
Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective


Abstract
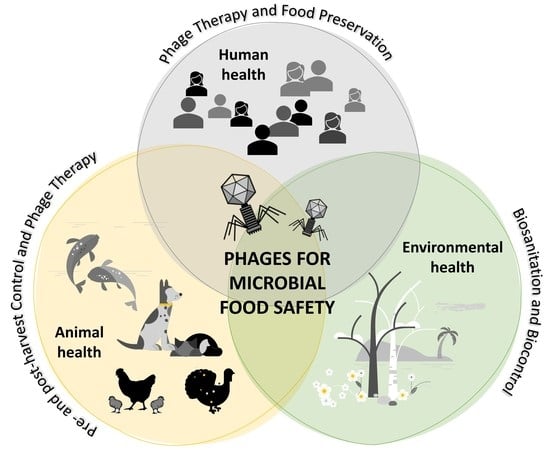
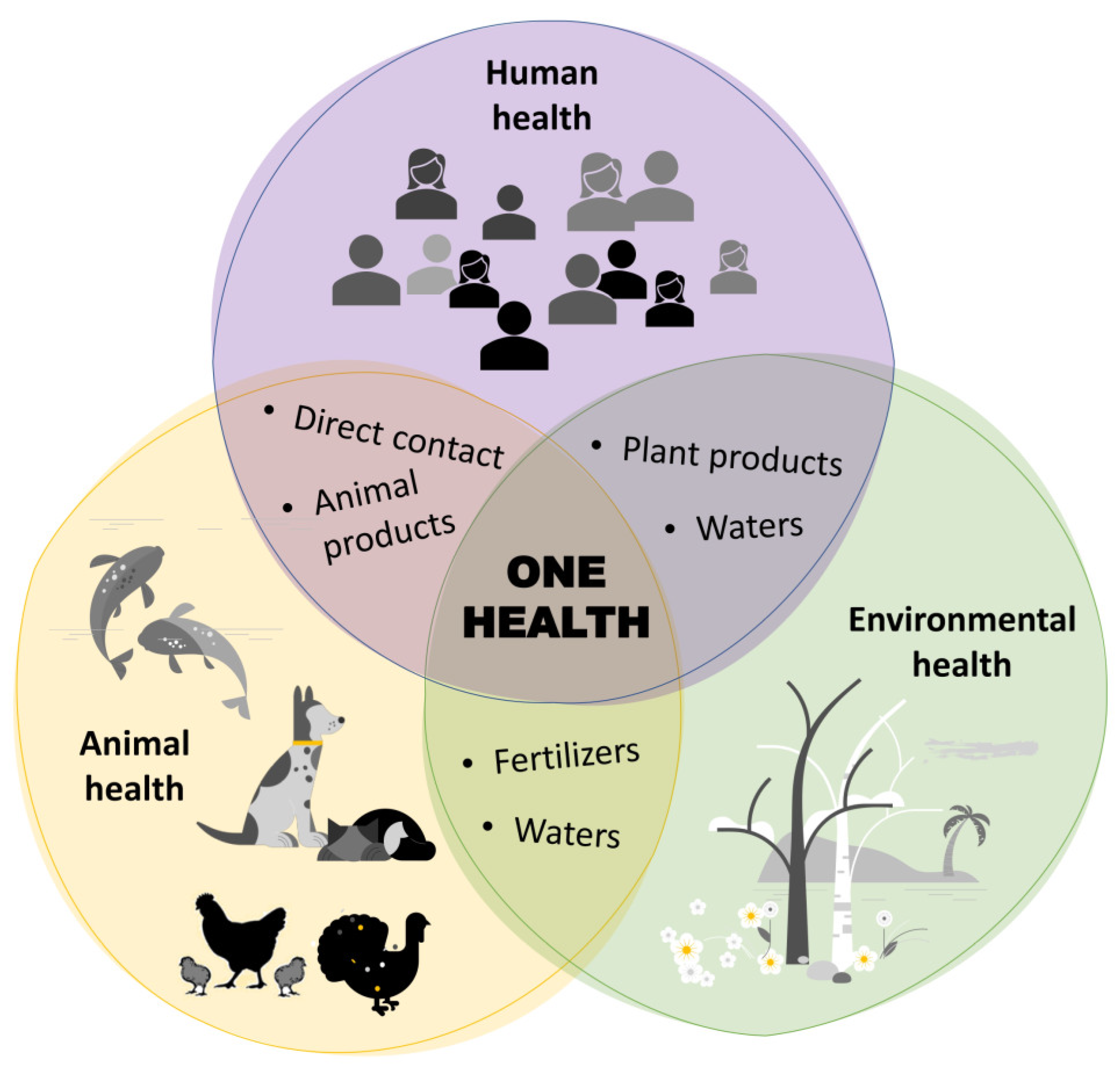
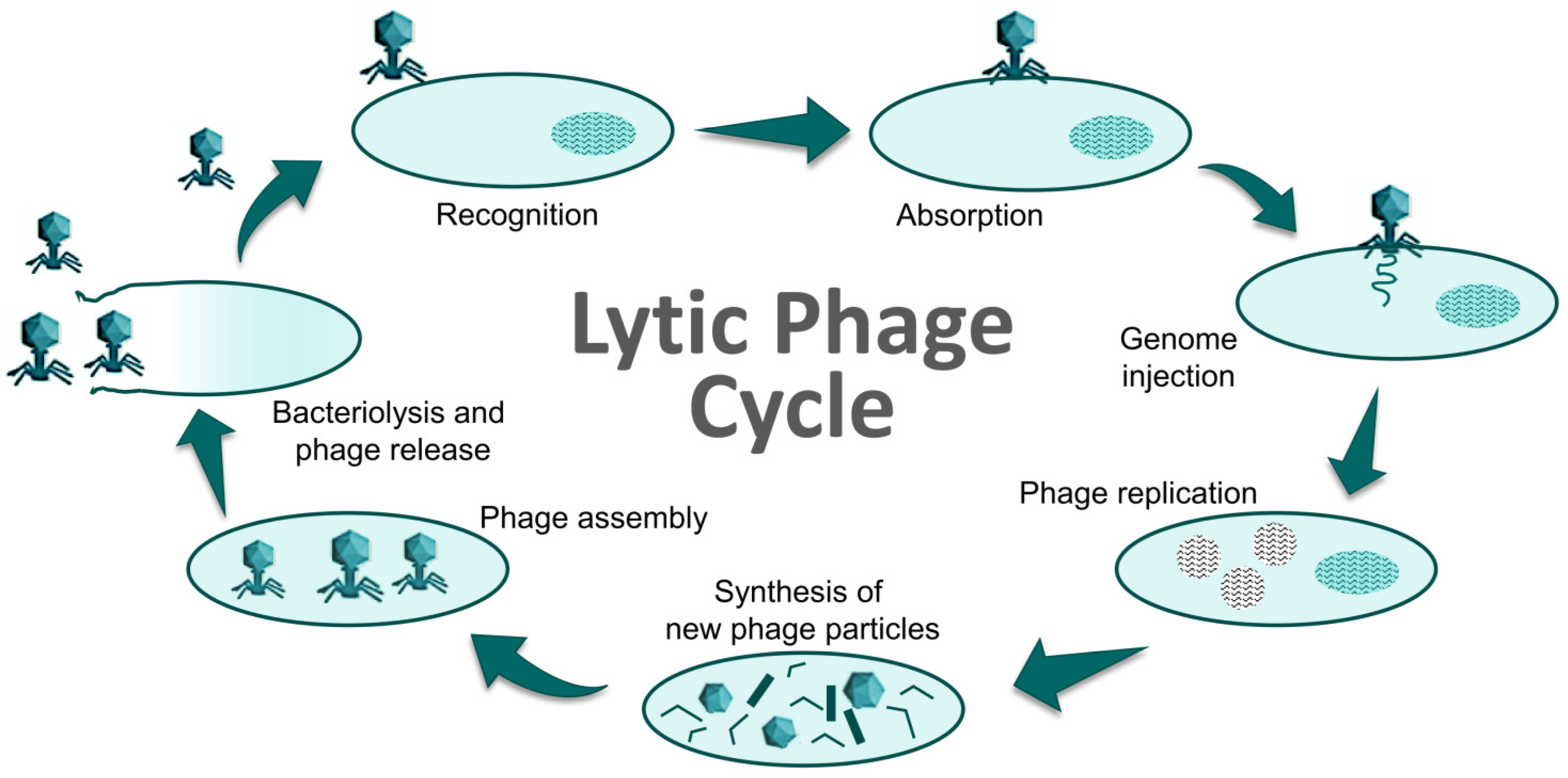
1. Introduction
2. Phage Biocontrol in Animal Husbandry for Food Production
2.1. Control of Campylobacter and Salmonella in Broilers

{kind=link}
{kind=link}
{kind=link}
| Animal (Age) | Bacteria Load 1,2 | Phage | Application Method and Dose 3 | Bacterial Reduction | Ref. |
|---|---|---|---|---|---|
| Campylobacter spp. | |||||
| Chickens (38 days old) | C. jejuni AMR * 108 (1) | ϕ16-izsam ϕ7-izsam | Oral (37 dpi); single dose; sequential application (24 h) 1 day before slaughtering. 107 108 | 1 log reduction 2 log reduction | [38] |
| Chickens (24 days old) | C. jejuni HPC5 108 (1) | Cocktail (2): CP20 GII CP30A GIII | Oral (4 dpi); single dose; 107 | 2.4 and 1.3 log reduction after 2 and 5 dpt | [39] |
| Chickens (25 days old) | C. jejuni HPC5 107 (1) C. jejuni GIIC8 107 (1) | CP34 GIII CP8 GIII CP8 GIII | Oral (5 dpi); single dose; 105–107–109 | 0.5–4 log reduction Marginal reductions Initial 5 log reduction and 2 log reduction after 5 dpt | [40] |
| Chickens (38 days old) | C. jejuni 2140CD1 107 (1) C. coli A11 106 (1) | Cocktail (3): ϕCcoIBB35 GII ϕCcoIBB37 GII ϕCcoIBB12 GII | Oral (7 dpi); single dose; 106 In feed (7dpi); single dose; 107 | 1.2 and 1.7 log reduction after 2 and 7 dpt 2 log reduction after 2 and 7 dpt | [41] |
| 3 field trials Chickens (36 days old) | C. jejuni 102–107 (1) | Cocktail (4): NCTC12672 GIII NCTC12673 GIII NCTC12674 GIII NCTC12678 GIII | Drinking water (7 dpi); single dose; 107 | Up to 3.2 log reduction in one field trial No reduction in two field trials | [42] |
| Chickens (47 days old) | Naturally colonized chickens | Cocktail (4): PH5, PH8, PH11, PH13 | Oral; single dose; 107 | 1.3 log reduction after 1 dpt | [43] |
| Chickens (10 days old) Chickens (32 days old) | C. jejuni C356 108–109 (1) | NCTC12671 GIII Cocktail (2): NCTC12671 GIII NCTC12669 GIII | Oral (5dpi); 5 doses (24 h interval); 1010–1011 Oral (7dpi); 4 doses (24 h interval); 1010–1011 | Initial 3 log reduction followed by 1 log reduction over 20 dpt Initial 1.5 log reduction followed by 1 log reduction over 20 dpt | [44] |
| Chickens (25 days old) | C. jejuni HPC5 107 (1) C. coli OR12 109 (1) | CP220 GII | Oral (5 dpi); single dose; 107 109 | 2 log reduction 2 log reduction | [45] |
| Chickens (27 days old) | C. jejuni 3871 109 (1) | CP14 GIII Cocktail (2): CP14 GIII CP81 GIII CP14 GIII CP68 GII | Oral (7 dpi); single dose; 5 × 108 Oral (7 dpi); single dose; 5 × 108 Oral (7 dpi); single dose; sequential application (24 h); 5 × 108–5 × 1010 | 1 log reduction after 3 dpt No reduction 3 log reduction after 3 dpt | [46] |
| Salmonella spp. | |||||
| Layer hens (6 weeks old) | S. Gallinarum KVCC BA00722 108 (2) | ST4 L13 SG3 | Feed additive 108 7 days before and 21 dpi | 50% reduction in liver and spleen after 7 dpi; 70% survival rate 75% and 50% reduction in liver and spleen after 7 and 14 dpi, respectively; 75% survival rate 25% and 50% reduction in liver and spleen after 7 and 14 dpi, respectively; 50% survival rate | [49] |
| Chickens (36 days old) | S. Enteritidis P125109 S. Typhimurium 4/74 108 (2) | Phage cocktail: ϕ151, ϕ25 ϕ10 | Oral; single dose 1011 | 1.53 log and 3.48 log reduction of S. Enteritidis and S. Typhimurium, respectively | [50] |
| Chickens (6-10 days) Chickens (31-35 days) | S. Enteritidis (PT4) 6 × 106 (2) | Phage cocktail: CNPSA1 CNPSA3 CNPSA4 | Early treatment after challenge: drinking water for 5 consecutive days (from 6 to 10 dpi) Later treatment after challenge: drinking water for 5 consecutive days (from 31 to 35 dpi) 109 | 1.08 log reduction after later treatment | [51] |
| Chickens (1 day old) (6 days old) | S. Enteritidis 103 (2) | Single phage or cocktail: CB4ϕ WT45ϕ | Cloacal drop 1 h pi: WT45ϕ: 109 Oral delivery 1 h pi: WT45ϕ: 108 CB4ϕ: 108 Cocktail: 108 | Reduction in Salmonella detection to 36% Reduction in Salmonella detection to 70%, 65%, and 45% after 1 dpt No significant differences after 48 h | [52] |
| Chickens (10 days old) | S. Enteritidis 105 (2) | Phage cocktail | Coarse spray or drinking water 108 | Reduction in Salmonella detection to 72.7% by aerosol-spray | [53] |
| Layer hens (40 weeks old) | S. Enteritidis (SENAR) 108 (2) | Phage cocktail: SP-1 STP-1 | Feed additive: 0.2% of the phage cocktail | 0.9, 0.57, and 0.38 log reduction in cecum, liver, and spleen at 7 dpt 0.86 log reduction in cecum at 6 dpt | [54] |
| Layer hens (60 weeks old) | Natural infection | Autophage (AP) Wild-type phage | Spray applications 108 Two single applications in 24 h intervals | 1.78 log reduction in feces samples Total elimination of Salmonella from the environment | [55] |
| Chickens (1 to 35 days) | S. Enteritidis 104 (2) | Bafasal (4 phages cocktail) | Feed additive daily 106 | 1 log reduction at day 35 of study | [56] |
