
Effective Therapeutic Verification of Crocin I, Geniposide, and Gardenia (Gardenia jasminoides Ellis) on Type 2 Diabetes Mellitus In Vivo and In Vitro



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Isolation and Identification of Geniposide and Crocetin
2.3. Inhibition of Enzyme Activity In Vitro
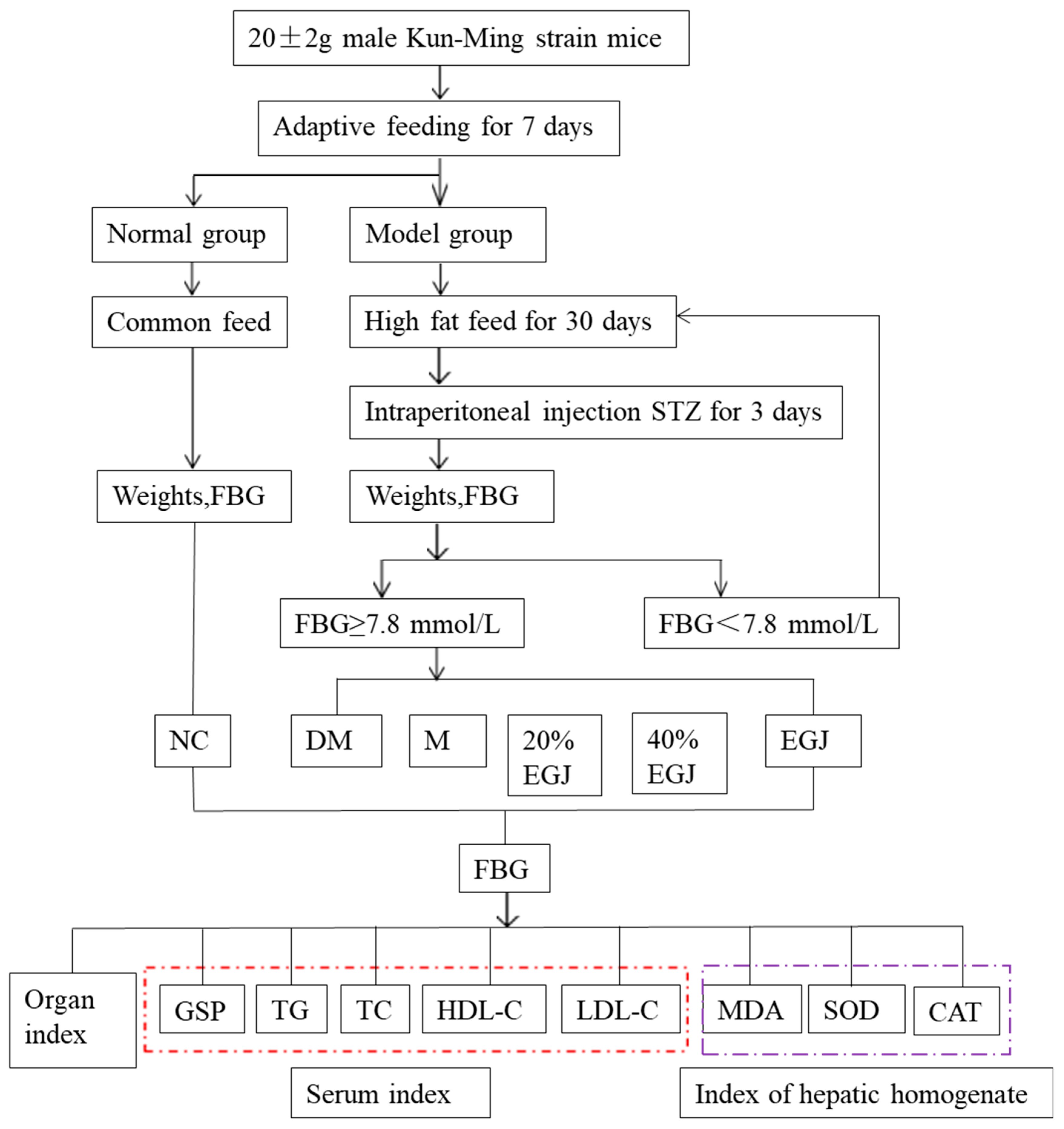
2.4. Animal Experiment
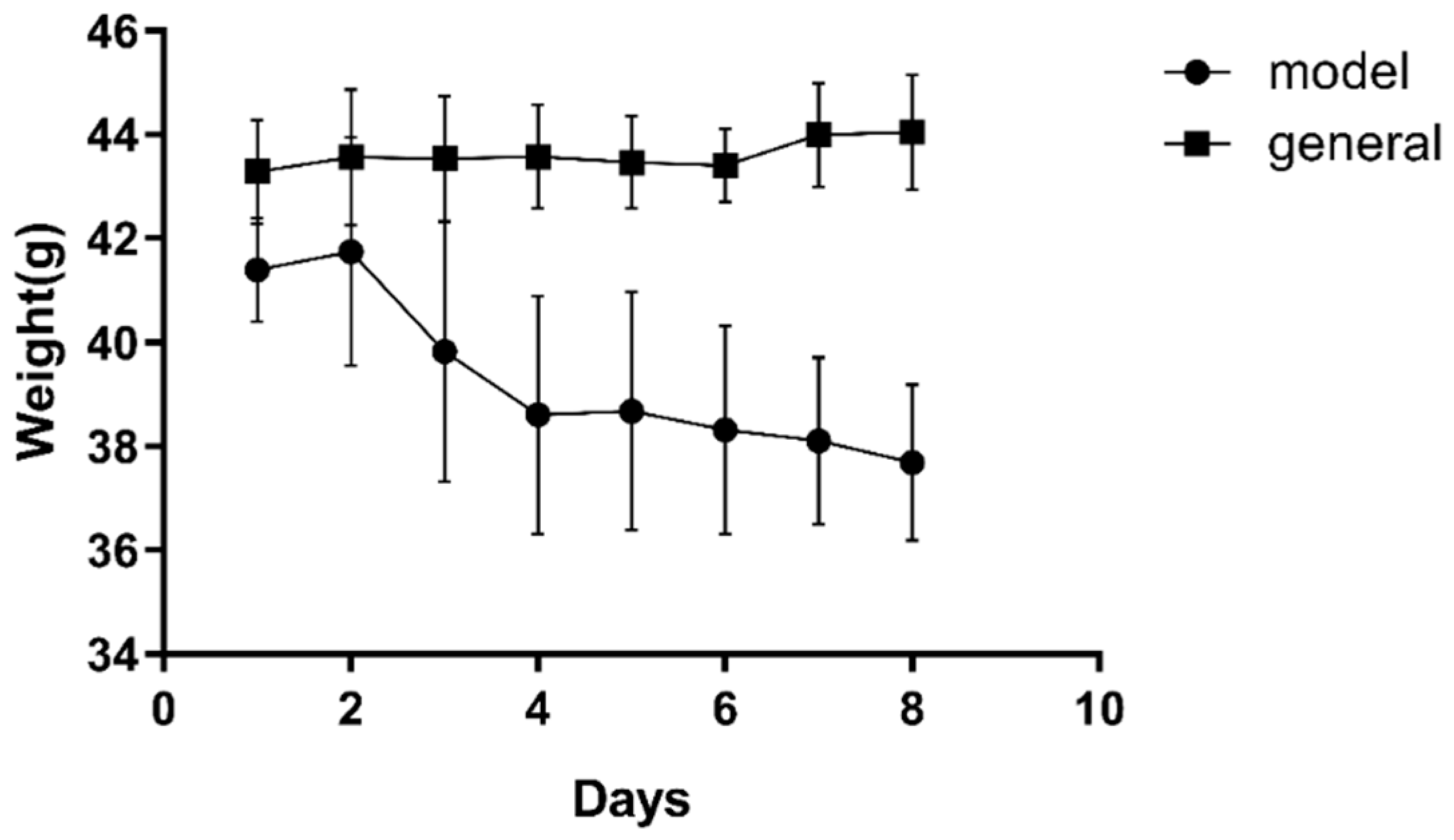
2.4.1. Establishment of the Type 2 Diabetes Mellitus (T2DM) Mouse Model and Drug Administration
2.4.2. Determination of the Body Weight, Food Intake, Water Intake, Blood Glucose, and Oral Glucose Tolerance
2.4.3. Biochemical Assays and the Detection of the Organ Indexes
2.5. Statistical Analysis
3. Results and Discussion
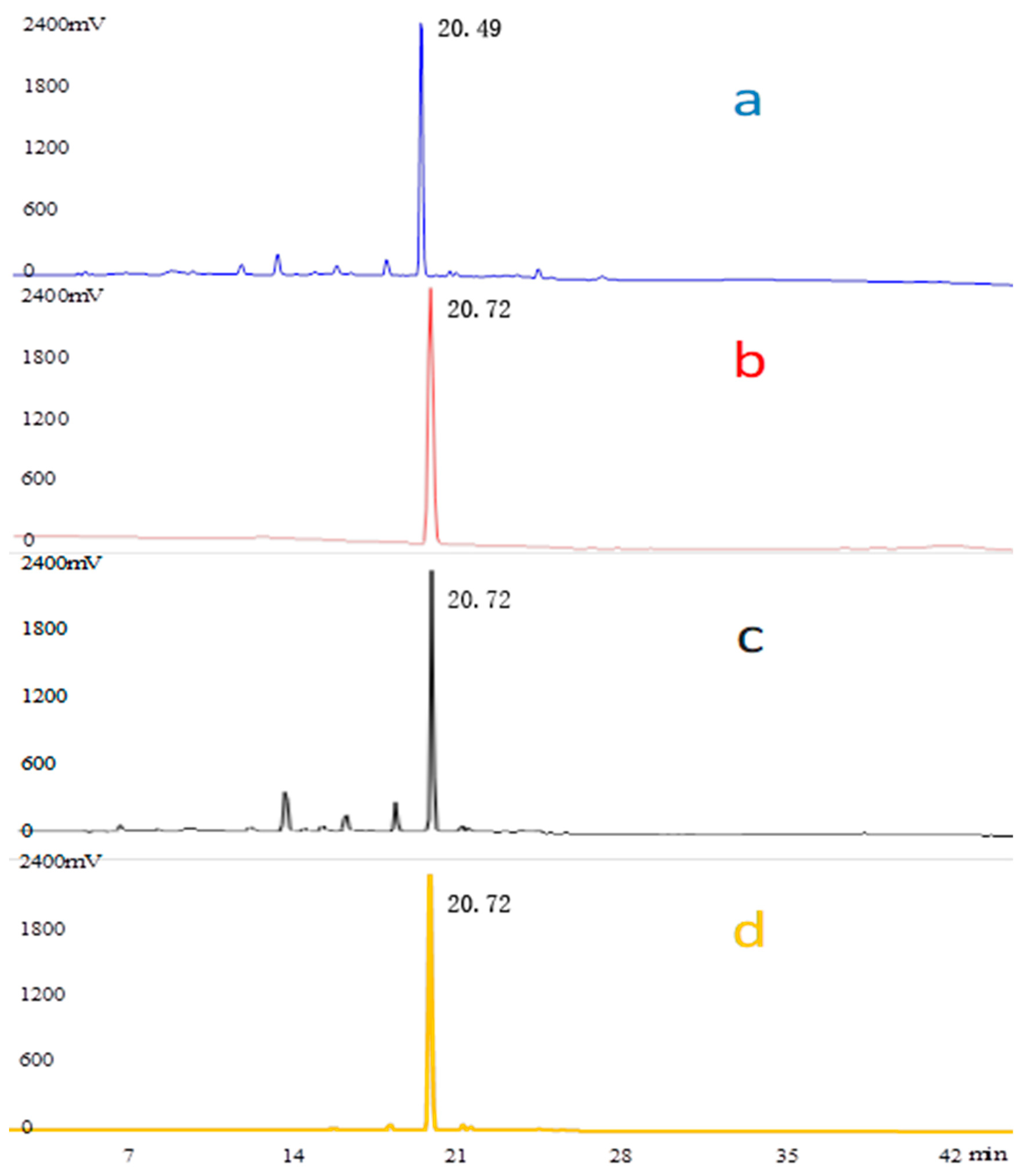
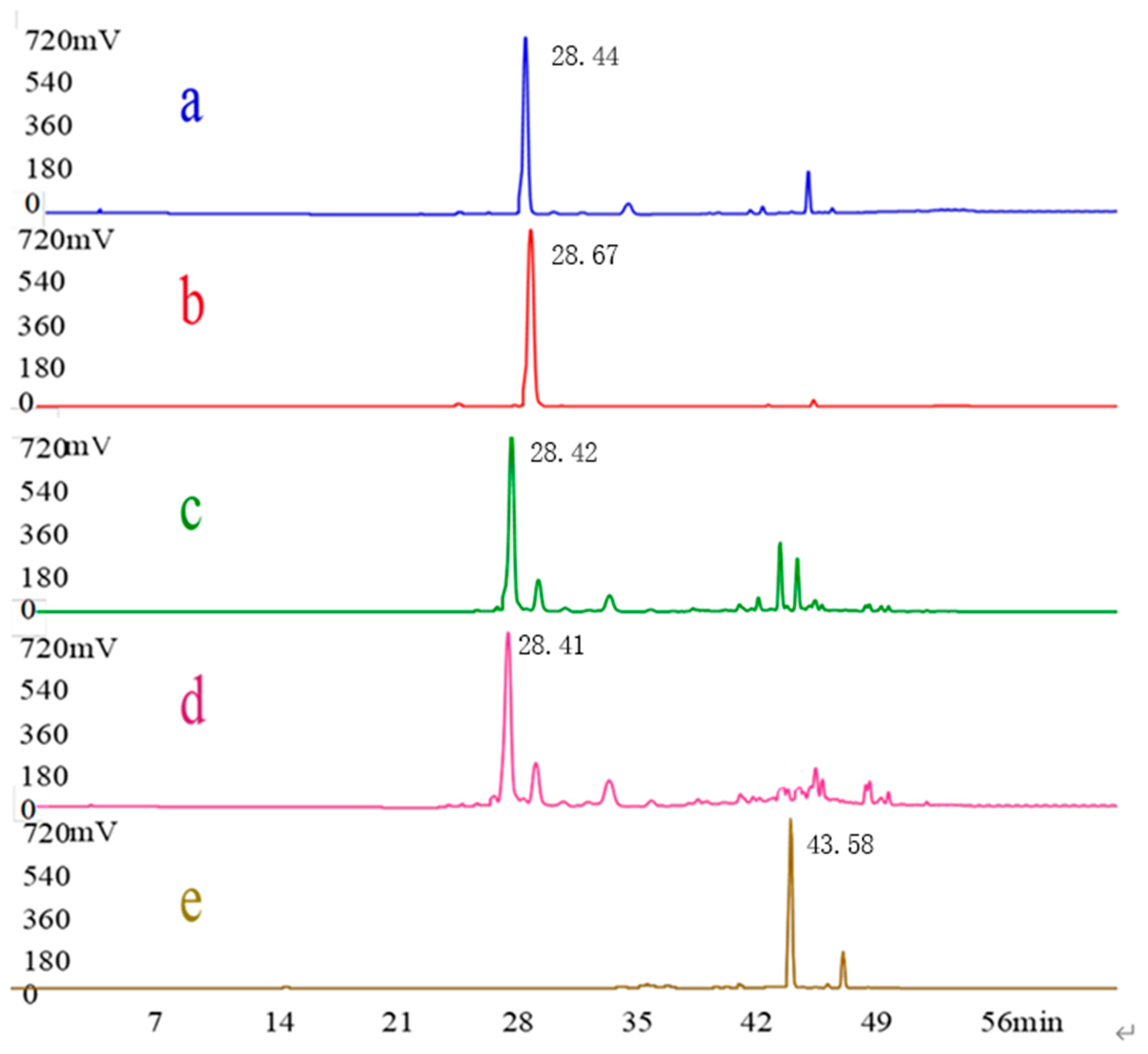
3.1. HPLC Analysis of Geniposide and Crocetin
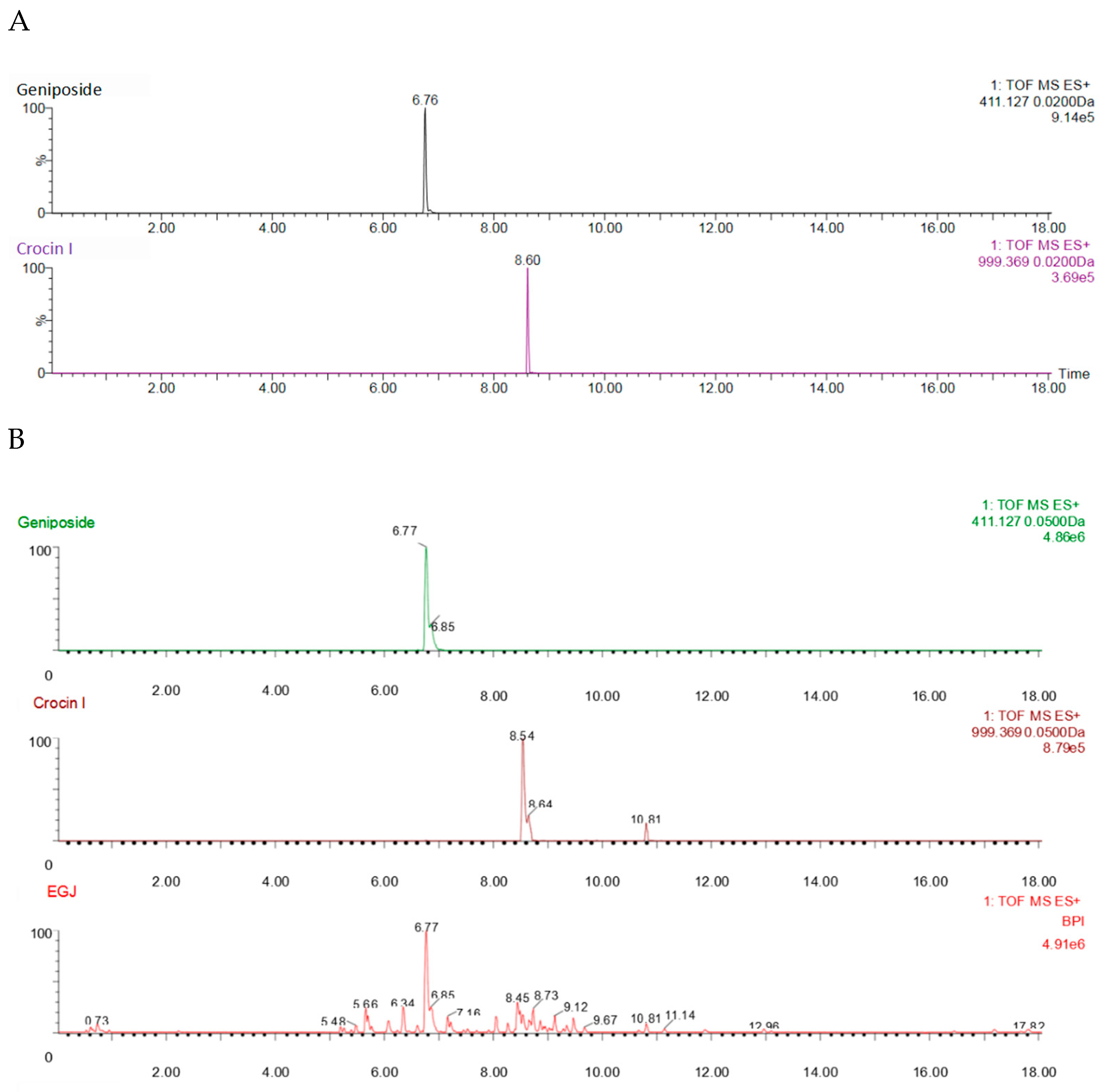
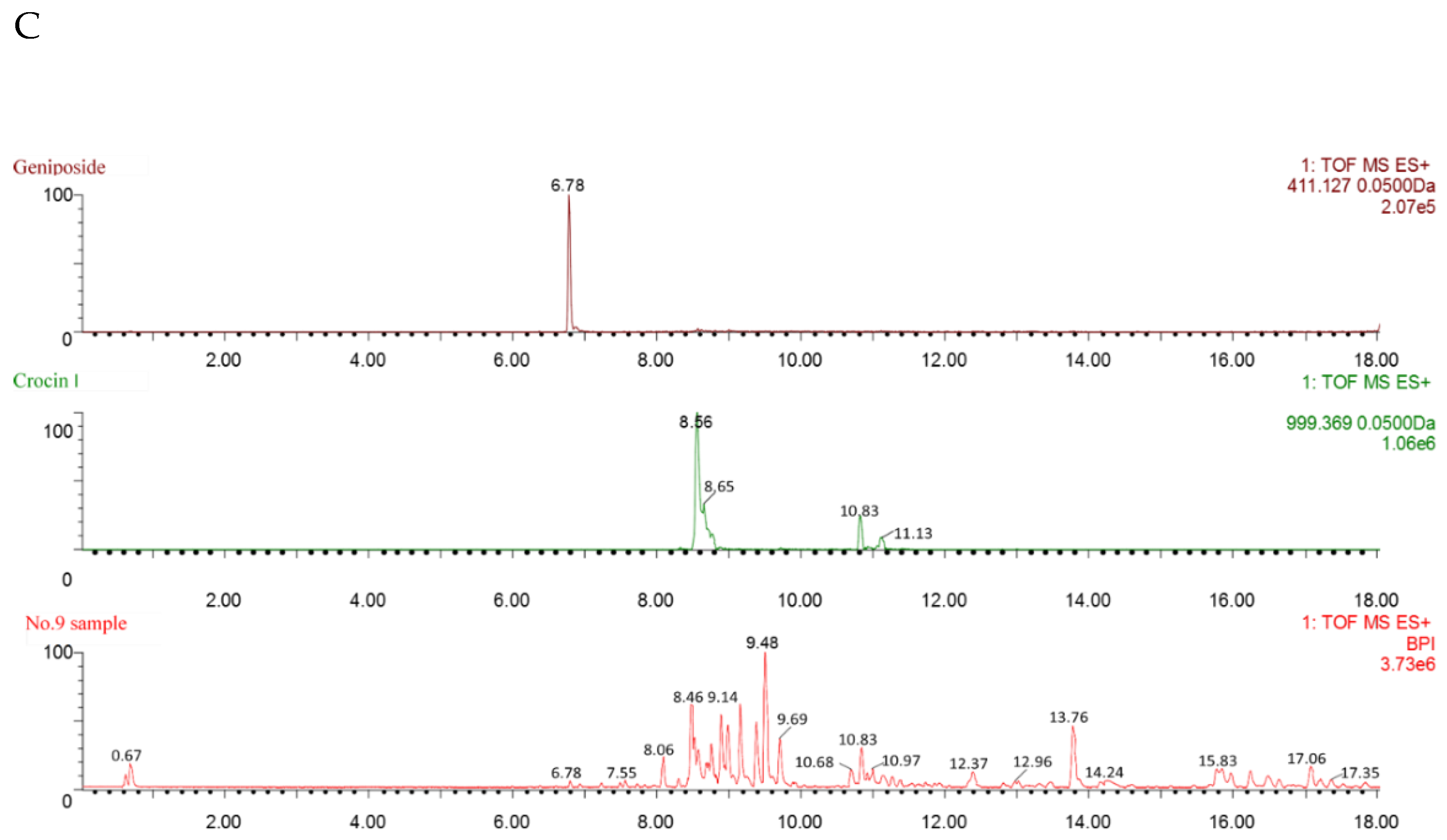
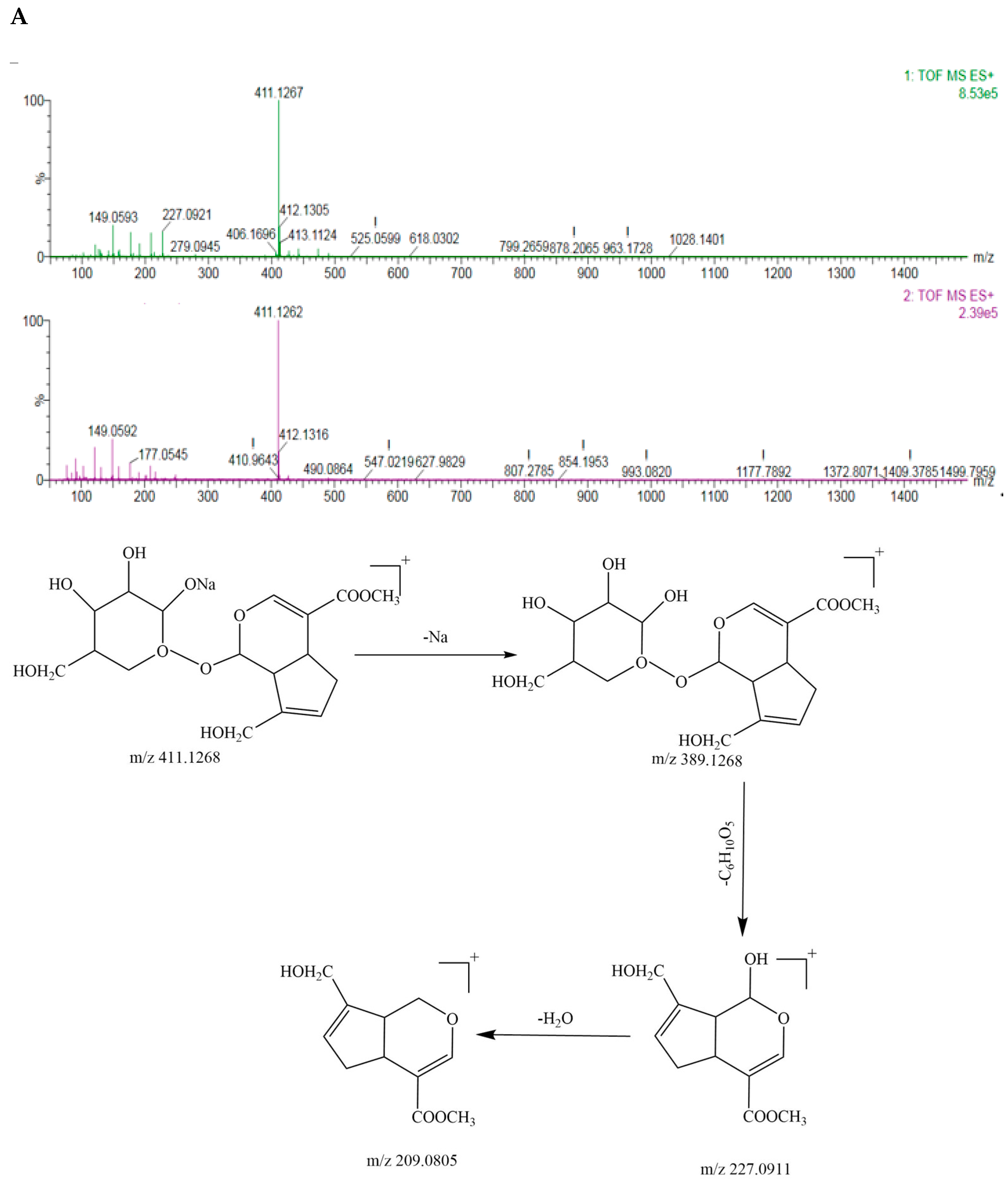
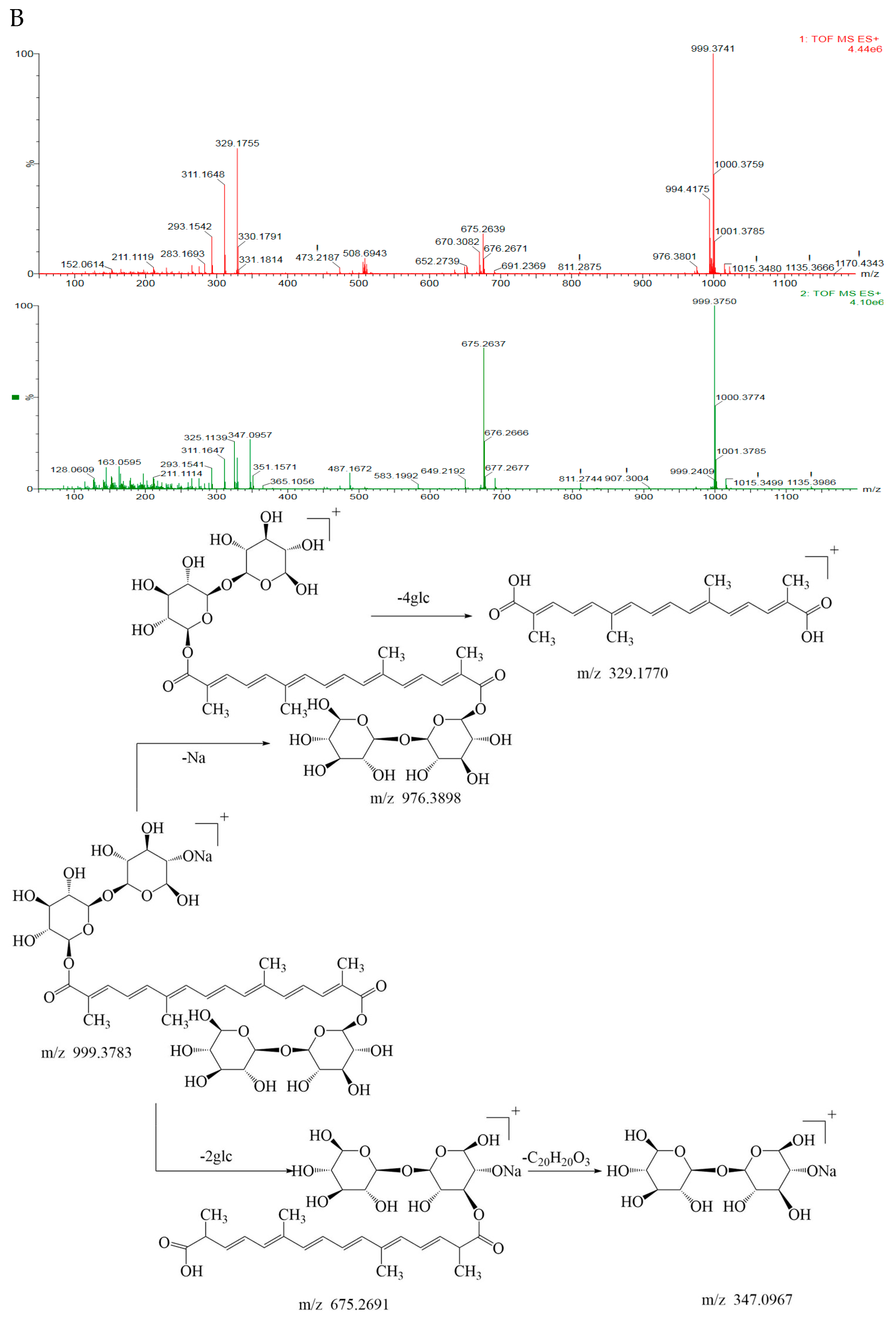
3.2. Identification of Geniposide and Crocetin Using UPLC-ESI-QTOF-MS/MS
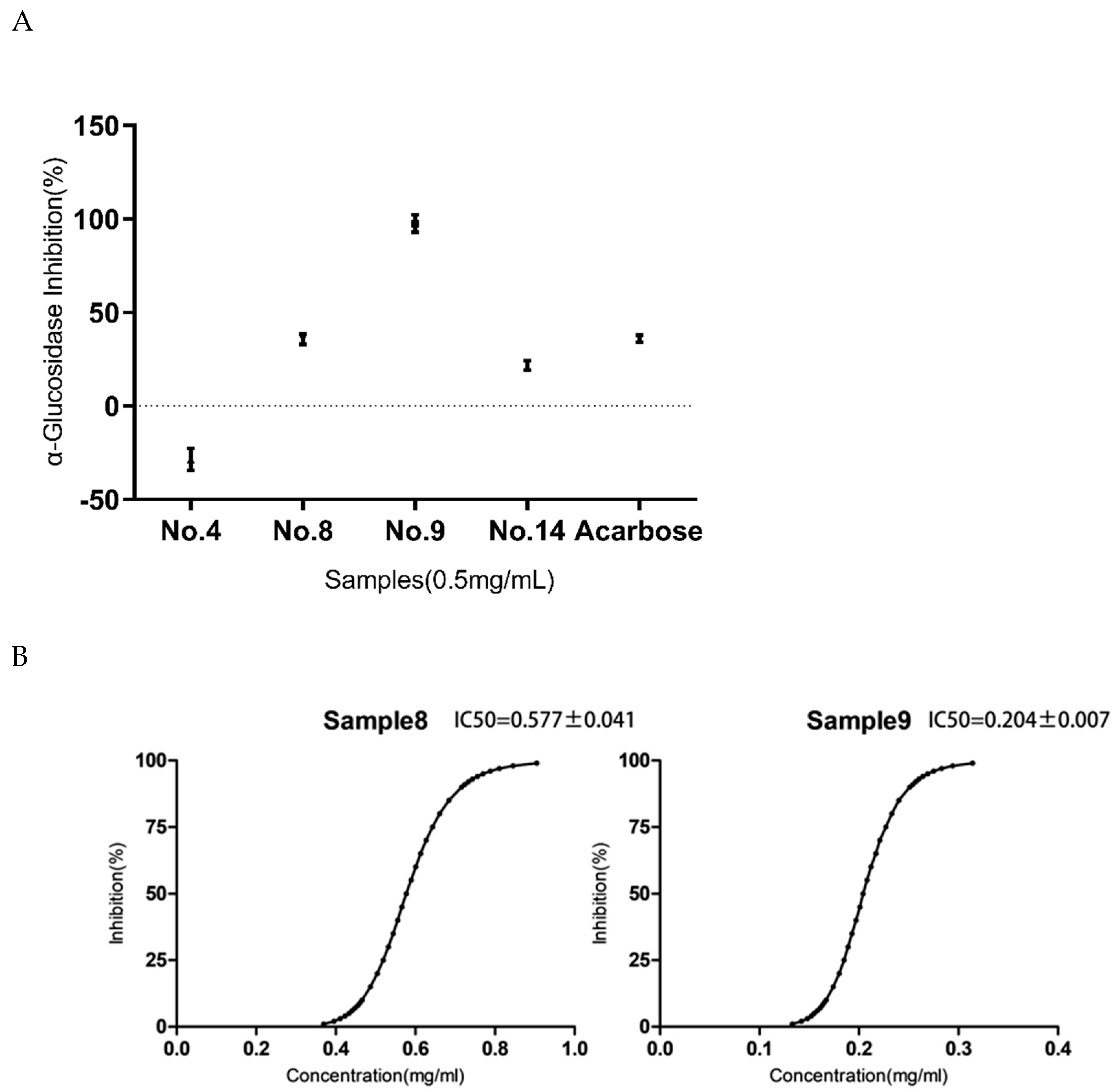
3.3. Screening of Geniposide and Crocetin on the α-Glucosidase Activity In Vitro
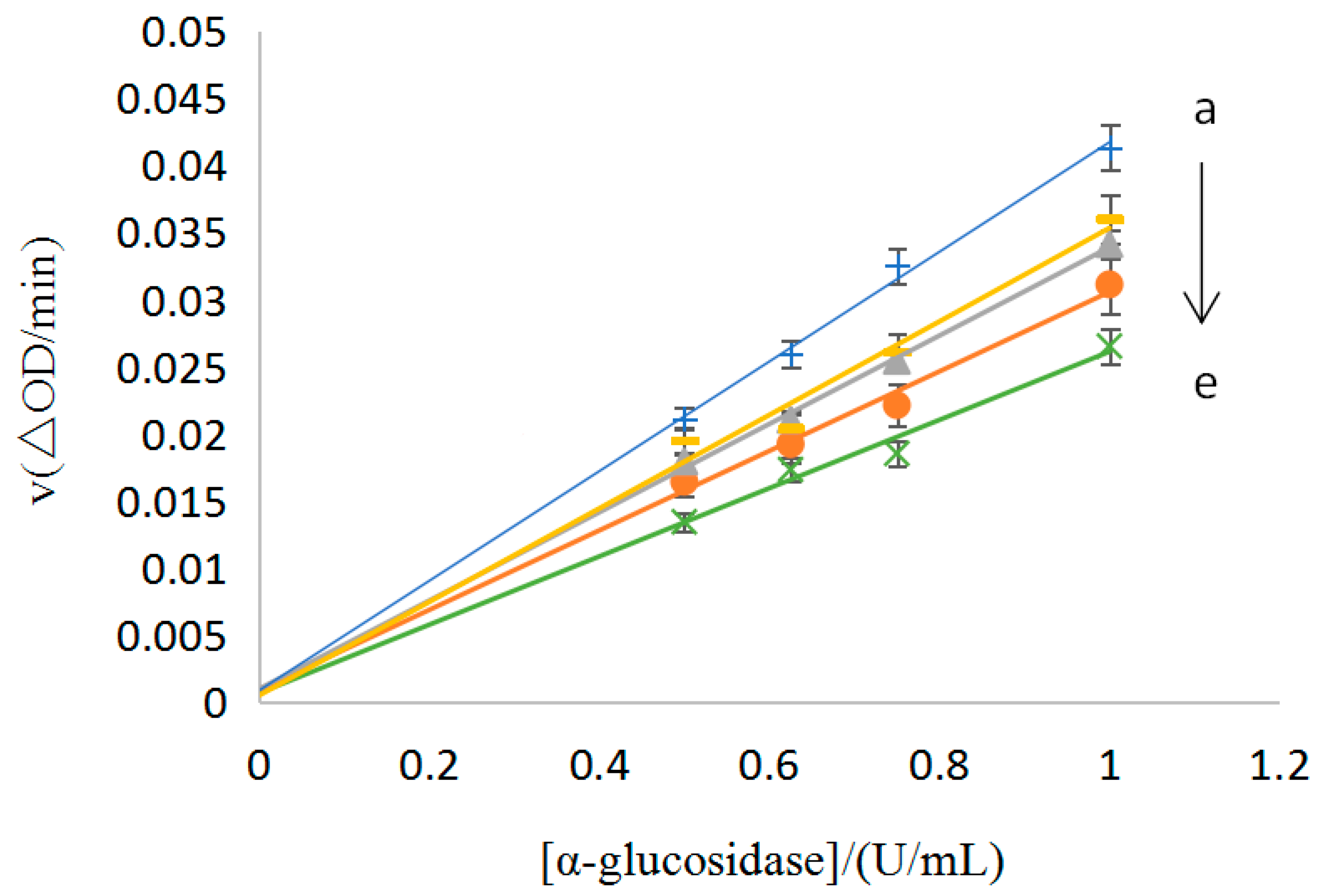
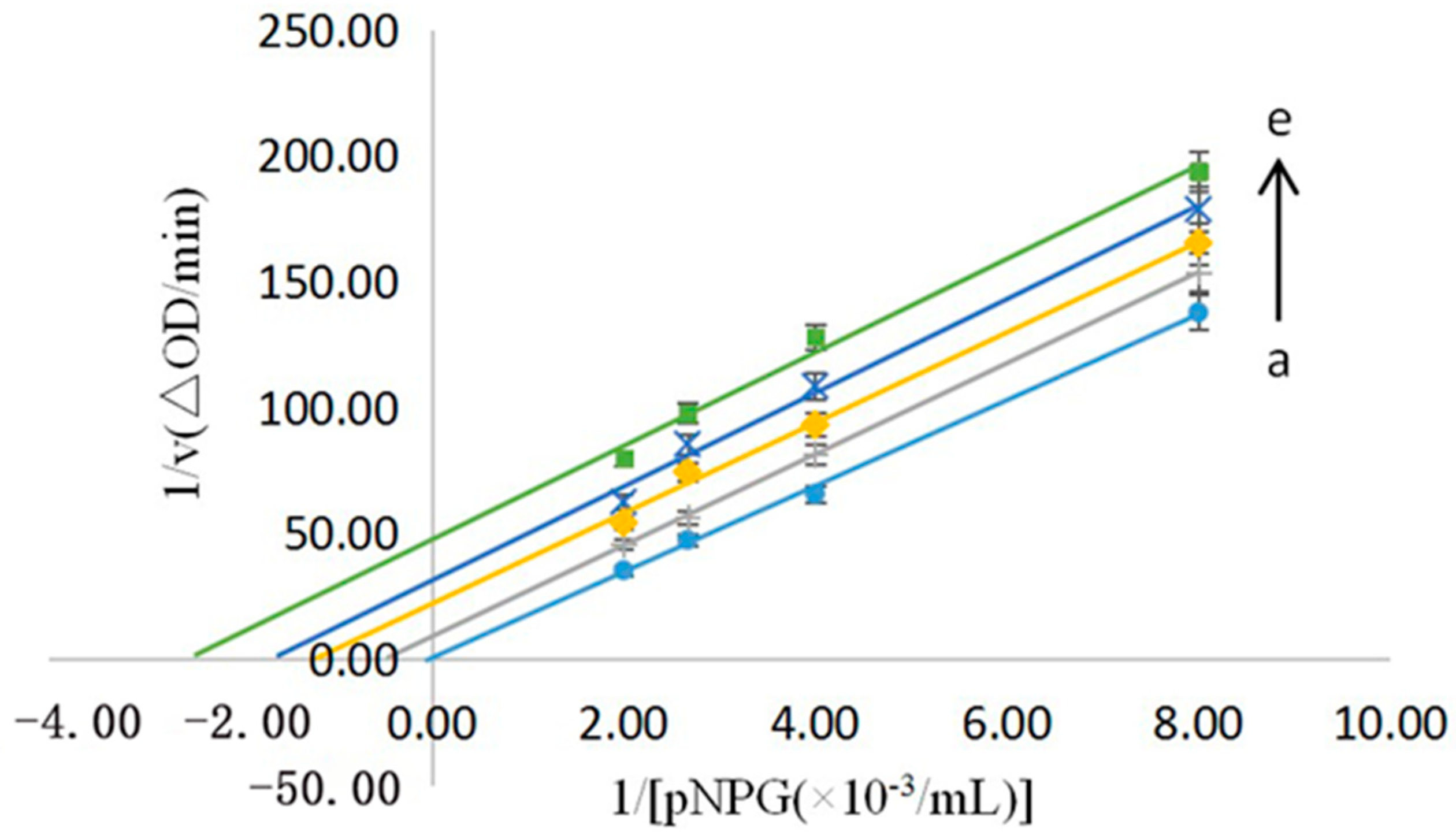
3.3.1. Type of Inhibition for the No.9 Sample
3.3.2. Mechanism of Inhibition for the No.9 Sample
3.4. Effects of the Crude EGJ, 20% EGJ, and 40% EGJ in the DM Male Mice
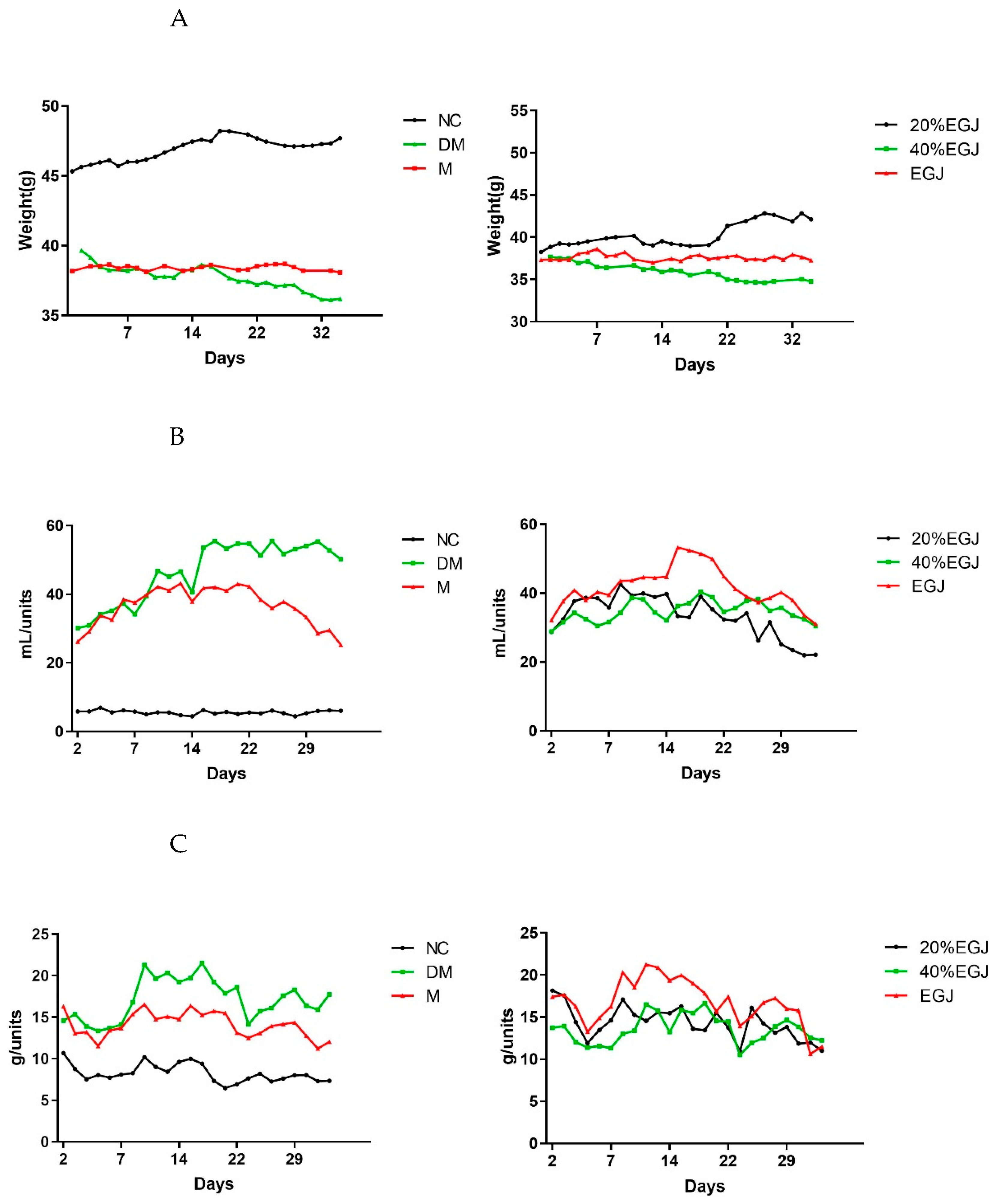
3.4.1. Effects of the Crude EGJ, 20% EGJ, and 40% EGJ on the Fasting Blood Glucose, Body Weight, and Water Intake
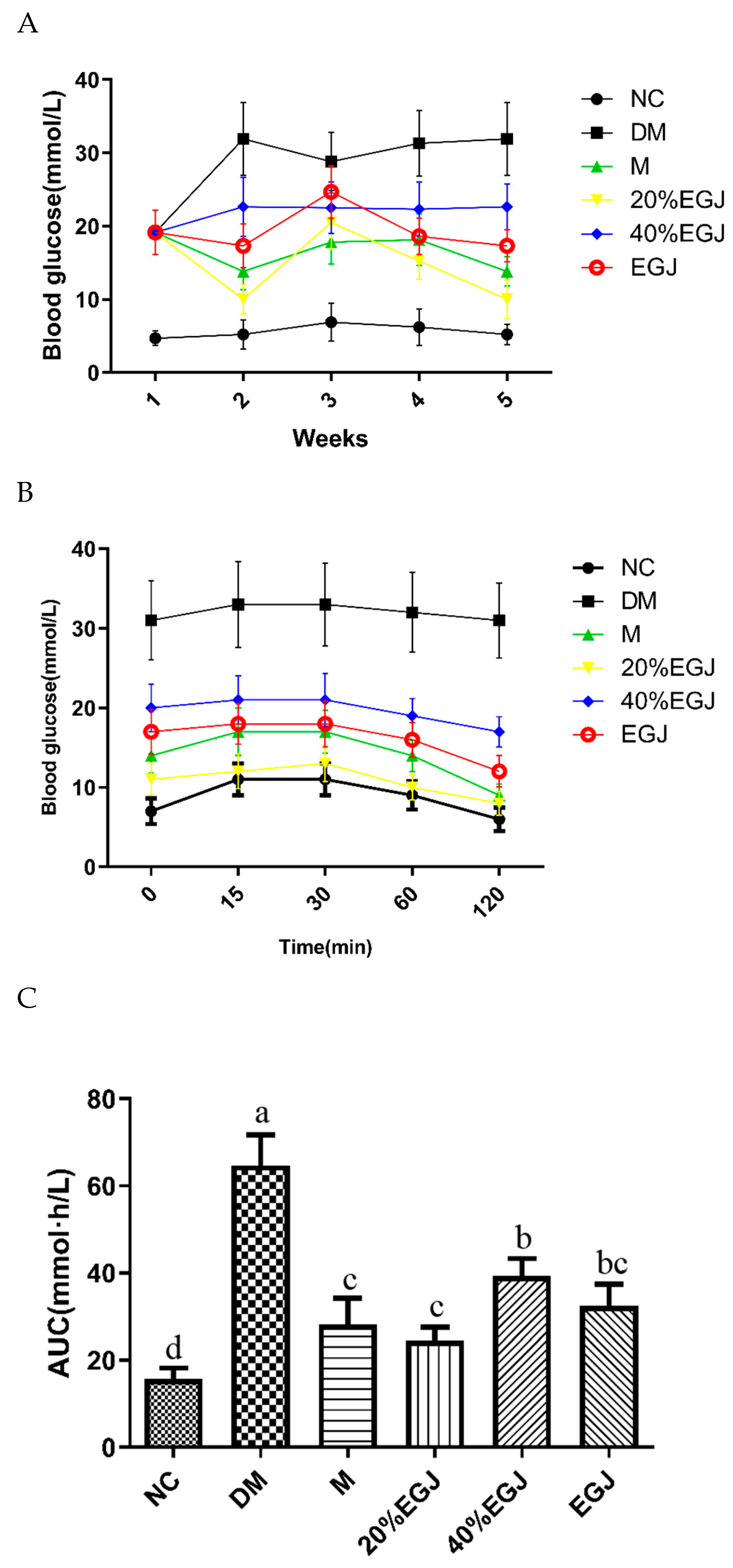
3.4.2. Effects of the Crude EGJ, 20% EGJ, and 40% EGJ on the FBG, OGTT, and AUC
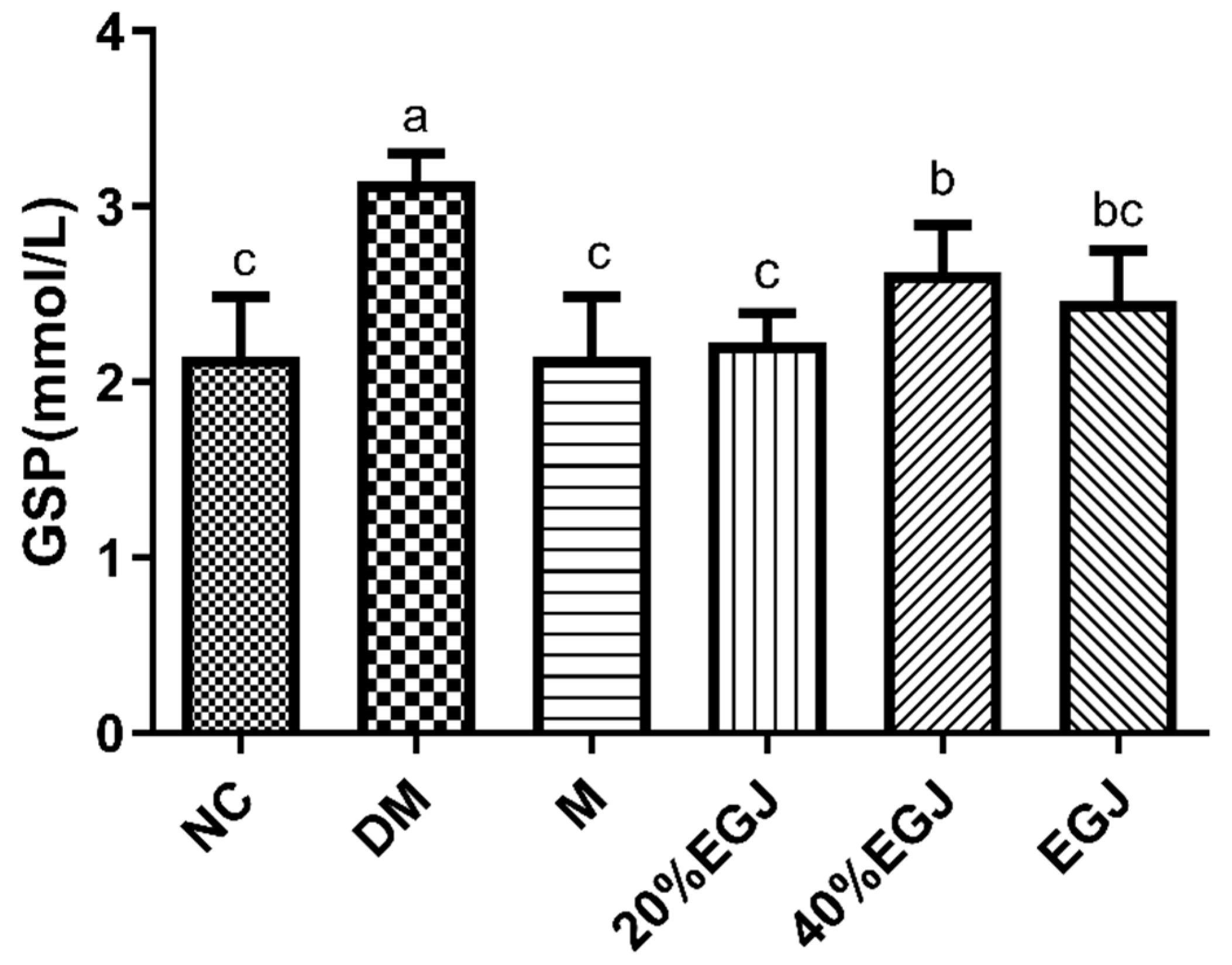
3.4.3. Effects of the Crude EGJ, 20% EGJ, and 40% EGJ on the Organ Indexes and GSP
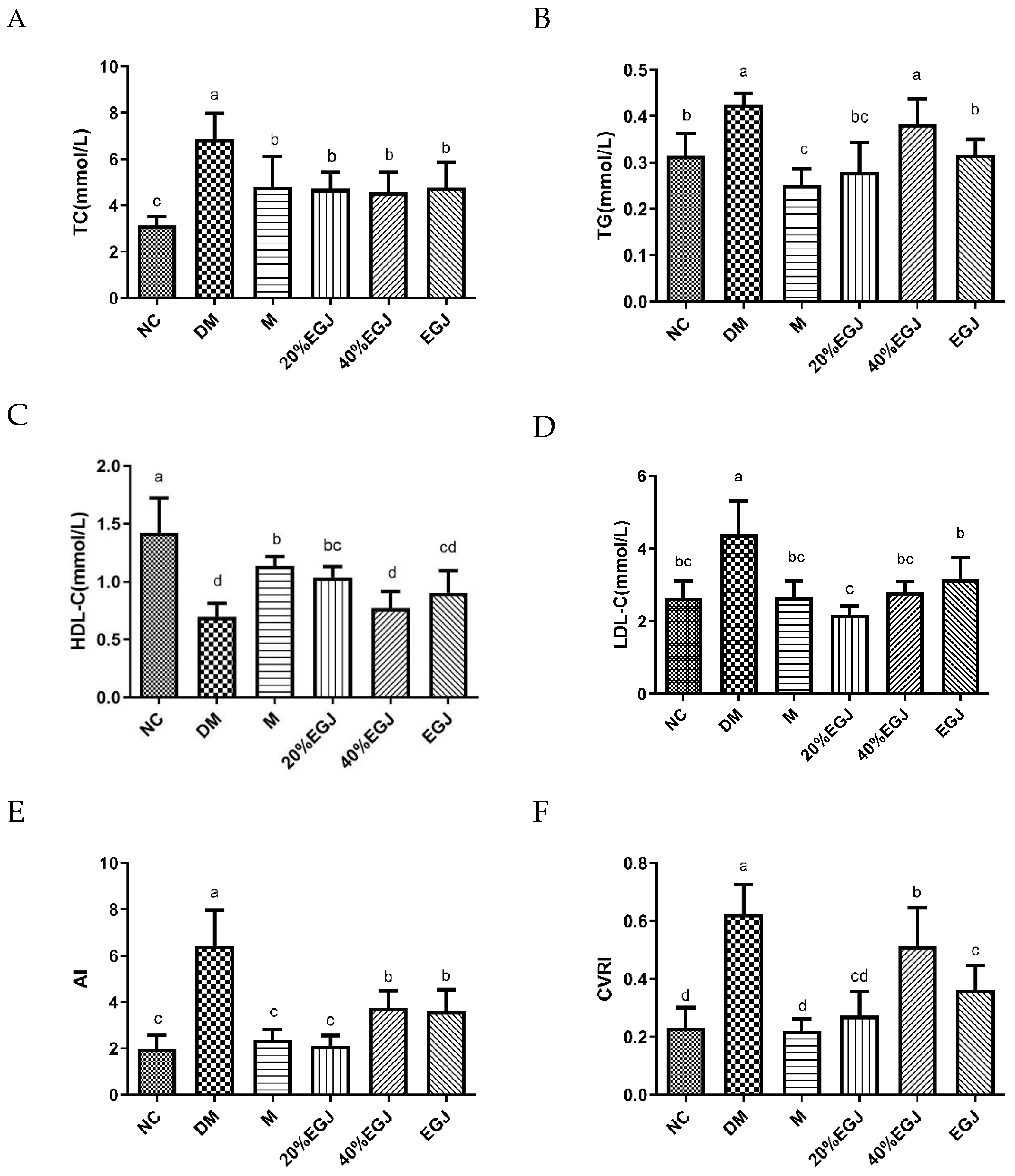
3.4.4. Effects of the Crude EGJ, 20% EGJ, and 40% EGJ on the Serum TC, TG, HDL-C, LDL-C, AI, and CVRI
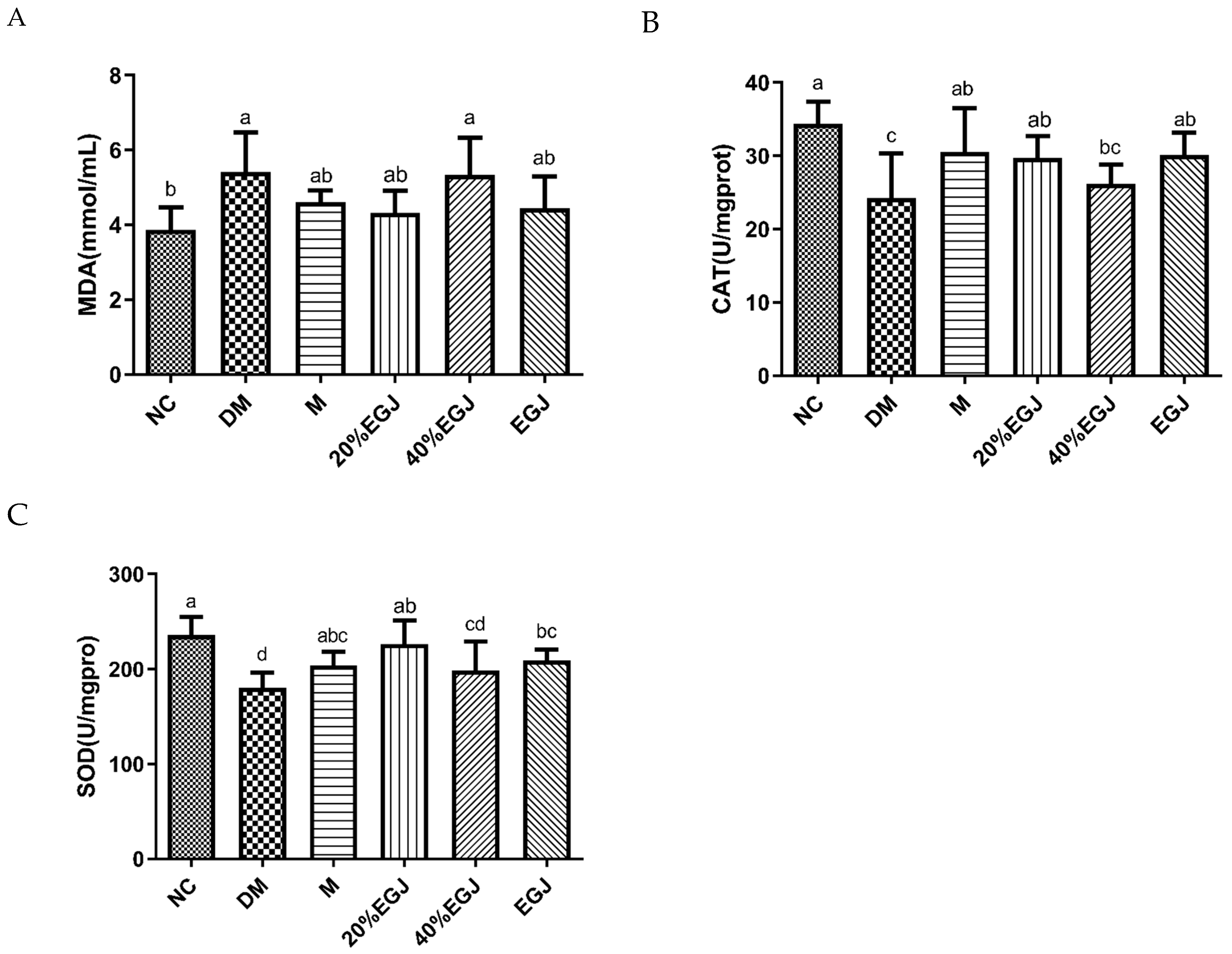
3.4.5. Effects of the Crude EGJ and Its 20% and 40% Fractions on Hepatic MDA, CAT, and SOD
