Exogenous Methyl Jasmonate (MeJA) Improves ‘Ruixue’ Apple Fruit Quality by Regulating Cell Wall Metabolism


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Treatments and Plant Materials
2.2. Detection of Physiological Characteristics
2.3. Determination of Cell Wall Component Content
2.4. Determination of Cell Wall Degradation-Related Enzyme Activities
2.5. Determination of Relative Gene Expression
2.6. Transient Expression of MdPL18 in Apple Fruit
2.7. Stable Overexpression of MdPL18 in Apple Calli
2.8. Statistical Analysis
3. Results
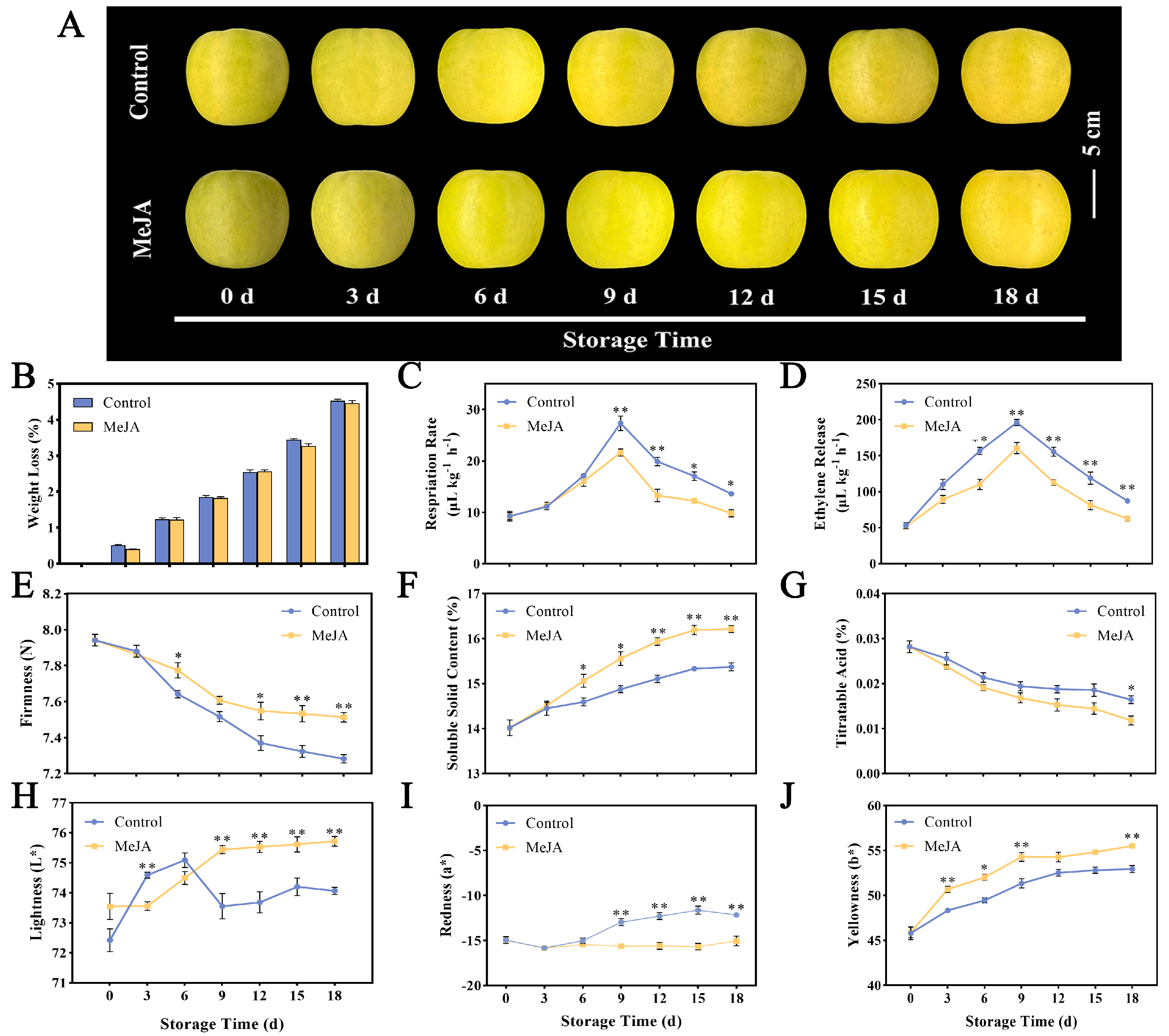
3.1. Changes in Quality Parameters of Apple Fruit Treated with Exogenous MeJA
3.2. Effects of Exogenous MeJA Treatment on Apple Fruit Cell Wall Components
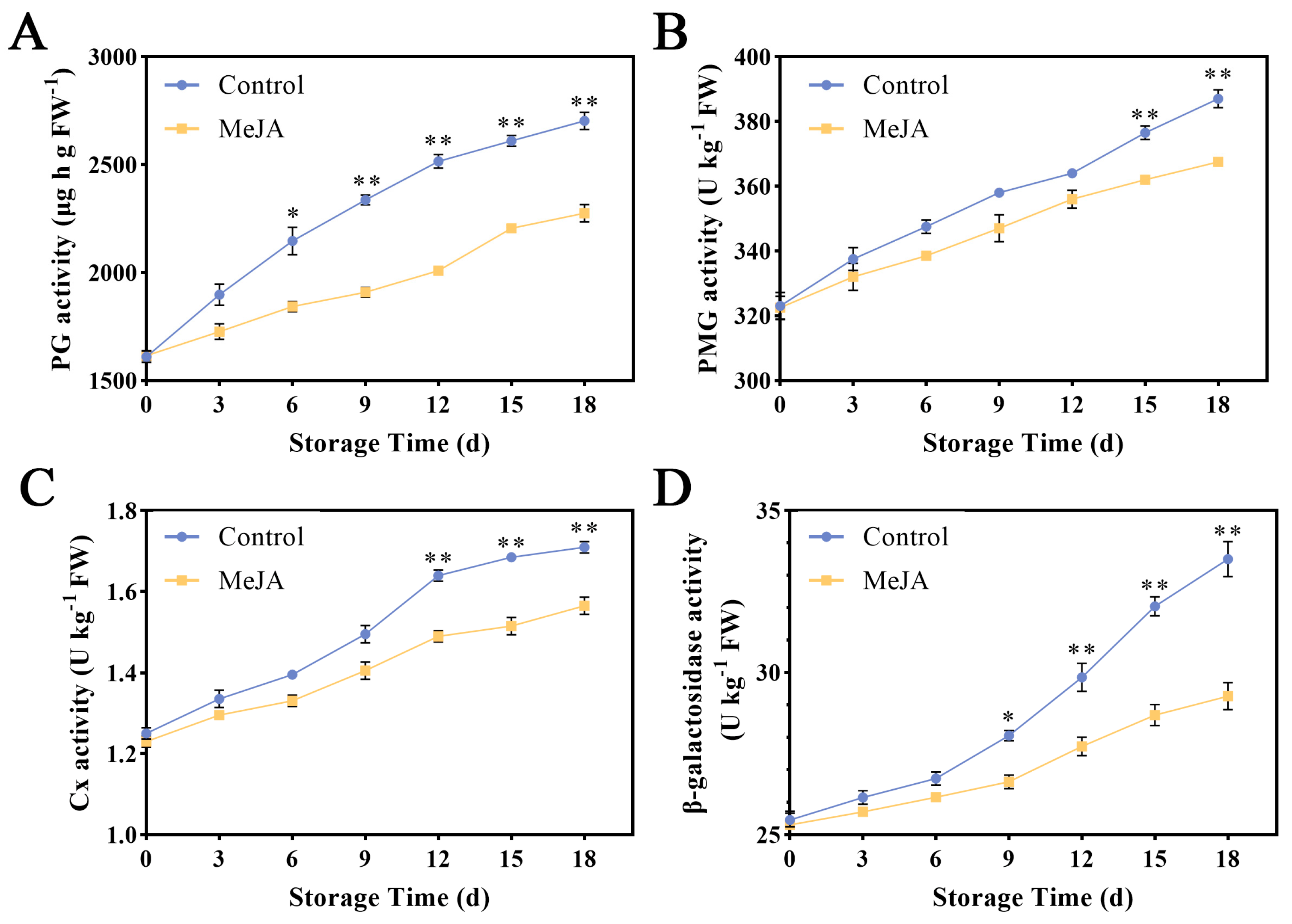
3.3. Effects of Exogenous MeJA Treatment on the Activities of Cell Wall-Degrading Enzymes in Apple Fruit
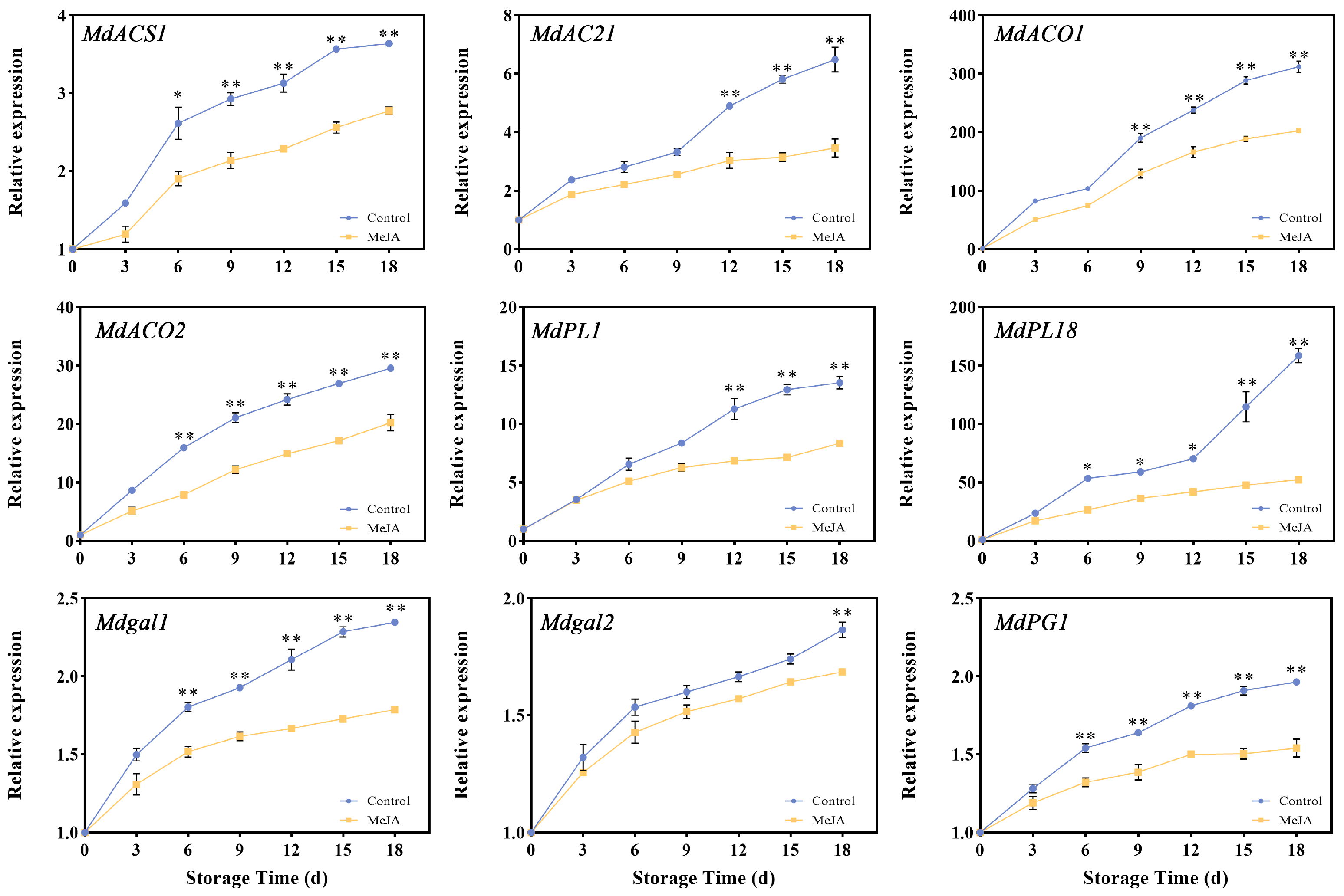
3.4. Effects of Exogenous MeJA Treatment on the Cell Wall- and Ethylene-Related Enzyme Genes
3.5. Correlation between the Candidate Gene MdPL18 (MD16G1070600) and Fruit Quality
3.6. Transient Overexpression of MdPL18 Promotes Ripening and Softening of ‘Ruixue’ Fruit
3.7. Effect of Stable Overexpression of MdPL18 in Apple Calli on Cell Wall Material Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Abbreviation | Definition |
| MeJA | methyl jasmonate |
| WSP | water-soluble pectin |
| CSP | chelator-soluble pectin |
| ISP | ion-soluble pectin |
| PME | pectin methylesterase |
| PG | polygalacturonase |
| β-Gal | glycosidase |
| PAL | phenylalanine ammonia-lyase |
| POD | peroxidase |
| PPO | polyphenol oxidase |
| ETH | ethylene |
| SSC | soluble solids content |
| TA | titratable acidity |
| HC | hemicellulose |
| CWM | cell wall material |
| PMG | polymethylgalacturonase |
| Cx | cellulase |