2.2. Control of Listeria Monocytogenes in Animals
2.3. Control of Vibrio spp. in Aquaculture
3. Phage Biocontrol at the Post-Harvest and Post-Slaughtering Stage
3.1. Campylobacter
| Food | Bacteria Load 1 | Phage | Application MOI * and Method | Result/Bacterial Reduction | Ref. |
|---|---|---|---|---|---|
| Campylobacter spp. | |||||
| Chicken skin | C. jejuni PT14 4 and 6 log | ϕ2: NCTC12674 GIII | MOI: 0.01–1 MOI: 10–103 spread on surface | Negligible reduction 1 log reduction after 30 min, 3 d and 5 d at 4 °C 2.5 log reduction after 5 d at −20 °C | [26] |
| Chicken skin | C. jejuni C222 4 log | NCTC12673 GIII | MOI: 102 spread on surface | 1 log reduction after 1 d at 4 °C | [94] |
| Chicken neck skin | C. jejuni NCTC12662 4 log | F356GIII F357 GIII F379 GIII Cocktail (2): F356 GIII F357 GIII | MOI: 103 spread on surface | 0.5 log reduction at 5 °C 0.5 log reduction at 5 °C Negligible reduction at 5 °C 0.7 log reduction after 1 d at 5 °C | [95] |
| Chicken meat | C. coli NCTC 126683 C. jejuni NCTC 11168 3 log | NCTC12684 GII CP81 GIII | MOI: 104 spread on surface | No reduction at 4 °C No reduction at 4 °C | [96] |
| Raw and cooked beef | C. jejuni 4 log | Cj6 | MOI: 104 spread on surface | 1.5 and 2 log reduction after 1 d at 5 °C in raw and cooked beef, respectively | [97] |
| Chicken meat | C. jejuni 4 log | CJ01 | MOI: 102 spread on surface | 1.7 log reduction after 2 d at 4 °C | [98] |
| Salmonella spp. | |||||
| Commercial broiler and turkey carcasses | S. Enteritidis (PT 13A) 20 CFU S. Enteritidis (PT 13A) 31 CFU S. Enteritidis host S. Enteritidis field (S9, S14) | PHL 4 72 wild-type phages | Broiler carcasses: MOI: 104 to 1010 spray Turkey carcasses: MOI: 106 to 108 rinsed | 50–100% reduction 60% reduction | [100] |
| Breast and eggs | S. Enteritidis LK5, UA1894 Breast: 106 Eggs: 107 | UAB_Phi 20 UAB_Phi78 UAB_Phi87 | 109 PFU (MOI: 103) rinse 1010 (MOI:103) spray | 2.0 log reduction 0.9 log reduction | [101] |
| Liquid eggs and chicken meat | S. Enteritidis Liquid eggs: 104 Chicken meat: 105 | SE07 | 1011 (MOI 107) Direct addition of 100 mL 1012 (MOI 107) spray | 2 log reduction after 12, 24, and 48 h 2 log reduction after 12, 24, and 48 h | [102] |
| Breast samples | S. Enteritidis ATCC13076 CVCC2184 4 × 105 | PA13076 PC2184 | Single phage: 4 × 109 (MOI: 104) Cocktail: 4 × 109 (MOI: 104) | 2 log reduction Phage PC2184 better than phage PA13076 at 4 °C and 25 °C 2 log reduction | [103] |
| Chicken breast | S. Typhimurium ATCC 14,028 S. Enteritidis ATCC 4931 S. Heidelberg ATCC 8326 3 logs | SalmoFresh TM (6 phages) | MOI: 106 spray | 0.7 and 0.9 log reduction on day 0 and 1, at 4 °C 1 log reduction on day 7 with modified atmosphere at 4 °C 0.8, 0.9, and 0.4 log reduction at 0, 4, and 8 h at room temperature, respectively | [104] |
| Chicken and turkey meat | S. Enteritidis ATCC 13,076 S. Typhimurium ATCC 6539 S. Heidelberg ATCC 8326 1.5 × 103 1.25 × 103 | SalmoLyse® | 2 × 106, 4 × 106, 9 × 106 MOI: 2 × 103, 3 × 103, 6 × 103 spray 9 × 106 and 2 × 107 MOI: 7 × 103, 1 × 104 spray | 60%, 71%, and 88% reduction from chicken meat at 2 × 106, 4 × 106, 9 × 106 PFU/mL, respectively 68% and 86% reduction from turkey meat at 106 and 107 PFU/g, respectively | [105] |
| Chicken meat | S. Typhimurium JCW-3001 S. Enteritidis VDL-133 S. Dublin SP-598 5 log | SalmoFREE® (6 phages) | 108, 109 (MOI: 103, 104) immersion | 1.9–2.0 log reduction in combination with plant-based essential oils | [106] |
| Chicken meat | S. Enteritidis 104 | PhageGuard S® (2 phages) | 107 (MOI: 103) immersion | 1.5 log reduction after 24 h | [107] |
| Listeria monocytogenes | |||||
| Raw salmon | 4 log 2 log | Listex™ P100 | MOI: 1, 10, 102, 103, 104 spread on surface MOI: 106 spread on surface | Marginal reductions at lower MOIs, but up to 3 log reduction at higher MOIs 1.4 log reduction (1 d) No regrowth after 10 d at 4 °C | [108] |
| Raw hake Raw salmon Smoked salmon | 3 log | Listex™ P100 | Automated spray MOI: 104 | 1.2 and 2.0 log reduction after 1 d and 7 d at 4°C (hake) 0.8 and 1.0 log reduction after 1 d and 7 d at 4°C (raw salmon) 0.8 and 1.6 log reduction after 1 d and 30 d at 4°C (smoked salmon) | [109] |
| Smoked salmon | 3 log | ListShield™ (6 phages) | MOI: 103 spray | 0.4 and 1 log reduction | [110] |
| RTE chicken breast roll | 2, 4, and 5 log | FWLLm1 | MOI: 105, 103, 102 spread on surface | Rapid 1.5–2.5 log at 5–30 °C. Regrowth prevented over 21 d at higher MOI and 5 °C (vacuum packed) | [111] |
| Cooked turkey and roast beef | 3 log | Listex™ P100 | MOI:104 spread on surface | 1.7 log and 2.1 log, respectively, after 28 d at 4 °C | [112] |
| Sliced cooked ham | 4 log | Listex™ P100 | MOI: 104 spread on surface | Rapid 1 log reduction 2 log reduction after 28 d at 4 °C | [113] |
| Dry-cured ham | 2, 3, 4 log | Listex™ P100 | MOI: 102–106 spread on surface | 2.5 log to undetectable (highest MOI) after 14 d at 4 °C | [114] |
| Milk | 5 log | Monophages LMP1 and LMP7 | MOI:10 addition to milk | 0.5–3.3 log at 4 °C | [115] |
