kim, J.H.; Cha, J.Y.; Kim, T.-K.; Jang, H.W.; Kim, D.-H.; Choi, Y.-S. Quality Characteristics of Meat Analogs through the Incorporation of Textured Vegetable Protein and Tenebrio Molitor Larvae in the Presence of Transglutaminase. Food Sci. Anim. Resour. 2024, 44, 1028–1039. [Google Scholar] [CrossRef]
- Pietrasik, Z.; Jarmoluk, A.; Shand, P.J. Effect of Non-Meat Proteins on Hydration and Textural Properties of Pork Meat Gels Enhanced with Microbial Transglutaminase. LWT-Food Sci. Technol. 2007, 40, 915–920. [Google Scholar] [CrossRef]
- Hui, T.; Xing, G. Effect of Transglutaminase Pre-Crosslinking Treatment Incorporated with Glucono-δ-Lactone on the Physicochemical and Digestive Properties of Tofu. Polymers 2022, 14, 2364. [Google Scholar] [CrossRef]
- Zheng, L.; Regenstein, J.M.; Teng, F.; Li, Y. Tofu Products: A Review of Their Raw Materials, Processing Conditions, and Packaging. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3683–3714. [Google Scholar] [CrossRef] [PubMed]
- PM, R.; Kumar GV, P.; Gaur, A.; Singh, L.; Hossain, S.; Alsulami, T.; Rafiq, S.M.; Nayik, G.A. Microbial Transglutaminase in Cashew-based Vegan Cheese: An Innovative Approach in Achieving Ideal Texture and Meltability. Int. J. Food Sci. Technol. 2024, 59, 8290–8297. [Google Scholar] [CrossRef]
- Wen, Y.; Kim, H.W.; Park, H.J. Effects of Transglutaminase and Cooking Method on the Physicochemical Characteristics of 3D-Printable Meat Analogs. Innov. Food Sci. Emerg. Technol. 2022, 81, 103114. [Google Scholar] [CrossRef]
- Amjanyakun, T.; Ponklang, W.; Butseekhot, S.; Ploypetchara, T.; Siricoon, S.; Sumsakul, S.; Auranwiwat, C.; Sorndech, W. Structural and Physicochemical Properties of Plant-Based Meat Analogues from Transglutaminase-Modified Soybean Protein. Food Res. 2024, 8, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Zeng, X.; Liang, H.; Li, J.; Zhou, B.; Wu, D.; Du, X.; Li, B. Construction of a Soybean Protein Isolate/Polysaccharide-Based Whole Muscle Meat Analog: Physical Properties and Freeze-Thawing Stability Study. Int. J. Biol. Macromol. 2024, 265, 131037. [Google Scholar] [CrossRef]
- Vijayan, P.; Song, Z.; Toy, J.Y.H.; Yu, L.L.; Huang, D. Effect of Transglutaminase on Gelation and Functional Proteins of Mung Bean Protein Isolate. Food Chem. 2024, 454, 139590. [Google Scholar] [CrossRef]
- Lee, E.-J.; Hong, G.-P. Effects of Microbial Transglutaminase and Alginate on the Water-Binding, Textural and Oil Absorption Properties of Soy Patties. Food Sci. Biotechnol. 2020, 29, 777–782. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variants Coding | Variability Factors | |
|---|---|---|
| Microbial Transglutaminase MTG [%] | Incubation Time T [h] | |
| 0_MTG_0T | 0 | 0 |
| 0_MTG_1.5T | 0 | 1.5 |
| 0_MTG_3T | 0 | 3 |
| 0.5_MTG_0T | 0.5 | 0 |
| 0.5_MTG_1.5T | 0.5 | 1.5 |
| 0.5_MTG_3T | 0.5 | 3 |
| 1_MTG_0T | 1 | 0 |
| 1_MTG_1.5T | 1 | 1.5 |
| 1_MTG_3T | 1 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimoch-Korzycka, A.; Krawczyk, A.; Król-Kilińska, Ż.; Kulig, D.; Bobak, Ł.; Jarmoluk, A. Influence of Microbial Transglutaminase on the Formation of Physico-Chemical Properties of Meat Analogs. Foods 2024, 13, 4085. https://doi.org/10.3390/foods13244085
Zimoch-Korzycka A, Krawczyk A, Król-Kilińska Ż, Kulig D, Bobak Ł, Jarmoluk A. Influence of Microbial Transglutaminase on the Formation of Physico-Chemical Properties of Meat Analogs. Foods. 2024; 13(24):4085. https://doi.org/10.3390/foods13244085
Chicago/Turabian StyleZimoch-Korzycka, Anna, Anna Krawczyk, Żaneta Król-Kilińska, Dominika Kulig, Łukasz Bobak, and Andrzej Jarmoluk. 2024. "Influence of Microbial Transglutaminase on the Formation of Physico-Chemical Properties of Meat Analogs" Foods 13, no. 24: 4085. https://doi.org/10.3390/foods13244085
APA StyleZimoch-Korzycka, A., Krawczyk, A., Król-Kilińska, Ż., Kulig, D., Bobak, Ł., & Jarmoluk, A. (2024). Influence of Microbial Transglutaminase on the Formation of Physico-Chemical Properties of Meat Analogs. Foods, 13(24), 4085. https://doi.org/10.3390/foods13244085