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AI | Atherosclerosis index |
| AUC | Area under the curve |
| CAT | Catalase |
| CVRI | Cardiovascular risk index |
| EGJ | Extract of gardenia (Gardenia jasminoides Ellis) |
| FBG | Fasting blood glucose |
| GSP | Glycosylated serum protein |
| HDL-C | High-density lipoprotein cholesterol |
| LDL-C | Low-density lipoprotein cholesterol |
| MDA | Malondialdehyde |
| OGTT | Oral glucose tolerance test |
| pNPG | The p-nitrophenyl α-D-glucosidase |
| SOD | Superoxide dismutase |
| TC | Total cholesterol |
| TG | Triglyceride |
| T2DM | Type 2 diabetes mellitus |
References
- Scully, T. Diabetes in Numbers. Nature 2012, 485, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Tonyan, Z.N.; Nasykhova, Y.A.; Danilova, M.M.; Glotov, A.S. Genetics of Macrovascular Complications in Type 2 Diabetes. World J. Diabetes 2021, 12, 1200–1219. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T. Immunomodulatory and Antioxidant Effects of Saffron Aqueous Extract (Crocus sativus L.) on Streptozotocin-Induced Diabetes in Rats. Indian Heart J. 2017, 69, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Papademetriou, V.; Lovato, L.; Tsioufis, C.; Cushman, W.; Applegate, W.B.; Mottle, A.; Punthakee, Z.; Nylen, E.; Doumas, M.; Accord Study Group. Effects of High Density Lipoprotein Raising Therapies on Cardiovascular Outcomes in Patients with Type 2 Diabetes Mellitus, with or without Renal Impairment: The Action to Control Cardiovascular Risk in Diabetes Study. Am. J. Nephrol. 2017, 45, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Dash, R.P.; Babu, R.J.; Srinivas, N.R. Reappraisal and Perspectives of Clinical Drug-Drug Interaction Potential of Alpha-Glucosidase Inhibitors Such as Acarbose, Voglibose and Miglitol in the Treatment of Type 2 Diabetes Mellitus. Xenobiotica 2018, 48, 89–108. [Google Scholar] [CrossRef]
