
Freshness Analysis of Raw Laver (Pyropia yenzoensis) Conserved under Supercooling Conditions


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Storage Methods
2.3. Drip Loss
2.4. Moisture Content
2.5. pH
2.6. Thiobarbituric Acid Reactive Substance (TBARS)
2.7. Volatile Basic Nitrogen (VBN)
2.8. Total Aerobic Count (TAC)
2.9. Color
2.10. Pigment Contents
2.11. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.12. Free Amino Acid Content
2.13. Statistical Analysis
3. Results and Discussion
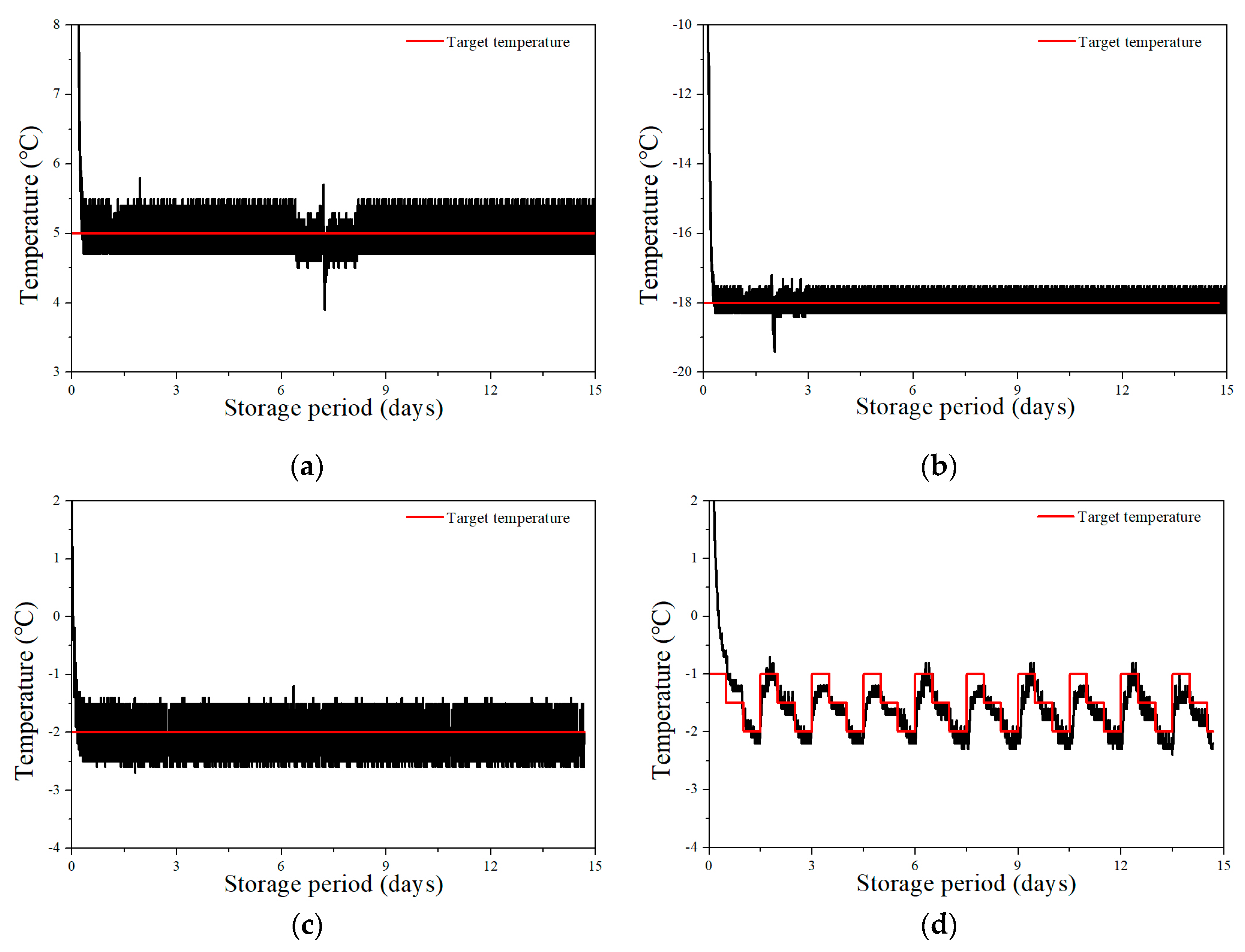
3.1. Time–Temperature Curves
3.2. Drip Loss, Moisture Content, and pH
3.2.1. Drip Loss
3.2.2. Moisture Content
3.2.3. pH
3.3. Color and Pigment Contents
3.3.1. Appearance and Color
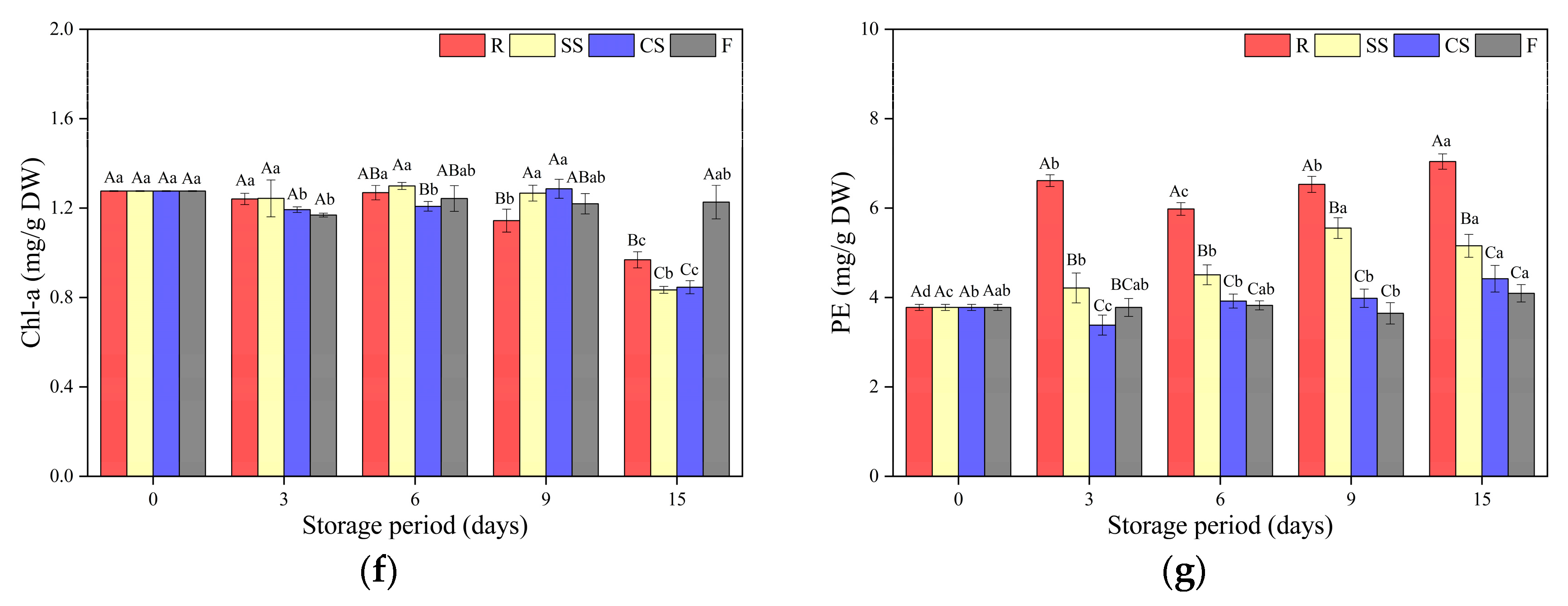
3.3.2. Pigment Content
3.4. TBARS, VBN, and TAC
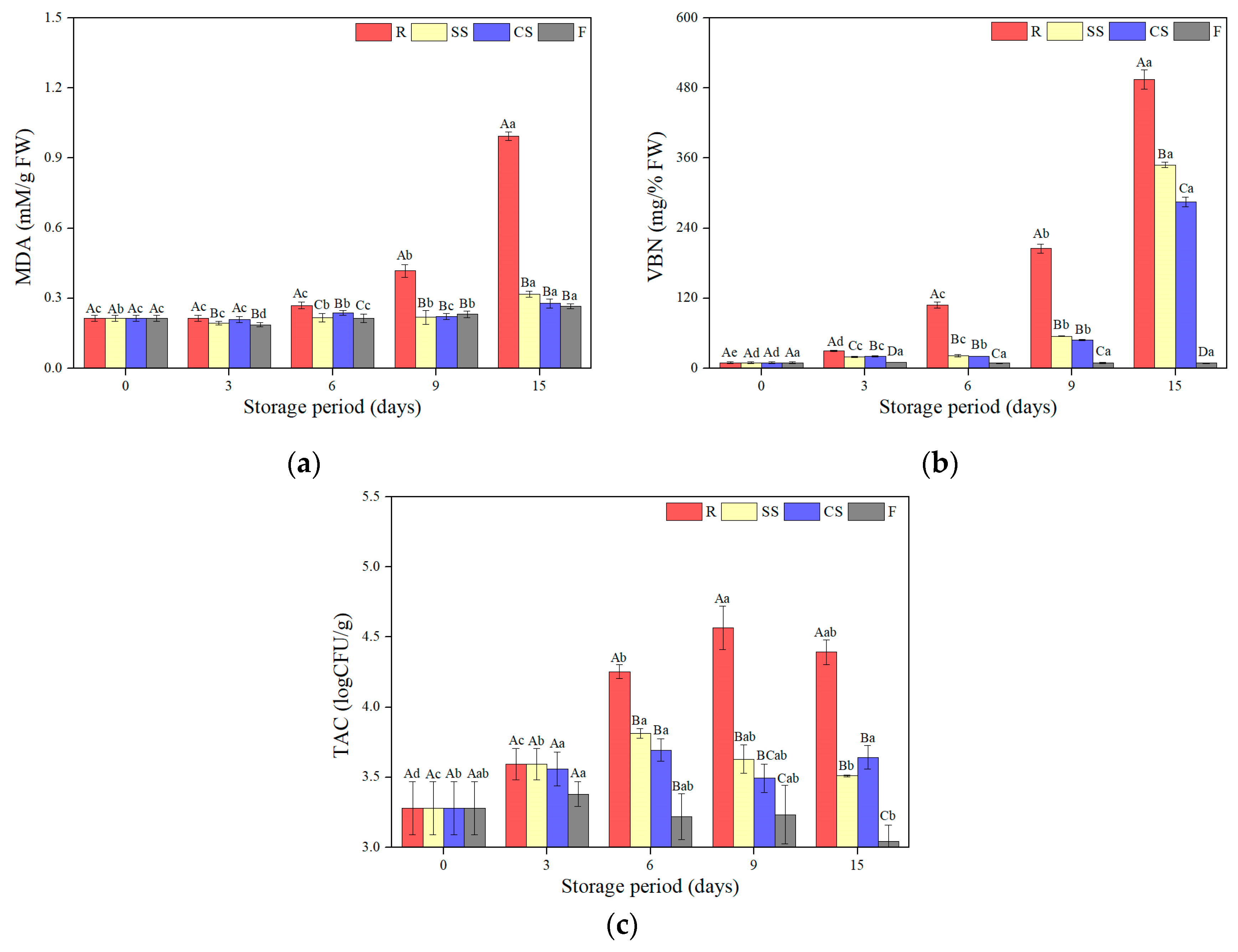
3.4.1. TBARS
3.4.2. VBN
3.4.3. TAC
3.5. SDS-PAGE
3.6. Free Amino Acid Concentration
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bito, T.; Teng, F.; Watanabe, F. Bioactive compounds of edible purple laver Porphyra sp. (Nori). J. Agric. Food Chem. 2017, 65, 10685–10692. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Hwang, J.H.; Oh, M.J.; Kim, M.Y.; Choi, M.R.; Park, W.M. Studies on the major nutritional components of commercial dried lavers (Porphyra yezoensis) cultivated in Korea. Korean J. Food Preserv. 2014, 21, 702–709. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, J.U.; Jang, Y.S. A study on the management efficiency of laver drying-processing company. J. Fish Bus. Adm. 2018, 49, 37–50. [Google Scholar] [CrossRef]
- Baek, E.Y. A study on the current state and problems of laver drying-processing industry. Korean Soc. Fish Sci. Educ. 2020, 32, 713–724. [Google Scholar]
