


Formation of Cysteine Adducts with Chlorogenic Acid in Coffee Beans

 ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Coffee Samples and Chemicals
2.1.1. Coffee Samples
2.1.2. Chemicals
2.2. Preliminary Analysis
2.2.1. Determination of the Antioxidative Capacity of the Coffee Bean Extracts
2.2.2. Free Amino Acids, Caffeine and Chlorogenic Acid Determination
Free Amino Acids
Analysis of Caffeine and CQA Derivatives
2.3. Analysis of CQA-Adducts
2.4. Effect of the pH Value and Concentration
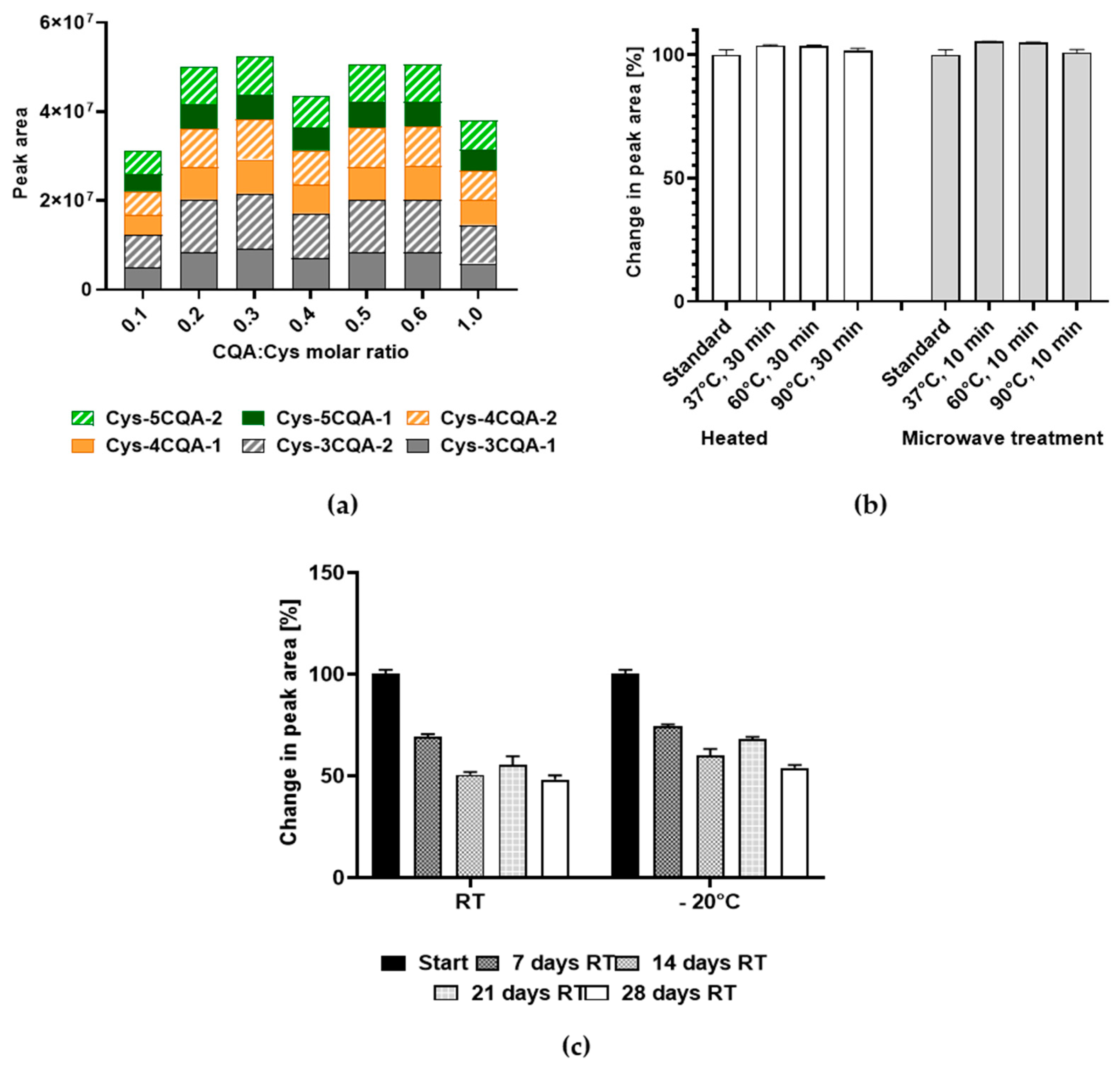
2.5. Stability of the Cys-CQA-Adducts
2.6. Determination of N-Acetylcysteine Modifications as a Function of pH Value
2.7. Determination of Glutathione-CQA Modifications as a Function of pH Value
2.8. Data Analysis
3. Results and Discussion
3.1. Preliminary Investigations
3.1.1. Dry Matter, Antioxidant Capacity, Protein and Caffeine Contents
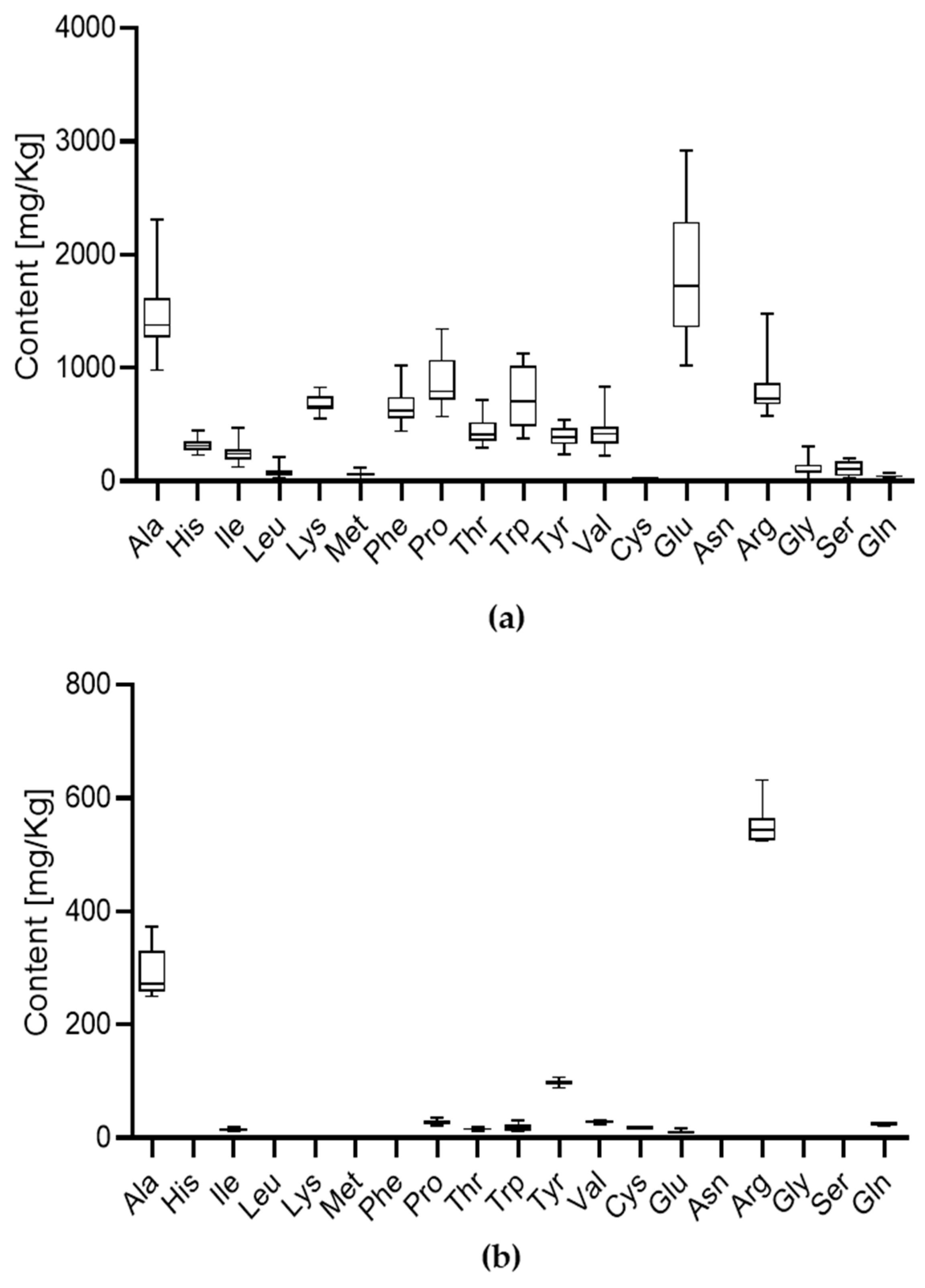
3.1.2. Content of Free Amino Acids
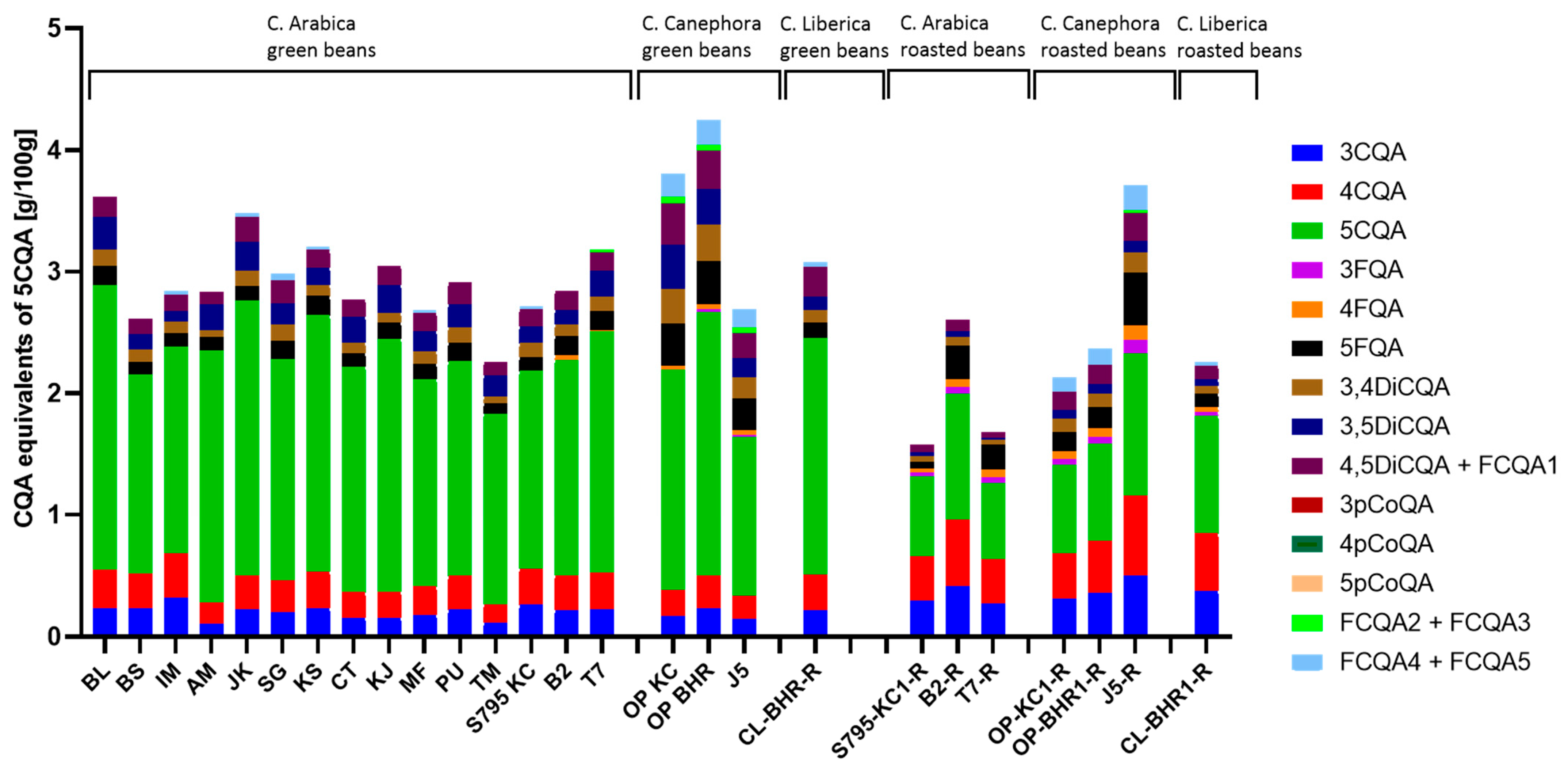
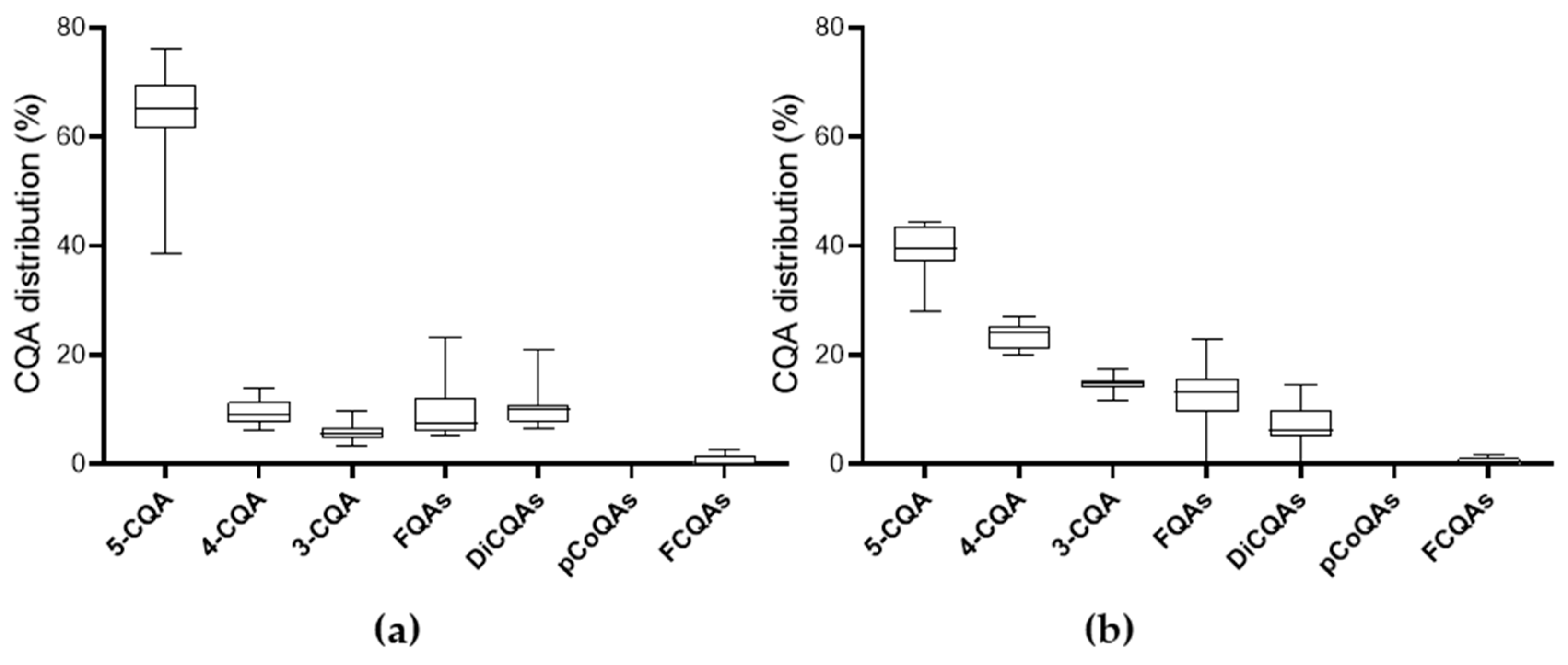