| “Queso fresco” cheese | 4 log | Listex™ P100 | MOI: 104 spread on surface | 2 log reduction | [116] |
| Soft cheeses | 3 log 1, 2 log | A511 | MOI: 105 in the smearing solution MOI: 106, 107 | 2.5–3 log reduction during the 21 d ripening period >6 log reduction (below the limit of detection) | [117] |
| Hard cheese | 4 log | ListShield™ (6 phages) | MOI: 104 spray | 0.7 log reduction | [110] |
| Lettuce Apple slices | 3 log 4 log | ListShield™ (6 phages) | MOI: 104, 105 spray MOI: 102 spray | 1.1 log reduction at higher MOI 1 log reduction | [110] |
| Fresh-cut apple and melon | 5.5 log | Cocktail (12 phages) LM-103 Cocktail (6 phages) LMP-102 | MOI: 102 spray | Below 0.4 log reduction in apple 2.0–4.6 log reduction in melon | [118] |
| Fresh-cut apple, pear, and melon slices. Apple, pear, and melon juices | 5 log 5 log | Listex™ P100 | MOI: 103 spread on surface MOI: 103 addition to juice | None, 0.6, and 1.5 log reduction in apple, pear, and melon slices after 8 d at 10 °C None, 2, and 8 log reduction in apple, pear, and melon juices after 8 d at 10 °C | [119] |
| Celery and enoki mushroom | 5 log | Mix of 3 phages: LMPC01 LMPC02 LMPC03 | MOI: 10 | 2.2 and 1.8 log reduction in celery and enoki mushroom after 7 d at 4 °C | [120] |
| Vibriospp. | |||||
| Oysters Crassostrea gigas | V. parahaemolyticus CRS 09-17, AMR* 1.6 × 106 CFU in each oyster | pVp-1 | 2 × 107 PFU/oyster (MOI: 10) surface of flesh | 6 log CFU/mL growth reduction after 12 h | [80] |
| Fresh fish flesh | V. parahaemolyticus FORC_023 3 × 104 | VPG01 | MOI: 1 MOI: 10 surface direct application | 1 log reduction (MOI: 1) Counts under the detection limit after 6 h (MOI: 10) | [81] |
| Cutting board | V. parahaemolyticus FORC_023 3 × 104 CFU/cm2 | VPG01 | MOI: 103 surface direct application | 3 log reduction in utensil surface | [81] |
| Raw fish flesh slices | V. parahaemolyticus FORC_023 3 × 104 | VPT02 | MOI of 0, 1, or 10 surface direct application | 2 log reduction after 6 h at 25 °C (MOI: 10) | [121] |
| Shrimp | V. parahaemolyticus F23 | F23s1 Recombinant endolysin ORF52 | MOI: 103 in vitro 20 µmol/L | Growth inhibition at 25 °C for 12 h Decreased OD600 after 60 min The endolysin also showed lytic activity against a panel of 23 drug-resistant V. parahaemolyticus | [122] |
| Manila clams | V. parahaemolyticus Vp-KF4 1 × 104 | Vpp2 | MOI of 1, 10, or 100 | 2.1 log reduction at 25 °C until 24 h No effect of treatment at 4 °C | [123] |
| Oysters | V. parahaemolyticus ATCC 17802) 104 | vB_VpaS_OMN | MOI: 103 surface direct application | 1 log and 2 log reduction after 48 and 72 h of incubation, respectively | [124] |
| Oysters | V. vulnificus 106 | Phage pool (9 phages): S1, P3, P38, P53, P65, P68, P108, P111, P147 | Unknown | 5 log reduction after 18 h of incubation at 4 °C | [125] |
| Abalone flesh | V. vulnificus MO6-24/O 2 × 103 | VVP001 | MOI: 105 MOI: 106 | 2.06 log reduction 2.51 log reduction | [126] |
3.2. Salmonella
3.3. Listeria Monocytogenes
3.4. Human Pathogenic Vibrio spp.
4. Challenges of Using Phages for Food Safety
| Challenge | Causes and/or Future Studies Needs | Refs. |
|---|---|---|
| Extreme host specificity |
| [144,145] |
| Potential development of phage resistance |
| [145,146,147] |
| Phage stability and administration routes |
| [148,149,150,151] |
| Mobilization of resistant genes between bacteria |
| [152,153] |
| Phage biobanking for immediate trials |
| [154] |
| Legislative approval |
| [155,156] |
| Consumer acceptance |
| [89] |
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Endersen, L.; O’Mahony, J.; Hill, C.; Ross, R.P.; McAuliffe, O.; Coffey, A. Phage Therapy in the Food Industry. Ann. Rev. Food Sci. Technol. 2014, 5, 327–349. [Google Scholar] [CrossRef]
- Mousavi, S.L.; Rasooli, I.; Nazarian, S.; Amani, J. Simultaneous detection of Escherichia coli O157:H7, toxigenic Vibrio cholerae, and Salmonella typhimurium by multiplex PCR. Iran. J. Clin. Infect. Dis. 2009, 4, 97–103. [Google Scholar]
- Gould, L.H.; Seys, S.; Everstine, K.; Norton, D.; Ripley, D.; Reimann, D.; Dreyfuss, M.; Chen, W.S.; Selman, C.A. Recordkeeping Practices of Beef Grinding Activities at Retail Establishments. J. Food Prot. 2011, 74, 1022–1024. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Tauxe, R.; Hednerg, C. The growing burden of foodborne outbreaks due to contaminated fresh produce: Risks and opportunities. Epidemiol. Infect. 2009, 137, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.L.; Lee, H.Y.; Mahyudin, N.A. Antimicrobial resistance of Escherichia coli and Staphylococcus aureus isolated from food handler’s hands. Food Control 2014, 44, 203–207. [Google Scholar] [CrossRef]
- WHO; FAO; OIE. Taking a Multisectoral, One Health Approach: A Tripartite Guide to Addressing Zoonotic Diseases in Countries. 2019. Available online: https://www.woah.org/app/uploads/2021/03/en-tripartitezoonosesguide-webversion.pdf (accessed on 3 August 2022).
