
Network Pharmacology Combined with an Experimental Validation Study to Reveal the Effect and Mechanism of Eucommia ulmoides Leaf Polysaccharide against Immunomodulation

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of ELP
2.2. Physiochemical Analysis of Eucommia ulmoides Leaf Polysaccharide
2.3. Phagocytic Activity of Macrophages
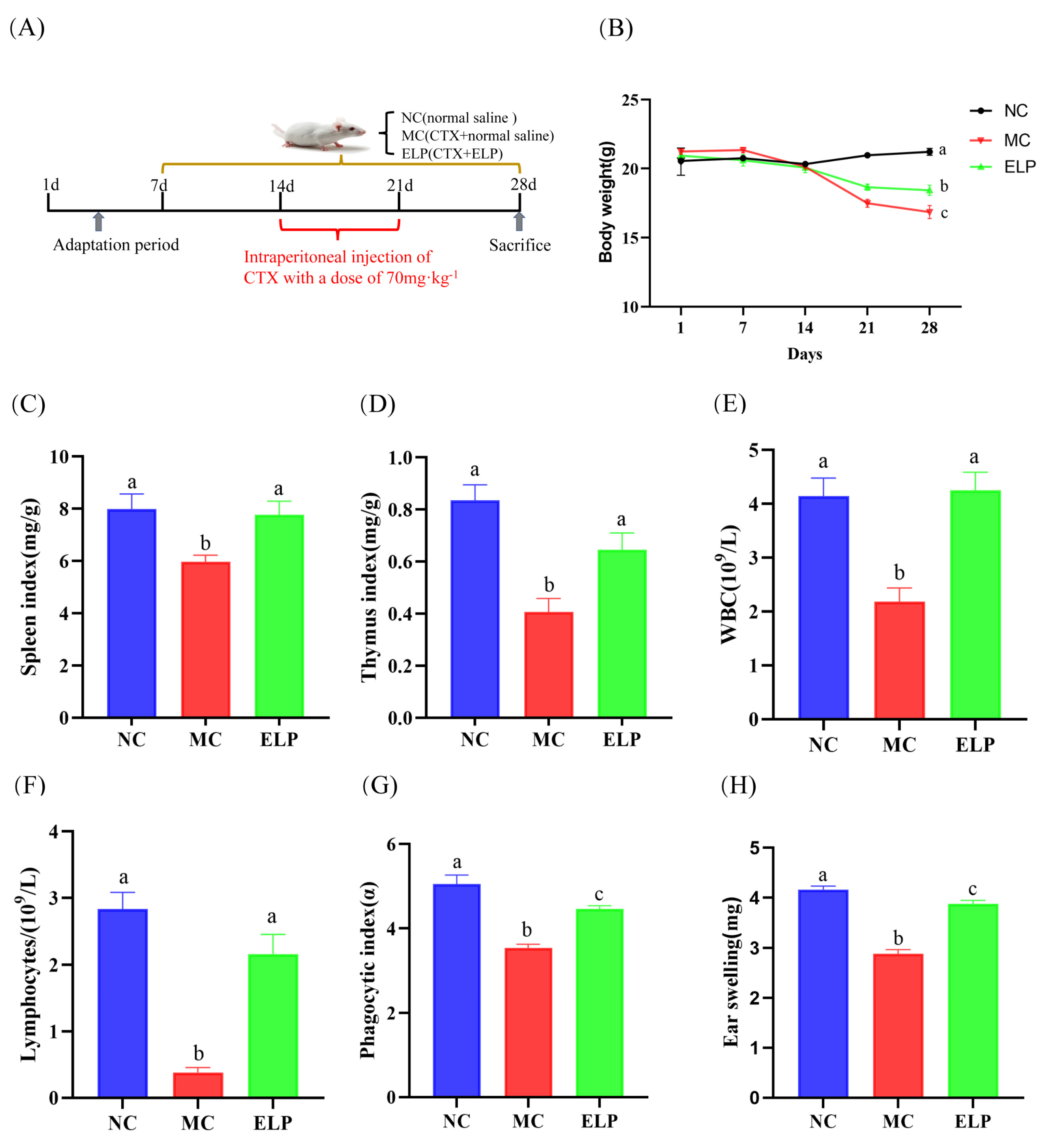
2.4. Experimental Design and Drug Administration
2.5. Determination of Immune Organs Indices
2.6. Histological Examinations of the Liver and Spleen
2.7. Determination of Cytokines in Serum
2.8. Peripheral Blood Leukocyte and Lymphocyte Counts
2.9. Determination of the Phagocytic Index
2.10. Delayed-Type Hypersensitivity (DTH) Test
2.11. qRT-PCR Analysis
2.12. Western Blot Assay
2.13. Network Pharmacology Analysis
2.14. Statistical Analysis
3. Results
3.1. Physicochemical Properties of Eucommia ulmoides Leaf Polysaccharide
3.2. Effects of ELP on Immunoregulation In Vitro
3.2.1. Effect of ELP in Macrophage Proliferation of Mice
3.2.2. Effect of ELP on Macrophage Phagocytosis in Mice
3.3. Effects of ELP on Immunoregulation In Vivo
3.3.1. Effects of ELP on Immune Organ Indexes
3.3.2. Peripheral Blood White Blood Cells (WBCs) and Lymphocytes
3.3.3. Phagocytic Index
3.3.4. DTH Reaction
3.3.5. Histological Analysis of Spleen and Liver
3.3.6. Effects of ELP on Serum Cytokine Levels
3.3.7. KEGG Pathway Enrichment
3.3.8. Effects of ELP on Gene Expression
3.3.9. Western Blot Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shyer, J.A.; Flavell, R.A.; Bailis, W. Metabolic signaling in T cells. Cell Res. 2020, 30, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Zhang, Y.; Chen, W.; Jia, R.; Yu, X.; Wang, Y.; Li, Y.; Liu, Y.; Ye, X.; Yu, L.; et al. Apios americana Medik flowers polysaccharide (AFP) alleviate Cyclophosphamide-induced immunosuppression in ICR mice. Int. J. Biol. Macromol. 2020, 144, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Palomares, O.; Akdis, M.; Martin-Fontecha, M.; Akdis, C.A. Mechanisms of immune regulation in allergic diseases: The role of regulatory T and B cells. Immunol. Rev. 2017, 278, 219–236. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Dai, Y.; Yuan, G.; Su, J.; Liu, X. Immunomodulatory Effects and Induction of Apoptosis by Different Molecular Weight Chitosan Oligosaccharides in Head Kidney Macrophages From Blunt Snout Bream (Megalobrama amblycephala). Front. Immunol. 2019, 10, 869. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gomez, A.; Carretero, M.; Weber, N.; Peterka, V.; To, A.; Titova, V.; Solis, G.; Osborn, O.; Petrascheck, M. A phenotypic Caenorhabditis elegans screen identifies a selective suppressor of antipsychotic-induced hyperphagia. Nat. Commun. 2018, 9, 5272. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Nan, L.; Xiao, C.; Ji, Q.; Li, K.; Wei, Q.; Liu, Y.; Bao, G. Optimum Preparation Method for Self-Assembled PEGylation Nano-Adjuvant Based on Rehmannia glutinosa Polysaccharide and Its Immunological Effect on Macrophages. Int. J. Nanomed. 2019, 14, 9361–9375. [Google Scholar] [CrossRef]