References
- Zhao, J.; Quan, P.; Liu, H.; Li, L.; Xing, L. Transcriptomic and Metabolic Analyses Provide New Insights into the Apple Fruit Quality Decline during Long-Term Cold Storage. J. Agric. Food Chem. 2020, 68, 4699–4716. [Google Scholar] [CrossRef]
- Chang, L.Y.; Brecht, J.K. Responses of 1-methylcyclopropene (1-MCP)-treated banana fruit to pre- and post-treatment ethylene exposure. Sci. Hortic. 2023, 309, 111636. [Google Scholar] [CrossRef]
- Fang, H.; Zuo, J.; Ma, Q.; Zhang, X.; Xu, Y.; Ding, S.; Wang, J.; Luo, Q.; Li, Y.; Wu, C.; et al. Phytosulfokine promotes fruit ripening and quality via phosphorylation of transcription factor DREB2F in tomato. Plant Physiol. 2024, kiae012. [Google Scholar] [CrossRef] [PubMed]
- Favre, L.; Hunter, D.A.; O’donoghue, E.M.; Erridge, Z.A.; Napier, N.J.; Somerfield, S.D.; Hunt, M.; McGhie, T.K.; Cooney, J.M.; Saei, A.; et al. Integrated multi-omic analysis of fruit maturity identifies biomarkers with drastic abundance shifts spanning the harvest period in ‘Royal Gala’ apple. Postharvest Biol. Technol. 2022, 193, 925–945. [Google Scholar] [CrossRef]
- Cornelius, S.; BarryJames, J. Giovannoni. Ethylene and Fruit Ripening. J. Plant Growth Regul. 2007, 26, 143–159. [Google Scholar] [CrossRef]
- Lim, S.; Lee, J.G.; Lee, E.J. Comparison of fruit quality and GC-MS-based metabolite profiling of kiwifruit ‘Jecy green’: Natural and exogenous ethylene-induced ripening. Food Chem. 2017, 234, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Tatsuki, M.; Endo, A.; Ohkawa, H. Influence of time from harvest to 1-MCP treatment on apple fruit quality and expression of genes for ethylene biosynthesis enzymes and ethylene receptors. Postharvest Biol. Technol. 2007, 43, 28–35. [Google Scholar] [CrossRef]