- Kim, B.H.; Lim, D.H.; Yi, J.H. Economical meaning and problem concerning industrial differentiation of laver industry. J. Fish Bus. Adm. 2016, 47, 047–061. [Google Scholar] [CrossRef]
- Green, L.A.; Neefus, C.D. The effects of short- and long-term freezing on Porphyra umbilicalis Kützing (Bangiales, Rhodophyta) blade viability. J. Exp. Mar. Biol. Ecol. 2014, 461, 499–503. [Google Scholar] [CrossRef]
- Park, D.H.; Lee, S.Y.; Kim, E.J.; Jo, Y.J.; Choi, M.J. Development of stepwise algorithm for supercooling storage of pork belly and chicken breast and its effect on freshness. Foods 2022, 11, 380. [Google Scholar] [CrossRef]
- Li, D.; Zhu, Z.; Sun, D.W. Effects of freezing on cell structure of fresh cellular food materials: A review. Trends Food Sci. Technol. 2018, 75, 46–55. [Google Scholar] [CrossRef]
- Park, D.H.; Lee, S.Y.; Lee, J.; Kim, E.J.; Jo, Y.J.; Kim, H.; Choi, M.J.; Hong, G.P. Stepwise cooling mediated feasible supercooling preservation to extend freshness of mackerel fillets. LWT Food Sci. Technol. 2021, 152, 112389. [Google Scholar] [CrossRef]
- Stonehouse, G.G.; Evans, J.A. The use of supercooling for fresh foods: A review. J. Food Eng. 2015, 148, 74–79. [Google Scholar] [CrossRef]
- Lee, S.Y.; Park, D.H.; Kim, E.J.; Kim, H.; Lee, Y.J.; Choi, M.J. Development of temperature control algorithm for supercooling storage of pork loin and its feasibility for improving freshness and extending shelf life. Food Sci. Anim. Resour. 2022, 42, 467–485. [Google Scholar] [CrossRef]
