

Associating Compositional, Nutritional and Techno-Functional Characteristics of Faba Bean (Vicia faba L.) Protein Isolates and Their Production Side-Streams with Potential Food Applications
 , , ,
, , , 
Abstract
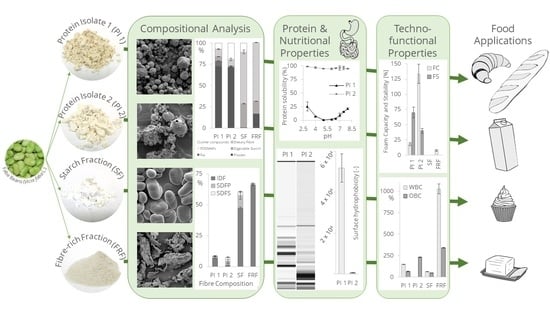
1. Introduction
2. Materials and Methods
2.1. Chemicals and Raw Materials
2.2. Preparation of Faba Bean Protein Ingredients
2.3. Compositional Analysis
2.4. Analysis of Antinutrients
2.4.1. Vicine and Convicine
2.4.2. Phytic Acid
2.4.3. Total Saponin Content
2.4.4. Condensed Tannins
2.4.5. Trypsin and Chymotrypsin Inhibitor Activity
2.5. Protein Analysis
2.5.1. Protein Solubility
2.5.2. Surface Hydrophobicity
2.5.3. Protein Profile
2.5.4. In Vitro Protein Digestibility
2.6. Microscopy
2.7. Techno-Functional Properties
2.7.1. Colour Measurement
2.7.2. pH and Total Titratable Acidity
2.7.3. Foaming Properties
2.7.4. Water and Oil Binding Capacity
2.7.5. Statistical Data Analysis
3. Results and Discussion
3.1. Compositional Analysis
3.2. Antinutritional Factors
3.2.1. Vicine and Convicine
3.2.2. Phytic Acid
3.2.3. Total Saponin Content
3.2.4. Condensed Tannins
3.2.5. Trypsin and Chymotrypsin Inhibitor Activity
3.3. Protein Analysis
3.3.1. Protein Solubility and Surface Hydrophobicity
3.3.2. Protein Profiles
3.3.3. In Vitro Protein Digestibility
3.4. Microscopy
3.5. Techno-Functional Properties
3.5.1. Colour Measurement
3.5.2. pH and Total Titratable Acidity
3.5.3. Foaming Properties
3.5.4. Water and Oil Binding Capacities
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations Department of Economic and Social Affairs, Population Division. World Population Prospects 2022: Summary of Results; United Nations: New York, NY, USA, 2022. [Google Scholar]
- United Nations. 2030 Agenda for Sustainable Development; United Nations: New York, NY, USA, 2015. [Google Scholar]
- Snowdon, R.J.; Wittkop, B.; Chen, T.W.; Stahl, A. Crop Adaptation to Climate Change as a Consequence of Long-Term Breeding. Theor. Appl. Genet. 2021, 134, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Crippa, M.; Solazzo, E.; Guizzardi, D.; Monforti-Ferrario, F.; Tubiello, F.N.; Leip, A. Food Systems Are Responsible for a Third of Global Anthropogenic GHG Emissions. Nat. Food 2021, 2, 198–209. [Google Scholar] [CrossRef]
- de Boer, J.; Aiking, H. Prospects for Pro-Environmental Protein Consumption in Europe: Cultural, Culinary, Economic and Psychological Factors. Appetite 2018, 121, 29–40. [Google Scholar] [CrossRef]
- Clune, S.; Crossin, E.; Verghese, K. Systematic Review of Greenhouse Gas Emissions for Different Fresh Food Categories. J. Clean. Prod. 2017, 140, 766–783. [Google Scholar] [CrossRef]
- Hall, C.; Hillen, C.; Robinson, J.G. Composition, Nutritional Value, and Health Benefits of Pulses. Cereal Chem. 2017, 94, 11–31. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on Healthy Diets from Sustainable Food Systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Andrade, C.; Olías, R.; Jiménez-López, J.C.; Clemente, A. Nutritional and Beneficial Effects of Grain Legumes on Human Health. Arbor 2016, 192, 1–13. [Google Scholar] [CrossRef]
- Legumes for Soil Health and Sustainable Management; Meena, R.S., Das, A., Singh Yadav, G., Lal, R., Eds.; Springer Nature Singapore Pte Ltd.: Varanasi, India, 2018; ISBN 9789811302527. [Google Scholar]
- Senbayram, M.; Wenthe, C.; Lingner, A.; Isselstein, J.; Steinmann, H.; Kaya, C.; Köbke, S. Legume-Based Mixed Intercropping Systems May Lower Agricultural Born N2O Emissions. Energy. Sustain. Soc. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Köpke, U.; Nemecek, T. Ecological Services of Faba Bean. F. Crop. Res. 2010, 115, 217–233. [Google Scholar] [CrossRef]
- Coda, R.; Melama, L.; Rizzello, C.G.; Curiel, J.A.; Sibakov, J.; Holopainen, U.; Pulkkinen, M.; Sozer, N. Effect of Air Classification and Fermentation by Lactobacillus Plantarum VTT E-133328 on Faba Bean (Vicia Faba L.) Flour Nutritional Properties. Int. J. Food Microbiol. 2015, 193, 34–42. [Google Scholar] [CrossRef]