- Wang, S.; Liu, C.; Su, X.; Chen, L.; Zhu, Z. Transcriptome analysis reveals key metabolic pathways and gene expression involving in cell wall polysaccharides-disassembling and postharvest fruit softening in custard apple (Annona squamosa L.). Int. J. Biol. Sci. 2023, 240, 141–158. [Google Scholar] [CrossRef]
- Bank, A.D. Cell wall disassembly in ripening fruit. Funct. Plant Biol. 2006, 33, 103–119. [Google Scholar] [CrossRef]
- Jarvis, M.C. Plant cell walls: Supramolecular assemblies. Food Hydrocoll. 2011, 25, 257–262. [Google Scholar] [CrossRef]
- Liu, M.P.; Wang, R.; Sun, W.W.; Han, W.J.; Li, G.; Zong, W.; Fu, J.M. Effects of postharvest calcium treatmenttreatment on the firmness of persimmon (Diospyros kaki) fruit based on a decline in WSP. Sci. Hortic. 2023, 307, 111490. [Google Scholar] [CrossRef]
- Creelman, R.A.; Mullet, J.E. Biosynthesis and Action of Jasmonates in Plants. Annu. Rev. Plant Biol. 1997, 48, 355–381. [Google Scholar] [CrossRef] [PubMed]
- Pandita, D. Jasmonates: Key players in plant stress tolerance. In Emerging Plant Growth Regulators in Agriculture; Academic Press: Cambridge, MA, USA, 2022; Chapter 5. [Google Scholar] [CrossRef]
- Dobritzsch, S.; Weyhe, M.; Schubert, R.; Dindas, J.; Hause, G.; Kopka, J.; Hause, B. Dissection of jasmonate functions in tomato stamen development by transcriptome and metabolome analyses. BMC Biol. 2015, 13, 28. [Google Scholar] [CrossRef]
- Cao, S.F.; Cai, Y.T.; Yang, Z.F.; Joyce, D.C.; Zheng, Y.H. Effect of MeJA treatment on polyamine, energy status and anthracnose rot of loquat fruit. Food Chem. 2014, 145, 86–89. [Google Scholar] [CrossRef]
- Mustafa, M.A.; Ali, A.; Seymour, G.; Tucker, G. Enhancing the antioxidant content of carambola (Averrhoa carambola) during cold storage and methyl jasmonate treatments. Postharvest Biol. Technol. 2016, 118, 79–86. [Google Scholar] [CrossRef]