- Addy, C.; Tatosian, D.; Glasgow, X.S.; Gendrano, I.N., III; Kauh, E.; Martucci, A.; Johnson-Levonas, A.O.; Selverian, D.; Matthews, C.Z.; Gutierrez, M.; et al. Pharmacokinetic and Pharmacodynamic Effects of Multiple-Dose Administration of Omarigliptin, a Once-Weekly Dipeptidyl Peptidase-4 Inhibitor, in Obese Participants with and without Type 2 Diabetes Mellitus. Clin. Ther. 2016, 38, 516–530. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, Q.; Chen, X.-Y.; Li, X.; Wang, Y.; Zhang, J.-L. Comparison and Screening of Bioactive Phenolic Compounds in Different Blueberry Cultivars: Evaluation of Anti-Oxidation and Alpha-Glucosidase Inhibition Effect. Food Res. Int. 2017, 100 Pt 1, 312–324. [Google Scholar] [CrossRef]
- Abou-Hany, H.O.; Atef, H.; Said, E.; Elkashef, H.A.; Salem, H.A. Crocin Mediated Amelioration of Oxidative Burden and Inflammatory Cascade Suppresses Diabetic Nephropathy Progression in Diabetic Rats. Chem.-Biol. Interact. 2018, 284, 90–100. [Google Scholar] [CrossRef]
- Lee, S.W.; Lim, J.M.; Bhoo, S.H.; Paik, Y.S.; Hahn, T.R. Colorimetric Determination of Amino Acids Using Genipin from Gardenia jasminoides. Anal. Chim. Acta 2003, 480, 267–274. [Google Scholar] [CrossRef]
- Chang, W.L.; Wang, H.Y.; Shi, L.S.; Lai, J.H.; Lin, H.C. Immunosuppressive Iridoids from the Fruits of Gardenia jasminoides. J. Nat. Prod. 2005, 68, 1683–1685. [Google Scholar] [CrossRef]
- Qi, Q.; Mao, Y.; Tian, Y.; Zhu, K.; Cha, X.; Wu, M.; Zhou, X. Geniposide Inhibited Endothelial-Mesenchymal Transition Via the Mtor Signaling Pathway in a Bleomycin-Induced Scleroderma Mouse Model. Am. J. Transl. Res. 2017, 9, 1025–1036. [Google Scholar] [PubMed]
- Lu, W.; Zhao, Y.; Kong, Y.; Zhang, W.; Ma, W.; Li, W.; Wang, K. Geniposide Prevents H2O2-Induced Oxidative Damage in Melanocytes by Activating the Pi3k-Akt Signalling Pathway. Clin. Exp. Dermatol. 2018, 43, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.; Hsu, C.-Y.; Chen, C.-Y.; Liu, H.-K. Fructus Corni Suppresses Hepatic Gluconeogenesis Related Gene Transcription, Enhances Glucose Responsiveness of Pancreatic Beta-Cells, and Prevents Toxin Induced Beta-Cell Death. J. Ethnopharmacol. 2008, 117, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-I.; Cheng, Y.-W.; Tzeng, C.-Y.; Lee, Y.-C.; Chang, Y.-N.; Lee, S.