- Mayer Labba, I.C.; Frøkiær, H.; Sandberg, A.S. Nutritional and Antinutritional Composition of Fava Bean (Vicia faba L., Var. Minor) Cultivars. Food Res. Int. 2021, 140, 110038. [Google Scholar] [CrossRef]
- Martineau-Côté, D.; Achouri, A.; Karboune, S.; L’Hocine, L. Faba Bean: An Untapped Source of Quality Plant Proteins and Bioactives. Nutrients 2022, 14, 1541. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.K.; Lucey, A.; Walter, J.; Zannini, E.; Arendt, E.K. Resistant Starch—An Accessible Fiber Ingredient Acceptable to the Western Palate. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2930–2955. [Google Scholar] [CrossRef] [PubMed]
- Cronin, P.; Joyce, S.A.; O’toole, P.W.; O’connor, E.M. Dietary Fibre Modulates the Gut Microbiota. Nutrients 2021, 13, 1655. [Google Scholar] [CrossRef] [PubMed]
- La Torre, D.; Verbeke, K.; Dalile, B. Dietary Fibre and the Gut–Brain Axis: Microbiota-Dependent and Independent Mechanisms of Action. Gut Microbiome 2021, 2, 1–26. [Google Scholar] [CrossRef]
- Kumar, J.; Rani, K.; Datt, C. Molecular Link between Dietary Fibre, Gut Microbiota and Health. Mol. Biol. Rep. 2020, 47, 6229–6237. [Google Scholar] [CrossRef] [PubMed]
- Khamassi, K.; Ben Jeddi, F.; Hobbs, D.; Irigoyen, J.; Stoddard, F.; O’Sullivan, D.M.; Jones, H. A Baseline Study of Vicine-Convicine Levels in Faba Bean (Vicia faba L.) Germplasm. Plant Genet. Resour. Characterisation Util. 2013, 11, 250–257. [Google Scholar] [CrossRef]
- Ivarsson, E.; Neil, M. Variations in Nutritional and Antinutritional Contents among Faba Bean Cultivars and Effects on Growth Performance of Weaner Pigs. Livest. Sci. 2018, 212, 14–21. [Google Scholar] [CrossRef]
- Felix, M.; Lopez-Osorio, A.; Romero, A.; Guerrero, A. Faba Bean Protein Flour Obtained by Densification: A Sustainable Method to Develop Protein Concentrates with Food Applications. LWT 2018, 93, 563–569. [Google Scholar] [CrossRef]
- Vogelsang-O’Dwyer, M.; Petersen, I.L.; Joehnke, M.S.; Sørensen, J.C.; Bez, J.; Detzel, A.; Busch, M.; Krueger, M.; O’Mahony, J.A.; Arendt, E.K.; et al. Comparison of Faba Bean Protein Ingredients Produced Using Dry Fractionation and Isoelectric Precipitation: Techno-Functional, Nutritional and Environmental Performance. Foods 2020, 9, 322. [Google Scholar] [CrossRef]
- Saldanha do Carmo, C.; Silventoinen, P.; Nordgård, C.T.; Poudroux, C.; Dessev, T.; Zobel, H.; Holtekjølen, A.K.; Draget, K.I.; Holopainen-Mantila, U.; Knutsen, S.H.; et al. Is Dehulling of Peas and Faba Beans Necessary Prior to Dry Fractionation for the Production of Protein- and Starch-Rich Fractions? Impact on Physical Properties, Chemical Composition and Techno-Functional Properties. J. Food Eng. 2020, 278, 109937. [Google Scholar] [CrossRef]
- Nivala, O.; Mäkinen, O.E.; Kruus, K.; Nordlund, E.; Ercili-Cura, D. Structuring Colloidal Oat and Faba Bean Protein Particles via Enzymatic Modification. Food Chem. 2017, 231, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Schieber, A. Side Streams of Plant Food Processing As a Source of Valuable Compounds: Selected Examples. Annu. Rev. Food Sci. Technol. 2017, 8, 97–112. [Google Scholar] [CrossRef]
- McClements, D.J.; Grossmann, L. The Science of Plant-based Foods: Constructing Next-generation Meat, Fish, Milk, and Egg Analogs. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4049–4100. [Google Scholar] [CrossRef]
- Andersen, K.E.; Sørensen, J.C.; Sørensen, H.; Sørensen, A.D. A Process for the Manufacture of a Product from a Plant. Material. Patent No. WO 2012/116703 A1, 01 March 2012. [Google Scholar]
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting Nitrogen into Protein—Beyond 6.25 and Jones’ Factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Mossé, J. Nitrogen to Protein Conversion Factor for Ten Cereals and Six Legumes or Oilseeds. A Reappraisal of Its Definition and Determination. Variation According to Species and to Seed Protein Content. J. Agric. Food Chem. 1990, 38, 18–24. [Google Scholar] [CrossRef]