- Gordillo-Altamirano, F.L.; Barr, J.J. Phage therapy in the postantibiotic era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef]
- O’Neil, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. UK, 2014. Available online: https://amr-review.org (accessed on 29 July 2022).
- Jamal, M.; Bukhari, S.M.A.U.S.; Andleeb, S.; Ali, M.; Raza, S.; Nawaz, M.A.; Hussain, T.; Rahman, S.U.; Shah, S.S.A. Bacteriophages: An overview of the control strategies against multiple bacterial infections in different fields. J. Basic Microbiol. 2019, 59, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Nagel, T.E.; Chan, B.K.; De Vos, D.; El-Shibiny, A.; Kang’ethe, E.K.; Makumi, A.; Pirnay, J.-P. The Developing World Urgently Needs Phages to Combat Pathogenic Bacteria. Front. Microbiol. 2016, 7, 882. [Google Scholar] [CrossRef] [PubMed]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage applications for food production and processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Bolton, D.; McAuliffe, O.; Coffey, A. Bacteriophages in food applications: From foe to friend. Annu. Rev. Food Technol. 2019, 15, 151–172. [Google Scholar] [CrossRef]
- Young, I.; Wang, I.; Roof, W.D. Phages will out: Strategies of host cell lysis. Trends Microbiol. 2000, 8, 120–128. [Google Scholar] [CrossRef]
- Bernhardt, T.G.; Wang, I.N.; Struck, D.K.; Young, R. Breaking free: “protein antibiotics” and phage lysis. Res. Microbiol. 2002, 153, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A.; Woolston, J.; Sulakvelidze, A. Phage Biocontrol Applications in Food Production and Processing. Curr. Issues Mol. Biol. 2021, 40, 267–302. [Google Scholar] [CrossRef] [PubMed]
- Pirnay, J.P.; Merabishvili, M.; Van Raemdonck, H.; DeVis, D.; Verbeken, G. Bacteriophage production in compliance with regulatory requirements. In Bacteriophage Therapy: From Lab to Clinical Practice; Azeredo, J., Sillankorva, S., Eds.; Humana Press: New York, NY, USA, 2017; Book Series: Methods in Molecular Biology; Volume 1693, pp. 233–252. [Google Scholar] [CrossRef]
- Alomari, M.M.M.; Dec, M.; Urban-Chmiel, R. Bacteriophages as an Alternative Method for Control of Zoonotic and Foodborne Pathogens. Viruses 2021, 13, 2348. [Google Scholar] [CrossRef]
- Kutateladze, M.; Adamia, R. Bacteriophages as potential new therapeutics to replace or supplement antibiotics. Trends Biotechnol. 2010, 28, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii Infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef]
- Doss, J.; Culbertson, K.; Hahn, D.; Camacho, J.; Barekzi, N. A Review of Phage Therapy against Bacterial Pathogens of Aquatic and Terrestrial Organisms. Viruses 2017, 9, 50. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, L.D.; Bisha, B. Phage-based biocontrol strategies to reduce foodborne pathogens in foods. Bacteriophage 2011, 1, 130–137. [Google Scholar] [CrossRef]
- Sillankorva, S.M.; Oliveira, H.; Azeredo, J. Bacteriophages and their role in food safety. Inter. J. Microbiol 2012, 12, 863945. [Google Scholar] [CrossRef]
- Mosimann, S.; Desiree, K.; Ebner, P. Efficacy of Phage Therapy in Poultry: A Systematic Review and Meta-Analysis. Poult. Sci. 2021, 100, 101472. [Google Scholar] [CrossRef]
- Summers, W.C. Bacteriophage therapy. Annu. Rev. Microbiol. 2001, 55, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.R.; March, J.B. Bacteriophages and biotechnology: Vaccines, gene therapy and antibacterials. Trends Biotechnol. 2006, 24, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Atterbury, R.J.; Connerton, P.L.; Dodd, C.E.; Rees, C.E.; Connerton, I.F. Application of host specific bacteriophages to the surface of chicken skin leads to a reduction in recovery of Campylobacter jejuni. Appl. Environ. Microbiol. 2003, 69, 6302–6306. [Google Scholar] [CrossRef] [PubMed]
- Bueno, E.; García, P.; Martínez, B.; Rodríguez, A. Phage inactivation of Staphylococcus aureus in fresh and hard-type cheeses. Int. J. Food Microbiol. 2012, 158, 23–27. [Google Scholar] [CrossRef] [PubMed]
- EFSA; ECDC. The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, 7666. [Google Scholar] [CrossRef]
- Sevilla-Navarro, S.; Catalá-Gregori, P.; Marin, C. Salmonella Bacteriophage Diversity According to Most Prevalent Salmonella Serovars in Layer and Broiler Poultry Farms from Eastern Spain. Animals 2020, 10, 1456. [Google Scholar] [CrossRef]
- Sifre, E.; Salha, B.A.; Ducournau, A.; Floch, P.; Chardon, H.; Megraud, F.; Lehours, P. EUCAST recommendations for antimicrobial susceptibility testing applied to the three main Campylobacter species isolated in humans. J. Microbiol. Methods 2015, 119, 206–213. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef]
- WHO. Global Priority List of Antibiotic Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 29 July 2022).
- Olson, E.G.; Micciche, A.C.; Rothrock, M.J., Jr.; Yang, Y.; Ricke, S.C. Application of Bacteriophages to Limit Campylobacter in Poultry Production. Front. Microbiol. 2022, 12, 458721. [Google Scholar] [CrossRef]
- EFSA BIOHAZ Panel (European Food Safety Authority Panel on Biological Hazards). Update and review of control options for Campylobacter in broilers at primary production. EFSA J. 2020, 18, 6090. [Google Scholar] [CrossRef]
- Mota-Gutierrez, J.; Lis, L.; Lasagabaster, A.; Nafarrate, I.; Ferrocino, I.; Cocolin, L.; Rantsiou, K. Campylobacter spp. prevalence and mitigation strategies in the broiler production chain. Food Microbiol. 2022, 104, 103998. [Google Scholar] [CrossRef] [PubMed]
- Nafarrate, I.; Mateo, E.; Miranda, K.; Lasagabaster, A. Isolation, host specificity and genetic characterization of Campylobacter specific bacteriophages from poultry and swine sources. Food Microbiol. 2021, 97, 103742. [Google Scholar] [CrossRef]
- Sails, A.D.; Wareing, D.R.A.; Bolton, F.J.; Fox, A.J.; Curry, A. Characterisation of 16 Campylobacter jejuni and C. coli typing bacteriophages. J. Med. Microbiol. 1998, 47, 123–128. [Google Scholar] [CrossRef]
- D’Angelantonio, D.; Scattolini, S.; Boni, A.; Neri, D.; Di Serafino, G.; Connerton, P.; Connerton, I.; Pomilio, F.; Di Giannatale, E.; Migliorati, G.; et al. Bacteriophage Therapy to Reduce Colonization of Campylobacter jejuni in Broiler Chickens before Slaughter. Viruses 2021, 13, 1428. [Google Scholar] [CrossRef] [PubMed]
- Richards, P.J.; Connerton, P.L.; Connerton, I. Phage Biocontrol of Campylobacter jejuni in Chickens Does Not Produce Collateral Effects on the Gut Microbiota. Front. Microbiol. 2019, 10, 476. [Google Scholar] [CrossRef] [PubMed]
- Loc-Carrillo, C.; Atterbury, R.J.; El Shibiny, A.; Connerton, P.L. Bacteriophage therapy to reduce Campylobacter jejuni colonization of broiler chickens. Appl. Environ. Microbiol. 2005, 71, 6554–6563. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.M.; Gannon, B.W.; Halfhide, D.E.; Santos, S.B.; Hayes, C.M.; Roe, J.M.; Azeredo, J. The in vivo efficacy of two administration routes of a phage cocktail to reduce numbers of Campylobacter coli and Campylobacter jejuni in chickens. BMC Microbiol. 2010, 10, 232. [Google Scholar] [CrossRef] [PubMed]
- Kittler, S.; Fischer, S.; Abdulmawjood, A.; Glünder, G.; Klein, G. Effect of Bacteriophage Application on Campylobacter jejuni Loads in Commercial Broiler Flocks. Appl. Environ. Microbiol. 2013, 79, 7525–7533. [Google Scholar] [CrossRef]
- Chinivasagam, H.N.; Estella, W.; Maddock, L.; Mayer, D.G.; Weyand, C.; Connerton, P.L.; Connerton, I.F. Bacteriophages to control Campylobacter in commercially farmed broiler chickens in Australia. Front. Microbiol. 2020, 11, 632. [Google Scholar] [CrossRef]
- Wagenaar, J.A.; Van Bergen, M.A.; Mueller, M.A.; Wassenaar, T.M.; Carlton, R.M. Phage therapy reduces Campylobacter jejuni colonization in broilers. Veter. Microbiol. 2005, 109, 275–283. [Google Scholar] [CrossRef]
- El-Shibiny, A.; Scott, A.; Timms, A.; Metawea, Y.; Connerton, P.; Connerton, I. Application of a Group II Campylobacter Bacteriophage To Reduce Strains of Campylobacter jejuni and Campylobacter coli Colonizing Broiler Chickens. J. Food Prot. 2009, 72, 733–740. [Google Scholar] [CrossRef]
- Hammerl, J.A.; Jäckel, C.; Alter, T.; Janzcyk, P.; Stingl, K.; Knüver, M.T.; Hertwig, S. Reduction of Campylobacter jejuni in broiler chicken by successive application of group II and group III phages. PLoS ONE 2014, 9, e114785. [Google Scholar] [CrossRef] [PubMed]
- WHO. Salmonella (Non-Typhoidal). Available online: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed on 11 August 2022).
