
Transforming Spent Coffee Grounds’ Hydrolysates with Yeast Lachancea thermotolerans and Lactic Acid Bacterium Lactiplantibacillus plantarum to Develop Potential Novel Alcoholic Beverages



Abstract
1. Introduction
2. Materials and Methods
2.1. SCG Hydrolysates Preparation
2.2. Inoculum Preparation
2.3. Microbial Fermentation Design
2.4. Total Soluble Solids Contents and pH Measurements
2.5. Analysis of Non-Volatile Composition
2.6. Analysis of Volatile Composition
2.7. Antioxidant Assays
2.8. Statistical Analysis
3. Results and Discussion
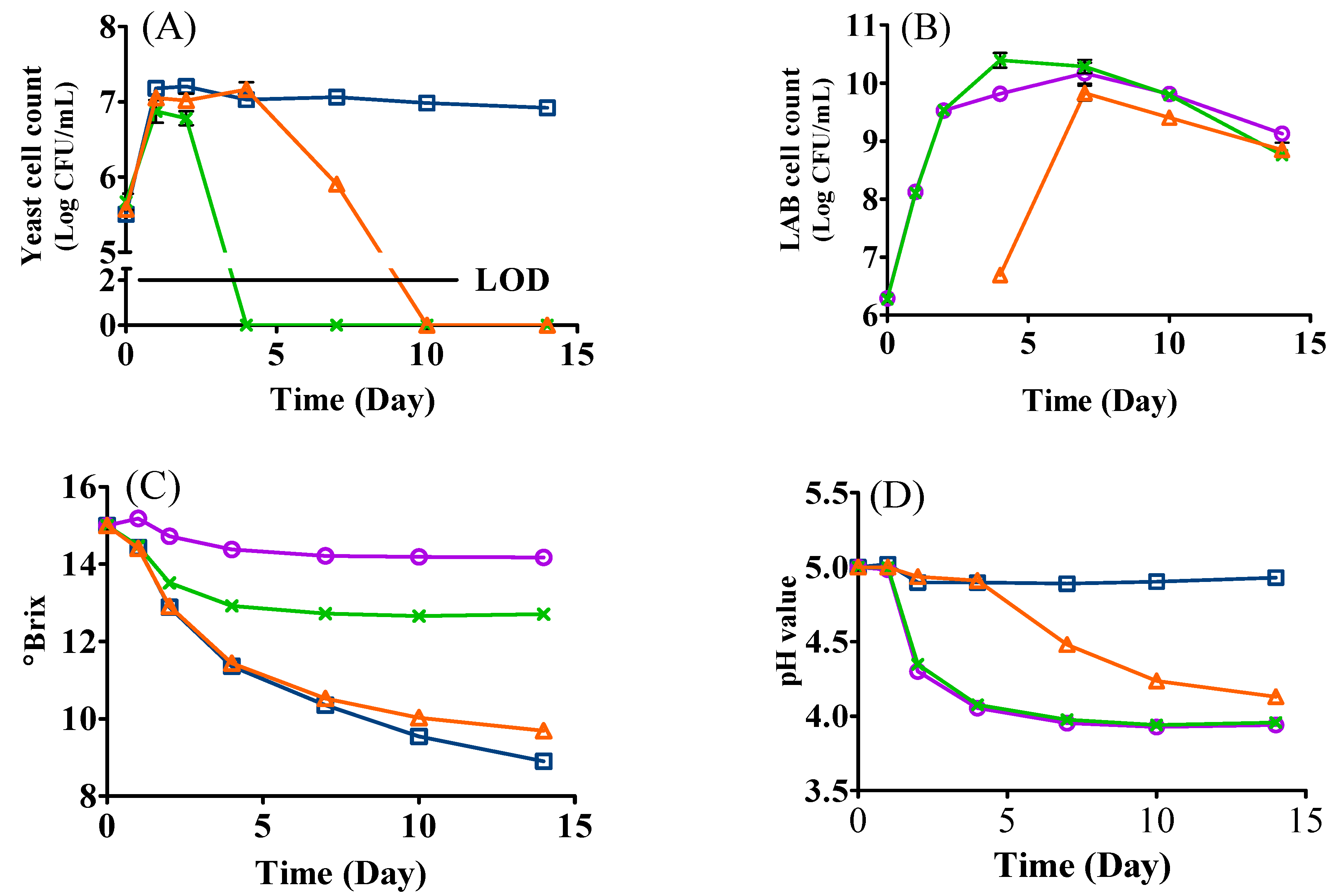
3.1. Microbial Growth
3.2. Changes in °Brix and pH
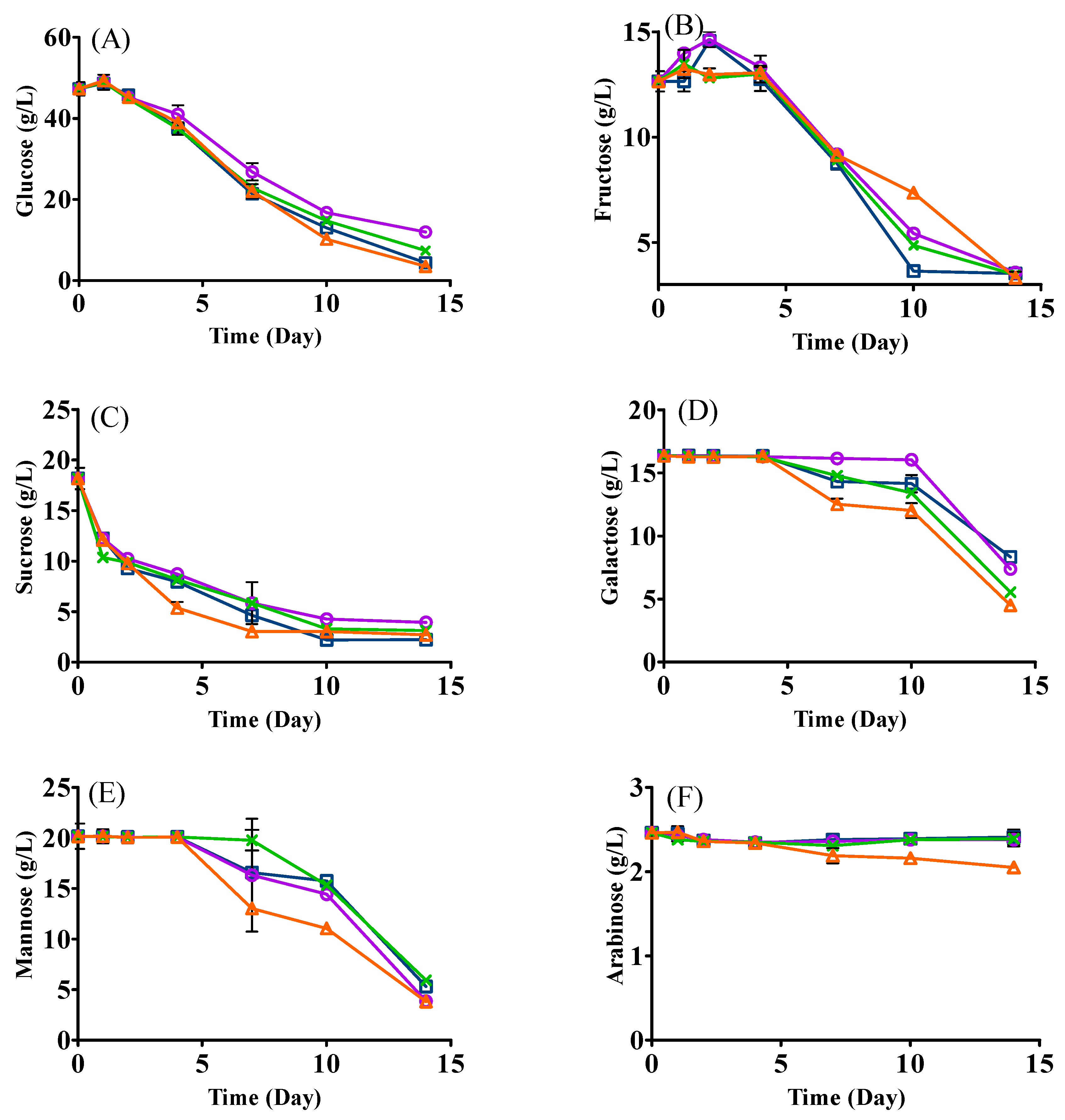
3.3. Changes in Sugars
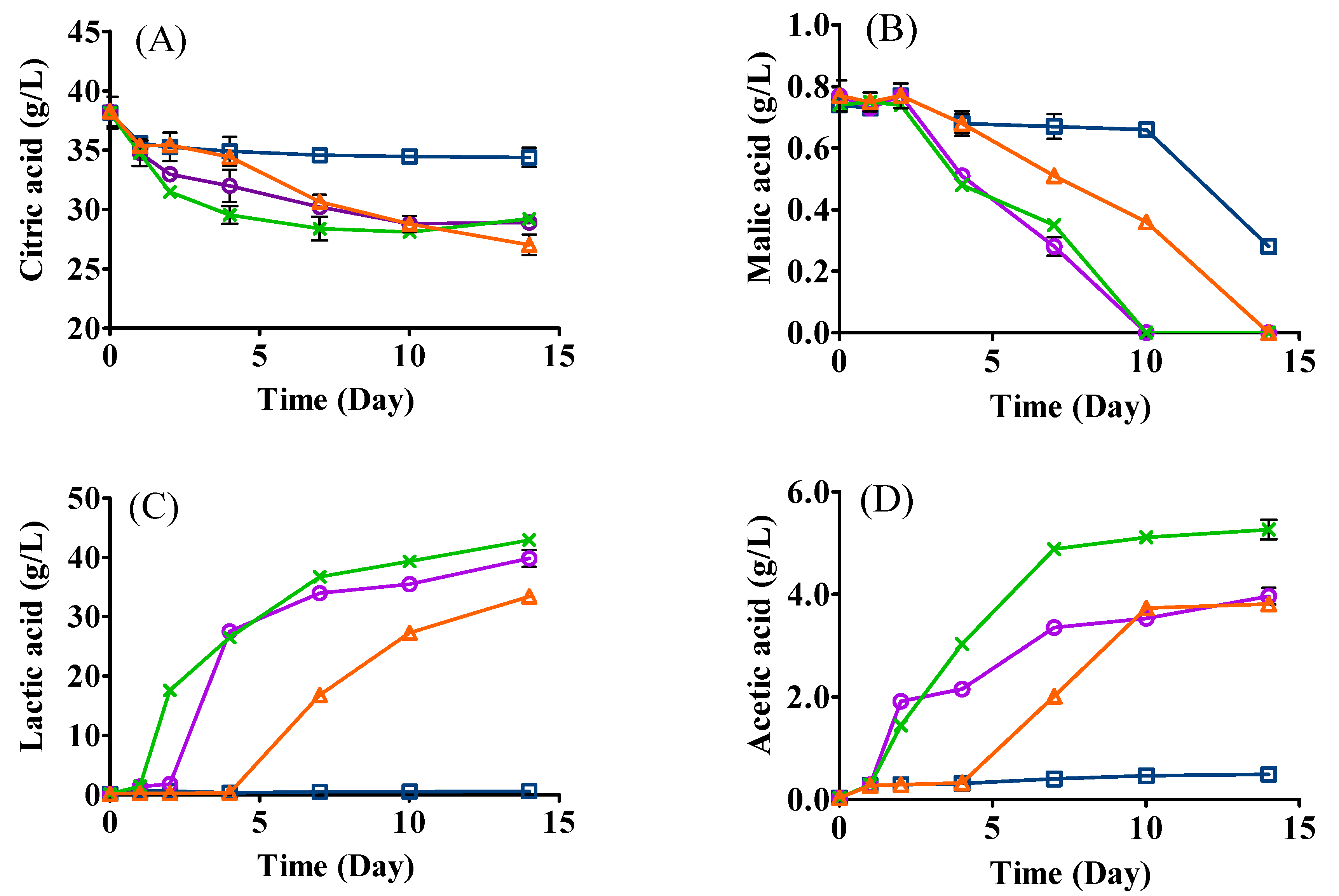
3.4. Changes in Organic Acids
3.5. Changes in Alkaloids, Phenolic Acids, and Antioxidant Capacity
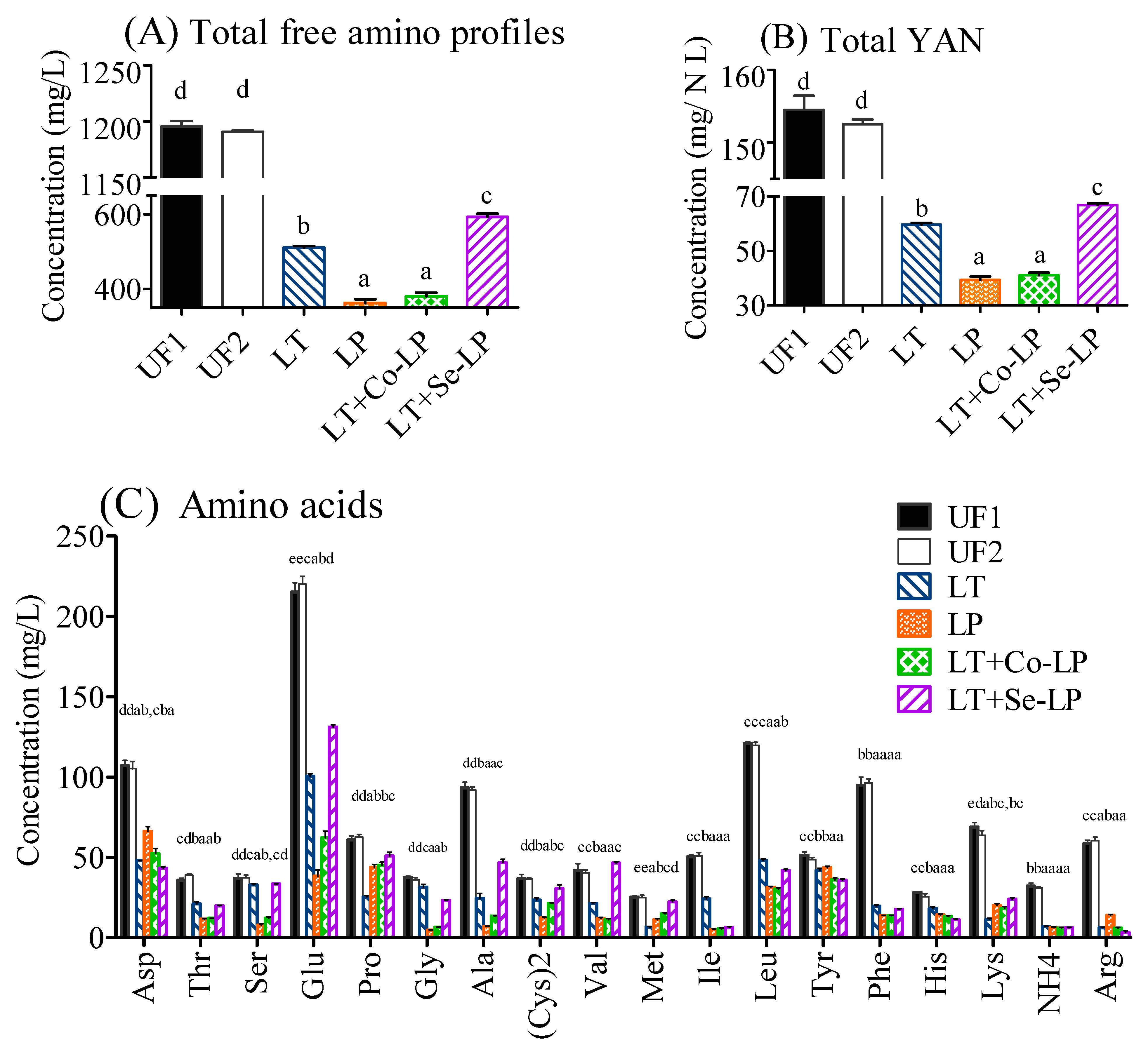
3.6. Changes in Amino Compounds
3.7. Changes in Volatiles and PCA Analysis
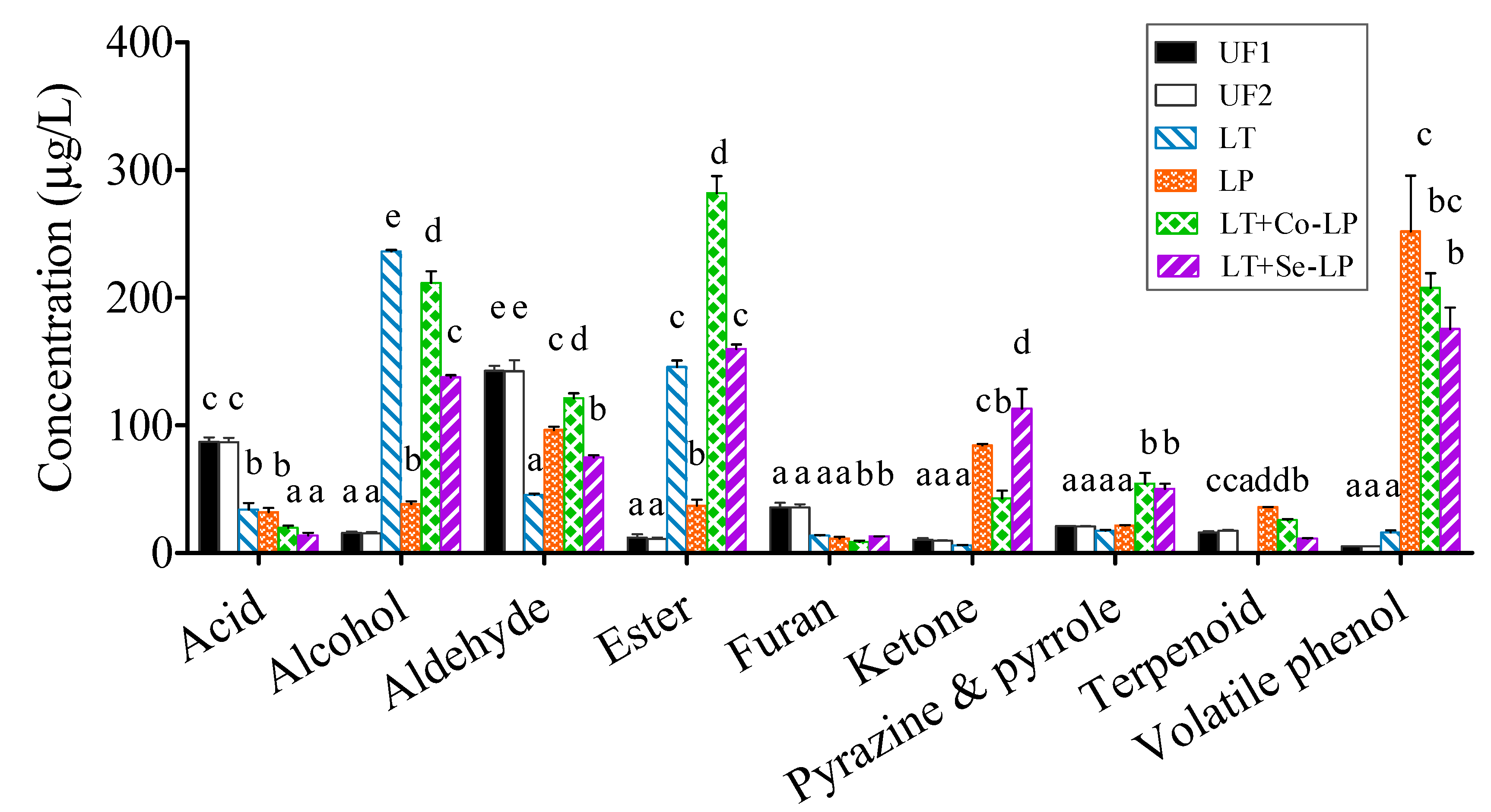
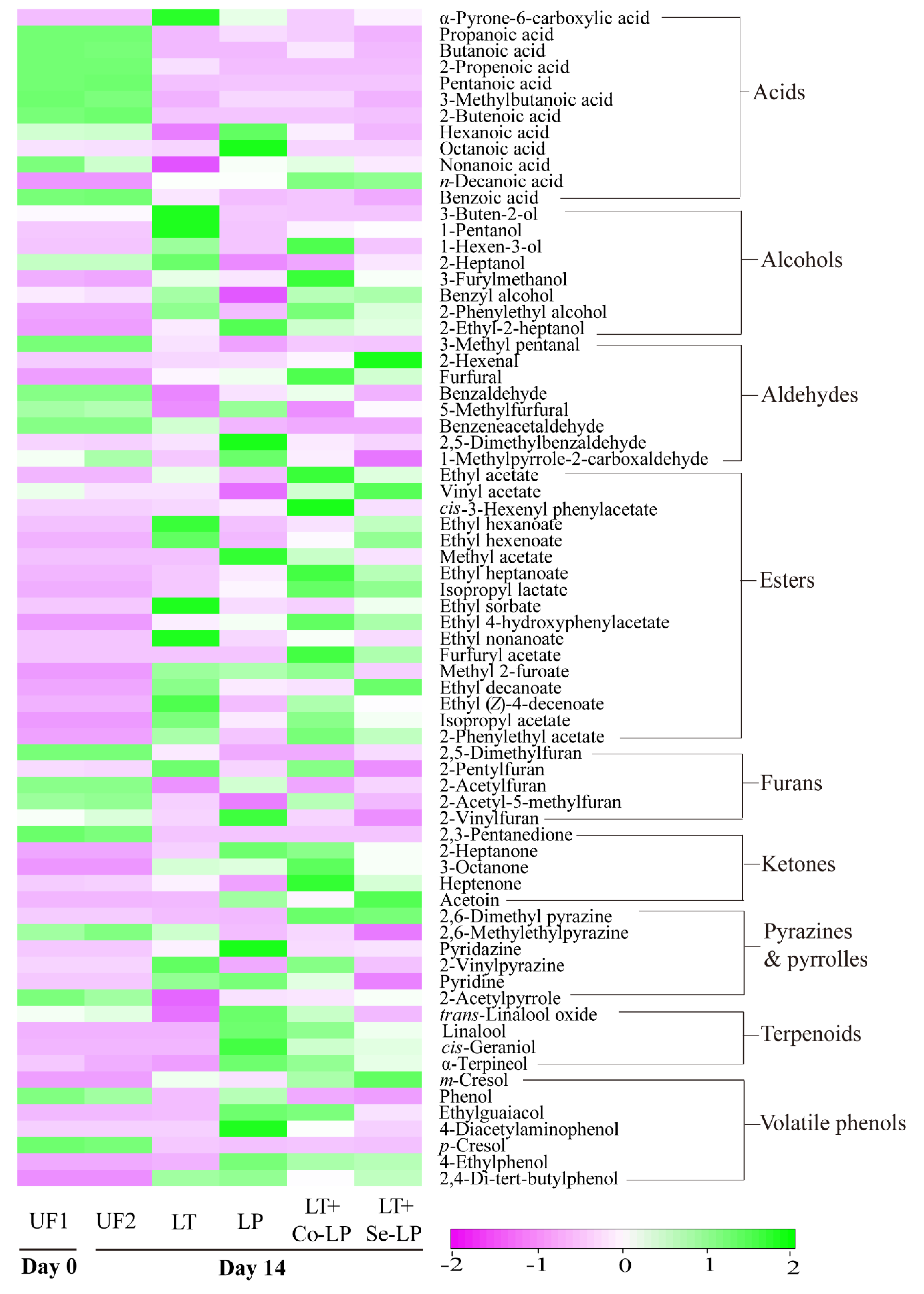
3.7.1. Changes in Volatile Components
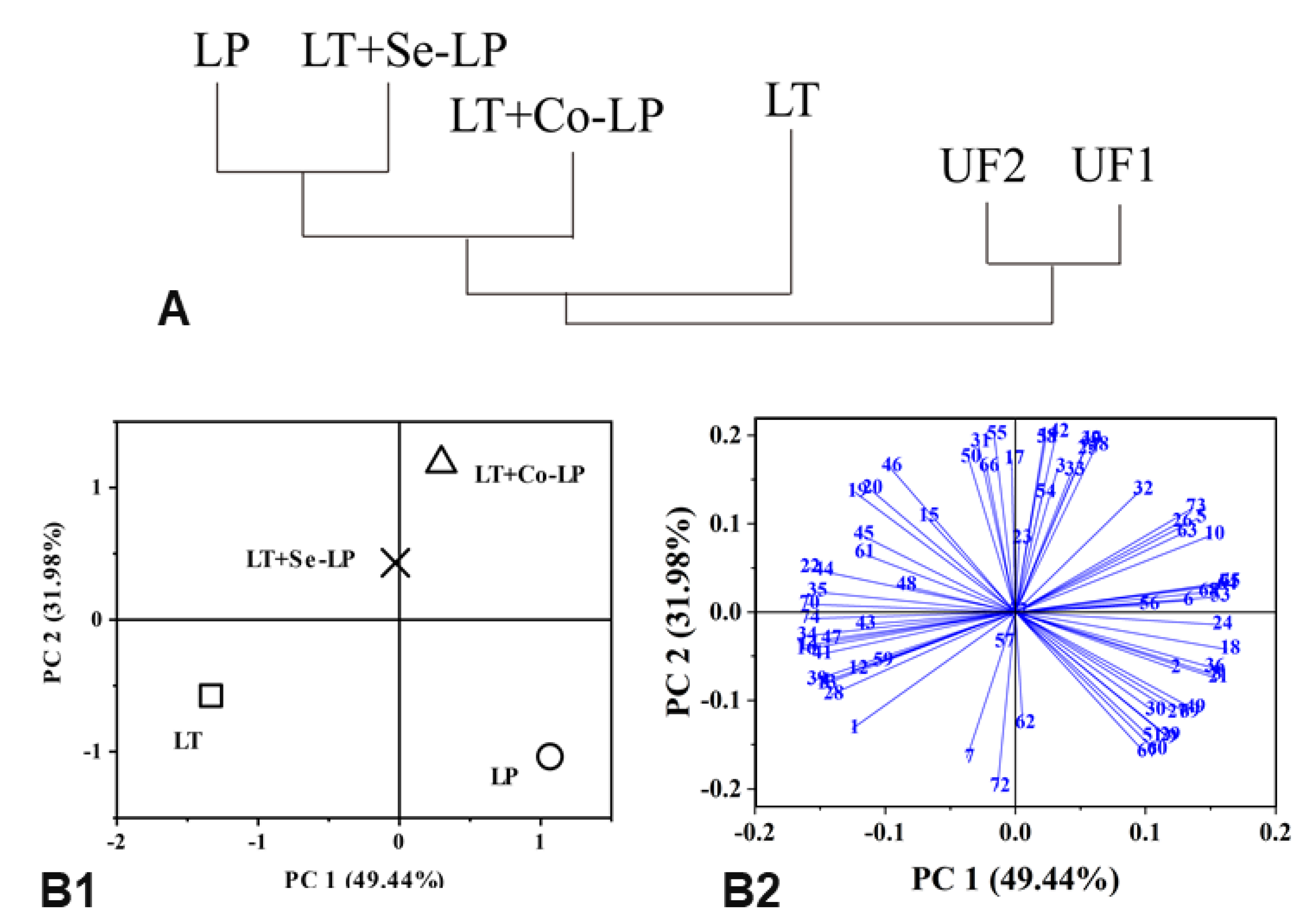
3.7.2. PCA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Escribano-Viana, R.; Portu, J.; Garijo, P.; López, R.; Santamaría, P.; López-Alfaro, I.; Gutiérrez, A.R.; González-Arenzana, L. Effect of the sequential inoculation of non-Saccharomyces/Saccharomyces on the anthocyans and stilbenes composition of tem-pranillo wines. Front. Microbiol. 2019, 10, 773. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef]
- Du Plessis, H.; Du Toit, M.; Nieuwoudt, H.; Van der Rijst, M.; Hoff, J.; Jolly, N. Modulation of Wine Flavor using Hanseniaspora uvarum in Combination with Different Saccharomyces cerevisiae, Lactic Acid Bacteria Strains and Malolactic Fermentation Strategies. Fermentation 2019, 5, 64. [Google Scholar] [CrossRef]