-C.; Tsai, C.-C.; Chen, J.-C.; Tzen, J.T.-C.; Chang, S.-L. Peroxisome Proliferator-Activated Receptor Activating Hypoglycemic Effect of Gardenia jasminoides Ellis Aqueous Extract and Improvement of Insulin Sensitivity in Steroid Induced Insulin Resistant Rats. BMC Complement. Altern. Med. 2014, 14, 30. [Google Scholar] [CrossRef]
- Li, S.; Liu, X.; Lei, J.; Yang, J.; Tian, P.; Gao, Y. Crocin Protects Podocytes against Oxidative Stress and Inflammation Induced by High Glucose through Inhibition of Nf-Kappa B. Cell. Physiol. Biochem. 2017, 42, 1481–1492. [Google Scholar] [CrossRef]
- Yin, F.; Liu, J.-H. Research and Application Progress of Gardenia jasminoides. Chin. Herb. Med. 2018, 10, 362–370. [Google Scholar] [CrossRef]
- Chen, T.; Kang, B.; Ren, L.; Chen, S.; Lin, H. The Optimized Parameters of Extracting Yellow Pigment from Gardenia (Gardenia jasminoides Ellis) Fruit. Chin. J. Trop. Crops 2010, 31, 1398–1402. [Google Scholar]
- Alshabi, A.M.; Shaikh, I.A. Antidiabetic and Antioxidant Potential of Gardenia latifolia in Type-2 Diabetic Rats Fed with High-Fat Diet Plus Low-Dose Streptozotocin. Saudi Med. J. 2022, 43, 881–890. [Google Scholar] [CrossRef]
- Zhang, J.-L.; Luo, C.-L.; Zhou, Q.; Zhang, Z.-C. Isolation and Identification of Two Major Acylated Anthocyanins from Purple Sweet Potato (Ipomoea batatas L. Cultivar Eshu No. 8) by Uplc-Qtof-Ms/Ms and Nmr. Int. J. Food Sci. Technol. 2018, 53, 1932–1941. [Google Scholar] [CrossRef]
- Luo, C.-L.; Zhou, Q.; Yang, Z.-W.; Wang, R.-D.; Zhang, J.-L. Evaluation of Structure and Bioprotective Activity of Key High Molecular Weight Acylated Anthocyanin Compounds Isolated from the Purple Sweet Potato (Ipomoea batatas L. Cultivar Eshu No.8). Food Chem. 2018, 241, 23–31. [Google Scholar] [CrossRef]
- Moras, B.; Loffredo, L.; Rey, S. Quality Assessment of Saffron (Crocus sativus L.) Extracts Via Uhplc-Dad-Ms Analysis and Detection of Adulteration Using Gardenia Fruit Extract (Gardenia jasminoides Ellis). Food Chem. 2018, 257, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Mojica, L.; Meyer, A.; Berhow, M.A.; de Mejia, E.G. Bean Cultivars (Phaseolus vulgaris L.) Have Similar High Antioxidant Capacity, in Vitro Inhibition of Alpha-Amylase and Alpha-Glucosidase While Diverse Phenolic Composition and Concentration. Food Res. Int. 2015, 69, 38–48. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Erejuwa, O.O.; Nwobodo, N.N.; Akpan, J.L.; Okorie, U.A.; Ezeonu, C.T.; Ezeokpo, B.C.; Nwadike, K.I.; Erhiano, E.; Wahab, M.S.A.; Sulaiman, S.A. Nigerian Honey Ameliorates Hyperglycemia and Dyslipidemia in Alloxan-Induced Diabetic Rats. Nutrients 2016, 8, 95. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, J.-L.; Shen, L.-H.; Feng, L.-J.; Zhou, Q. Inhibition Mechanism of Diacylated Anthocyanins from Purple Sweet Potato (Ipomoea batatas L.) against Alpha-Amylase and Alpha-Glucosidase. Food Chem. 2021, 359, 129934. [Google Scholar] [CrossRef]
- Zhang, G.-S.; Yang, H.-D.; Yue, X.-X.; Liu, Z.-X.; Xu, C.-M. Study on Enzymatic Combined Chemical Demulsification Process of Emulsion from Enzyme-Assisted Aqueous Extraction of Pumpkin Seed Oil. Food Mach. 2018, 10, 139–178. [Google Scholar]
- Hirano, T.; Yamashita, S.; Takahashi, M.; Hashimoto, H.; Mori, Y.; Goto, M. Anagliptin, a Dipeptidyl Peptidase-4 Inhibitor, Decreases Macrophage Infiltration and Suppresses Atherosclerosis in Aortic and Coronary Arteries in Cholesterol-Fed Rabbits. Metab.-Clin. Exp. 2016, 65, 893–903. [Google Scholar] [CrossRef]
- Vo, A.Q.; Feng, X.; Morott, J.T.; Pimparade, M.B.; Tiwari, R.V.; Zhang, F.; Repka, M.A. A Novel Floating Controlled Release Drug Delivery System Prepared by Hot-Melt Extrusion. Eur. J. Pharm. Biopharm. 2016, 98, 108–121. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, J.; Yu, J.; Chen, X.; Zhang, S.; Cai, Y.; Li, L. A New Functionality Study of Vanillin as the Inhibitor for Alpha-Glucosidase and Its Inhibition Kinetic Mechanism. Food Chem. 2021, 353, 129448. [Google Scholar] [CrossRef]
- Tan, S.Y.; Wong, J.L.M.; Sim, Y.J.; Wong, S.S.; Elhassan, S.A.M.; Tan, S.H.; Lim, G.P.L.; Tay, N.W.R.; Annan, N.C.; Bhattamisra, S.K.; et al. Type 1 and 2 Diabetes Mellitus: A Review on Current Treatment Approach and Gene Therapy as Potential Intervention. Diabetes Metab. Syndr. 2019, 13, 364–372. [Google Scholar] [CrossRef]