- Dawoud, T.M.; Davis, M.L.; Park, S.H.; Kim, S.A.; Kwon, Y.M.; Jarvis, N.; O’Bryan, C.A.; Shi, Z.; Crandall, P.G.; Ricke, S.C. The Potential Link between Thermal Resistance and Virulence in Salmonella: A Review. Front. Vet. Sci. 2017, 4, 93. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.S.; Jeong, J.; Lee, J.; Kim, S.; Min, W.; Myung, H. Therapeutic Effects of Bacteriophages against Salmonella Gallinarum Infection in Chickens. J. Microbiol. Biotechnol. 2013, 23, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Atterbury, R.J.; van Bergen, M.A.P.; Ortiz, F.; Lovell, M.A.; Harris, J.A.; de Boer, A.; Wagenaar, J.A.; Allen, V.M.; Barrow, P.A. Bacteriophage Therapy to Reduce Salmonella Colonization of Broiler Chickens. Appl. Environ. Microbiol. 2007, 73, 4543–4549. [Google Scholar] [CrossRef] [PubMed]
- Vaz, C.S.L.; Voss-Rech, D.; Alves, L.; Coldebella, A.; Brentano, L.; Trevisol, I.M. Effect of Time of Therapy with Wild-Type Lytic Bacteriophages on the Reduction of Salmonella Enteritidis in Broiler Chickens. Vet. Microbiol. 2020, 240, 108527. [Google Scholar] [CrossRef]
- Andreatti, R.L.; Higgins, J.P.; Higgins, S.E.; Gaona, G.; Wolfenden, A.D.; Tellez, G.; Hargis, B.M. Ability of bacteriophages isolated from different sources to reduce Salmonella enterica serovar Enteritidis in vitro and in vivo. Poult. Sci. 2007, 86, 1904–1909. [Google Scholar] [CrossRef]
- Borie, C.; Albala, I.; Sánchez, P.; Sánchez, M.L.; Ramírez, S.; Navarro, C.; Morales, M.A.; Retamales, J.; Robeson, J. Bacteriophage treatment reduces Salmonella colonization of infected chickens. Avian Dis. 2008, 52, 64–67. [Google Scholar] [CrossRef]
- Adhikari, P.A.; Cosby, D.E.; Cox, N.A.; Lee, J.H.; Kim, W.K. Effect of dietary bacteriophage supplementation on internal organs, fecal excretion, and ileal immune response in laying hens challenged by Salmonella Enteritidis. Poult. Sci. 2017, 96, 3264–3271. [Google Scholar] [CrossRef]
- Sevilla-Navarro, S.; Marín, C.; Cortés, V.; García, C.; Vega, S.; Catalá-Gregori, P. Autophage as a control measure for Salmonella in laying hens. Poult. Sci. 2018, 97, 4367–4373. [Google Scholar] [CrossRef]
- Wójcik, E.A.; Stańczyk, M.; Wojtasik, A.; Kowalska, J.D.; Nowakowska, M.; Łukasiak, M.; Bartnicka, M.; Kazimierczak, J.; Dastych, J. Comprehensive Evaluation of the Safety and Efficacy of BAFASAL® Bacteriophage Preparation for the Reduction of Salmonella in the Food Chain. Viruses 2020, 12, 742. [Google Scholar] [CrossRef] [PubMed]
- EFSA; ECDC. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from hu-mans, animals and food in 2019–2020. EFSA J. 2022, 20, 7209. [Google Scholar] [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine, 6th Revision 2018, Ranking of Medically Important Antimicrobials for Risk Management of Antimicrobial Resistance Due to Non-Human Use. 2018. Available online: https://apps.who.int/iris/bitstream/handle/10665/312266/9789241515528-eng.pdf (accessed on 29 July 2022).
- Carvalho, C.; Costa, A.R.; Silva, F.; Oliveira, A. Bacteriophages and Their Derivatives for the Treatment and Control of Food-Producing Animal Infections. Crit. Rev. Microbiol. 2017, 43, 583–601. [Google Scholar] [CrossRef] [PubMed]
- Nabil, N.M.; Tawakol, M.M.; Hassan, H.M. Assessing the Impact of Bacteriophages in the Treatment of Salmonella in Broiler Chickens. Infect. Ecol. Epidemiol. 2018, 8, 1539056. [Google Scholar] [CrossRef] [PubMed]
- Żbikowska, K.; Michalczuk, M.; Dolka, B. The Use of Bacteriophages in the Poultry Industry. Animals 2020, 10, 872. [Google Scholar] [CrossRef]
- Lim, T.-H.; Lee, D.-H.; Lee, Y.-N.; Park, J.-K.; Youn, H.-N.; Kim, M.-S.; Lee, H.-J.; Yang, S.-Y.; Cho, Y.-W.; Lee, J.-B.; et al. Efficacy of Bacteriophage Therapy on Horizontal Transmission of Salmonella Gallinarum on Commercial Layer Chickens. Avian Dis. 2011, 55, 435–438. [Google Scholar] [CrossRef]
- Nakari, U.M.; Rantala, L.; Pihlajasaari, A.; Toikkanen, S.; Johansson, T.; Hellsten, C.; Raulo, S.M.; Kuusi, M.; Siitonen, A.; Rimhanen-Finne, R. Investigation of increased listeriosis revealed two fishery production plants with persistent Listeria contamination in Finland in 2010. Epidemiol. Infect. 2014, 142, 2261–2269. [Google Scholar] [CrossRef]
- Skowron, K.; Kwiecinska, J.; Grudlewska, K.; Swieca, A.; Paluszak, Z.; Bauza, J.; Wałecka, E.; Gospodarek, E. The occurrence, transmission, virulence and antibiotic resistance of Listeria monocytogenes in fish processing plant. Int. J. Food Microbiol. 2018, 282, 71–83. [Google Scholar] [CrossRef]
- Morwal, S.; Sharma, S.K. Bacterial zoonosis—A public health importance. J. Dairy Vet. Anim. Res. 2017, 5, 56–59. [Google Scholar] [CrossRef]
- Loessner, M.; Carlton, R.M. Virulent Phages to Control Listeria monocytogenes in Foodstuffs and in Food Processing Plants. US Patent US7438901B2, 28 August 2008. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2004004495 (accessed on 13 October 2022).