- McCleary, B.V.; McNally, M.; Rossiter, P.; Aman, P.; Amrein, T.; Arnouts, S.; Arrigoni, E.; Bauer, L.; Bavor, C.; Brunt, K.; et al. Measurement of Resistant Starch by Enzymatic Digestion in Starch and Selected Plant Materials: Collaborative Study. J. AOAC Int. 2002, 85, 1103–1111. [Google Scholar] [CrossRef]
- Hoehnel, A.; Salas García, J.; Coffey, C.; Zannini, E.; Arendt, E.K. Comparative Study of Sugar Extraction Procedures for HPLC Analysis and Proposal of an Ethanolic Extraction Method for Plant-Based High-Protein Ingredients. J. Sci. Food Agric. 2021, 102, 5055–5064. [Google Scholar] [CrossRef]
- Ispiryan, L.; Heitmann, M.; Hoehnel, A.; Zannini, E.; Arendt, E.K. Optimization and Validation of an HPAEC-PAD Method for the Quantification of FODMAPs in Cereals and Cereal-Based Products. J. Agric. Food Chem. 2019, 67, 4384–4392. [Google Scholar] [CrossRef] [PubMed]
- McCleary, B.V.; McLoughlin, C. Determination of Insoluble, Soluble, and Total Dietary Fiber in Foods Using a Rapid Integrated Procedure of Enzymatic-Gravimetric-Liquid Chromatography: First Action 2022.01. J. AOAC Int. 2023, 106, 127–145. [Google Scholar] [CrossRef] [PubMed]
- Pontonio, E.; Dingeo, C.; Di Cagno, R.; Blandino, M.; Gobbetti, M.; Rizzello, C.G. Brans from Hull-Less Barley, Emmer and Pigmented Wheat Varieties: From by-Products to Bread Nutritional Improvers Using Selected Lactic Acid Bacteria and Xylanase. Int. J. Food Microbiol. 2020, 313, 108384. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.R.; Hsieh, S.C.; Huang, H.Y.; Chou, C.C. Effect of Lactic Fermentation on the Total Phenolic, Saponin and Phytic Acid Contents as Well as Anti-Colon Cancer Cell Proliferation Activity of Soymilk. J. Biosci. Bioeng. 2013, 115, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Nionelli, L.; Montemurro, M.; Pontonio, E.; Verni, M.; Gobbetti, M.; Rizzello, C.G. Pro-Technological and Functional Characterization of Lactic Acid Bacteria to Be Used as Starters for Hemp (Cannabis sativa L.) Sourdough Fermentation and Wheat Bread Fortification. Int. J. Food Microbiol. 2018, 279, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Price, M.L.; Van Scoyoc, S.; Butler, L.G. A Critical Evaluation of the Vanillin Reaction as an Assay for Tannin in Sorghum Grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Liu, K.; Seegers, S.; Cao, W.; Wanasundara, J.; Chen, J.; da Silva, A.E.; Ross, K.; Franco, A.L.; Vrijenhoek, T.; Bhowmik, P.; et al. An International Collaborative Study on Trypsin Inhibitor Assay for Legumes, Cereals, and Related Products. JAOCS J. Am. Oil Chem. Soc. 2021, 98, 375–390. [Google Scholar] [CrossRef]
- Alonso, R.; Aguirre, A.; Marzo, F. Effects of Extrusion and Traditional Processing Methods on Antinutrients and in Vitro Digestibility of Protein and Starch in Faba and Kidney Beans. Food Chem. 2000, 68, 159–165. [Google Scholar] [CrossRef]
- Hayakawa, S.; Nakai, S. Relationships of Hydrophobicity and Net Charge to the Solubility of Milk and Soy Proteins. J. Food Sci. 1985, 50, 486–491. [Google Scholar] [CrossRef]
- Karaca, A.C.; Low, N.; Nickerson, M. Emulsifying Properties of Chickpea, Faba Bean, Lentil and Pea Proteins Produced by Isoelectric Precipitation and Salt Extraction. Food Res. Int. 2011, 44, 2742–2750. [Google Scholar] [CrossRef]
- Amagliani, L.; O’Regan, J.; Kelly, A.L.; O’Mahony, J.A. Composition and Protein Profile Analysis of Rice Protein Ingredients. J. Food Compos. Anal. 2017, 59, 18–26. [Google Scholar] [CrossRef]
- Joehnke, M.S.; Lametsch, R.; Sørensen, J.C. Improved in Vitro Digestibility of Rapeseed Napin Proteins in Mixtures with Bovine Beta-Lactoglobulin. Food Res. Int. 2019, 123, 346–354. [Google Scholar] [CrossRef]