- Liu, Y.; Yuan, W.; Lu, Y.; Liu, S.Q. Biotransformation of spent coffee grounds by fermentation with monocultures of Saccha-romyces cerevisiae and Lachancea thermotolerans aided by yeast extracts. LWT-Food Sci. Technol. 2021, 138, 110751–110762. [Google Scholar] [CrossRef]
- Lu, Y.; Huang, D.; Lee, P.R.; Liu, S.Q. Effects of cofermentation and sequential inoculation of Saccharomyces bayanus and Torulaspora delbruckii on durian wine composition. Int. J. Food Sci. Technol. 2015, 50, 2653–2663. [Google Scholar] [CrossRef]
- Lu, Y.; Huang, D.; Lee, P.R.; Liu, S.Q. Assessment of volatile and non-volatile compounds in durian wines fermented with four commercial non-Saccharomyces yeasts. J. Sci. Food Agric. 2016, 96, 1511–1521. [Google Scholar] [CrossRef]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The Oenological Potential of Hanseniaspora uvarum in Simultaneous and Sequential Co-fermentation with Saccharomyces cerevisiae for Industrial Wine Production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef]
- Brizuela, N.; Tymczyszyn, E.E.; Semorile, L.C.; Valdes, D.; Hens, L.; Delfederico, L.; Hollmann, A.; Bravo-ferrada, B. Lactoba-cillus plantarum as a malolactic starter culture in winemaking: A new (old) player? Electron. J. Biotechnol. 2019, 38, 10–18. [Google Scholar] [CrossRef]
- Liu, Y.; Seah, R.H.; Abdul Rahaman, M.S.; Lu, Y.; Liu, S.Q. Concurrent inoculations of Oenococcus oeni and Lachancea thermo-tolerans: Impacts on non-volatile and volatile components of spent coffee grounds hydrolysates. LWT-Food Sci. Technol. 2021, 148, 111795–111804. [Google Scholar] [CrossRef]
- Lu, Y.; Chua, J.Y.; Voon, M.K.W.; Huang, D.; Lee, P.R.; Liu, S.Q. Effects of different inoculation regimes of Torulaspora delbrueckii and Oenococcus oeni on fermentation kinetics and chemical constituents of durian wine. S. Afr. J. Enol. Vitic. 2017, 38, 273–285. [Google Scholar] [CrossRef]