- Zhu, L.; Yu, H.; Dai, X.; Yu, M.; Yu, Z. Effect of methyl jasmonate on the quality and antioxidant capacity by modulating ascorbate-glutathione cycle in peach fruit. Sci. Hortic. 2022, 303, 111216. [Google Scholar] [CrossRef]
- Chang, L.L.; Zhang, Y.T.; Wang, G.X.; Dong, J.; Zhong, C.F.; Wang, L.N.; Li, T.H. The effects of exogenous methyl jasmonate on FaNES1 gene expression and the biosynthesis of volatile terpenes in strawberry (Fragaria × ananassa Duch.) fruit. J. Hortic. Sci. Biotech. 2013, 88, 393–398. [Google Scholar] [CrossRef]
- Jiang, J.J.; Yao, L.N.; Yu, Y.J.; Lv, M.L.; Miao, Y.; Cao, J.S. PECTATE LYASE-LIKE10 is associated with pollen wall development in Brassica campestris. J. Integr. Plant Biol. 2014, 56, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yeats, T.H.; Uluisik, S.; Rose, J.K.C.; Seymour, G.B. Fruit softening: Revisiting the role of pectin. Trends Plant Sci. 2018, 23, 302–310. [Google Scholar] [CrossRef]
- Wieczorek, K.; Elashry, A.; Quentin, M.; Grundler, F.M.; Favery, B.; Seifert, G.J.; Bohlmann, H. A distinct role of pectate lyases in the formation of feeding structures induced by cyst and root-knot nematodes. Sci. Rep. 2014, 27, 901–912. [Google Scholar] [CrossRef]
- Ke, X.; Wang, H.; Li, Y.; Zhu, B.; Zang, Y.; He, Y.; Cao, J.; Zhu, Z.; Yu, Y. Genome-wide identification and analysis of polygalacturonase genes in Solanum lycopersicum. Int. J. Mol. Sci. 2018, 19, 2290. [Google Scholar] [CrossRef] [PubMed]
- Marín-Rodríguez, M.C.; Orchard, J.; Seymour, G.B. Pectate lyases cell wall degradation and fruit softening. J. Exp. Bot. 2002, 53, 2115–2119. [Google Scholar] [CrossRef] [PubMed]