- Joehnke, M.S.; Rehder, A.; Sørensen, S.; Bjergegaard, C.; Sørensen, J.C.; Markedal, K.E. In Vitro Digestibility of Rapeseed and Bovine Whey Protein Mixtures. J. Agric. Food Chem. 2018, 66, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Jeske, S.; Bez, J.; Arendt, E.K.; Zannini, E. Formation, Stability, and Sensory Characteristics of a Lentil-Based Milk Substitute as Affected by Homogenisation and Pasteurisation. Eur. Food Res. Technol. 2019, 245, 1519–1531. [Google Scholar] [CrossRef]
- Boeck, T.; Zannini, E.; Sahin, A.W.; Bez, J.; Arendt, E.K. Nutritional and Rheological Features of Lentil Protein Isolate for Yoghurt-like Application. Foods 2021, 10, 1692. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Miravalles, L.; Jeske, S.; Bez, J.; Detzel, A.; Busch, M.; Krueger, M.; Wriessnegger, C.L.; O’Mahony, J.A.; Zannini, E.; Arendt, E.K. Membrane Filtration and Isoelectric Precipitation Technological Approaches for the Preparation of Novel, Functional and Sustainable Protein Isolate from Lentils. Eur. Food Res. Technol. 2019, 245, 1855–1869. [Google Scholar] [CrossRef]
- Boye, J.I.; Aksay, S.; Roufik, S.; Ribéreau, S.; Mondor, M.; Farnworth, E.; Rajamohamed, S.H. Comparison of the Functional Properties of Pea, Chickpea and Lentil Protein Concentrates Processed Using Ultrafiltration and Isoelectric Precipitation Techniques. Food Res. Int. 2010, 43, 537–546. [Google Scholar] [CrossRef]
- Fernández-Quintela, A.; Macarulla, M.T.; Del Barrio, A.S.; Martínez, J.A. Composition and Functional Properties of Protein Isolates Obtained from Commercial Legumes Grown in Northern Spain. Plant Foods Hum. Nutr. 1997, 51, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Ray, H.; Georges, F. A Genomic Approach to Nutritional, Pharmacological and Genetic Issues of Faba Bean (Vicia faba): Prospects for Genetic Modifications. GM Crops 2010, 1, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.H.C. Oleosins and Oil Bodies in Seeds and Other Organs. Plant Physiol. 1996, 110, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, K.L.; Vickers, Z.M. Taste Intensities of Oil-in-Water Emulsions with Varying Fat Content. J. Sens. Stud. 2002, 17, 379–390. [Google Scholar] [CrossRef]
- Lockyer, S.; Nugent, A.P. Health Effects of Resistant Starch. Nutr. Bull. 2017, 42, 10–41. [Google Scholar] [CrossRef]
- Chambers, L.; McCrickerd, K.; Yeomans, M.R. Optimising Foods for Satiety. Trends Food Sci. Technol. 2015, 41, 149–160. [Google Scholar] [CrossRef]
- Howarth, N.C.; Saltzman, E.; Roberts, S.B. Dietary Fiber and Weight Regulation. Nutr. Rev. 2001, 59, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Crépon, K.; Marget, P.; Peyronnet, C.; Carrouée, B.; Arese, P.; Duc, G. Nutritional Value of Faba Bean (Vicia faba L.) Seeds for Feed and Food. F. Crop. Res. 2010, 115, 329–339. [Google Scholar] [CrossRef]
- Luzzatto, L.; Arese, P. Favism and Glucose-6-Phosphate Dehydrogenase Deficiency. N. Engl. J. Med. 2018, 378, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, R.R.; Muduuli, D.S.; Frohlich, A.A. Purification and Some Properties of Vicine and Convicine Isolated from Faba Bean (Vicia faba L.) Protein Concentrate. J. Agric. Food Chem. 1983, 31, 839–844. [Google Scholar] [CrossRef]
- Delfini, M.; Di Nola, A.; Carnovale, E.; Lepri, A.; Gaggelli, E.; Russo, N. 13C NMR Study of the Natural Glycosides Vicine and Convicine. Spectrosc. Lett. 1990, 23, 657–667. [Google Scholar] [CrossRef]
- Pulkkinen, M.; Gautam, M.; Lampi, A.M.; Ollilainen, V.; Stoddard, F.; Sontag-Strohm, T.; Salovaara, H.; Piironen, V. Determination of Vicine and Convicine from Faba Bean with an Optimized High-Performance Liquid Chromatographic Method. Food Res. Int. 2015, 76, 168–177. [Google Scholar] [CrossRef]
- Luo, Y.W.; Xie, W.H. Effect of Different Processing Methods on Certain Antinutritional Factors and Protein Digestibility in Green and White Faba Bean (Vicia faba L.). CYTA-J. Food 2013, 11, 43–49. [Google Scholar] [CrossRef]
- Carnovale, E.; Lugaro, E.; Lombardi-Boccia, G. Phytic Acid in Faba Bean and Pea: Effect on Protein Availability. Cereal Chem. 1988, 65, 114–117. [Google Scholar]