- G-Alegría, E.; López, I.; Ruiz-Sanz, J.I.; Sã¡enz, J.; Fernã¡ndez, E.; Zarazaga, M.; Dizy, M.; Torres, C.; Larrea, F.R. High tolerance of wild Lactobacillus plantarum and Oenococcus oeni strains to lyophilisation and stress environmental conditions of acid pH and ethanol. FEMS Microbiol. Lett. 2004, 230, 53–61. [Google Scholar] [CrossRef]
- Krieger-Weber, S.; Heras, J.M.; Suarez, C. Lactobacillus plantarum, a New Biological Tool to Control Malolactic Fermentation: A Review and an Outlook. Beverages 2020, 6, 23. [Google Scholar] [CrossRef]
- Tufariello, M.; Capozzi, V.; Spano, G.; Cantele, G.; Venerito, P.; Mita, G.; Grieco, F. Effect of Co-Inoculation of Candida zemplinina, Saccharomyces cerevisiae and Lactobacillus plantarum for the Industrial Production of Negroamaro Wine in Apulia (Southern Italy). Microorganisms 2020, 8, 726. [Google Scholar] [CrossRef]
- Russo, P.; Englezos, V.; Capozzi, V.; Pollon, M.; Segade, S.R.; Rantsiou, K.; Spano, G.; Cocolin, L. Effect of mixed fermentations with Starmerella bacillaris and Saccharomyces cerevisiae on management of malolactic fermentation. Food Res. Int. 2020, 134, 109246. [Google Scholar] [CrossRef]
- Urbina, Á.; Calderón, F.; Benito, S. The Combined Use of Lachancea thermotolerans and Lactiplantibacillus plantarum (former Lactobacillus plantarum) in Wine Technology. Foods 2021, 10, 1356. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, Y.; Liu, S.Q. The potential of spent coffee grounds hydrolysates fermented with Torulaspora delbrueckii and Pichia kluyveri for developing an alcoholic beverage: The yeasts growth and chemical compounds modulation by yeast extracts. Curr. Res. Food Sci. 2021, 4, 489–498. [Google Scholar] [CrossRef]
- Sampaio, A.; Dragone, G.; Vilanova, M.; Oliveira, J.M.; Teixeira, J.A.; Mussatto, S.I. Production, chemical characterization, and sensory profile of a novel spirit elaborated from spent coffee ground. LWT-Food Sci. Technol. 2013, 54, 557–563. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-Throughput Assay of Oxygen Radical Absorbance Capacity (ORAC) Using a Multichannel Liquid Handling System Coupled with a Microplate Fluorescence Reader in 96-Well Format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Bartle, L.; Sumby, K.; Sundstrom, J.; Jiranek, V. The microbial challenge of winemaking: Yeast-bacteria compatibility. FEMS Yeast Res. 2019, 19, 1–16. [Google Scholar] [CrossRef]