- Uluisik, S.; Chapman, N.H.; Smith, R.; Poole, M.; Adams, G.; Gillis, R.B.; Besong, T.M.; Sheldon, J.; Stiegelmeyer, S.; Perez, L. Genetic improvement of tomato by targeted control of fruit softening. Nat. Biotechnol. 2016, 34, 950–952. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Doménech, N.; Jiménez-Bemudez, S.; Matas, A.J.; Rose, J.K.; Munoz-Blanco, J.; Mercado, J.A.; Quesada, M.A. Antisense inhibition of a pectate lyase gene supports a role for pectin depolymerization in strawberry fruit softening. J. Exp. Bot. 2008, 59, 2769–2779. [Google Scholar] [CrossRef]
- Lv, J.Y.; Zhang, M.Y.; Zhang, J.H.; Ge, Y.H.; Li, C.Y.; Meng, K.; Li, J.R. Effects of methyl jasmonate on expression of genes involved in ethylene biosynthesis and signaling pathway during postharvest ripening of apple fruit. Sci. Hortic. 2018, 229, 157–166. [Google Scholar] [CrossRef]
- Yang, X.T.; Song, J.; Du, L.N.; Charles, F.; Leslie, C.P.; Sherry, F.; Zhang, Z.Q. Ethylene and 1-MCP regulate major volatile biosynthetic pathways in apple fruit. Food Chem. 2016, 194, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Melton, L.D.; Smith, B.G. Isolation of Plant Cell Walls and Fractionation of Cell Wall Polysaccharides. Curr. Protoc. Food Anal. Chem. 2001, E3.1.1–E3.1.23. [Google Scholar] [CrossRef]
- Li, X.L.; Su, Q.F.; Jia, R.J.; Wang, Z.D.; Fu, J.H.; Guo, J.H.; Zhao, Z.Y. Comparison of cell wall changes of two different types of apple cultivars during fruit development and ripening. J. Inter. Agric. 2023, 22, 2705–2718. [Google Scholar] [CrossRef]
- Payasi, A.; Misra, P.C.; Sanwal, G.G. Effect of phytohormones on pectate lyase activity in ripening Musa acuminata—ScienceDirect. Plant Physiol. Biochem. 2004, 42, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://meme-suite.org/meme/tools/meme (accessed on 1 May 2024).
- Park, W.T.; Kim, Y.B.; Seo, J.M.; Kim, S.J.; Chung, E.; Lee, J.H.; Park, S.U. Accumulation of Anthocyanin and Associated Gene Expression in Radish Sprouts Exposed to Light and Methyl Jasmonate. Chem. Rev. 2013, 61, 4127–4132. [Google Scholar] [CrossRef]