- Luo, Y.; Gu, Z.; Han, Y.; Chen, Z. The Impact of Processing on Phytic Acid, in Vitro Soluble Iron and Phy/Fe Molar Ratio of Faba Bean (Vicia faba L.). J. Sci. Food Agric. 2009, 89, 861–866. [Google Scholar] [CrossRef]
- Dhull, S.B.; Kidwai, M.K.; Noor, R.; Chawla, P.; Rose, P.K. A Review of Nutritional Profile and Processing of Faba Bean (Vicia faba L.). Legum. Sci. 2021, 4, e129. [Google Scholar] [CrossRef]
- Liener, I.E. Implications of Antinutritional Components in Soybean Foods. Crit. Rev. Food Sci. Nutr. 1994, 34, 31–67. [Google Scholar] [CrossRef] [PubMed]
- Fekadu Gemede, H. Antinutritional Factors in Plant Foods: Potential Health Benefits and Adverse Effects. Int. J. Nutr. Food Sci. 2014, 3, 284. [Google Scholar] [CrossRef]
- Kareem, O.; Ali, T.; Dar, L.A.; Mir, S.A.; Rashid, R.; Nazli, N.; Gulzar, T.; Bader, G.N. Positive Health Benefits of Saponins from Edible Legumes: Phytochemistry and Pharmacology. In Edible Plants in Health and Diseases; Masoodi, M.H., Rehman, M.U., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2022; Volume II, pp. 279–289. ISBN 9789811649585. [Google Scholar]
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. The Biological Action of Saponins in Animal Systems: A Review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Milgate, J.; Roberts, D.C.K. The Nutritional & Biological Significance of Saponins. Nutr. Res. 1995, 15, 1223–1249. [Google Scholar] [CrossRef]
- Güçlü-Üstündağ, Ö.; Mazza, G. Saponins: Properties, Applications and Processing. Crit. Rev. Food Sci. Nutr. 2007, 47, 231–258. [Google Scholar] [CrossRef] [PubMed]
- Barros, F.; Awika, J.M.; Rooney, L.W. Interaction of Tannins and Other Sorghum Phenolic Compounds with Starch and Effects on in Vitro Starch Digestibility. J. Agric. Food Chem. 2012, 60, 11609–11617. [Google Scholar] [CrossRef] [PubMed]
- Avilés-Gaxiola, S.; Chuck-Hernández, C.; Serna Saldívar, S.O. Inactivation Methods of Trypsin Inhibitor in Legumes: A Review. J. Food Sci. 2018, 83, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Losso, J.N. The Biochemical and Functional Food Properties of the Bowman-Birk Inhibitor. Crit. Rev. Food Sci. Nutr. 2008, 48, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Mu, K.; Arntfield, S.D.; Nickerson, M.T. Changes in Levels of Enzyme Inhibitors during Soaking and Cooking for Pulses Available in Canada. J. Food Sci. Technol. 2017, 54, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Duijsens, D.; Gwala, S.; Pallares, A.P.; Pälchen, K.; Hendrickx, M.; Grauwet, T. How Postharvest Variables in the Pulse Value Chain Affect Nutrient Digestibility and Bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5067–5096. [Google Scholar] [CrossRef]
- Liu, K. Method Development and Optimization for Measuring Chymotrypsin and Chymotrypsin Inhibitor Activities. J. Food Sci. 2022, 87, 2018–2033. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.Y.; Ng, T.B.; Rao, P.F. A Bowman-Birk-Type Trypsin-Chymotrypsin Inhibitor from Broad Beans. Biochem. Biophys. Res. Commun. 2001, 289, 91–96. [Google Scholar] [CrossRef]
- Aguilera, Y.; Esteban, R.M.; Benítez, V.; Mollá, E.; Martín-Cabrejas, M.A. Starch, Functional Properties, and Microstructural Characteristics in Chickpea and Lentil as Affected by Thermal Processing. J. Agric. Food Chem. 2009, 57, 10682–10688. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.R.; Sorgentini, D.A.; Anon, M.C. Relation between Solubility and Surface Hydrophobicity as an Indicator of Modifications during Preparation Processes of Commercial and Laboratory-Prepared Soy Protein Isolates. J. Agric. Food Chem. 2000, 48, 3159–3165. [Google Scholar] [CrossRef] [PubMed]
- van Gils, J.H.M.; Gogishvili, D.; van Eck, J.; Bouwmeester, R.; van Dijk, E.; Abeln, S. How Sticky Are Our Proteins? Quantifying Hydrophobicity of the Human Proteome. Bioinforma. Adv. 2022, 2, vbac002. [Google Scholar] [CrossRef] [PubMed]
- Dill, K.A. Theory for the folding and stability of globular proteins. Biochemistry 1985, 24, 1501–1509. [Google Scholar] [CrossRef] [PubMed]