- Li, T.; Zhao, X.; Wang, X.; Wang, Z.; Tian, C.; Shi, W.; Qi, Y.; Wei, H.; Song, C.; Xue, H.; et al. Characterization and Preliminary application of phage isolated from Listeria monocytogenes. Front. Vet. Sci. 2022, 9, 946814. [Google Scholar] [CrossRef]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef]
- Neetoo, H.; Reega, K.; Manoga, Z.S.; Nazurally, N.; Bhoyroo, V.; Allam, M.; Jaufeerally-Fakim, Y.; Ghoorah, A.W.; Jaumdally, W.; Hossen, A.M.; et al. Prevalence, genomic characterization, and risk assessment of human pathogenic Vibrio Species in Seafood. J. Food Prot. 2022, 85, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nat. Rev. Dis. Prim. 2018, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, M.; LewisOscar, F.; Thajuddin, N.; Pugazhendhi, A.; Nithya, C. In vitro and in vivo biofilm forming Vibrio spp: A significant threat in aquaculture. Process Biochem. 2020, 94, 213–223. [Google Scholar] [CrossRef]
- Plaza, N.; Castillo, D.; Pérez-Reytor, D.; Higuera, G.; García, K.; Bastías, R. Bacteriophages in the control of pathogenic vibrios. Electron. J. Biotechnol. 2018, 31, 24–33. [Google Scholar] [CrossRef]
- Cascarano, M.C.; Stavrakidis-Zachou, O.; Mladineo, I.; Thompson, K.D.; Papandroulakis, N.; Katharios, P. Mediterranean Aquaculture in a Changing Climate: Temperature Effects on Pathogens and Diseases of Three Farmed Fish Species. Pathogens 2021, 10, 1205. [Google Scholar] [CrossRef]
- Harrison, J.; Nelson, K.; Morcrette, H.; Morcrette, C.; Preston, J.; Helmer, L.; Titball, R.W.; Butler, C.S.; Wagley, S. The increased prevalence of Vibrio species and the first reporting of Vibrio jasicida and Vibrio rotiferianus at UK shellfish sites. Water Res. 2022, 211, 117942. [Google Scholar] [CrossRef]
- Scarano, C.; Spanu, C.; Ziino, G.; Pedonese, F.; Dalmasso, A.; Spanu, V.; Virdis, S.; De Santis, E.P.L. Antibiotic resistance of Vibrio species isolated from Sparus aurata reared in Italian mariculture. New Microbiol. 2014, 37, 329–337. [Google Scholar]
- Elmahdi, S.; DaSilva, L.V.; Parveen, S. Antibiotic resistance of Vibrio parahaemolyticus and Vibrio vulnificus in various countries: A review. Food Microbiol. 2016, 57, 128–134. [Google Scholar] [CrossRef]
- Yano, Y.; Hamano, K.; Satomi, M.; Tsutsui, I.; Ban, M.; Aue-umneoy, D. Prevalence and antimicrobial susceptibility of Vibrio species related to food safety isolated from shrimp cultured at inland ponds in Thailand. Food Control 2014, 38, 30–36. [Google Scholar] [CrossRef]
- European Commission. DG Maritime Affairs and Fisheries. Strategic Guidelines for a More Sustainable and Competitive EU Aquaculture for the Period 2021 to 2030; Document 52021DC0236, COM/2021/236 final; European Commission: Brussels, Belgium, 2021.
- Onarinde, B.A.; Dixon, R.A. Prospects for Biocontrol of Vibrio parahaemolyticus Contamination in Blue Mussels (Mytilus edulus)-A Year-Long Study. Front. Microbiol. 2018, 9, 1043. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.W.; Kim, H.J.; Yun, S.K.; Chai, J.Y.; Park, S.C. Eating oysters without risk of vibriosis: Application of a bacteriophage against Vibrio parahaemolyticus in oysters. Int. J. Food Microbiol. 2014, 188, 31–35. [Google Scholar] [CrossRef]
- Lee, J.H.; Oh, M.; Kim, B.S. Phage biocontrol of zoonotic food-borne pathogen Vibrio parahaemolyticus for seafood safety. Food Control 2023, 144, 109334. [Google Scholar] [CrossRef]
- Vinod, M.G.; Shivu, M.M.; Umesha, K.R.; Rajeeva, B.C.; Krohneb, G. Isolation of Vibrio harveyi bacteriophage with a potential for biocontrol of luminous vibriosis in hatchery environments. Aquaculture 2006, 255, 117–124. [Google Scholar] [CrossRef]
- Droubogiannis, S.; Katharios, P. Genomic and biological profile of a novel Vibrio phage, Virtus, which improves survival of Sparus aurata larvae challenged with Vibrio harveyi. Pathogens 2022, 11, 630. [Google Scholar] [CrossRef] [PubMed]
- Higuera, G.; Bastías, R.; Tsertsvadze, G.; Romero, J.; Espejo, R.T. Recently discovered Vibrio anguillarum phages can protect against experimentally induced vibriosis in Atlantic salmon, Salmo salar. Aquaculture 2013, 395, 128–133. [Google Scholar] [CrossRef]
- Silva, Y.J.; Costa, L.; Pereira, C.; Mateus, C.; Cunha, Â.; Calado, R.; Gomes, N.; Pardo, M.A.; Hernandez, I.; Almeida, A. Phage Therapy as an Approach to Prevent Vibrio anguillarum Infections in Fish Larvae Production. PLoS ONE 2014, 9, e114197. [Google Scholar] [CrossRef]
- Lomelí-Ortega, C.O.; Martínez-Díaz, S.F. Phage therapy against Vibrio parahaemolyticus infection in the whiteleg shrimp (Litopenaeus vannamei) larvae. Aquaculture 2014, 434, 208–211. [Google Scholar] [CrossRef]
- Karunasagar, I.; Shivu, M.M.; Girisha, S.K.; Krohne, G.; Karunasagar, I. Biocontrol of pathogens in shrimp hatcheries using bacteriophages. Aquaculture 2007, 268, 288–292. [Google Scholar] [CrossRef]
- Nakai, T.; Park, S.C. Bacteriophage therapy of infectious diseases in aquaculture. Res. Microbiol. 2002, 153, 13–18. [Google Scholar] [CrossRef]
- Lasagabaster, A.; Jiménez, E.; Lehnherr, T.; Miranda-Cadena, K.; Lehnherr, H. Bacteriophage biocontrol to fight Listeria outbreaks in seafood. Food Chem. Toxicol. 2020, 145, 111682. [Google Scholar] [CrossRef] [PubMed]
- Sulakvelidze, A. Using lytic bacteriophages to eliminate or significantly reduce contamination of food by foodborne bacterial pathogens. J. Sci. Food Agric. 2013, 93, 3137–3146. [Google Scholar] [CrossRef] [PubMed]
- Bolocan, A.S.; Callanan, J.; Forde, A.; Ross, P.; Hill, C. Phage therapy targeting Escherichia coli-a story with no end? FEMS Microbiol. Lett. 2016, 363, fnw256. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Fernández, L.; Martínez, B.; Ruas-Madiedo, P.; García, P.; Rodríguez, A. Real-Time Assessment of Staphylococcus aureus Biofilm Disruption by Phage-Derived Proteins. Front. Microbiol. 2017, 8, 1632. [Google Scholar] [CrossRef]
- Hagens, S.; Loessner, M.J. Application of bacteriophages for detection and control of foodborne pathogens. Appl. Microbiol. Biotechnol. 2007, 76, 513–519. [Google Scholar] [CrossRef]
- Goode, D.; Allen, V.M.; Barrow, P.A. Reduction of experimental Salmonella and Campylobacter contamination of chicken skin by application of lytic bacteriophages. Appl. Environ. Microbiol. 2003, 69, 5032–5036. [Google Scholar] [CrossRef]
- Zampara, A.; Sørensen, M.C.; Elsser-Gravesen, A.; Brøndsted, L. Significance of phage-host interactions for biocontrol of Campylobacter jejuni in food. Food Control 2017, 73, 1169–1175. [Google Scholar] [CrossRef]
- Orquera, S.; Golz, G.; Hertwig, S.; Hammerl, J.; Sparborth, D.; Joldic, A.; Alter, T. Control of Campylobacter spp. and Yersinia enterocolitica by virulent bacteriophages. J. Mol. Genet. Med. 2012, 6, 273–278. [Google Scholar] [CrossRef]
- Bigwood, T.; Hudson, J.A.; Billington, C.; Carey-Smith, G.V.; Heinemann, J.A. Phage inactivation of foodborne pathogens on cooked and raw meat. Food Microbiol. 2008, 25, 400–406. [Google Scholar] [CrossRef]
- Thung, T.Y.; Lee, E.; Mahyudin, N.A.; Radzi, C.W.J.; Mazlan, N.; Tan, C.W.; Radu, S. Partial characterization and in vitro evaluation of a lytic bacteriophage for biocontrol of Campylobacter jejuni in mutton and chicken meat. J. Food Saf. 2020, 40, e12770. [Google Scholar] [CrossRef]
- Siringan, P.; Connerton, P.L.; Payne, R.J.H.; Connerton, I.F. Bacteriophage-mediated dispersal of Campylobacter jejuni biofilms. Appl. Environ. Microbiol. 2011, 77, 3320–3326. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Higgins, S.E.; Guenther, K.L.; Huff, W.; Donoghue, A.M.; Donoghue, D.J.; Hargis, B.M. Use of a Specific Bacteriophage Treatment to Reduce Salmonella in Poultry Products. Poult. Sci. 2005, 84, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Spricigo, D.A.; Bardina, C.; Cortés, P.; Llagostera, M. Use of a Bacteriophage Cocktail to Control Salmonella in Food and the Food Industry. Int. J. Food Microbiol. 2013, 165, 169–174. [Google Scholar] [CrossRef]