- Alejandra, M.E.; Pedro, J.Z.; Salvador, C.; Fabian, G.; Domingo, M.R.; Daniel, V.; Maria, S. Preharvest application of methyl jasmonate (MeJA) in two plum cultivars. 1. Improvement of fruit growth and quality attributes at harvest. Postharvest Biol. Technol. 2014, 98, 98–105. [Google Scholar] [CrossRef]
- Kucuker, E.; Ozturk, B.; Celik, S.M.; Aksit, H. Pre-harvest spray application of methyl jasmonate plays an important role in fruit ripening, fruit quality and bioactive compounds of Japanese plums. Sci. Hortic. 2014, 176, 162–169. [Google Scholar] [CrossRef]
- Li, Z.; Min, D.; Fu, X.; Zhao, X.; Wang, J.; Zhang, X.; Li, F.; Li, X. The roles of SlMYC2 in regulating ascorbate-glutathione cycle mediated by methyl jasmonate in postharvest tomato fruits under cold stress. Sci. Hortic. 2021, 288, 110406. [Google Scholar] [CrossRef]
- Kondo, S.; Setha, S.; Rudell, D.R.; Buchanan, D.A.; Mattheis, J.P. Aroma volatile biosynthesis in apples affected by 1-MCP and methyl jasmonate. Postharvest Biol. Technol. 2005, 36, 61–68. [Google Scholar] [CrossRef]
- Han, Y.L.; Chen, C.; Yan, Z.M.; Li, J.; Wang, Y.H. The methyl jasmonate accelerates the strawberry fruits ripening process. Sci. Hortic. 2019, 249, 250–256. [Google Scholar] [CrossRef]
- Wang, H.; Wu, X.; Fan, C.; Li, G.J.; Dong, T.T. Retardation of postharvest softening of blueberry fruit by methyl jasmonate is correlated with altered cell wall modification and energy metabolism. Sci. Hortic. 2021, 276, 109752. [Google Scholar] [CrossRef]
- Jing, S.Y.; Liu, Y.; Liu, H.F.; Li, N.; Li, L. Methyl jasmonate regulates protective enzyme activities to improve resistance to Venturia nashicola in pear (Pyrus bretschneideri Rehd.). Eur. J. Plant Pathol. 2020, 158, 789–797. [Google Scholar] [CrossRef]