- Sangmanee, P.; Hongpattarakere, T. Inhibitory of multiple antifungal components produced by Lactobacillus plantarum K35 on growth, aflatoxin production and ultrastructure alterations of Aspergillus flavus and Aspergillus parasiticus. Food Control 2014, 40, 224–233. [Google Scholar] [CrossRef]
- Plumed-Ferrer, C.; Koistinen, K.M.; Tolonen, T.L.; Lehesranta, S.J.; Karenlampi, S.O.; Makimattila, E.; Von Wright, A. Com-parative study of sugar fermentation and protein expression patterns of two Lactobacillus plantarum strains grown in three different media. Appl. Environ. Microbiol. 2008, 74, 5349–5358. [Google Scholar] [CrossRef]
- Stevens, M.J.A.; Molenaar, D.; De Jong, A.; De Vos, W.M.; Kleerebezem, M. σ54-mediated control of the mannose phos-photransferase sytem in Lactobacillus plantarum impacts on carbohydrate metabolism. Microbiology 2010, 156, 695–707. [Google Scholar] [CrossRef]
- Kennes, C.; Dubourguler, H.C.; Albagnac, G.; Nyns, E.J. Citrate metabolism by Lactobacillus plantarum isolated from orange juice. J. Appl. Bacteriol. 1991, 70, 380–384. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Sedewitz, B.; Schleifer, K.H.; Götz, F. Physiological role of pyruvate oxidase in the aerobic metabolism of Lactobacillus plantarum. J. Bacteriol. 1984, 160, 462–465. [Google Scholar] [CrossRef]
- Landete, J.M.; Ferrer, S.; Monedero, V.; Zúñiga, M. Malic Enzyme and Malolactic Enzyme Pathways Are Functionally Linked but Independently Regulated in Lactobacillus casei BL23. Appl. Environ. Microbiol. 2013, 79, 5509–5518. [Google Scholar] [CrossRef]
- Liu, S.-Q.; Holland, R.; McJarrow, P.; Crow, V. Serine metabolism in Lactobacillus plantarum. Int. J. Food Microbiol. 2003, 89, 265–273. [Google Scholar] [CrossRef]