3.1.3. Content of Chlorogenic Acid Derivatives
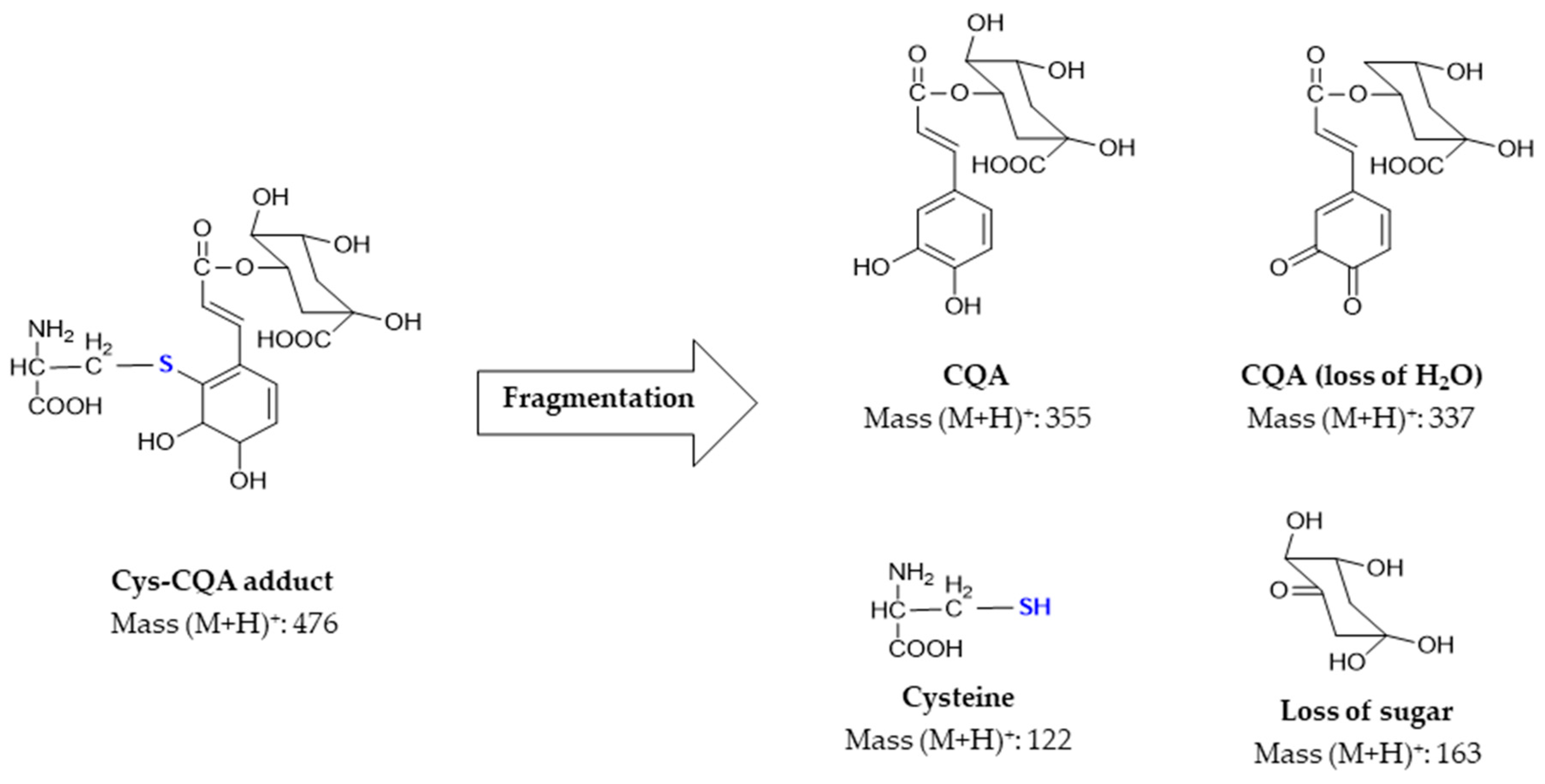
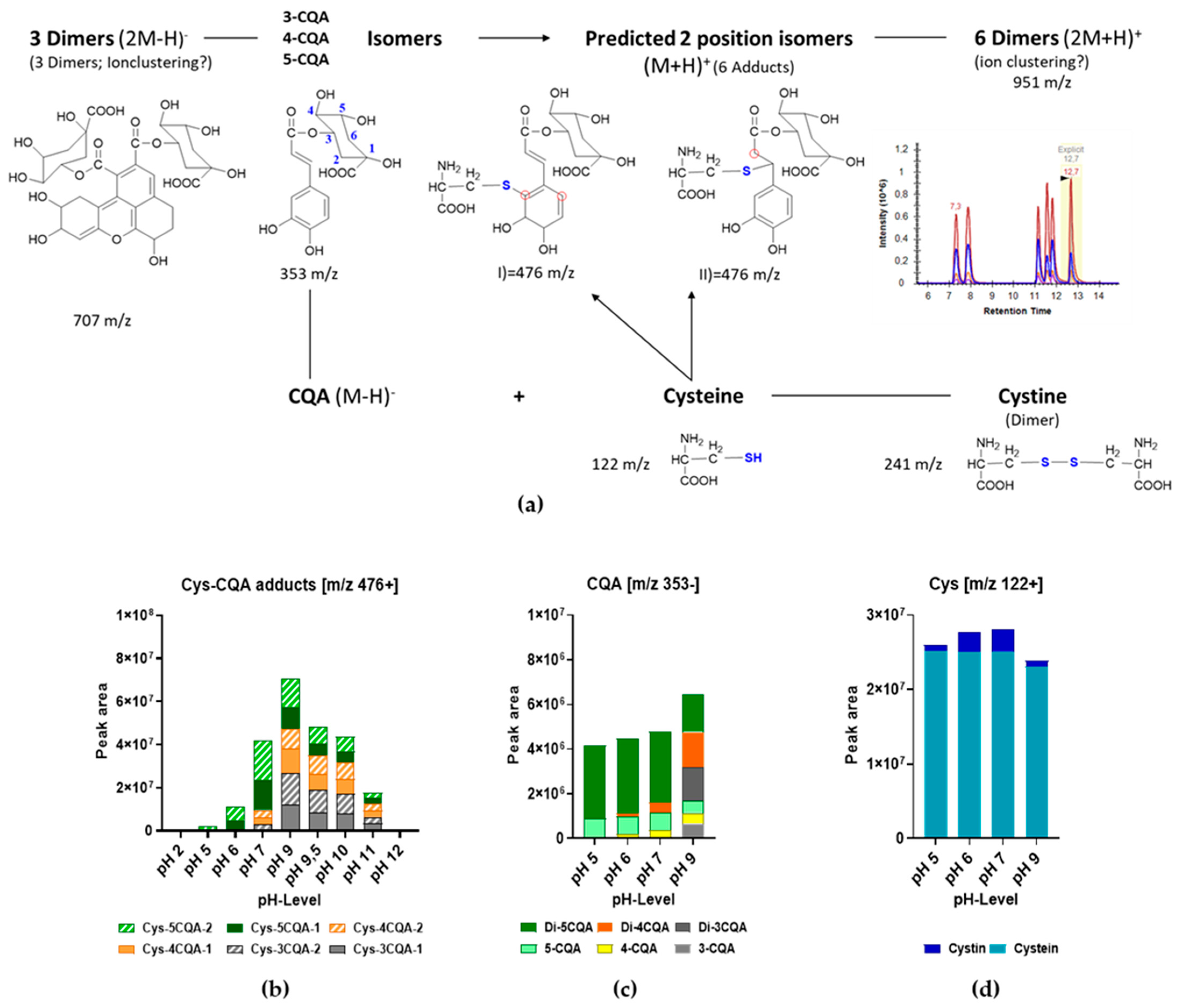
3.2. Formation of Cysteine CQA Adducts
3.3. Effect of pH on Cys-CQA Adduct Formation
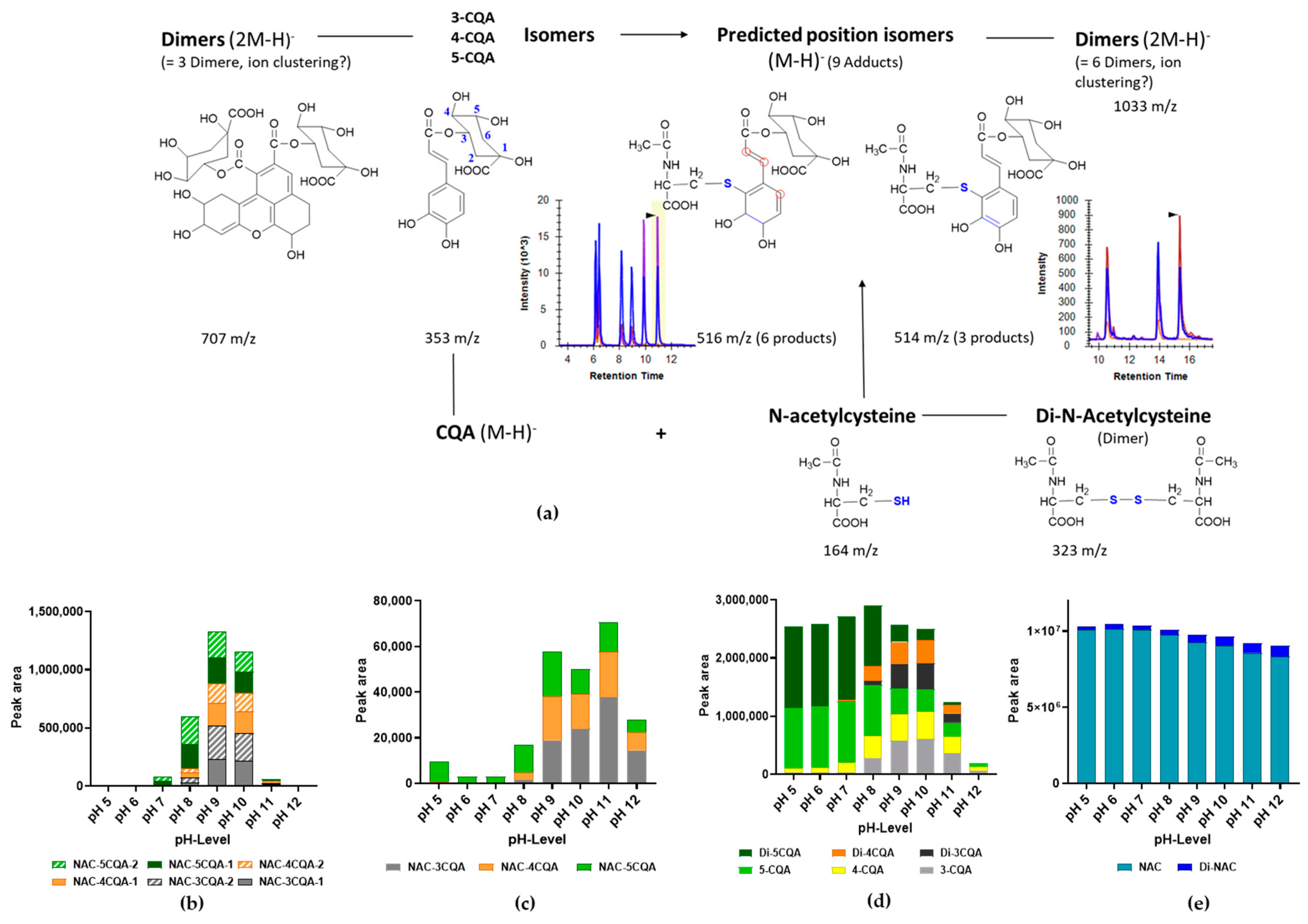
3.4. Formation of CQA Adducts with Other Thiol-Containing Reactants
3.5. Identification of Cysteine-Chlorogenic Acid Adducts in Coffee Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anthony, F.; Bertrand, B.; Etienne, H.; Lashermes, P. Coffea and Psilanthus. In Wild Crop Relatives: Genomic and Breeding Resources: Plantation and Ornamental Crops; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 41–61. [Google Scholar] [CrossRef]
- Campos, G.F.A.; Sagu, S.T.; Celis, P.S.; Rawel, H.M. Comparison of Batch and Continuous Wet-Processing of Coffee: Changes in the Main Compounds in Beans, by-Products and Wastewater. Foods 2020, 9, 1135. [Google Scholar] [CrossRef] [PubMed]
- Farah, A.; Santos, T.F. The Coffee Plant and Beans: An Introduction. In Coffee in Health and Disease Prevention; Preedy, V.R., Ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: New York, NY, USA, 2014. [Google Scholar]
- Rattan, S.; Parande, A.K.; Nagaraju, V.D.; Ghiwari, G.K. A comprehensive review on utilization of wastewater from coffee processing. Environ. Sci. Pollut. Res. 2015, 22, 6461–6472. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J. Kaffee: Konsum und Anbau—Statistiken und Daten. Available online: https://de.statista.com/themen/171/kaffee/#topicOverview (accessed on 3 June 2023).