- Dill, K.A. Dominant Forces in Protein Folding. Biochemistry 1990, 29, 7133–7155. [Google Scholar] [CrossRef]
- Arntfield, S.D.; Murray, E.D. The Influence of Processing Parameters on Food Protein Functionality I. Differential Scanning Calorimetry as an Indicator of Protein Denaturation. Can. Inst. Food Sci. Technol. J. 1981, 14, 289–294. [Google Scholar] [CrossRef]
- Majzoobi, M.; Abedi, E.; Farahnaky, A.; Aminlari, M. Functional Properties of Acetylated Glutenin and Gliadin at Varying PH Values. Food Chem. 2012, 133, 1402–1407. [Google Scholar] [CrossRef]
- Geerts, M.E.J.; Dekkers, B.L.; van der Padt, A.; van der Goot, A.J. Aqueous Fractionation Processes of Soy Protein for Fibrous Structure Formation. Innov. Food Sci. Emerg. Technol. 2018, 45, 313–319. [Google Scholar] [CrossRef]
- Warsame, A.O.; Michael, N.; O’Sullivan, D.M.; Tosi, P. Identification and Quantification of Major Faba Bean Seed Proteins. J. Agric. Food Chem. 2020, 68, 8535–8544. [Google Scholar] [CrossRef]
- Lampi, A.M.; Yang, Z.; Mustonen, O.; Piironen, V. Potential of Faba Bean Lipase and Lipoxygenase to Promote Formation of Volatile Lipid Oxidation Products in Food Models. Food Chem. 2020, 311, 125982. [Google Scholar] [CrossRef] [PubMed]
- Sáenz De Miera, L.E.; Ramos, J.; Pérez De La Vega, M. A Comparative Study of Convicilin Storage Protein Gene Sequences in Species of the Tribe Vicieae. Genome 2008, 51, 511–523. [Google Scholar] [CrossRef]
- Horstmann, C.; Schlesier, B.; Otto, A.; Kostka, S.; Müntz, K. Polymorphism of Legumin Subunits from Field Bean (Vicia faba L. Var. minor) and Its Relation to the Corresponding Multigene Family. Theor. Appl. Genet. 1993, 86, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed Storage Proteins: Structures and Biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Monge, R.; Lopez-Torrejón, G.; Pascual, C.Y.; Varela, J.; Martin-Esteban, M.; Salcedo, G. Vicilin and Convicilin Are Potential Major Allergens from Pea. Clin. Exp. Allergy 2004, 34, 1747–1753. [Google Scholar] [CrossRef]
- Loveday, S.M. Food Proteins: Technological, Nutritional, and Sustainability Attributes of Traditional and Emerging Proteins. Annu. Rev. Food Sci. Technol. 2019, 10, 311–339. [Google Scholar] [CrossRef]
- Longstaff, M.; McNAB, J.M. The Inhibitory Effects of Hull Polysaccharides and Tannins of Field Beans (Vicia faba L.) on the Digestion of Amino Acids, Starch and Lipid and on Digestive Enzyme Activities in Young Chicks. Br. J. Nutr. 1991, 65, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Wichers, H.J.; Hettinga, K.A. Heat-Induced Unfolding Facilitates Plant Protein Digestibility during in Vitro Static Infant Digestion. Food Chem. 2022, 375, 131878. [Google Scholar] [CrossRef]
- Ma, R.; Lin, Z.; Wu, Y.; Gao, Z.; Hu, B.; Xu, L.; Fang, Y.; Nishinari, K. Modulating the in Vitro Gastric Digestion of Heat-Induced Beta-Lactoglobulin Aggregates: Incorporation with Polysaccharide. Food Chem. 2021, 354, 129506. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.J.; Chen, X.D.; Pearce, D. Surface Composition of Industrial Spray-Dried Milk Powders. 2. Effects of Spray Drying Conditions on the Surface Composition. J. Food Eng. 2009, 94, 169–181. [Google Scholar] [CrossRef]
- Amagliani, L.; O’Regan, J.; Kelly, A.L.; O’Mahony, J.A. Physical and Flow Properties of Rice Protein Powders. J. Food Eng. 2016, 190, 1–9. [Google Scholar] [CrossRef]
- Alamilla-Beltrán, L.; Chanona-Pérez, J.J.; Jiménez-Aparicio, A.R.; Gutiérrez-Lopez, G.F. Description of Morphological Changes of Particles along Spray Drying. J. Food Eng. 2005, 67, 179–184. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; Hoover, R.; Donner, E.; Liu, Q.; Jaiswal, S.; Chibbar, R.; Nantanga, K.K.M.; Seetharaman, K. Structure of Faba Bean, Black Bean and Pinto Bean Starches at Different Levels of Granule Organization and Their Physicochemical Properties. Food Res. Int. 2011, 44, 2962–2974. [Google Scholar] [CrossRef]