- Cakar, U.; Grozdanic, N.; Pejin, B.; Vasic, V.; Cakar, M.; Petrovic, A.; Djordjevic, B. Impact of Vinification Procedure on Fruit Wine Inhibitory Activity against Alpha-Glucosidase. Food Biosci. 2018, 25, 1–7. [Google Scholar] [CrossRef]
- Cakar, U.; Grozdanic, N.; Petrovic, A.; Pejin, B.; Nastasijevic, B.; Markovic, B.; Dordevic, B. Fruit Wines Inhibitory Activity against Alpha-Glucosidase. Curr. Pharm. Biotechnol. 2017, 18, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Wan, Y.; Li, L.; Pan, T. Studies on the Inhibition Kinetics and Interaction Mechanism of Gardenia Yellow on Starch Digestive Enzyme. J. Chin. Inst. Food Sci. Technol. 2021, 21, 38–47. [Google Scholar]
- Wu, S.-Y.; Wang, G.-F.; Liu, Z.-Q.; Rao, J.-J.; Lue, L.; Xu, W.; Wu, S.-G.; Zhang, J.-J. Effect of Geniposide, a Hypoglycemic Glucoside, on Hepatic Regulating Enzymes in Diabetic Mice Induced by a High-Fat Diet and Streptozotocin. Acta Pharmacol. Sin. 2009, 30, 202–208. [Google Scholar] [CrossRef]
- Jiang, X.-Q.; Shen, S.-L.; Li, W.-Z.; Xu, X.-K.; Yin, F. Molecular Mechanism of Geniposide in Regulating Glut2 Glycosylation in Pancreatic Beta Cells. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Med. 2021, 46, 3643–3649. [Google Scholar]
- Gao, S.; Feng, Q. The Beneficial Effects of Geniposide on Glucose and Lipid Metabolism: A Review. Drug Des. Dev. Ther. 2022, 16, 3365–3383. [Google Scholar] [CrossRef] [PubMed]
- Khorasani, M.K.; Ahangarpour, A.; Khorsandi, L. Effects of Crocin and Metformin on Methylglyoxal-Induced Reproductive System Dysfunction in Diabetic Male Mice. Clin. Exp. Reprod. Med. 2021, 48, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Gu, M. Crocin Improves Insulin Sensitivity and Ameliorates Adiposity by Regulating Ampk-Cdk5-Ppar Gamma Signaling. Biomed Res. Int. 2020, 2020, 9136282. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt/min | Proposed Formula | Relative Molecular Mass/Da | [M+H]+ | Fragment Ions | ppm | Tentative Identification | Crude EGJ | No.9 Sample | |