- Huang, L.; Lyu, Q.; Zheng, W.; Yang, Q.; Cao, G. Traditional application and modern pharmacological research of Eucommia ulmoides Oliv. Chin. Med. 2021, 16, 73. [Google Scholar] [CrossRef]
- Wang, C.Y.; Tang, L.; He, J.W.; Li, J.; Wang, Y.Z. Ethnobotany, Phytochemistry and Pharmacological Properties of Eucommia ulmoides: A Review. Am. J. Chin. Med. 2019, 47, 259–300. [Google Scholar] [CrossRef]
- Zhu, M.Q.; Sun, R.C. Eucommia ulmoides Oliver: A Potential Feedstock for Bioactive Products. J. Agric. Food Chem. 2018, 66, 5433–5438. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Li, Y.; Song, M.; Li, Y.; Yang, A.; Zhang, Y.; Wang, D.; Hu, M. Aucubin slows the development of osteoporosis by inhibiting osteoclast differentiation via the nuclear factor erythroid 2-related factor 2-mediated antioxidation pathway. Pharm. Biol. 2021, 59, 1556–1565. [Google Scholar] [CrossRef]
- He, X.; Wang, J.; Li, M.; Hao, D.; Yang, Y.; Zhang, C.; He, R.; Tao, R. Eucommia ulmoides Oliv.: Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2014, 151, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.Y.; Wang, J.Y.; Wang, K.; Zhang, Y.; Liu, K.; Chen, X.Y.; Yuan, Y. Inhibition of Rheumatoid Arthritis Using Bark, Leaf, and Male Flower Extracts of Eucommia ulmoides. Evid. Based Complement Altern. Med. 2020, 2020, 3260278. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.B.; Yang, J.; Zhi, H.; Hu, X.; Yang, Y.; Zhang, L.Z.; Liu, Q.Q.; Feng, Y.Y.; Wu, D.Y.; Li, H.Y. Eucommia ulmoides Leaf Polysaccharide in Conjugation with Ovalbumin Act as Delivery System Can Improve Immune Response. Pharmaceutics 2021, 13, 1384. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.P.; Lu, W.; Ku, K.M.; Zhang, L.H.; Lei, L.L.; Zong, W. Ultrasonic-assisted extraction and antioxidant activity of polysaccharides from Eucommia ulmoides leaf. Pak. J. Pharm. Sci. 2020, 33, 581–588. [Google Scholar] [CrossRef]
- Kong, Y.F.; Hu, Y.L.; Li, J.M.; Cai, J.T.; Qiu, Y.H.; Dong, C.H. Anti-inflammatory Effect of a Novel Pectin Polysaccharide From Rubus chingii Hu on Colitis Mice. Front. Nutr. 2022, 9, 868657. [Google Scholar] [CrossRef]
- Guo, L.; Dai, H.W.; Ma, J.Y.; Wang, J.M.; Hua, Y.; Zhou, L.Y. Isolation, structure characteristics and antioxidant activity of two water-soluble polysaccharides from Lenzites betulina. BMC Chem. 2021, 15, 19. [Google Scholar] [CrossRef]
- Chen, W.X.; Zhang, W.Y.; Shen, W.B.; Wang, K.C. Effects of the acid polysaccharide fraction isolated from a cultivated Cordyceps sinensis on macrophages in vitro. Cell. Immunol. 2010, 262, 69–74. [Google Scholar] [CrossRef]
- Tang, C.; Sun, J.; Zhou, B.; Jin, C.H.; Liu, J.; Kan, J.; Qian, C.L.; Zhang, N.F. Effects of polysaccharides from purple sweet potatoes on immune response and gut microbiota composition in normal and cyclophosphamide treated mice. Food Funct. 2018, 9, 937–950. [Google Scholar] [CrossRef]
- Lee, G.H.; Lee, H.Y.; Park, S.A.; Shin, T.S.; Chae, H.J. Eucommia ulmoides Leaf Extract Ameliorates Steatosis Induced by High-fat Diet in Rats by Increasing Lysosomal Function. Nutrients 2019, 11, 426. [Google Scholar] [CrossRef]
- Zhao, Z.; Yu, Z.; Hou, Y.; Zhang, L.; Fu, A. Improvement of cognitive and motor performance with mitotherapy in aged mice. Int. J. Biol. Sci. 2020, 16, 849–858. [Google Scholar] [CrossRef]
- Wang, H.; Yang, S.; Wang, Y.; Jiang, T.; Li, S.; Lv, Z. Immunoenhancement Effects of Glycosaminoglycan from Apostichopus japonicus: In Vitro and In Cyclophosphamide-Induced Immunosuppressed Mice Studies. Mar. Drugs 2017, 15, 347. [Google Scholar] [CrossRef]
- Qu, D.; Hu, H.; Lian, S.; Sun, W.; Si, H. The Protective Effects of Three Polysaccharides From Abrus cantoniensis Against Cyclophosphamide-Induced Immunosuppression and Oxidative Damage. Front. Vet. Sci. 2022, 9, 870042. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhong, L.; Li, G.; Huang, R.; Wang, Q.; Wang, Z.; Zhang, C.; Chen, B.; Jiang, T.; Zhang, W. Pre-treatment neutrophils count as a prognostic marker to predict chemotherapeutic response and survival outcomes in glioma: A single-center analysis of 288 cases. Am. J. Transl. Res. 2020, 12, 90–104. [Google Scholar] [PubMed]
- Mittendorf, E.A.; Ardavanis, A.; Symanowski, J.; Murray, J.L.; Shumway, N.M.; Litton, J.K.; Hale, D.F.; Perez, S.A.; Anastasopoulou, E.A.; Pistamaltzian, N.F.; et al. Primary analysis of a prospective, randomized, single-blinded phase II trial evaluating the HER2 peptide AE37 vaccine in breast cancer patients to prevent recurrence. Ann. Oncol. 2016, 27, 1241–1248. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Rabbani, S.I.; Asdaq, S.M.B.; Mohzari, Y.; Alrashed, A.; Najib Alajami, H.; Othman Aljohani, A.; Ali Al Mushtawi, A.; Sultan Alenazy, M.; Fahad Alamer, R.; et al. Morus alba Prevented the Cyclophosphamide Induced Somatic and Germinal Cell Damage in Male Rats by Ameliorating the Antioxidant Enzyme Levels. Molecules 2021, 26, 1226. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Du, M.; Zhou, T.Y.; Ai, W.Z.; Zhang, Z.S.; Xiang, X.W.; Zhou, Y.F.; Wen, Z.S. Polysaccharides from Hemp Seed Protect against Cyclophosphamide-Induced Intestinal Oxidative Damage via Nrf2-Keap1 Signaling Pathway in Mice. Oxidative Med. Cell. Longev. 2020, 2020, 1813798. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wan, F.; Zhao, L.; Meng, X.; Li, B. Potential Immunomodulatory Activity of a Selected Strain Bifidobacterium bifidum H3-R2 as Evidenced in vitro and in Immunosuppressed Mice. Front. Microbiol. 2020, 11, 2089. [Google Scholar] [CrossRef]
- Torretta, S.; Scagliola, A.; Ricci, L.; Mainini, F.; Di Marco, S.; Cuccovillo, I.; Kajaste-Rudnitski, A.; Sumpton, D.; Ryan, K.M.; Cardaci, S. D-mannose suppresses macrophage IL-1beta production. Nat. Commun. 2020, 11, 6343. [Google Scholar] [CrossRef]
- Wang, S.; Huang, S.; Ye, Q.H.; Zeng, X.F.; Yu, H.T.; Qi, D.S.; Qiao, S.Y. Prevention of Cyclophosphamide-Induced Immunosuppression in Mice with the Antimicrobial Peptide Sublancin. J. Immunol. Res. 2018, 2018, 4353580. [Google Scholar] [CrossRef]
- Lin, S.; Yan, Y.Y.; Wu, Y.L.; Wang, M.F.; Zhu, Y.Y.; Wang, X.Z. Development of a novel score for the diagnosis of bacterial infection in patients with acute-on-chronic liver failure. World J. Gastroenterol. 2020, 26, 4857–4865. [Google Scholar] [CrossRef]
- Gurven, M.; Kraft, T.S.; Alami, S.; Adrian, J.C.; Linares, E.C.; Cummings, D.; Rodriguez, D.E.; Hooper, P.L.; Jaeggi, A.V.; Gutierrez, R.Q.; et al. Rapidly declining body temperature in a tropical human population. Sci. Adv. 2020, 6, eabc6599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhu, J.H.; Xu, H.; Huang, X.P.; Liu, X.D.; Deng, C.Q. Five Active Components Compatibility of Astragali Radix and Angelicae Sinensis Radix Protect Hematopoietic Function Against Cyclophosphamide-Induced Injury in Mice and t-BHP-Induced Injury in HSCs. Front. Pharmacol. 2019, 10, 936. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, Y.; Bo, J.; Han, W.; Wang, Y.; Zhang, L.; Wu, X.; Yu, S.; Liu, R. Autologous cytokine-induced killer (CIK) cell immunotherapy combined with cyclophosphamide in five patients with POEMS syndrome. Clin. Exp. Immunol. 2016, 184, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Artaza-Irigaray, C.; Molina-Pineda, A.; Aguilar-Lemarroy, A.; Ortiz-Lazareno, P.; Limon-Toledo, L.P.; Pereira-Suarez, A.L.; Rojo-Contreras, W.; Jave-Suarez, L.F. E6/E7 and E6(*) From HPV16 and HPV18 Upregulate IL-6 Expression Independently of p53 in Keratinocytes. Front. Immunol. 2019, 10, 1676. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.B.; Yang, W.T.; Shi, C.W.; Feng, B.; Huang, K.Y.; Zhao, G.X.; Li, Q.Y.; Xie, J.; Huang, H.B.; Jiang, Y.L.; et al. Immune responses induced by recombinant Lactobacillus plantarum expressing the spike protein derived from transmissible gastroenteritis virus in piglets. Appl. Microbiol. Biotechnol. 2018, 102, 8403–8417. [Google Scholar] [CrossRef]
- Sasaki, K.; Himeno, A.; Nakagawa, T.; Sasaki, Y.; Kiyonari, H.; Iwai, K. Modulation of autoimmune pathogenesis by T cell-triggered inflammatory cell death. Nat. Commun. 2019, 10, 3878. [Google Scholar] [CrossRef]
- Yan, H.J.; Jiang, N.; Hu, L.T.; Xu, Q.; Peng, X.D.; Yang, H.; Zhao, W.Y.; Lyu, L.Y.; Wang, L.M.; Che, C.Y. CD23 mediated the induction of pro-inflammatory cytokines Interleukin-1 beta and tumor necrosis factors-alpha in Aspergillus fumigatus keratitis. Chin. Med. J. 2021, 134, 1001–1003. [Google Scholar] [CrossRef]
- Bai, R.B.; Zhang, Y.J.; Fan, J.M.; Jia, X.S.; Li, D.; Wang, Y.P.; Zhou, J.; Yan, Q.; Hu, F.D. Immune-enhancement effects of oligosaccharides from Codonopsis pilosula on cyclophosphamide induced immunosuppression in mice. Food Funct. 2020, 11, 3306–3315. [Google Scholar] [CrossRef]
- Tan, Z.J.; Chen, L.L.; Ren, Y.C.; Jiang, X.H.; Gao, W. Neuroprotective effects of FK866 against traumatic brain injury: Involvement of p38/ERK pathway. Ann. Clin. Transl. Neur. 2020, 7, 742–756. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward Primer | Reverse Primer |
|---|---|---|
| TNF-α | AGTCCGGGCAGGTCTACTTT | GTCACTGTCCCAGCATCTTGT |
| IL-1β | TGACGGACCCCAAAAGATGA | TCTCCACAGCCACAATGAGT |
| IL-6 | CACTTCACAAGTCGGAGGCT | CTGCAAGTGCATCATCGTTGT |
| GAPDH | AGGTTGTCTCCTGCGACTGCA | GTGGTCCAGGGTTTCTTACTCC |
| NO. | Compound | OB/% | DL |
|---|---|---|---|
| MOL000382 | Arabinose | 1.87 | 0.02 |
| MOL011643 | Galacturonic acid | 40.39 | 0.06 |
| MOL000814 | Galactose | 47.94 | 0.04 |
| MOL000424 | Rhamnose | 50.50 | 0.04 |
| MOL000734 | Glucose | 24.44 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, E.; Tang, P.; Zhu, X.; Lv, M.; Wang, S.; Xue, Y.; Li, C.; Zhao, S. Network Pharmacology Combined with an Experimental Validation Study to Reveal the Effect and Mechanism of Eucommia ulmoides Leaf Polysaccharide against Immunomodulation. Foods 2023, 12, 1062. https://doi.org/10.3390/foods12051062
Cui E, Tang P, Zhu X, Lv M, Wang S, Xue Y, Li C, Zhao S. Network Pharmacology Combined with an Experimental Validation Study to Reveal the Effect and Mechanism of Eucommia ulmoides Leaf Polysaccharide against Immunomodulation. Foods. 2023; 12(5):1062. https://doi.org/10.3390/foods12051062
Chicago/Turabian StyleCui, Enhui, Pan Tang, Xiaoyan Zhu, Mengyuan Lv, Shuai Wang, Yuhuan Xue, Cixia Li, and Shanting Zhao. 2023. "Network Pharmacology Combined with an Experimental Validation Study to Reveal the Effect and Mechanism of Eucommia ulmoides Leaf Polysaccharide against Immunomodulation" Foods 12, no. 5: 1062. https://doi.org/10.3390/foods12051062
APA StyleCui, E., Tang, P., Zhu, X., Lv, M., Wang, S., Xue, Y., Li, C., & Zhao, S. (2023). Network Pharmacology Combined with an Experimental Validation Study to Reveal the Effect and Mechanism of Eucommia ulmoides Leaf Polysaccharide against Immunomodulation. Foods, 12(5), 1062. https://doi.org/10.3390/foods12051062