- Lu, Y.; Chua, J.-Y.; Huang, D.; Lee, P.-R.; Liu, S.-Q. Biotransformation of chemical constituents of durian wine with simultaneous alcoholic fermentation by Torulaspora delbrueckii and malolactic fermentation by Oenococcus oeni. Appl. Microbiol. Biotechnol. 2016, 100, 8877–8888. [Google Scholar] [CrossRef]
- Dudley, E.; Steele, J. Succinate production and citrate catabolism by Cheddar cheese nonstarter lactobacilli. J. Appl. Microbiol. 2005, 98, 14–23. [Google Scholar] [CrossRef]
- Martín-García, A.; Balmaseda, A.; Bordons, A.; Reguant, C. Effect of the inoculation strategy of non-Saccharomyces yeasts on wine malolactic fermentation. OENO One 2020, 54, 101–108. [Google Scholar] [CrossRef]
- Joshi, J.G.; Handler, P. Metabolism of Trigonelline. J. Biol. Chem. 1962, 237, 3185–3188. [Google Scholar] [CrossRef]
- Dash, S.S.; Gummadi, S.N. Catabolic pathways and biotechnological applications of microbial caffeine degradation. Biotechnol. Lett. 2006, 28, 1993–2002. [Google Scholar] [CrossRef]
- Glück, M.; Lingens, F. Studies on the microbial production of theobromine and heteroxanthine from caffeine. Appl. Microbiol. Biotechnol. 1987, 25, 334–340. [Google Scholar] [CrossRef]
- Crépin, L.; Nidelet, T.; Sanchez, I.; Dequin, S.; Camarasa, C. Sequential Use of Nitrogen Compounds by Saccharomyces cerevisiae during Wine Fermentation: A Model Based on Kinetic and Regulation Characteristics of Nitrogen Permeases. Appl. Environ. Microbiol. 2012, 78, 8102–8111. [Google Scholar] [CrossRef]
- Knockaert, D.; Raes, K.; Wille, C.; Struijs, K.; Van Camp, J. Metabolism of ferulic acid during growth of Lactobacillus plantarum and Lactobacillus collinoides. J. Sci. Food Agric. 2012, 92, 2291–2296. [Google Scholar] [CrossRef]