- Chieng, D.; Canovas, R.; Segan, L.; Sugumar, H.; Voskoboinik, A.; Prabhu, S.; Ling, L.H.; Lee, G.; Morton, J.B.; Kaye, D.M.; et al. The impact of coffee subtypes on incident cardiovascular disease, arrhythmias, and mortality: Long-term outcomes from the UK Biobank. Eur. J. Prev. Cardiol. 2022, 29, 2240–2249. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.; Rocha, T.S.; Prudencio, S.H. Potential of green and roasted coffee beans and spent coffee grounds to provide bioactive peptides. Food Chem. 2021, 348, 129061. [Google Scholar] [CrossRef] [PubMed]
- Rawel, H.M.; Kulling, S.E. Nutritional contribution of coffee, cacao and tea phenolics to human health. J. Verbraucherschutz Leb. 2007, 2, 399–406. [Google Scholar] [CrossRef]
- Esquivel, P.; Jimenez, V.M. Functional properties of coffee and coffee by-products. Food Res. Int. 2012, 46, 488–495. [Google Scholar] [CrossRef]
- Clifford, M. Chlorogenic acids—Their complex nature and routine determination in coffee beans. Food Chem. 1979, 4, 63–71. [Google Scholar] [CrossRef]
- Clifford, M.N. Chlorogenic acids and other cinnamates—Nature, occurrence, dietary burden, absorption and metabolism. J. Sci. Food Agric. 2000, 80, 1033–1043. [Google Scholar] [CrossRef]
- Rawel, H.M.; Rohn, S. Nature of hydroxycinnamate-protein interactions. Phytochem. Rev. 2010, 9, 93–109. [Google Scholar] [CrossRef]
- Farah, A.; Monteiro, M.C.; Calado, V.; Franca, A.S.; Trugo, L.C. Correlation between cup quality and chemical attributes of Brazilian coffee. Food Chem. 2006, 98, 373–380. [Google Scholar] [CrossRef]
- Jaiswal, R.; Kuhnert, N. Hierarchical scheme for liquid chromatography/multi-stage spectrometric identification of 3,4,5-triacyl chlorogenic acids in green Robusta coffee beans. Rapid Commun. Mass Spectrom. 2010, 24, 2283–2294. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.; Kuhnert, N. Identification and characterization of five new classes of chlorogenic acids in burdock (Arctium lappa L.) roots by liquid chromatography/tandem mass spectrometry. Food Funct. 2011, 2, 63–71. [Google Scholar] [CrossRef]
- Awwad, S.; Issa, R.; Alnsour, L.; Albals, D.; Al-Momani, I. Quantification of Caffeine and Chlorogenic Acid in Green and Roasted Coffee Samples Using HPLC-DAD and Evaluation of the Effect of Degree of Roasting on Their Levels. Molecules 2021, 26, 7502. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.; Matei, M.F.; Golon, A.; Witt, M.; Kuhnert, N. Understanding the fate of chlorogenic acids in coffee roasting using mass spectrometry based targeted and non-targeted analytical strategies. Food Funct. 2012, 3, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Yabuta, G.; Koizumi, Y.; Namiki, K.; Hida, M.; Namiki, M. Structure of green pigment formed by the reaction of caffeic acid esters (or chlorogenic acid) with a primary amino compound. Biosci. Biotechnol. Biochem. 2001, 65, 2121–2130. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Homann, T.; Kreisel, J.; Khalil, M.; Puhlmann, R.; Kruse, H.P.; Rawel, H. Characterization and modeling of the interactions between coffee storage proteins and phenolic compounds. J. Agric. Food Chem. 2012, 60, 11601–11608. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Homann, T.; Khalil, M.; Kruse, H.P.; Rawel, H. Milk whey protein modification by coffee-specific phenolics: Effect on structural and functional properties. J. Agric. Food Chem. 2013, 61, 6911–6920. [Google Scholar] [CrossRef] [PubMed]
- Bongartz, V.; Brandt, L.; Gehrmann, M.L.; Zimmermann, B.F.; Schulze-Kaysers, N.; Schieber, A. Evidence for the Formation of Benzacridine Derivatives in Alkaline-Treated Sunflower Meal and Model Solutions. Molecules 2016, 21, 91. [Google Scholar] [CrossRef] [PubMed]
- Namiki, M.; Yabuta, G.; Koizumi, Y.; Yano, M. Development of free radical products during the greening reaction of caffeic acid esters (or chlorogenic acid) and a primary amino compound. Biosci. Biotechnol. Biochem. 2001, 65, 2131–2136. [Google Scholar] [CrossRef] [PubMed]