- Thung, T.Y.; Krishanthi Jayarukshi Kumari Premarathne, J.M.; San Chang, W.; Loo, Y.Y.; Chin, Y.Z.; Kuan, C.H.; Tan, C.W.; Basri, D.F.; Radzi, C.W.J.W.M.; Radu, S. Use of a Lytic Bacteriophage to Control Salmonella Enteritidis in Retail Food. LWT 2017, 78, 222–225. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, P.; Zhang, H.; Zhou, Y.; Zhang, L.; Wang, R. Bio-Control of Salmonella Enteritidis in Foods Using Bacteriophages. Viruses 2015, 7, 4836–4853. [Google Scholar] [CrossRef]
- Sukumaran, A.T.; Nannapaneni, R.; Kiess, A.; Sharma, C.S. Reduction of Salmonella on chicken breast fillets stored under aerobic or modified atmosphere packaging by the application of lytic bacteriophage preparation SalmoFreshTM. Poult. Sci. 2016, 95, 668–675. [Google Scholar] [CrossRef]
- Soffer, N.; Abuladze, T.; Woolston, J.; Li, M.; Hanna, L.F.; Heyse, S.; Charbonneau, D.; Sulakvelidze, A. Bacteriophages safely reduce Salmonella contamination in pet food and raw pet food ingredients. Bacteriophage 2016, 6, e1220347. [Google Scholar] [CrossRef]
- Moon, S.H.; Waite-Cusic, J.; Huang, E. Control of Salmonella in chicken meat using a combination of a commercial bacteriophage and plant-based essential oils. Food Control 2020, 110, 106984. [Google Scholar] [CrossRef]
- Hagens, S.; de Vegt, B.; Peterson, R. Efficacy of a Commercial Phage Cocktail in Reducing Salmonella Contamination on Poultry Products: Laboratory Data and Industrial Trial Data. Meat Muscle Biol. 2018, 2, 156. [Google Scholar] [CrossRef]
- Soni, K.A.; Nannapaneni, R. Bacteriophage significantly reduces Listeria monocytogenes on raw salmon fillet tissue. J. Food Prot. 2010, 73, 32–38. [Google Scholar] [CrossRef]
- Baños, A.; García-López, J.D.; Núñez, C.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Biocontrol of Listeria monocytogenes in fish by enterocin AS-48 and Listeria lytic bacteriophage P100. LWT 2016, 66, 672–677. [Google Scholar] [CrossRef]
- Perera, M.N.; Abuladze, T.; Li, M.; Woolston, J.; Sulakvelidze, A. Bacteriophage cocktail significantly reduces or eliminates Listeria monocytogenes contamination on lettuce, apples, cheese, smoked salmon and frozen foods. Food Microbiol. 2015, 52, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Bigot, B.; Lee, W.J.; McIntyre, L.; Wilson, T.; Hudson, J.A.; Billington, C.; Heinemann, J. Control of Listeria monocytogenes growth in a ready-to-eat poultry product using a bacteriophage. Food Microbiol. 2011, 28, 1448–1452. [Google Scholar] [CrossRef] [PubMed]
- Chibeu, A.; Agius, L.; Gao, A.; Sabour, P.M.; Kropinski, A.M.; Balamurugan, S. Efficacy of bacteriophage LISTEX™P100 combined with chemical antimicrobials in reducing Listeria monocytogenes in cooked turkey and roast beef. Int. J. Food Microbiol. 2013, 167, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Holck, A.; Berg, J. Inhibition of Listeria monocytogenes in cooked ham by virulent bacteriophages and protective cultures. Appl. Environ. Microbiol. 2009, 75, 6944–6946. [Google Scholar] [CrossRef]
- Iacumin, L.; Manzano, M.; Comi, G. Phage inactivation of Listeria monocytogenes on San Daniele dry-cured ham and elimination of biofilms from equipment and working environments. Microorganisms 2016, 4, 4. [Google Scholar] [CrossRef]
- Lee, S.; Kim, M.G.; Lee, H.S.; Heo, S.; Kwon, M.; Kim, G. Isolation and Characterization of Listeria phages for Control of Growth of Listeria monocytogenes in Milk. Korean J. Food Sci. Anim. Resour. 2017, 37, 320–328. [Google Scholar] [CrossRef]
- Soni, K.A.; Desai, M.; Oladunjove, A.; Skrobot, F.; Nannapaneni, R. Reduction of Listeria monocytogenes in queso fresco cheese by a combination of listericidal and listeriostatic GRAS antimicrobials. Int. J. Food Microbiol. 2012, 155, 82–88. [Google Scholar] [CrossRef]
- Guenther, S.; Loessner, M.J. Bacteriophage biocontrol of Listeria monocytogenes on soft ripened white mold and red- smear cheeses. Bacteriophage 2011, 1, 94–100. [Google Scholar] [CrossRef]
- Leverentz, B.; Conway, W.S.; Camp, M.J.; Janisiewicz, W.J.; Abuladze, T.; Yang, M.; Saftner, R.; Sulakvelidze, A. Biocontrol of Listeria monocytogenes on Fresh-Cut Produce by Treatment with Lytic Bacteriophages and a Bacteriocin. Appl. Environ. Microbiol. 2003, 69, 4519–4526. [Google Scholar] [CrossRef]
- Oliveira, M.; Viñas, I.; Colàs, P.; Anguera, M.; Usall, J.; Abadias, M. Effectiveness of a bacteriophage in reducing Listeria monocytogenes on fresh-cut fruits and fruit juices. Food Microbiol. 2014, 38, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Byun, K.H.; Han, S.H.; Choi, M.W.; Park, S.H.; Ha, S.D. Isolation, characterization, and application of bacteriophages to reduce and inhibit Listeria monocytogenes in celery and enoki mushroom. Food Control 2022, 135, 108826. [Google Scholar] [CrossRef]
- You, H.J.; Lee, J.H.; Oh, M.; Hong, S.Y.; Kim, D.; Noh, J.; Kim, M.; Kim, B.S. Tackling Vibrio parahaemolyticus in ready-to-eat raw fish flesh slices using lytic phage VPT02 isolated from market oyster. Food Res. Int. 2021, 150, 110779. [Google Scholar] [CrossRef]
- Xia, H.; Yang, H.; Yan, N.; Hou, W.; Wang, H.; Wang, X.; Wang, H.; Zhou, M. Bacteriostatic effects of phage F23s1 and its endolysin on Vibrio parahaemolyticus. J. Basic Microbiol. 2022, 62, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.J.; Hong, J.; Lee, N.; Chun, H.S.; Kim, H.Y.; Choi, S.W.; Ok, G. Growth inhibitory effect of bacteriophages isolated from western and southern coastal areas of Korea against Vibrio parahaemolyticus in Manila clams. Appl. Biol. Chem. 2016, 59, 359–365. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Z.; Zhou, Y.; Bao, H.; Wang, R.; Li, T.; Pang, M.; Sun, L.; Zhou, X. Application of a Phage in Decontaminating Vibrio Parahaemolyticus in Oysters. Int. J. Food Microbiol. 2018, 275, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Pelon, W.; Luftig, R.B.; Johnston, K.H. Vibrio vulnificus load reduction in oysters after combined exposure to Vibrio vulnificus--specific bacteriophage and to an oyster extract component. J. Food Prot. 2005, 68, 1188–1191. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, Y.T.; Kim, H.B.; Choi, S.H.; Lee, J.H. Characterization of bacteriophage VVP001 and its application for the inhibition of V. vulnificus causing seafood-borne diseases. Food Microbiol. 2021, 94, 103630. [Google Scholar] [CrossRef]
- Aguilera, M.; Martínez, S.; Tello, M.; Gallardo, M.J.; García, V. Use of Cocktail of Bacteriophage for Salmonella Typhimurium Control in Chicken Meat. Foods 2022, 11, 1164. [Google Scholar] [CrossRef]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial Biofilms in the Food Industry—A Comprehensive Review. Int. J. Environ. Res. Public Health 2021, 18, 14. [Google Scholar] [CrossRef]
- Korzeniowski, P.; Śliwka, P.; Kuczkowski, M.; Mišić, D.; Milcarz, A.; Kuźmińska-Bajor, M. Bacteriophage cocktail can effectively control Salmonella biofilm in poultry housing. Front. Microbiol. 2022, 13, 2317. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Vermeulen, A.; Gysemans, K.P.M.; Bernaerts, K.; Geeraerd, A.H.; Van Impe, J.F.; Debevere, J.; Devlieghere, F. Influence of pH, water activity and acetic acid concentration on Listeria monocytogenes at 7 °C: Data collection for the development of a growth/no growth model. Int. J. Food Microbiol. 2007, 114, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.B.; Mujahid, S.; Orsi, R.H.; Bergholz, T.M.; Wiedmann, M.; Boor, K.; Destro, M. Contributions of σB and PrfA to Listeria monocytogenes salt stress under food relevant conditions. Int. J. Food Microbiol. 2014, 177, 98–108. [Google Scholar] [CrossRef]