- Kang, T.; You, Y.; Jun, S. Supercooling preservation technology in food and biological samples: A review focused on electric and magnetic field applications. Food Sci. Biotechnol. 2020, 29, 303–321. [Google Scholar] [CrossRef]
- Kim, E.J. Development of Novel Low-Temperature Preservation Technology to Extend Freshness and Wholesomeness of Foods. Doctoral Dissertation, Research Information Sharing Service, Konkuk University, Seoul, Republic of Korea, February 2021. [Google Scholar]
- Fukuma, Y.; Yamane, A.; Itoh, T.; Tsukamasa, Y.; Ando, M. Application of supercooling to long-term storage of fish meat. Fish Sci. 2012, 78, 451–461. [Google Scholar] [CrossRef]
- Kim, Y.; Hong, G.P. Effects of artificial supercooling followed by slow freezing on the microstructure and qualities of pork loin. Korean J. Food Sci. Anim. Resour. 2016, 36, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Obluchinskaya, E.; Daurtseva, A. Effects of air drying and freezing and long-term storage on phytochemical composition of brown seaweeds. J. Appl. Phycol. 2020, 32, 4235–4249. [Google Scholar] [CrossRef]
- Davison, I.R.; Dudgeon, S.R.; Ruan, H.M. Effect of freezing on seaweed photosynthesis. Mar. Ecol. Prog. Ser. 1989, 58, 123–131. [Google Scholar] [CrossRef]
- Wang, W.-J.; Wang, F.-J.; Zhu, J.-Y.; Sun, X.-T.; Yao, C.-Y.; Xu, P. Freezing tolerance of Porphyra yezoensis (Bangiales, Rhodophyta) gametophyte assessed by chlorophyll fluorescence. J. Appl. Phycol. 2011, 23, 1017–1022. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 19th ed.; AOAC International: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Chappelle, E.; Kim, M.; McMurtrey, J., III. Ratio analysis of reflectance spectra (RARS): An algorithm for the remote estimation of the concentrations of chlorophyll A, chlorophyll B, and carotenoids in soybean leaves. Remote Sens. Environ. 1992, 39, 239–247. [Google Scholar] [CrossRef]