|---|---|---|---|---|---|---|---|---|---|---|
| Measured Value | Theoretical Value | ESI (+) | ||||||||
| 1 | 5.265 | C16H24O11 | 392.1385 | 393.1385 | 393.1397 | 415.1232[M+Na] | −1.2 | Shanzhiside | + | − |
| 431.0829[M+K] | ||||||||||
| 397.1082[M+Na-H2O] | ||||||||||
| 2 | 5.481 | C17H24O11 | 404.1319 | 375.1460[M+H-H2O] | 6-alpha-Hydroxy geniposide | + | − | |||
| 427.1217[M+Na] | ||||||||||
| 443.0999[M+K] | ||||||||||
| 409.2032[M+Na-H2O] | ||||||||||
| 3 | 5.666 | C17H24O11 | 404.1319 | 405.1559 | 405.1563 | 427.1218[M+Na] | −0.4 | Feretoside | + | − |
| 443.0805[M+K] | ||||||||||
| 409.1115[M+Na-H2O] | ||||||||||
| 4 | 5.760 | C17H24O11 | 404.1319 | 405.1554 | 405.1563 | 427.1212[M+Na] | −0.9 | Feretoside | + | − |
| 443.1036[M+K] | ||||||||||
| 5 | 6.034 | C16H18O9 | 354.0951 | 355.1003 | 355.1029 | 731.1856[2M+Na] | −2.6 | Chlorogenic acid | + | − |
| 393.0562[M+K] | ||||||||||
| 377.0871[M+Na] | ||||||||||
| 163.0306[M+H-QA] | ||||||||||
| 135.0393[M+H-QA-CO] | ||||||||||
| 6 | 6.339 | C23H34O15 | 550.1898 | 551.1967 | 551.1976 | 573.1804[M+Na] | −0.9 | Genipin-1-O-β-D-gentian glycosides | + | − |
| 589.1512[M+K] | ||||||||||
| 389.1396[M+H-glc] | ||||||||||
| 227.0854[M+H-2glc] | ||||||||||
| 7 | 6.817 | C17H24O10 | 388.1369 | 411.1262[M+Na] | Geniposide | + | + | |||
| 777.2932[M+2Na] | ||||||||||
| 8 | 8.278 | C27H30O14 | 578.1636 | 579.3737 | 579.3714 | 271.0556[M-rha-glc] | 2.3 | Apigenin-7-O-rutinoside | + | + |
| 9 | 8.494 | C32H40O17 | 696.2265 | 697.2177 | 697.2144 | 719.2186[M+Na] | 3.3 | 6”-O-P-coumaroyl genipingentiobioside | + | + |
| 735.1713[M+K] | ||||||||||
| 309.0976[M+H-gen] | ||||||||||
| 10 | 8.668 | C44H64O24 | 976.3788 | 977.3801 | 977.3786 | 999.3741[M+Na] | 1.5 | Corcin I | + | + |
| 675.2639[M+Na-2glc] | ||||||||||
| 347.0957[M-2glc-C20H20O3] | ||||||||||
| 329.1755[M-4glc] | ||||||||||
| 11 | 9.031 | C38H54O19 | 814.3174 | 815.3259 | 815.3255 | 675.2301[M+Na-2glc] | 0.4 | Crocin II | + | + |
| 347.0930[M-2glc-C20H20O3] | ||||||||||
| 329.1750[M-4glc] | ||||||||||
| 12 | 9.110 | C22H22O11 | 462.1162 | 463.1225 | 463.124 | 947.2373[2M+Na] | −1.5 | 2-O-Cinnamoyl-glucogallin | + | + |