- Santamaría, L.; Reverón, I.; de Felipe, F.L.; Rivas, B.D.L.; Muñoz, R. Ethylphenol Formation by Lactobacillus plantarum: Identification of the Enzyme Involved in the Reduction of Vinylphenols. Appl. Environ. Microbiol. 2018, 84, e01064-18. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Antioxidant Property of Coffee Components: Assessment of Methods that Define Mechanisms of Action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Vilanova, M.; Ugliano, M.; Varela, C.; Siebert, T.; Pretorius, I.S.; Henschke, P.A. Assimilable nitrogen utilisation and production of volatile and non-volatile compounds in chemically defined medium by Saccharomyces cerevisiae wine yeasts. Appl. Microbiol. Biotechnol. 2007, 77, 145–157. [Google Scholar] [CrossRef]
- Virdis, C.; Sumby, K.; Bartowsky, E.; Jiranek, V. Lactic Acid Bacteria in Wine: Technological Advances and Evaluation of Their Functional Role. Front. Microbiol. 2021, 11, 612118. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
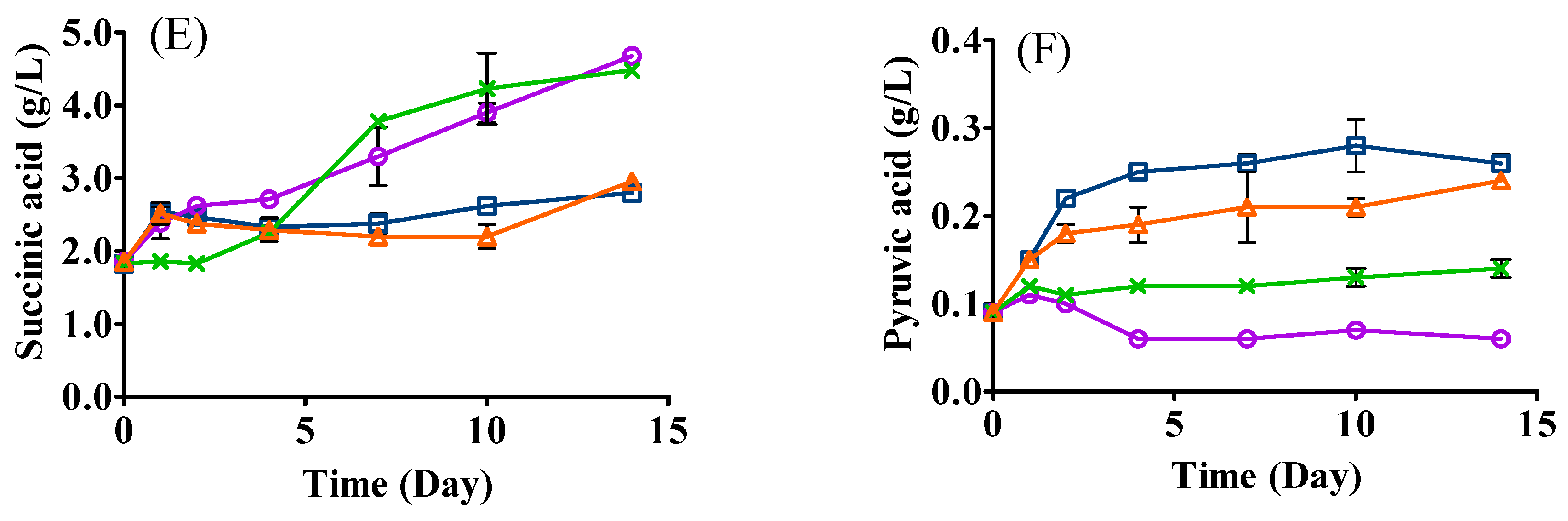{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day 0 | Day 14 | |||||
|---|---|---|---|---|---|---|
| UF1 | UF2 | LT | LP | LT+Co-LP | LT+Se-LP | |
| pH | 5.00 ± 0.00 d | 5.00 ± 0.00 d | 4.93 ± 0.02 c | 3.94 ± 0.01 a | 3.96 ± 0.01 a | 4.13 ±0.01 c |
| °Brix | 15.02 ± 0.01 e | 15.00 ± 0.01 e | 8.90 ± 0.02 a | 14.17 ± 0.04 d | 12.70 ± 0.05 c | 9.69 ± 0.03 b |
| Ethanol (%, v/v) | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 4.92 ± 0.28 c | 0.00 ± 0.00 a | 1.03 ± 0.01 b | 3.20 ± 0.14 c |
| Sugars (g/L) | ||||||
| Fructose | 12.65 ± 0.48 b | 12.36 ± 0.06 b | 3.53 ± 0.04 a | 3.58 ± 0.15 a | 3.47 ± 0.17 a | 3.33 ± 0.29 a |
| Glucose | 47.33 ± 1.67 d | 47.31 ± 0.47 d | 4.40 ± 0.12 a | 12.02 ± 0.82 c | 7.40 ± 0.08 b | 3.53 ± 0.20 a |
| Sucrose | 18.18 ± 1.06 c | 18.29 ± 0.28 c | 2.23 ± 0.03 a | 3.94 ± 0.16 b | 3.13 ± 0.10 ab | 2.72± 0.02 ab |
| Mannose | 20.17 ± 1.25 d | 20.18 ± 0.22 d | 5.31 ± 0.27 bc | 3.86 ± 0.05 ab | 5.93 ± 0.14 c | 3.82 ± 0.05 a |
| Galactose | 16.35 ± 0.31 e | 16.37 ± 0.37 e | 8.33 ± 0.17 d | 7.37 ± 0.16 c | 5.55 ± 0.09 b | 4.50 ± 0.22 a |