- Blennow, A.; Hansen, M.; Schulz, A.; Jørgensen, K.; Donald, A.M.; Sanderson, J. The Molecular Deposition of Transgenically Modified Starch in the Starch Granule as Imaged by Functional Microscopy. J. Struct. Biol. 2003, 143, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Glaring, M.A.; Koch, C.B.; Blennow, A. Genotype-Specific Spatial Distribution of Starch Molecules in the Starch Granule: A Combined CLSM and SEM Approach. Biomacromolecules 2006, 7, 2310–2320. [Google Scholar] [CrossRef] [PubMed]
- Nasar-Abbas, S.M.; Siddique, K.H.M.; Plummer, J.A.; White, P.F.; Harris, D.; Dods, K.; D’Antuono, M. Faba Bean (Vicia faba L.) Seeds Darken Rapidly and Phenolic Content Falls When Stored at Higher Temperature, Moisture and Light Intensity. LWT 2009, 42, 1703–1711. [Google Scholar] [CrossRef]
- Al-Dabbas, M.M.; Al-Ismail, K.; Taleb, R.A.; Ibrahim, S. Acid-Base Buffering Properties of Five Legumes and Selected Food in Vitro. Am. J. Agric. Biol. Sci. 2010, 5, 154–160. [Google Scholar] [CrossRef]
- Chung, C.; Sher, A.; Rousset, P.; Decker, E.A.; McClements, D.J. Formulation of Food Emulsions Using Natural Emulsifiers: Utilization of Quillaja Saponin and Soy Lecithin to Fabricate Liquid Coffee Whiteners. J. Food Eng. 2017, 209, 1–11. [Google Scholar] [CrossRef]
- Narsimhan, G.; Xiang, N. Role of Proteins on Formation, Drainage, and Stability of Liquid Food Foams. Annu. Rev. Food Sci. Technol. 2018, 9, 45–63. [Google Scholar] [CrossRef] [PubMed]
- Amagliani, L.; Schmitt, C. Globular Plant Protein Aggregates for Stabilization of Food Foams and Emulsions. Trends Food Sci. Technol. 2017, 67, 248–259. [Google Scholar] [CrossRef]
- Munialo, C.D.; Martin, A.H.; Van Der Linden, E.; De Jongh, H.H.J. Fibril Formation from Pea Protein and Subsequent Gel Formation. J. Agric. Food Chem. 2014, 62, 2418–2427. [Google Scholar] [CrossRef] [PubMed]
- Makri, E.; Papalamprou, E.; Doxastakis, G. Study of Functional Properties of Seed Storage Proteins from Indigenous European Legume Crops (Lupin, Pea, Broad Bean) in Admixture with Polysaccharides. Food Hydrocoll. 2005, 19, 583–594. [Google Scholar] [CrossRef]
- Cornet, S.H.V.; Snel, S.J.E.; Lesschen, J.; van der Goot, A.J.; van der Sman, R.G.M. Enhancing the Water Holding Capacity of Model Meat Analogues through Marinade Composition. J. Food Eng. 2021, 290, 110283. [Google Scholar] [CrossRef]
- Grasso, N.; Alonso-Miravalles, L.; O’Mahony, J.A. Composition, Physicochemical and Sensorial Properties of Commercial Plant-Based Yogurts. Foods 2020, 9, 252. [Google Scholar] [CrossRef] [PubMed]
- Escamilla-Silva, E.M.; Guzmán-Maldonado, S.H.; Cano-Medinal, A.; González-Alatorre, G. Simplified Process for the Production of Sesame Protein Concentrate. Differential Scanning Calorimetry and Nutritional, Physicochemical and Functional Properties. J. Sci. Food Agric. 2003, 83, 972–979. [Google Scholar] [CrossRef]
- Zayas, J.F. Oil and Fat Binding Properties Of Proteins. In Functionality of Proteins in Food; Zayas, J.F., Ed.; Springer: New York, NY, USA, 1997; pp. 228–259. ISBN 9783642624759. [Google Scholar]
- Kinsella, J.E. Functional Properties of Soy Proteins. J. Am. Oil Chem. Soc. 1979, 56, 242–258. [Google Scholar] [CrossRef]
- Sathe, S.K.; Deshpande, S.S.; Salunkhe, D.K. Functional Properties of Winged Bean [Psophocarpus tetragonolobus (L.) DC] Proteins. J. Food Sci. 1982, 47, 503–509. [Google Scholar] [CrossRef]
- Kinsella, J.E. Functional Properties of Proteins in Foods: A Survey. Crit. Rev. Food Sci. Nutr. 1976, 7, 219–280. [Google Scholar] [CrossRef]
- Zhang, Z.; Tian, X.; Wang, P.; Jiang, H.; Li, W. Compositional, Morphological, and Physicochemical Properties of Starches from Red Adzuki Bean, Chickpea, Faba Bean, and Baiyue Bean Grown in China. Food Sci. Nutr. 2019, 7, 2485–2494. [Google Scholar] [CrossRef] [PubMed]
- Blackwood, A.D.; Salter, J.; Dettmar, P.W.; Chaplin, M.F. Dietary Fibre, Physicochemical Properties and Their Relationship to Health. J. R. Soc. Promot. Health 2000, 120, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Rosell, C.M.; Rojas, J.A.; Benedito de Barber, C. Influence of Hydrocolloids on Dough Rheology and Bread Quality. Food Hydrocoll. 2001, 15, 75–81. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PI 1 | PI 2 | SF | FRF | |