- Makariti, I.P.; Grivokostopoulos, N.C.; Skandamis, P.N. Effect οf οxygen availability and pH οn adaptive acid tolerance response of immobilized Listeria monocytogenes in structured growth media. Food Microbiol. 2021, 99, 103826. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Langsrud, S.; Møretrø, T. Microbial diversity and ecology of biofilms in food industry environments associated with Listeria monocytogenes persistence. Curr. Opin. Food Sci. 2021, 37, 171–178. [Google Scholar] [CrossRef]
- Forauer, E.; Tongyu Wu, S.; Etter, A.J. Listeria monocytogenes in the retail deli environment: A review. Food Control 2021, 119, 107443. [Google Scholar] [CrossRef]
- Belias, A.; Sullivan, G.; Wiedmann, M.; Ivanek, R. Factors that contribute to persistent Listeria in food processing facilities and relevant interventions: A rapid review. Food Control 2022, 133, 108579. [Google Scholar] [CrossRef]
- Carlton, R.M.; Noordman, W.H.; Biswas, B.; De Meester, E.D.; Loessner, M.J. Bacteriophage P100 for control of Listeria monocytogenes in foods: Genome sequence, bioinformatic analyses, oral toxicity study and application. Regul. Toxicol. Pharmacol. 2005, 43, 301–312. [Google Scholar] [CrossRef]
- Schmuki, M.M.; Erne, D.; Loessner, M.J.; Klumpp, J. Bacteriophage P70: Unique morphology and unrelatedness to other Listeria bacteriophages. J. Virol. 2012, 86, 13099–13102. [Google Scholar] [CrossRef]
- Zhou, C.; Zhu, M.; Wang, Y.; Yang, Z.; Ye, M.; Wu, L.; Bao, H.; Pang, M.; Zhou, Y.; Wang, R.; et al. Broad host range phage vBLmoM-SH3-3 reduces the risk of Listeria contamination in two types of ready-to-eat food. Food Control 2020, 108, 106830. [Google Scholar] [CrossRef]
- Kawacka, I.; Olejnik-Schmidt, A.; Schmidt, M.; Sip, A. Effectiveness of phage-based inhibition of Listeria monocytogenes in food products and food processing environments. Microorganisms 2020, 8, 1764. [Google Scholar] [CrossRef] [PubMed]
- EFSA BIOHAZ Panel. Scientific opinion on the evaluation of the safety and efficacy of ListexTM P100 for reduction of pathogens on different ready-to-eat (RTE) food products. EFSA J. 2016, 14, e04565. [Google Scholar] [CrossRef]
- Gray, J.A.; Chandry, P.S.; Kaur, M.; Kocharunchitt, C.; Bowman, J.P.; Fox, E.M. Novel Biocontrol Methods for Listeria monocytogenes Biofilms in Food Production Facilities. Front. Microbiol. 2018, 9, 605. [Google Scholar] [CrossRef] [PubMed]
- Brives, C.; Pourraz, J. Phage therapy as a potential solution in the fight against AMR: Obstacles and possible futures. Palgrave Commun. 2020, 6, 100. [Google Scholar] [CrossRef]
- Ferriol-González, C.; Domingo-Calap, P. Phages for Biofilm Removal. Antibiotics 2020, 9, 268. [Google Scholar] [CrossRef]
- Mateus, L.; Costa, L.; Silva, Y.J.; Pereira, C.; Cunha, A.; Almeida, A. Efficiency of phage cocktails in the inactivation of Vibrio in aquaculture. Aquaculture 2014, 424–425, 167–173. [Google Scholar] [CrossRef]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef]
- Wandro, S.; Ghatbale, P.; Attai, H.; Hendrickson, C.; Samillano, C.; Suh, J.; Pride, D.T.; Whiteson, K. Phage cocktails can prevent the evolution of phage-resistant Enterococcus. BioRxiv 2021. [Google Scholar] [CrossRef]
- Kaur, S.; Kumari, A.; Kumari Negi, A.; Galav, V.; Thakur, S.; Agrawal, M.; Sharma, V. Nanotechnology Based Approaches in Phage Therapy: Overcoming the Pharmacological Barriers. Front. Pharmacol. 2021, 12, 699054. [Google Scholar] [CrossRef]
- Loh, B.; Gondil, V.S.; Manohar, P.; Khan, F.M.; Yang, H.; Leptihn, S. Encapsulation and delivery of therapeutic phages. Appl. Environ. Microbiol 2020, 87, e01979-20. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.A.; Liu, H.; Wang, Q.; Zhong, F.; Guo, Q.; Balamurugan, S. Use of encapsulated bacteriophages to enhance farm to fork food safety. Crit. Rev. Food Sci. 2017, 57, 2801–2810. [Google Scholar] [CrossRef] [PubMed]
- Danis-Wlodarczyk, K.; Dąbrowska, K.; Abedon, S.T. Phage Therapy: The Pharmacology of Antibacterial Viruses. Curr. Issues Mol. Biol. 2021, 40, 81–164. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rubio, L.; Jofre, J.; Muniesa, M. Is Genetic Mobilization Considered When Using Bacteriophages in Antimicrobial Therapy? Antibiotics 2017, 6, 32. [Google Scholar] [CrossRef]
- Abedon, S.T.; Danis-Wlodarczyk, K.M.; Wozniak, D.J. Phage Cocktail Development for Bacteriophage Therapy: Toward Improving Spectrum of Activity Breadth and Depth. Pharmaceuticals 2021, 14, 1019. [Google Scholar] [CrossRef] [PubMed]
- Nagel, T.; Musila, L.; Muthoni, M.; Nikolich, M.; Nakavuma, J.L.; Clokie, M.R. Phage banks as potential tools to rapidly and cost-effectively manage antimicrobial resistance in the developing world. Curr. Opin. Virol. 2022, 53, 101208. [Google Scholar] [CrossRef]
- Domingo-Calap, P.; Delgado-Martínez, J. Bacteriophages: Protagonists of a Post-Antibiotic Era. Antibiotics 2018, 7, 66. [Google Scholar] [CrossRef]
- Leptihn, S.; Loh, B. Complexity, challenges and costs of implementing phage therapy. Future Microbiol. 2022, 17, 643–646. [Google Scholar] [CrossRef]
- Vázquez, R.; Díez-Martínez, R.; Domingo-Calap, P.; García, P.; Gutiérrez, D.; Muniesa, M.; Ruiz-Ruigómez, M.; Sanjuán, R.; Tomás, M.; Tormo-Mas, M.; et al. Essential topics for the regulatory consideration of phages as clinically valuable therapeutic agents: A perspective from Spain. Microorganisms 2022, 10, 717. [Google Scholar] [CrossRef]


| Animal | Bacteria Load 1 | Phage | Application Method and Dose 2 | Bacterial Reduction | Ref. |
|---|---|---|---|---|---|
| Mussels | V. parahaemolyticus, naturally infected | Phage cocktail: VP10 | Immersion ~103 | Undetectable levels in seawater, sediment, or mussels after 48 h | [79] |
| Oysters | V. parahaemolyticus CRS 09-17, AMR * 2.7 × 106 | pVp-1 | Immersion 1.6 × 107 | Growth reduction >5 log after 72 h | [80] |
| Aquatic crustacean Artemia franciscana | V. parahaemolyticus FORC_023 104 | VPG01 | Immersion 103, 104, 105 | 2 log reduction Increased survival from 10% to 40% with higher phage concentration (104 and 105 PFU/mL). | [81] |
| Shrimps (Penaeus monodon) larvae | V. harveyi 105 | Bacteriophage of V. harveyi | Immersion 2 × 105 | 3 log reduction in bacterial counts Increased larvae survival from 17% to 86% More effective than antibiotics (40% survival) | [82] |
| Gilthead seabream larvae | V. harveyi VH2 106 | Virtus | Immersion 107 | Increased survival of larvae: from <6% to >40% | [83] |
| Atlantic salmon | V. anguillarum PF4 5 × 105 | CHOED | Immersion (100L) 5 × 105 Immersion (100L) 106 Immersion (farm conditions) 5 × 107 | Increased survival from 5% to 70% after 10 d Increased survival from 5% to 100% after 10 d Increased survival from 65% to 100% after 9 d (protection up to 20 d) | [84] |
| Zebrafish larvae | V. anguillarum 106 | VP-2 phage | Immersion 108 | Increased survival from 83% to 98% after 72 h | [85] |
| Whiteleg shrimp larvae | V. parahaemolyticus ATCC 17802 2 × 106 | A3S Vpms1 | Immersion 105, 106,107 | Increased survival of larvae from <60% to 80% (A3S phage), depending on the conditions | [86] |
| Shrimps (Penaeus monodon) | V. harveyi | Viha10 Viha8 | Immersion 2 × 106 | Increased survival of larvae to >86% vs. ~65% survival with antibiotics | [87] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavilla, M.; Domingo-Calap, P.; Sevilla-Navarro, S.; Lasagabaster, A. Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective. Foods 2023, 12, 552. https://doi.org/10.3390/foods12030552
Lavilla M, Domingo-Calap P, Sevilla-Navarro S, Lasagabaster A. Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective. Foods. 2023; 12(3):552. https://doi.org/10.3390/foods12030552
Chicago/Turabian StyleLavilla, Maria, Pilar Domingo-Calap, Sandra Sevilla-Navarro, and Amaia Lasagabaster. 2023. "Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective" Foods 12, no. 3: 552. https://doi.org/10.3390/foods12030552
APA StyleLavilla, M., Domingo-Calap, P., Sevilla-Navarro, S., & Lasagabaster, A. (2023). Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective. Foods, 12(3), 552. https://doi.org/10.3390/foods12030552