- Beer, S.; Eshel, A. Determining phycoerythrin and phycocyanin concentrations in aqueous crude extracts of red algae. Mar. Freshw. Res. 1985, 36, 785–792. [Google Scholar] [CrossRef]
- Shin, H.; Kim, H.T.; Choi, M.J.; Ko, E.Y. Effects of bromelain and double emulsion on the physicochemical properties of pork loin. Food Sci. Anim. Resour. 2019, 39, 888–902. [Google Scholar] [CrossRef]
- Nayyar, D.; Skonberg, D.I. Contrasting effects of two storage temperatures on the microbial, physicochemical, and sensory properties of two fresh red seaweeds, Palmaria palmata and Gracilaria tikvahiae. J. Appl. Phycol. 2019, 31, 731–739. [Google Scholar] [CrossRef]
- Son, S.M.; Kwon, H.O.; Lee, J.H. Physicochemical composition of Capsosiphon fulvescens according to drying methods. J. Korean Soc. Food Sci. Nutr. 2011, 40, 1582–1588. [Google Scholar] [CrossRef]
- Choi, M.J.; Abduzukhurov, T.; Park, D.H.; Kim, E.J.; Hong, G.P. Effects of deep freezing temperature for long-term storage on quality characteristics and freshness of lamb meat. Korean J. Food Sci. Anim. Resour. 2018, 38, 959–969. [Google Scholar] [CrossRef]
- Jo, H.; Surh, J. Comparison of the antioxidant activities of sulfur-fortified onions stored at room temperature and 1 °C. J. Korean Soc. Food Sci. Nutr. 2020, 49, 1218–1229. [Google Scholar] [CrossRef]
- Zhu, D.; Liang, J.; Liu, H.; Cao, X.; Ge, Y.; Li, J. Sweet cherry softening accompanied with moisture migration and loss during low-temperature storage. J. Sci. Food Agric. 2018, 98, 3651–3658. [Google Scholar] [CrossRef]