|---|---|---|---|---|
| Protein | 72.64 ± 0.31 a | 71.37 ± 0.93 a | 0.18 ± 0.01 c | 16.70 ± 0.08 b |
| Fat 1 | 6.24 ± 0.18 a | 0.78 ± 0.04 b | n.d. | n.d. |
| Digestible Starch | 5.32 ± 0.28 c | 1.00 ± 0.07 d | 28.46 ± 0.37 a | 15.06 ± 0.22 b |
| Resistant Starch 2 | 0.33 ± 0.01 c | n.d. | 55.41 ± 3.31 a | 9.70 ± 0.44 b |
| Σ Total Starch | 5.65 ± 0.27 c | 1.00 ± 0.07 d | 83.87 ± 3.07 a | 24.76 ± 0.61 b |
| Total Dietary Fibre | 8.69 ± 0.67 c | 7.64 ± 1.53 c | 62.28 ± 0.76 b | 69.09 ± 0.24 a |
| FODMAPs 3 | n.d. | 0.07 ± 0.00 a | 0.06 ± 0.00 a | n.d. |
| PI 1 | PI 2 | SF | FRF | |
|---|---|---|---|---|
| Vicine (mg/g DM) | n.d. 1 | 2.18 ± 0.11 a | n.d. 1 | n.d. 1 |
| Convicine (mg/g DM) | 0.30 ± 0.03 a | 1.52 ± 0.19 b | n.d. 1 | n.d. 1 |
| Phytic acid (g/100 g DM) | 0.39 ± 0.04 a | 0.14 ± 0.02 b | 0.008 ± 0.011 c | 0.16 ± 0.01 b |
| Saponins (mg/g DM) | n.d. 1 | n.d. 1 | 0.97 ± 0.08 a | 4.94 ± 0.08 b |
| Condensed tannins (CE mg/g DM) | 7.19 ± 1.45 a | n.d. 1 | 3.23 ± 0.54 a | 0.55 ± 0.00 b |
| Trypsin inhibitors (TIU/mg DM) | 2.72 ± 0.04 a | 206.73 ± 12.01 b | n.d. 1 | 3.38 ± 0.08 c |
| Chymotrypsin inhibitors (CIU/mg DM) | 1.65 ± 0.76 a | 14.61 ± 1.11 b | 0.85 ± 0.10 a | 1.63 ± 0.07 a |
| Protein Isolates | Pepsin Digestibility (1 h) | Total Digestibility (2 h) |
|---|---|---|
| PI 1 | 5.31 ± 0.03 a | 23.30 ± 1.18 a |
| PI 2 | 1.98 ± 0.29 b | 9.59 ± 0.13 a |
| BSA (reference) | 5.75 ± 0.11 c | 23.85 ± 0.15 a |
| PI 1 | PI 2 | SF | FRF | ||
|---|---|---|---|---|---|
| WI | (%) | 81.21 ± 0.95 c | 88.10 ± 0.21 b | 96.52 ± 0.50 a | 80.21 ± 0.74 d |
| pH | (-) | 5.14 ± 0.04 c | 4.57 ± 0.00 d | 8.02 ± 0.06 a | 6.36 ± 0.02 b |
| TTA | (mL/g) | 4.80 ± 0.05 b | 6.67 ± 0.55 a | 0.02 ± 0.00 d | 0.45 ± 0.01 c |
| FC | (%) | 18.06 ± 2.41 b | 133.33 ± 15.73 a | 0.00 ± 0.00 d | 7.92 ± 1.34 c |
| FS | (%) | 70.00 ± 8.66 a | 39.79 ± 4.09 b | - 2 | 0.00 ± 0.00 c |
| WBC | (%) | 149.80 ± 0.61 b | - 1 | 63.67 ± 1.28 c | 1028.92 ± 56.90 a |
| OBC | (%) | 65.32 ± 0.33 c | 229.13 ± 4.42 b | 49.43 ± 1.55 d | 342.35 ± 4.96 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krause, M.; Sørensen, J.C.; Petersen, I.L.; Duque-Estrada, P.; Cappello, C.; Tlais, A.Z.A.; Di Cagno, R.; Ispiryan, L.; Sahin, A.W.; Arendt, E.K.; et al. Associating Compositional, Nutritional and Techno-Functional Characteristics of Faba Bean (Vicia faba L.) Protein Isolates and Their Production Side-Streams with Potential Food Applications. Foods 2023, 12, 919. https://doi.org/10.3390/foods12050919
Krause M, Sørensen JC, Petersen IL, Duque-Estrada P, Cappello C, Tlais AZA, Di Cagno R, Ispiryan L, Sahin AW, Arendt EK, et al. Associating Compositional, Nutritional and Techno-Functional Characteristics of Faba Bean (Vicia faba L.) Protein Isolates and Their Production Side-Streams with Potential Food Applications. Foods. 2023; 12(5):919. https://doi.org/10.3390/foods12050919
Chicago/Turabian StyleKrause, Magdalena, Jens Christian Sørensen, Iben Lykke Petersen, Patrícia Duque-Estrada, Claudia Cappello, Ali Zein Alabiden Tlais, Raffaella Di Cagno, Lilit Ispiryan, Aylin W. Sahin, Elke K. Arendt, and et al. 2023. "Associating Compositional, Nutritional and Techno-Functional Characteristics of Faba Bean (Vicia faba L.) Protein Isolates and Their Production Side-Streams with Potential Food Applications" Foods 12, no. 5: 919. https://doi.org/10.3390/foods12050919
APA StyleKrause, M., Sørensen, J. C., Petersen, I. L., Duque-Estrada, P., Cappello, C., Tlais, A. Z. A., Di Cagno, R., Ispiryan, L., Sahin, A. W., Arendt, E. K., & Zannini, E. (2023). Associating Compositional, Nutritional and Techno-Functional Characteristics of Faba Bean (Vicia faba L.) Protein Isolates and Their Production Side-Streams with Potential Food Applications. Foods, 12(5), 919. https://doi.org/10.3390/foods12050919