- Schilling, S.; Sigolotto, C.I.; Carle, R.; Schieber, A. Characterization of covalent addition products of chlorogenic acid quinone with amino acid derivatives in model systems and apple juice by high-performance liquid chromatography/electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2008, 22, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Poojary, M.M.; Hellwig, M.; Henle, T.; Lund, M.N. Covalent bonding between polyphenols and proteins: Synthesis of caffeic acid-cysteine and chlorogenic acid-cysteine adducts and their quantification in dairy beverages. Food Chem. 2023, 403, 134406. [Google Scholar] [CrossRef]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [PubMed]
- Belitz, H.-D.; Grosch, W.; Schieberle, P. Food Chemistry, 6th ed.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 2009, pp. 969–1003. [Google Scholar]
- Bekedam, E.K.; Loots, M.J.; Schols, H.A.; Van Boekel, M.A.; Smit, G. Roasting effects on formation mechanisms of coffee brew melanoidins. J. Agric. Food Chem. 2008, 56, 7138–7145. [Google Scholar] [CrossRef] [PubMed]
- Bekedam, E.K.; Schols, H.A.; van Boekel, M.A.; Smit, G. High molecular weight melanoidins from coffee brew. J. Agric. Food Chem. 2006, 54, 7658–7666. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xian, F.; Ou, J.; Jiang, K.; Zheng, J.; Ou, S.; Liu, F.; Rao, Q.; Huang, C. Formation and Identification of Six Amino Acid—Acrylamide Adducts and Their Cytotoxicity Toward Gastrointestinal Cell Lines. Front. Nutr. 2022, 9, 902040. [Google Scholar] [CrossRef] [PubMed]
- Arnold, U.; Ludwig, E. Analysis of free amino acids in green coffee beans. II. Changes of the amino acid content in arabica coffees in connection with post-harvest model treatment. Z. Lebensm. Unters. Forsch. 1996, 203, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Farah, A.; Donangelo, C.M. Phenolic compounds in coffee. Braz. J. Plant Physiol. 2006, 18, 23–36. [Google Scholar] [CrossRef]
- Jeszka-Skowron, M.; Sentkowska, A.; Pyrzyńska, K.; De Peña, M.P. Chlorogenic acids, caffeine content and antioxidant properties of green coffee extracts: Influence of green coffee bean preparation. Eur. Food Res. Technol. 2016, 242, 1403–1409. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.-J.; Jeong, H.-J.; Moon, H.; Kim, H.-N.; Cho, J.-H.; Lee, J.-E.; Hong, K.S.; Hong, Y.-S. Assessment of green coffee bean metabolites dependent on coffee quality using a 1H NMR-based metabolomics approach. Food Res. Int. 2015, 67, 175–182. [Google Scholar] [CrossRef]
- Farah, A.; de Paulis, T.; Trugo, L.C.; Martin, P.R. Effect of roasting on the formation of chlorogenic acid lactones in coffee. J. Agric. Food Chem. 2005, 53, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, J.; Sun, J.; Qiu, S.; Chu, B.; Fang, R.; Li, L.; Gong, J.; Zheng, F. Degradation kinetics and isomerization of 5-O-caffeoylquinic acid under ultrasound: Influence of epigallocatechin gallate and vitamin C. Food Chem. X 2021, 12, 100147. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jongberg, S.; Andersen, M.L.; Davies, M.J.; Lund, M.N. Quinone-induced protein modifications: Kinetic preference for reaction of 1,2-benzoquinones with thiol groups in proteins. Free Radic. Biol. Med. 2016, 97, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Drucker, C.T.; Cicali, A.R.; Roberts, A.M.P.; Hughey, C.A.; Senger, L.W. Identification of alkaline-induced thiolyl-chlorogenic acid conjugates with cysteine and glutathione. Food Chem. 2023, 423, 136267. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Poyatos, M.D.P.; Ruiz-Medina, A.; Zengin, G.; Llorent-Martinez, E.J. Phenolic Characterization, Antioxidant Activity, and Enzyme Inhibitory Properties of Berberis thunbergii DC. Leaves: A Valuable Source of Phenolic Acids. Molecules 2019, 24, 4171. [Google Scholar] [CrossRef] [PubMed]
- Jongberg, S.; Lund, M.N.; Otte, J. Dissociation and reduction of covalent beta-lactoglobulin-quinone adducts by dithiothreitol, tris(2-carboxyethyl)phosphine, or sodium sulfite. Anal. Biochem. 2015, 478, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Campos, G.A.F.; Kruizenga, J.G.K.T.; Sagu, S.T.; Schwarz, S.; Homann, T.; Taubert, A.; Rawel, H.M. Effect of the Post-Harvest Processing on Protein Modification in Green Coffee Beans by Phenolic Compounds. Foods 2022, 11, 159. [Google Scholar] [CrossRef] [PubMed]