| Arabinose | 2.46 ± 0.06 b | 2.49 ± 0.08 b | 2.41 ± 0.09 b | 2.38 ± 0.08 b | 2.39 ± 0.05 b | 2.05 ± 0.03 a |
| Total | 117.25 ± 1.01 e | 117.11 ± 0.56 e | 26.26 ± 0.32 b | 33.21 ± 0.70 d | 27.94 ± 0.24 c | 20.37 ± 0.08 a |
| Glycerol (g/L) | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 4.27 ± 0.25 d | 0.00 ± 0.00 a | 1.07 ± 0.04 b | 2.52 ± 0.17 c |
| Organic acids (g/L) | ||||||
| Citric acid | 38.16 ± 1.33 c | 38.26 ± 1.25 c | 34.39 ± 0.81 b | 28.91 ± 0.06 a | 29.24 ± 0.32 a | 27.02 ± 0.87 a |
| α-Ketoglutaric acid (mg/L) | 33.70 ± 1.27 b | 31.37 ± 2.98 b | 19.72 ± 0.70 a | 18.04 ± 1.16 a | 31.79 ± 0.97 a | 34.36 ± 0.83 a |
| Malic acid | 0.46 ± 0.04 b | 0.45 ± 0.04 b | 0.10 ± 0.00 a | 0.08 ± 0.01 a | 0.08 ± 0.01 a | 0.08 ± 0.01 a |
| Pyruvic acid | 0.09 ± 0.01 b | 0.09 ± 0.00 b | 0.26 ± 0.01 e | 0.06 ± 0.00 a | 0.14 ± 0.01 c | 0.24 ± 0.00 d |
| Succinic acid | 1.83 ± 0.04 a | 1.85 ± 0.01 a | 2.80 ± 0.05 b | 4.68 ± 0.00 c | 4.48 ± 0.08 c | 2.96 ± 0.01 b |
| Lactic acid | 0.14 ± 0.00 a | 0.14 ± 0.01 a | 0.58 ± 0.01 a | 39.84 ± 1.42 c | 42.92 ± 0.36 d | 33.41 ± 0.60 b |
| Acetic acid | 0.13 ± 0.00 a | 0.13 ± 0.05 a | 0.49 ± 0.02 b | 3.96 ± 0.16 c | 5.26 ± 0.19 d | 3.81 ± 0.08 c |
| Total | 40.37 ± 1.38 a | 40.49 ± 1.26 a | 38.55 ± 0.90 a | 77.49 ± 1.64 c | 82.07 ± 1.00 d | 67.49 ± 1.56 b |
| Day 0 | Day 14 | |||||
|---|---|---|---|---|---|---|
| UF1 | UF2 | LT | LP | LT+Co-LP | LT+Se-LP | |
| Alkaloids | ||||||
| Trigonelline | 530.82 ± 4.07 b | 529.69 ± 3.66 b | 532.13 ± 5.48 b | 518.24 ± 10.18 b | 452.72 ± 5.72 a | 507.62 ± 2.48 b |
| Caffeine | 1181.26 ± 11.65 b | 1178.82 ± 1.90 b | 1151.97 ± 1.39 b | 806.02 ± 7.69 a | 800.26± 1.20 a | 788.15 ± 2.46 a |
| Theobromine | 124.75 ± 1.38 d | 122.92 ± 2.10 d | 40.92 ± 0.90 a | 103.24 ± 1.32 c | 100.15 ± 0.23 c | 77.90 ± 1.05 b |
| Theophylline | 73.66 ± 1.83 b | 73.43 ± 1.51 b | 33.97 ± 0.84 a | 64.05 ± 0.07 b | 64.92 ± 0.37 b | 92.10 ± 3.75 c |
| Phenolic acids | ||||||