- Sánchez-García, F.; Hernández, I.; Palacios, V.M.; Roldán, A.M. Freshness quality and shelf life evaluation of the seaweed Ulva rigida through physical, chemical, microbiological, and sensory methods. Foods 2021, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jia, S.; Hong, H.; Zhang, L.; Zhuang, S.; Sun, X.; Liu, X.; Luo, Y. Assessment of bacterial contributions to the biochemical changes of chill-stored blunt snout bream (Megalobrama amblycephala) fillets: Protein degradation and volatile organic compounds accumulation. Food Microbiol. 2020, 91, 103495. [Google Scholar] [CrossRef]
- Noda, H. Health benefits and nutritional properties of nori. J. Appl. Phycol. 1993, 5, 255–258. [Google Scholar] [CrossRef]
- Heaton, J.W.; Marangoni, A.G. Chlorophyll degradation in processed foods and senescent plant tissues. Trends Food Sci. Technol. 1996, 7, 8–15. [Google Scholar] [CrossRef]
- Lee, K.H.; Ryuk, J.H.; Jeong, I.H.; Jung, W.J. Quality changes of dried lavers during processing and storage 3. Changes in pigments, trypsin indigestible substrates (TIS) and dietary fiber content during roasting and storage. Korea. J. Fish. Aquat. Sci. 1990, 23, 280–288. [Google Scholar]
- Klotz, A.V.; Glazer, A.N. Characterization of the bilin attachment sites in R-phycoerythrin. J. Biol. Chem. 1985, 260, 4856–4863. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-S.; Noh, B.-S. Quality Evaluation of dried laver (Porphyra yenzoensis Ueda) using electronic nose based on metal oxide sensor or GC with SAW sensor during storage. Korean J. Food Sci. Technol. 2002, 34, 947–953. [Google Scholar]
- Bowler, C.; Montagu, M.V.; Inzé, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant. Physiol. Plant. Mol. