
Antioxidant Activity of Crocodile Oil (Crocodylus siamensis) on Cognitive Function in Rats
 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Fatty Acid Composition
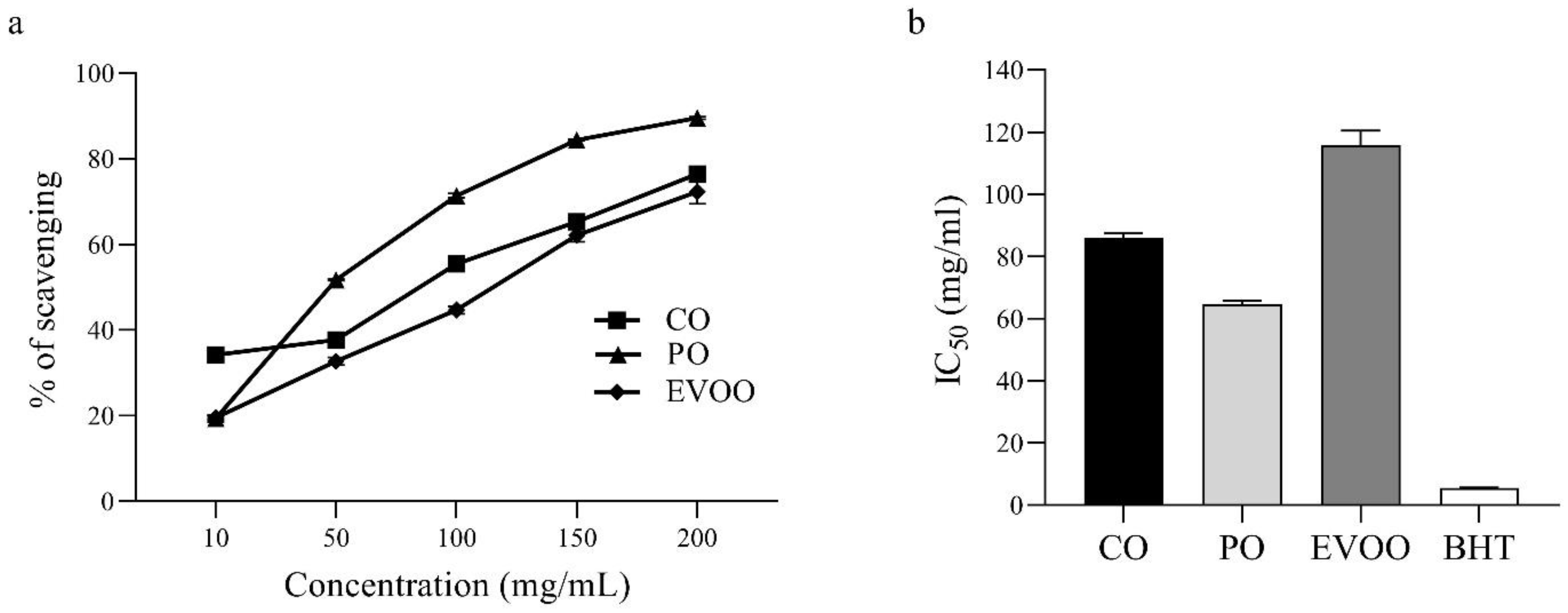
2.3. Free Radicals Scavenging Activity of CO In Vitro
2.4. Animals
2.5. Experimental Design
2.6. Food Intake, Energy Intake, Bodyweight Gain, and Bodyweight of Rats
2.7. Morris Water Maze (MWM) Test
2.8. Sample Collection
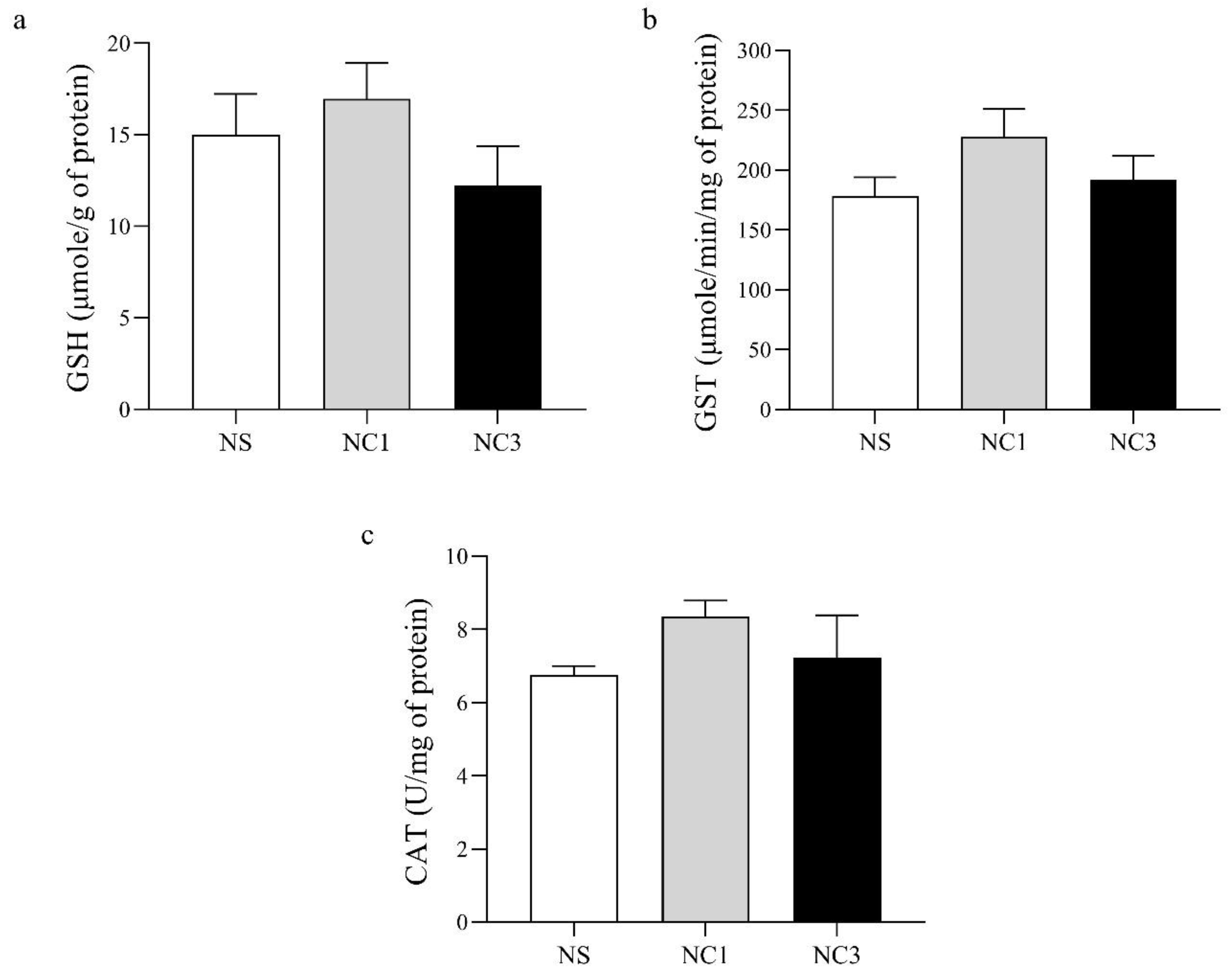
2.9. Antioxidant Markers
2.9.1. GSH
2.9.2. Glutathione-S-Transferase (GST)
2.9.3. Catalase (CAT)
2.10. Proteomic Analysis
2.10.1. Sample Preparation
2.10.2. LC-MS/MS
2.11. Data and Statistical Analysis
3. Results
3.1. Fatty Acid Profiles of CO
3.2. In Vitro Antioxidant Activity of CO Using the DPPH Assay
3.3. Effect of CO on Food Intake, Energy Intake, Body Weight Gain, and Body Weight
3.4. Effect of CO on Serum Lipid Profiles
3.5. Effect of CO on Spatial Learning and Memory
3.6. Effect of CO on Markers of Antioxidant Activity in the Brain
3.7. Effect of CO on Expression of Antioxidant-Related Proteins in the Brain
3.8. Effect of CO on Expression of Cognition-Related Proteins in the Brain
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kanwatanakid-Savini, C.; Pliosungnoen, M.; Pattanavibool, A.; Thorbjarnarson, J.B.; Limlikhitaksorn, C.; Platt, S.G. A survey to determine the conservation status of Siamese crocodiles in Kaeng Krachan national park, Thailand. Herpetol. Conserv. Biol. 2012, 7, 157–168. [Google Scholar]
- Preecharram, S.; Jearranaiprepame, P.; Daduang, S.; Temsiripong, Y.; Somdee, T.; Fukamizo, T.; Svasti, J.; Araki, T.; Thammasirirak, S. Isolation and characterization of crocosin, an antibacterial compound from crocodile (Crocodylus siamensis) plasma. Anim. Sci. J. 2010, 81, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Buthelezi, S.; Southway, C.; Govinden, U.; Bodenstein, J.; Toit, K. An investigation of the antimicrobial and anti-inflammatory activities of crocodile oil. J. Ethnopharmacol. 2012, 143, 325–330. [Google Scholar] [CrossRef]
- Praduptong, A.; Siruntawineti, J.; Chaeychomsri, S.; Srimangkornkaew, P.; Chaeychomsri, W. Acute oral toxicity testing of Siamese crocodile (Crocodylus siamensis) oil in Wistar rats. Biosci. Disc. 2018, 9, 409–415. [Google Scholar]
- Li, H.; Chen, L.; Hu, Y.; Qin, Y.; Liang, G.; Xiong, Y.; Chen, Q. Crocodile oil enhances cutaneous burn wound healing and reduces scar formation in rats. Acad. Emerg. Med. 2012, 19, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, X.; Huang, S.; Xiong, Y.; Zhang, Z.; Zheng, Y.; Chen, Q.; Chen, Q. Repair function of essential oil from Crocodylus Siamensis (Schneider, 1801) on the burn wound healing via up-regulated growth factor expression and anti-inflammatory effect. J. Ethnopharmacol. 2021, 264, 113286. [Google Scholar] [CrossRef] [PubMed]
- Ngernjan, M.; Ontawong, A.; Lailerd, N.; Mengamphan, K.; Sarapirom, S.; Amornlerdpison, D. Crocodile oil modulates inflammation and immune responses in LPS-stimulated RAW 264.7 macrophages. Molecules 2022, 27, 3784. [Google Scholar] [CrossRef] [PubMed]
- Parunyakul, K.; Srisuksai, K.; Santativongchai, P.; Pongchairerk, U.; Ampawong, S.; Tulayakul, P.; Fungfuang, W. The first study on the effect of crocodile oil from Crocodylus siamensis on hepatic mitochondrial function for energy homeostasis in rats. Vet. world 2022, 15, 986–997. [Google Scholar] [CrossRef]
- Santativongchai, P.; Srisuksai, K.; Parunyakul, K.; Thiendedsakul, P.; Lertwatcharasarakul, P.; Fungfuang, W.; Tulayakul, P. Effects of crocodile oil (Crocodylus siamensis) on liver enzymes: Cytochrome P450 and glutathione s-transferase activities in high fat diet rats. Vet. Med. Int. 2022, 2022, 9990231. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Khan, F.; Garg, V.K.; Singh, A.K.; Kumar, T. Role of free radicals and certain antioxidants in the management of huntington’s disease: A review. J. Anal. Pharm. Res. 2018, 7, 386–392. [Google Scholar] [CrossRef]
- Rahman, M.; Islam, B.; Biswas, M.; Alam, K. In vitro antioxidant and free radical scavenging activity of different parts of Tabebuia pallida growing in Bangladesh. BMC Res. Notes. 2015, 8, 621. [Google Scholar] [CrossRef]
- Wu, Z.; Tan, B.; Liu, Y.; Dunn, J.; Guerola, P.M.; Tortajada, M.; Cao, Z.; Ji, P. Chemical composition and antioxidant properties of essential oils from peppermint, native spearmint and scotch spearmint. Molecules. 2019, 24, 2825. [Google Scholar] [CrossRef] [PubMed]
- Akmal, N.L.H.B.I.; Roy, A. Free radical scavenging activity of fish oil—An in-vitro study. Int. J. Pharm. Sci. Res. 2017, 8, 3872–3875. [Google Scholar]
- Nagababu, E.; Rifkind, J.M.; Sesikeran, B.; Lakshmaiah, N. Assessment of antioxidant activities of eugenol by in vitro and in vivo methods. Methods Mol. Biol. 2010, 610, 165–180. [Google Scholar]
- Gülçin, İ.; Elmastaş, M.; Aboul-Enein, H.Y. Antioxidant activity of clove oil—A powerful antioxidant source. Arab. J. Chem. 2012, 5, 489–499. [Google Scholar] [CrossRef]
- Mbah, C.J.; Orabueze, I.; Okorie, N.H. Antioxidants properties of natural and synthetic chemical compounds: Therapeutic effects on biological system. Acta Sci. Pharm. Sci. 2019, 3, 28–42. [Google Scholar] [CrossRef]
- Muriach, M.; Flores-Bellver, M.; Romero, F.J.; Barcia, J.M. Diabetes and the brain: Oxidative stress, inflammation, and autophagy. Oxid. Med. Cell Longev. 2014, 2014, 9. [Google Scholar] [CrossRef]
- Fai Poon, H.; Calabrese, V.; Scapagnini, G.; Butterfield, D.A. Free radicals and brain aging. Clin. Geriatr. Med. 2004, 20, 329–359. [Google Scholar] [CrossRef]
- Nantitanon, W.; Chowwanapoonpohn, S.; Okonogi, S. Antioxidant and antimicrobial activities of Hyptis suaveolens essential oil. Sci. Pharm. 2007, 75, 35–46. [Google Scholar] [CrossRef]
- Caleja, C.; Barros, L.; Antonio, A.L.; Oliveira, B.P.P.; Ferreira, I. A comparative study between natural and synthetic antioxidants: Evaluation of their performance after incorporation into biscuits. Food Chem. 2017, 216, 342–346. [Google Scholar] [CrossRef]
- Blasi, F.; Cossignani, L. An overview of natural extracts with antioxidant activity for the improvement of the oxidative stability and shelf life of edible oils. Processes 2020, 8, 956. [Google Scholar] [CrossRef]
- Santativongchai, P.; Fungfuang, W.; Boonyawiwat, V.; Pongchairerk, U.; Tulayakul, P. Comparison of physicochemical properties and fatty acid composition of crocodile oil (Crocodylus siamensis) extracted by using various extraction methods. Int. J. Food Prop. 2020, 23, 1465–1474. [Google Scholar] [CrossRef]
- Linn, T.W.; Kobroob, A.; Ngernjan, M.; Amornlerdpison, D.; Lailerd, N.; Wongmekiat, O. Crocodile oil disrupts mitochondrial homeostasis and exacerbates diabetic kidney injury in spontaneously diabetic Torii rats. Biomolecules 2022, 12, 1068. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Yen, P.; Chang, S.; Cheng, P.; Lo, Y.; Liao, V.H. Antioxidant activities of both oleic acid and Camellia tenuifolia seed oil are regulated by the transcription factor DAF-16/FOXO in Caenorhabditis elegans. PLoS ONE 2016, 11, e0157195. [Google Scholar]
- Emekli-Alturfan, E.; Kasikci, E.; Yarat, A. Effects of oleic acid on the tissue factor activity, blood lipids, antioxidant and oxidant parameter of streptozotocin induced diabetic rats fed a high-cholesterol diet. Med. Chem. Res. 2010, 19, 1011–1024. [Google Scholar] [CrossRef]
- Sakurai, K.; Shen, C.; Shiraishi, I.; Inamura, N.; Hisatsune, T. Consumption of oleic acid on the preservation of cognitive functions in Japanese elderly individuals. Nutrients 2021, 13, 284. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Chen, S.; Xu, X.; Zhu, M.; Zhao, W.; Liu, K.; Liu, K. Isolation of linoleic acid from Sambucus williamsii seed oil extracted by high pressure fluid and its antioxidant, antiglycemic, hypolipidemic activities. Int. J. Food Eng. 2015, 11, 383–391. [Google Scholar] [CrossRef]
- Yang, B.; Zhou, Y.; Wu, M.; Li, X.; Mai, K.; Ai, Q. ω-6 polyunsaturated fatty acids (linoleic acid) activate both autophagy and antioxidation in a synergistic feedback loop via TOR-dependent and TOR-independent signaling pathways. Cell Death Dis. 2020, 11, 607. [Google Scholar] [CrossRef]
- Hennebelle, M.; Morgan, R.K.; Sethi, S.; Zhang, Z.; Chen, H.; Grodzki, A.C.; Lein, P.J.; Taha, A.Y. Linoleic acid-derived metabolites constitute the majority of oxylipins in the rat pup brain and stimulate axonal growth in primary rat cortical neuron-glia co-cultures in a sex-dependent manner. J. Neurochem. 2020, 152, 195–207. [Google Scholar] [CrossRef]
- Li, R.; Chaicherdsakul, T.; Kunathigan, V.; Roytrakul, S.; Paemanee, A.; Kittisenachai, S. Shotgun proteomic analysis of germinated rice (Oryza sativa L.) under salt stress. Appl. Sci. Eng. Prog. 2020, 13, 76–85. [Google Scholar] [CrossRef]
- McLean, F.H.; Campbell, F.M.; Sergi, D.; Grant, C.; Morris, A.C.; Hay, E.A.; MacKenzie, A.; Mayer, C.D.; Langston, R.F.; Williams, L.M. Early and reversible changes to the hippocampal proteome in mice on a high-fat diet. Nutr. Metab. 2019, 16, 57. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Hossain, S.; Abdullah, N.; Aminudin, N. Brain proteomics links oxidative stress with metabolic and cellular stress response proteins in behavioural alteration of Alzhimer’s disease model rats. AIMS Neurosci. 2019, 6, 299–315. [Google Scholar] [CrossRef]
- Naphatthalung, J.; Cing, L.S.; Kanokwiroon, K.; Radenahmad, N.; Jansakul, C. Effect of six-week consumption of lard or palm oil on blood pressure and blood vessel H2S in middle-aged male rats. FFHD 2018, 8, 307–322. [Google Scholar] [CrossRef]
- Gong, H.; Han, Y.W.; Sun, L.; Zhang, Y.; Zhang, E.Y.; Li, Y.; Zhang, T.M. The effects of energy intake of four different feeding patterns in rats. Exp. Biol. Med. 2016, 241, 52–59. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Lacoste, M.G.; Ponce, I.T.; Golini, R.L.; Delgado, S.M.; Anzulovich, A.C. Aging modified daily variation of antioxidant enzymes and oxidative status in the hippocampus. Exp. Gerontol. 2017, 88, 42–50. [Google Scholar] [CrossRef]
- Ozcelebi, H.; Ari, F.; Dere, E. Glutathione S-transferase activity in tissues of rats exposed to fenarimol. Braz. Arch. Biol. Technol. 2021, 64, e21200751. [Google Scholar] [CrossRef]
- Mokhtari-Zaer, A.; Hosseini, M.; Boskabady, M.H. The effects of exercise on depressive- and anxiety-like behaviors as well as lung and hippocampus oxidative stress in ovalbumin-sensitized juvenile rats. Respir. Physiol. Neurobiol. 2018, 248, 55–62. [Google Scholar] [CrossRef]
- Losuwannarak, N.; Maiuthed, A.; Kitkumthorn, N.; Leelahavanichkul, A.; Roytrakul, S.; Chanvorachote, P. Gigantol targets cancer stem cells and destabilizes tumor via the suppression of the PI3K/AKT and JAK/STAT pathways in ectopic lung cancer xenografts. Cancers 2019, 11, 2032. [Google Scholar] [CrossRef] [PubMed]
- Johansson, C.; Samskog, J.; Sundström, L.; Wadensten, H.; Björkesten, L.; Flensburg, J. Differential expression analysis of Escherichia coli proteins using a novel software for relative quantitation of LC-MS/MS data. Proteomics 2006, 6, 4475–4485. [Google Scholar] [CrossRef] [PubMed]
- Thorsell, A.; Portelius, E.; Blennow, K.; Westman-Brinkmalm, A. Evaluation of sample fractionation using micro-scale liquid-phase isoelectric focusing on mass spectrometric identification and quantitation of proteins in a SILAC experiment. Rapid. Commun. Mass Sp. 2007, 21, 771–778. [Google Scholar] [CrossRef]
- Saravanan, N.; Haseeb, A.; Ehtesham, N.Z.; Ghafoorunissa. Differential effects of dietary saturated and trans-fatty acids on expression of genes associated with insulin sensitivity in rat adipose tissue. Eur. J. Endocrinol. 2005, 153, 159–165. [Google Scholar] [CrossRef]
- Yıldız, M.; Eroldoğan, T.O.; Ofori-Mensah, S.; Engin, K.; Ali Baltacı, M. The effects of fish oil replacement by vegetable oils on growth performance and fatty acid profile of rainbow trout: Re-feeding with fish oil finishing diet improved the fatty acid composition. Aquaculture 2018, 488, 123–133. [Google Scholar] [CrossRef]
- Duval, C.; Augé, N.; Frisach, M.; Casteilla, L.; Salvayre, R.; Nègre-Salvayre, A. Mitochondrial oxidative stress is modulated by oleic acid via epidermal growth factor receptor-dependent activation of glutathione peroxidase. Biochem. J. 2002, 367, 889–894. [Google Scholar] [CrossRef]
- Solfrizzi, V.; Capurso, C.; D’Introno, A.; Colacicco, A.M.; Frisardi, V.; Santamato, A.; Ranieri, M.; Fiore, P.; Vendemiale, G.; Seripa, D.; et al. Dietary fatty acids, age-related cognitive decline, and mild cognitive impairment. J. Nutr. Health Aging 2008, 12, 382–386. [Google Scholar] [CrossRef]
- Moon, J.; Shibamoto, T. Antioxidant assays for plant and food components. J. Agric. Food Chem. 2009, 57, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Bai, K.; Xu, W.; Zhang, J.; Kou, T.; Niu, Y.; Wan, X.; Zhang, L.; Wang, C.; Wang, T. Assessment of free radical scavenging activity of dimethylglycine sodium salt and its role in providing protection against lipopolysaccharide-induced oxidative stress in mice. PLoS ONE 2016, 11, e0155393. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.A.; Dek, M.S.P.; Tan, C.P.; Zainudin, M.A.M.; Fang, E.K.W. Changes of major antioxidant compounds and radical scavenging activity of palm oil and rice bran oil during deep-frying. Antioxidants 2014, 3, 205–215. [Google Scholar] [CrossRef]
- Ismail, M.; Alsalahi, A.; Imam, M.U.; Ooi, D.J.; Khaza’ai, H.; Aljaberi, M.A.; Shamsudin, M.N.; Idrus, Z. Safety and neuroprotective efficacy of palm oil and tocotrienol-rich fraction from palm oil: A systematic review. Nutrients 2020, 12, 521. [Google Scholar] [CrossRef] [PubMed]
- Praduptong, A.; Srimangkornkaew, P.; Chaeychomsri, S.; Thitipramote, N.; Chaeychomsri, W.; Siruntawineti, J. Antioxidant activity of mixture herbal oil from Siamese crocodile oil (Crocodylus siamensis), turmerica (Curcuma longa), black galingale (Kaempferia parviflora) and plai (Zingiber cassumunar Roxb). Prawarun Agr. J. 2020, 17, 159–170. [Google Scholar]
- Makaba, S.; Tingginehe, R.M.; Ruru, Y. The effectiveness of crocodile oil extract ointment on the treatment of burns in mice (Mus musculus). Med. Legal Update 2021, 21, 1114–1121. [Google Scholar]
- Snow, S.J.; Henriquez, A.R.; Fenton, J.I.; Goeden, T.; Fisher, A.; Vallanat, B.; Angrish, M.; Richards, J.E.; Schladweiler, M.C.; Cheng, W.; et al. Diets enriched with coconut, fish, or olive oil modify peripheral metabolic effects of ozone in rats. Toxicol. Appl. Pharmacol. 2021, 410, 115337. [Google Scholar] [CrossRef] [PubMed]
- Pellizzon, M.; Buison, A.; Ordiz, F.; Santa Ana, L.; Jen, C. Effects of dietary fatty acids and exercise on body-weight regulation and metabolism in rats. Obes. Res. 2002, 10, 947–955. [Google Scholar] [CrossRef]
- Rolland, V.; Roseau, S.; Fromentin, G.; Nicolaidis, S.; Tomé, D.; Even, P.C. Body weight, body composition, and energy metabolism in lean and obese Zucker rats fed soybean oil or butter. Am. J. Clin. Nutr. 2002, 75, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Witters, L.A.; Kemp, B.E. Insulin activation of acetyl-CoA carboxylase accompanied by inhibition of the 5’-AMP-activated protein kinase. J. Biol. Chem. 1992, 267, 2864–2867. [Google Scholar] [CrossRef]
- Bes-Rastrollo, M.; Sánchez-Villegas, A.; de la Fuente, C.; de Irala, J.; Martínez, J.A.; Marínez-González, M.A. Olive oil consumption and weight change: The SUN prospective cohort study. Lipids 2006, 41, 249–256. [Google Scholar] [CrossRef]
- García-Cerro, S.; Rueda, N.; Vidal, V.; Puente, A.; Campa, V.; Lantigua, S.; Narcís, O.; Velasco, A.; Bartesaghi, R.; Martínez-Cué, C. Prenatal administration of oleic acid or linolenic acid reduces neuromorphological and cognitive alterations in Ts65dn down syndrome mice. J. Nutr. 2020, 150, 1631–1643. [Google Scholar] [CrossRef]
- Raatz, S.K.; Conrad, Z.; Johnson, L.K.; Picklo, M.J.; Jahns, L. Relationship of the reported intakes of fat and fatty acids to body weight in US adults. Nutrients 2017, 9, 438. [Google Scholar] [CrossRef]
- Yang, J.; Chang, J.; Chen, C.; Yeh, C.; Chien, Y. Effects of different amounts and types of dietary fatty acids on the body weight, fat accumulation, and lipid metabolism in hamsters. Nutrition 2016, 32, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Allman-Farinelli, M.A.; Gomes, K.; Favaloro, E.J.; Petocz, P. A diet rich in high-oleic-acid sunflower oil favorably alters low-density lipoprotein cholesterol, triglyceride, and factor VII coagulant activity. J. Am. Diet Assoc. 2005, 105, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Bogensberger, B.; Benčič, A.; Knüppel, S.; Boeing, H.; Hoffmann, G. Effects of oils and solid fats on blood lipids: A systematic review and network meta-analysis. J. Lipid Res. 2018, 59, 1771–1782. [Google Scholar] [CrossRef]
- Qi, K.; Fan, C.; Jiang, J.; Zhu, H.; Jiao, H.; Meng, Q.; Deckelbaum, R.J. Omega-3 fatty acid containing diets decrease plasma triglyceride concentrations in mice by reducing endogenous triglyceride synthesis and enhancing the blood clearance of triglyceride-rich particles. Clin. Nutr. 2008, 27, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, M.F.; Afify Amer, M.M.; El-Saadany, S.S.; Abd El-Fatah El-Masry, R.; El-Said Awad, A. Changes in lipid profile by vegetable oil blends rich in polyunsaturated fatty acids in rats with hypercholesterolemia. Food Sci. Technol. Int. 2009, 15, 119–130. [Google Scholar] [CrossRef]
- Franssen, R.; Visser, M.E.; Kuivenhoven, J.A.; Kastelein, J.J.P.; Dallinga-Thie, G.M.; Stroes, E.S.G. Role of lipoprotein lipase in triglyceride metabolism: Potential therapeutic target. Future Lipodol. 2008, 3, 385–397. [Google Scholar]
- Nogoy, K.M.C.; Kim, H.J.; Lee, Y.; Zhang, Y.; Yu, J.; Lee, D.H.; Li, X.Z.; Smith, S.B.; Seong, H.A.; Choi, S.H. High dietary oleic acid in olive oil-supplemented diet enhanced omega-3 fatty acid in blood plasma of rats. Food Sci. Nutr. 2020, 8, 3617–3625. [Google Scholar] [CrossRef]
- Yıldırım, E.; Çınar, M.; Yalçınkaya, İ.; Ekici, H.; Atmaca, N.; Güncüm, E. Effect of cocoa butter and sunflower oil supplementation on performance, immunoglobulin, and antioxidant vitamin status of rats. Biomed. Res. Int. 2014, 2014, 606575. [Google Scholar] [CrossRef]
- Wichai, T.; Pannangrong, W.; Welbat, J.; Chaichun, A.; Sripanidkulchai, K.; Sripanidkulchai, B. Effects of aged garlic extract on spatial memory and oxidative damage in the brain of amyloid-β induced rats. Songklanakarin J. Sci. Technol. 2019, 41, 311–318. [Google Scholar]
- MacDonald-Wicks, L.; McEvoy, M.; Magennis, E.; Schofield, P.W.; Perrerson, A.J.; Zacharia, K. Dietary long-chain fatty acids and cognitive performance in older Australian adults. Nutrients 2019, 11, 711. [Google Scholar] [CrossRef]
- Khan, R.A. Effects of Launaea procumbens on brain antioxidant enzymes and cognitive performance of rat. BMC Complement Altern. Med. 2012, 12, 219. [Google Scholar] [CrossRef]
- Saleh, S.R.; Masry, A.M.; Ghareeb, D.A.; Newairy, A.A.; Sheta, E.; Maher, A.M. Trichoderma reesei fungal degradation boosted the potentiality of date pit extract in fighting scopolamine-induced neurotoxicity in male rats. Sci. Rep. 2021, 11, 14872. [Google Scholar] [CrossRef]
- Pitozzi, V.; Jacomelli, M.; Catelan, D.; Servili, M.; Taticchi, A.; Biggeri, A.; Dolara, P.; Giovannelli, L. Long-term dietary extra-virgin olive oil rich in polyphenols reverses age-related dysfunctions in motor coordination and contextual memory in mice: Role of oxidative stress. Rejuvenation Res. 2012, 15, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Pase, C.S.; Teixeira, A.M.; Roversi, K.; Dias, V.T.; Calabrese, F.; Molteni, R.; Franchi, S.; Panerai, A.E.; Riva, M.A.; Burger, M.E. Olive oil-enriched diet reduces brain oxidative damages and ameliorates neurotrophic factor gene expression in different life stages of rats. J. Nutr. Biochem. 2015, 26, 1200–1207. [Google Scholar] [CrossRef]
- Liu, J.; Su, G.; Gao, J.; Tian, Y.; Liu, X.; Zhang, Z. Effects of peroxiredoxin 2 in neurological disorder: A review of its molecular mechanisms. Neurochem. Res. 2020, 45, 720–730. [Google Scholar] [CrossRef]
- Choi, J.H.; Kim, D.W.; Yoo, D.Y.; Jeong, H.J.; Kim, W.; Jung, H.; Nam, S.M.; Kim, J.H.; Yoon, Y.S.; Choi, S.Y.; et al. Repeated administration of PEP-1-Cu,Zn-superoxide dismutase and PEP-1-peroxiredoxin-2 to senescent mice induced by D-galactose improves the hippocampal functions. Neurochem. Res. 2013, 38, 2046–2055. [Google Scholar] [CrossRef]
- Ock, J.; Han, H.S.; Hong, S.H.; Lee, S.Y.; Han, Y.; Kwon, B.; Suk, K. Obovatol attenuates microglia-mediated neuroinflammation by modulating redox regulation. Br. J. Pharmacol. 2010, 159, 1646–1662. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.; Sebaï, H.; Gadacha, W.; Boughattas, N.A.; Reinberg, A.; Mossadok, B.A. Catalase activity and rhythmic patterns in mouse brain, kidney and liver. Comp Biochem. Physiol. B Biochem. Mol. Biol. 2006, 145, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Yang, S. Brain globins in physiology and pathology. Med. Gas. Res. 2016, 6, 154–163. [Google Scholar]
- Brown, N.; Alkhayer, K.; Clements, R.; Singhal, N.; Gregory, R.; Azzam, S.; Li, S.; Freeman, E.; McDonough, J. Neuronal hemoglobin expression and its relevance to multiple sclerosis neuropathology. J. Mol. Neurosci. 2016, 59, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Richter, F.; Meurers, B.H.; Zhu, C.; Medvedeva, V.P.; Chesselet, M. Neurons express hemoglobin α- and β-chains in rat and human brains. J. Comp. Neurol. 2009, 515, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Nishi, H.; Inagi, R.; Kato, H.; Tanemoto, M.; Kojima, I.; Son, D.; Fujita, T.; Nangaku, M. Hemoglobin is expressed by mesangial cells and reduces oxidant stress. J. Am. Soc. Nephrol. 2008, 19, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Zou, R.; Tao, J.; Qiu, J.; Shi, W.; Zou, M.; Chen, W.; Li, W.; Zhou, N.; Wang, S.; Ma, L.; et al. Ndufs1 deficiency aggravates the mitochondrial membrane potential dysfunction in pressure overload-induced myocardial hypertrophy. Oxid. Med. Cell Longev. 2021, 2021, 5545261. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Fabuel, I.; Le Douce, J.; Logan, A.; James, A.M.; Bonvento, G.; Murphy, M.P.; Almeida, A.; Bolanos, J.P. Complex I assembly into supercomplexes determines differential mitochondrial ROS production in neurons and astrocytes. PNAS 2016, 113, 13063–13068. [Google Scholar] [CrossRef]
- Hirano, G.; Izumi, H.; Yasuniwa, Y.; Shimajiri, S.; Ke-Yong, W.; Sasagiri, Y.; Kusaba, H.; Matsumoto, K.; Hasegawa, T.; Akimoto, M.; et al. Involvement of riboflavin kinase expression in cellular sensitivity against cisplatin. Int. J. Oncol. 2011, 38, 893–902. [Google Scholar]
- Chidawanyika, T.; Supattapone, S. Hydrogen peroxide-induced cell death in mammalian cells. J. Cell Signal. 2021, 2, 206–211. [Google Scholar]
- Yadav, M.; Rai, N.; Yadav, H.S. The role of peroxidase in the enzymatic oxidation of phenolic compounds to quinones from Luffa aegyptiaca (gourd) fruit juice. Green Chem. Lett. Rev. 2017, 10, 154–161. [Google Scholar] [CrossRef]
- Khan, A.A.; Rahmani, A.H.; Aldebasi, Y.H.; Aly, S.M. Biochemical and pathological studies on peroxidases-an updated review. Glob. J. Health. Sci. 2014, 6, 87–98. [Google Scholar] [CrossRef]
- Singhal, A.; Morris, V.B.; Labhasetwar, V.; Ghorpade, A. Nanoparticle-mediated catalase delivery protects human neurons from oxidative stress. Cell Death Dis. 2013, 4, e903. [Google Scholar] [CrossRef]
- Bahrami, S.; Shahriari, A.; Tavalla, M.; Azadmanesh, S.; Hamidinejat, H. Blood levels of oxidant/antioxidant parameters in rats infected with Toxoplasma gondii. Oxid. Med. Cell Longev. 2016, 2016, 8045969. [Google Scholar] [CrossRef]
- Kumagai, A.; Fujita, A.; Yokoyama, T.; Nonobe, Y.; Hasaba, Y.; Sasaki, T.; Itoh, Y.; Koura, M.; Suzuki, O.; Adachi, S.; et al. Altered Actions of memantine and NMDA-induced currents in a new Grid2-deleted mouse line. Genes 2014, 5, 1095–1114. [Google Scholar] [CrossRef] [PubMed]
- Bye, C.M.; McDonald, R.J. A specific role of hippocampal NMDA receptors and Arc protein in rapid encoding of novel environmental representations and a more general long-term consolidation function. Front. Behav. Neurosci. 2019, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Collingridge, G.L.; Bliss, T.V.P. NMDA receptors-their role in long-term potentiation. Trends Neurosci. 1987, 10, 288–293. [Google Scholar] [CrossRef]
- Keller, E.A.; Borghese, C.M.; Carrer, H.F.; Ramirez, O.A. The learning capacity of high or low performance rats is related to the hippocampus NMDA receptors. Brain Res. 1992, 576, 162–164. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Liang, M.; Yu, C.; Wei, Z. Liprin-α-mediated assemblies and their roles in synapse formation. Front. Cell Dev. Biol. 2021, 9, 653381. [Google Scholar] [CrossRef]
- Mao, X.W.; Sandberg, L.B.; Gridley, D.S.; Herrmann, E.C.; Zhang, G.; Raghavan, R.; Zubarev, R.A.; Zhang, B.; Stodieck, L.S.; Ferguson, V.L.; et al. Proteomic analysis of mouse brain subjected to spaceflight. Int. J. Mol. Sci. 2019, 20, 7. [Google Scholar] [CrossRef]
- Wong, M.Y.; Liu, C.; Wang, S.S.H.; Roquas, A.C.F.; Fowler, S.C.; Kaeser, P.S. Liprin-α3 controls vesicle docking and exocytosis at the active zone of hippocampal synapses. PNAS 2018, 115, 2234–2239. [Google Scholar] [CrossRef]
- Ishimoto, T.; Ninomiya, K.; Inoue, R.; Koike, M.; Uchiyama, Y.; Mori, H. Mice lacking BCAS1, a novel myelin-associated protein, display hypomyelination, schizophrenia-like abnormal behaviors, and upregulation of inflammatory genes in the brain. Glia 2017, 65, 727–739. [Google Scholar] [CrossRef]
- Fields, R.D.; Bukalo, O. Myelin makes memories. Nat. Neurosci. 2020, 23, 469–470. [Google Scholar] [CrossRef]
- Hosseinzadeh, Z.; Moazedi, A.A.; Chinipardez, R. The effect of palmitic acid on spatial learning and extinction in adult male rat. Pak. J. Biol. Sci. 2007, 10, 2653–2658. [Google Scholar]
- Melo, H.M.; Silva, G.S.; Sant’Ana, M.R.; Teixeira, C.V.L.; Clarke, J.R.; Coreixas, V.M.; de Melo, B.C.; Fortuna, J.T.S.; Forny-Germano, L.; Henrique Ledo, J.; et al. Palmitate is increased in the cerebrospinal fluid of humans with obesity and induces memory impairment in mice via pro-inflammatory TNF-α. Cell Rep. 2020, 30, 2180–2194. [Google Scholar] [CrossRef] [PubMed]
- Ugbaja, R.N.; James, A.S.; Ugwor, E.I.; Akamo, A.J.; Thomas, F.C.; Kosoko, A.M. Lycopene suppresses palmitic acid-induced brain oxidative stress, hyperactivity of some neuro-signalling enzymes, and inflammation in female Wistar rat. Sci. Rep. 2021, 11, 15038. [Google Scholar] [CrossRef] [PubMed]
- Franzoni, F.; Scarfo, G.; Guidotti, S.; Fusi, J.; Asomov, M.; Pruneti, C. Oxidative stress and cognitive decline: The neuroprotective role of natural antioxidants. Front. Neurosci. 2021, 15, 729757. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Composition (%) |
|---|---|
| Lauric acid (C12:0) | 0.11 ± 0.036 |
| Myristic acid (C14:0) | 0.57 ± 0.060 |
| Pentadecanoic acid (C15:0) | 0.09 ± 0.010 |
| Palmitic acid (C16:0) | 19.92 ± 0.307 |
| Heptadecanoic acid (C17:0) | 0.16 ± 0.010 |
| Stearic acid (C18:0) | 5.42 ± 0.029 |
| Arachidic acid (C20:0) | 0.13 ± 0.012 |
| Myristoleic acid (C14:1) | 0.10 ± 0.006 |
| Palmitoleic acid (C16:1) | 3.83 ± 0.137 |
| cis-10-Heptadecenoic acid (C17:1n10) | 0.11 ± 0.010 |
| cis-9-Oleic acid (C18:1n9) | 41.07 ± 0.549 |
| Linoleic acid (C18:2 n6c) | 21.08 ± 0.180 |
| alpha-Linoleic acid (ALA, C18:3n3) | 0.96 ± 0.049 |
| gamma-Linoleic acid (C18:3n6) | 0.18 ± 0.006 |
| cis-11-Eicosenoic acid (C20:1n9) | 0.41 ± 0.026 |
| cis-11,14-Eicosadienoic acid (C20:2) | 0.27 ± 0.035 |
| cis-8,11,14-Eicosatrienoic acid (C20:3n6) | 0.27 ± 0.035 |
| Arachidonic acid (C20:4n6) | 0.82 ± 0.092 |
| Docosahexaenoic acid (DHA, C22:6n3) | 0.22 ± 0.015 |
| Others Total saturated fatty acids Total unsaturated fatty acids | 4.27 ± 0.234 26.40 ± 0.40 69.33 ± 0.642 |
| Parameter | Group | ||
|---|---|---|---|
| NS | NC1 | NC3 | |
| Food intake (g/rat/day) | 19.00 ± 0.02 | 18.49 ± 0.21 ** | 16.25 ± 0.06 ** |
| Energy intake (kcal/rat/day) | 63.34 ± 0.73 | 65.94 ± 1.36 | 66.13 ± 0.92 |
| Body weight gain (g) | 35.50 ± 2.81 | 41.14 ± 5.54 | 49.67 ± 12.20 |
| Parameter | Group | ||
|---|---|---|---|
| NS | NC1 | NC3 | |
| Cholesterol (mg/dL) | 76.13 ± 2.28 | 73.58 ± 7.63 | 69.43 ± 5.92 |
| Triglycerides (mg/dL) | 189.10 ± 33.86 | 100.58 ± 11.13 * | 98.42 ± 6.87 ** |
| HDL-C (mg/dL) | 39.05 ± 2.70 | 42.74 ± 6.40 | 38.02 ± 3.86 |
| LDL-C (mg/dL) | 10.83 ± 1.98 | 13.02 ± 0.81 | 12.70 ± 2.73 |
| Group | Accession Number | Gene Name | Protein Name | Function | Peptide |
|---|---|---|---|---|---|
| NC1 | A0A0G2JSH9 | Prdx2 | Peroxiredoxin-2 | Antioxidant activity; Thioredoxin-peroxidase activity | QITVNDLPVGR |
| NC3 | P04762 | Cat Cas1 | Catalase | Antioxidant activity; Catalase activity; Oxidoreductase activity | GKANL |
| P01946 | Hba1 Hba-a1 | Hemoglobin subunit alpha-1/2 | Oxygen binding; Oxygen-carrier activity | LRVDPVNFK | |
| Shared in NC1–NC3 group | Q66HF1 | Ndufs1 | NADH-ubiquinone oxidoreductase 75-kDa subunit | Electron-transfer activity; NADH-dehydrogenase activity | KPMVVLGSSALQR |
| Q6AYA7 | Rfk | Riboflavin kinase | ATP binding; Riboflavin-kinase activity; | GFGRGSK | |
| A0A0G2JWB6 | Pxdn | Peroxidase | Peroxidase activity | EIQPGAFR |
| Accession Number | Gene Name | Protein Name | Function | Peptide |
|---|---|---|---|---|
| Q63226 | Grid2 | Glutamate receptor ionotropic, delta-2 | Glutamate-receptor activity; Ionotropic glutamate-receptor activity; Ligand-gated ion-channel activity | LENNMR |
| Q91Z79 | Ppfia3 | Liprin-alpha-3 | Neurotransmitter secretion; Regulation of short-term neuronal; synaptic plasticity; Exocytosis of synaptic vesicles | MNDDHNK |
| A0A0G2K079 | Bcas1 | Breast carcinoma-amplified sequence 1 homolog | Myelination | DPEDTK |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srisuksai, K.; Parunyakul, K.; Santativongchai, P.; Phaonakrop, N.; Roytrakul, S.; Tulayakul, P.; Fungfuang, W. Antioxidant Activity of Crocodile Oil (Crocodylus siamensis) on Cognitive Function in Rats. Foods 2023, 12, 791. https://doi.org/10.3390/foods12040791
Srisuksai K, Parunyakul K, Santativongchai P, Phaonakrop N, Roytrakul S, Tulayakul P, Fungfuang W. Antioxidant Activity of Crocodile Oil (Crocodylus siamensis) on Cognitive Function in Rats. Foods. 2023; 12(4):791. https://doi.org/10.3390/foods12040791
Chicago/Turabian StyleSrisuksai, Krittika, Kongphop Parunyakul, Pitchaya Santativongchai, Narumon Phaonakrop, Sittiruk Roytrakul, Phitsanu Tulayakul, and Wirasak Fungfuang. 2023. "Antioxidant Activity of Crocodile Oil (Crocodylus siamensis) on Cognitive Function in Rats" Foods 12, no. 4: 791. https://doi.org/10.3390/foods12040791
APA StyleSrisuksai, K., Parunyakul, K., Santativongchai, P., Phaonakrop, N., Roytrakul, S., Tulayakul, P., & Fungfuang, W. (2023). Antioxidant Activity of Crocodile Oil (Crocodylus siamensis) on Cognitive Function in Rats. Foods, 12(4), 791. https://doi.org/10.3390/foods12040791