- Gupta, R.; Mehta, G.; Deswal, D.; Sharma, S.; Singh, A. Cellulases and Their Biotechnological Applications. In Biotechnology for Environmental Management and Resource Recovery; Springer: New Delhi, India, 2013; pp. 89–106. [Google Scholar] [CrossRef]
- Li, T.; Jiang, Z.Y.; Zhang, L.C.; Tan, D.M.; Wei, Y.; Yuan, H.; Wang, A.D. Apple (Malus domestica) MdERF2 negatively affects ethylene biosynthesis during fruit ripening by suppressing MdACS1 transcription. Plant J. 2016, 88, 735–748. [Google Scholar] [CrossRef]
- Schaffer, R.J.; Friel, E.N.; Souleyre, E.J.; Bolitho, K.; Thodey, K.; Ledger, S.; Bowen, J.H.; Ma, J.-H.; Nain, B.; Cohen, D.; et al. A Genomics Approach Reveals That Aroma Production in Apple Is Controlled by Ethylene Predominantly at the Final Step in Each Biosynthetic Pathway. Plant Physiol. 2007, 144, 1899–1912. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Wang, B.; Gong, Y.; Yan, X.; Chen, X.; Zhong, Y.; Zhao, Z. Exogenous Methyl Jasmonate (MeJA) Improves ‘Ruixue’ Apple Fruit Quality by Regulating Cell Wall Metabolism. Foods 2024, 13, 1594. https://doi.org/10.3390/foods13111594
Ding X, Wang B, Gong Y, Yan X, Chen X, Zhong Y, Zhao Z. Exogenous Methyl Jasmonate (MeJA) Improves ‘Ruixue’ Apple Fruit Quality by Regulating Cell Wall Metabolism. Foods. 2024; 13(11):1594. https://doi.org/10.3390/foods13111594
Chicago/Turabian StyleDing, Xiaoyi, Bin Wang, Yubo Gong, Xueqing Yan, Xinxin Chen, Yuanwen Zhong, and Zhengyang Zhao. 2024. "Exogenous Methyl Jasmonate (MeJA) Improves ‘Ruixue’ Apple Fruit Quality by Regulating Cell Wall Metabolism" Foods 13, no. 11: 1594. https://doi.org/10.3390/foods13111594
APA StyleDing, X., Wang, B., Gong, Y., Yan, X., Chen, X., Zhong, Y., & Zhao, Z. (2024). Exogenous Methyl Jasmonate (MeJA) Improves ‘Ruixue’ Apple Fruit Quality by Regulating Cell Wall Metabolism. Foods, 13(11), 1594. https://doi.org/10.3390/foods13111594