| Chlorogenic acid | 135.70 ± 1.58 d | 135.13 ± 1.30 d | 53.67 ± 1.29 b | 120.33 ± 5.33 c | 26.19 ± 0.45 a | 24.69 ± 1.11 a |
| Caffeic acid | 595.47 ± 5.67 b | 596.58 ± 6.26 b | 646.46 ± 13.56 c | 16.20 ± 1.01 a | 37.81 ± 1.40 a | 11.31 ± 1.63 a |
| Ferulic acid | 13.19 ± 0.20 b | 13.06 ± 0.22 b | 27.97 ± 1.78 c | 7.66 ± 0.19 a | 10.36 ± 0.28 ab | 10.21 ± 0.70 ab |
| p-Coumaric acid | 7.55 ± 0.02 c | 7.60 ± 0.05 c | 11.85 ± 0.11 d | 6.04 ± 0.02 b | 5.65 ± 0.06 a | 6.16 ± 0.01 b |
| Antioxidant capacity | ||||||
| DPPH (μmol TE/L) | 4.58 ± 0.10 c | 4.31 ± 0.13 c | 4.74 ± 0.37 c | 3.61 ± 0.27 b | 3.34 ± 0.01 a | 3.66 ± 0.22 b |
| ORAC (mmol TE/L) | 707.00 ± 42.22 a | 687.61 ± 88.99 a | 756.56 ± 92.71 a | 827.05 ± 74.28 a | 703.67 ± 1.66 a | 684.27 ± 83.52 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Lu, Y.; Liu, S.-Q. Transforming Spent Coffee Grounds’ Hydrolysates with Yeast Lachancea thermotolerans and Lactic Acid Bacterium Lactiplantibacillus plantarum to Develop Potential Novel Alcoholic Beverages. Foods 2023, 12, 1161. https://doi.org/10.3390/foods12061161
Liu Y, Lu Y, Liu S-Q. Transforming Spent Coffee Grounds’ Hydrolysates with Yeast Lachancea thermotolerans and Lactic Acid Bacterium Lactiplantibacillus plantarum to Develop Potential Novel Alcoholic Beverages. Foods. 2023; 12(6):1161. https://doi.org/10.3390/foods12061161
Chicago/Turabian StyleLiu, Yunjiao, Yuyun Lu, and Shao-Quan Liu. 2023. "Transforming Spent Coffee Grounds’ Hydrolysates with Yeast Lachancea thermotolerans and Lactic Acid Bacterium Lactiplantibacillus plantarum to Develop Potential Novel Alcoholic Beverages" Foods 12, no. 6: 1161. https://doi.org/10.3390/foods12061161
APA StyleLiu, Y., Lu, Y., & Liu, S.-Q. (2023). Transforming Spent Coffee Grounds’ Hydrolysates with Yeast Lachancea thermotolerans and Lactic Acid Bacterium Lactiplantibacillus plantarum to Develop Potential Novel Alcoholic Beverages. Foods, 12(6), 1161. https://doi.org/10.3390/foods12061161