- Rao, N.Z.; Fuller, M. Acidity and Antioxidant Activity of Cold Brew Coffee. Sci. Rep. 2018, 8, 16030. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coffee Samples | Dry Matter (g/100 g) | Antioxidant Capacity (g/100 g) | Protein Content (g/100 g) | Caffeine Content | ||
|---|---|---|---|---|---|---|
| UV (g/100 g) | MS (g/100 g) | |||||
| Green coffee beans | BL | 91.14 ± 0.16 a | 3.03 ± 0.19 a | 13.29 ± 0.06 a | 0.826 ± 0.001 a | 1.337 ± 0.003 a |
| BS | 91.36 ± 0.02 a,b | 3.02 ± 0.03 a | 13.62 ± 0.09 a | 0.593 ± 0.002 a | 0.967 ± 0.001 a | |
| IM | 91.81 ± 0.01 b | 2.95 ± 0.26 a | 12.68 ± 0.16 b | 0.684 ± 0.004 a | 1.206 ± 0.003 a | |
| AM | 91.35 ± 0.14 a,b | 3.12 ± 0.10 a | 11.28 ± 0.05 c | 0.520 ± 0.002 a | 0.856 ± 0.003 b | |
| JK | 91.52 ± 0.02 a,b | 2.91 ± 0.20 a | 12.31 ± 0.10 d | 0.702 ± 0.008 a | 1.142 ± 0.001 a,b | |
| SG | 91.07 ± 0.03 a | 3.28 ± 0.21 b | 12.34 ± 0.08 d | 0.619 ± 0.000 a | 1.014 ± 0.001 a,b | |
| KS | 90.94 ± 0.04 a | 3.00 ± 0.15 a | 12.39 ± 0.02 b,d | 0.576 ± 0.000 a | 0.998 ± 0.008 a,b | |
| CT | 90.01 ± 0.09 c | 2.36 ± 0.18 c | 12.35 ± 0.08 b | 0.635 ± 0.003 a | 1.040 ± 0.005 a,b | |
| KJ | 90.04 ± 0.02 c | 3.29 ± 0.19 b | 11.49 ± 0.02 c | 0.688 ± 0.004 a | 1.093 ± 0.002 a,b | |
| MF | 91.03 ± 0.02 a,d | 2.54 ± 0.06 c | 11.94 ± 0.20 e | 0.538 ± 0.00 a | 0.961 ± 0.008 a,b | |
| PU | 91.06 ± 0.06 a | 2.56 ± 0.07 c | 12.57 ± 0.18 b | 0.651 ± 0.001 a | 1.058 ± 0.004 a,b | |
| TM | 90.30 ± 0.01 c,e | 2.71 ± 0.13 c | 11.72 ± 0.08 e | 0.531 ± 0.002 a | 0.865 ± 0.006 b | |
| S795 KC | 92.84 ± 0.23 f | 2.71 ± 0.04 c | 12.72 ± 0.17 b | 0.543 ± 0.002 a | 0.750 ± 0.002 b | |
| B2 | 90.59 ± 0.04 d,e | 2.18 ± 0.05 c | 13.11 ± 0.11 a | 0.785 ± 0.000 a | 0.794 ± 0.007 b | |
| T7 | 92.02 ± 0.01 b | 2.50 ± 0.04 c | 13.09 ± 0.16 a | 0.859 ± 0.002 a | 0.887 ± 0.003 b | |
| OP KC | 92.56 ± 0.34 f | 3.39 ± 0.13 b | 15.87 ± 0.22 f | 0.857 ± 0.005 a | 1.515 ± 0.051 a | |
| OP BHR | 93.16 ± 0.15 f | 3.23 ± 0.03 a,b | 16.20 ± 0.25 f | 1.153 ± 0.007 a | 1.792 ± 0.014 c | |
| J5 | 88.89 ± 0.08 g | 2.77 ± 0.26 a,d | 15.32 ± 0.01 g | 0.944 ± 0.001 a | 0.965 ± 0.006 a,b | |
| CL-BHR | 93.17 ± 0.14 f | 2.32 ± 0.16 c | 12.72 ± 0.43 b | 0.537 ± 0.002 a | 0.804 ± 0.006 b | |
| Roasted coffee beans | S795-KC-R | 95.83 ± 0.14 h | 2.75 ± 0.09 a,d | 12.88 ± 0.24 a,b | 0.633 ± 0.001 a | 0.965 ± 0.004 a,b |
| B2-R | 98.65 ± 0.05 i | 3.13 ± 0.30 a,b | 13.18 ± 0.04 a | 1.389 ± 0.003 b | 1.428 ± 0.001 a,c | |
| T7-R | 95.64 ± 0.06 h | 2.53 ± 0.07 c,d | 13.55 ± 0.10 a | 1.309 ± 0.009 b | 1.344 ± 0.001 a,b | |
| OP KC-R | 96.26 ± 0.20 j | 3.58 ± 0.12 b | 16.40 ± 0.59 f | 1.053 ± 0.002 a,b | 1.659 ± 0.010 a,c | |
| OP BHR-R | 95.94 ± 0.12 h | 3.34 ± 0.21 a,b | 16.29 ± 0.27 f | 1.111 ± 0.002 a,b | 1.557 ± 0.035 a,c | |
| J5-R | 98.07 ± 0.11 k | 4.31 ± 0.18 e | 15.60 ± 0.02 g | 2.549 ± 0.009 c | 2.572 ± 0.010 d | |
| CL-BHR-R | 95.96 ± 0.24 h,j | 3.41 ± 0.25 a,b | 12.77 ± 0.29 b | 0.643 ± 0.003 a | 0.859 ± 0.002 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sagu, S.T.; Ulbrich, N.; Morche, J.R.; Nichani, K.; Özpinar, H.; Schwarz, S.; Henze, A.; Rohn, S.; Rawel, H.M. Formation of Cysteine Adducts with Chlorogenic Acid in Coffee Beans. Foods 2024, 13, 1660. https://doi.org/10.3390/foods13111660
Sagu ST, Ulbrich N, Morche JR, Nichani K, Özpinar H, Schwarz S, Henze A, Rohn S, Rawel HM. Formation of Cysteine Adducts with Chlorogenic Acid in Coffee Beans. Foods. 2024; 13(11):1660. https://doi.org/10.3390/foods13111660
Chicago/Turabian StyleSagu, Sorel Tchewonpi, Nina Ulbrich, Johanna Rebekka Morche, Kapil Nichani, Haydar Özpinar, Steffen Schwarz, Andrea Henze, Sascha Rohn, and Harshadrai M. Rawel. 2024. "Formation of Cysteine Adducts with Chlorogenic Acid in Coffee Beans" Foods 13, no. 11: 1660. https://doi.org/10.3390/foods13111660
APA StyleSagu, S. T., Ulbrich, N., Morche, J. R., Nichani, K., Özpinar, H., Schwarz, S., Henze, A., Rohn, S., & Rawel, H. M. (2024). Formation of Cysteine Adducts with Chlorogenic Acid in Coffee Beans. Foods, 13(11), 1660. https://doi.org/10.3390/foods13111660