| 485.2435[M+Na] | ||||||||||
| 13 | 11.134 | C21H18O11 | 446.0849 | 447.0956 | 447.0927 | 469.1128[M+Na] | 2.9 | Baicalin | + | + |
| 271.0462[M-glc] | ||||||||||
| 14 | 12.374 | C21H21NO4 | 351.1471 | 352.2179 | 352.1849 | 337.1254[M-CH3] | 33 | Palmatine | − | + |
| 308.1244[M-CH3-H-CO] | ||||||||||
| Group | Liver (g) | Kidney (g) | Liver Index (%) | Kidney Index (%) |
|---|---|---|---|---|
| NC | 1.533 ± 0.266 b | 0.553 ± 0.0482 b | 3.166 ± 0.546 d | 1.141 ± 0.099 d |
| DM | 1.780 ± 0.327 a | 0.642 ± 0.058 a | 5.021 ± 0.932 a | 1.812 ± 0.134 a |
| M | 1.493 ± 0.156 b | 0.513 ± 0.074 b | 3.6137 ± 0.571 cd | 1.304 ± 0.311 cd |
| 20%EGJ | 1.674 ± 0.279 ab | 0.573 ± 0.097 b | 3.876 ± 0.649 bc | 1.326 ± 0.226 bc |
| 40%EGJ | 1.585 ± 0.369 ab | 0.556 ± 0.116 b | 4.259 ± 0.9931 b | 1.494 ± 0.401 b |
| EGJ | 1.496 ± 0.2632 b | 0.531 ± 0.0301 b | 4.187 ± 0.7367 bc | 1.485 ± 0.089 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Zhang, S.; Chen, L.; Liu, Y.; Shen, L.; Zhang, J. Effective Therapeutic Verification of Crocin I, Geniposide, and Gardenia (Gardenia jasminoides Ellis) on Type 2 Diabetes Mellitus In Vivo and In Vitro. Foods 2023, 12, 1668. https://doi.org/10.3390/foods12081668
Zhou H, Zhang S, Chen L, Liu Y, Shen L, Zhang J. Effective Therapeutic Verification of Crocin I, Geniposide, and Gardenia (Gardenia jasminoides Ellis) on Type 2 Diabetes Mellitus In Vivo and In Vitro. Foods. 2023; 12(8):1668. https://doi.org/10.3390/foods12081668
Chicago/Turabian StyleZhou, Haibo, Sen Zhang, Lianghua Chen, Yimei Liu, Luhong Shen, and Jiuliang Zhang. 2023. "Effective Therapeutic Verification of Crocin I, Geniposide, and Gardenia (Gardenia jasminoides Ellis) on Type 2 Diabetes Mellitus In Vivo and In Vitro" Foods 12, no. 8: 1668. https://doi.org/10.3390/foods12081668
APA StyleZhou, H., Zhang, S., Chen, L., Liu, Y., Shen, L., & Zhang, J. (2023). Effective Therapeutic Verification of Crocin I, Geniposide, and Gardenia (Gardenia jasminoides Ellis) on Type 2 Diabetes Mellitus In Vivo and In Vitro. Foods, 12(8), 1668. https://doi.org/10.3390/foods12081668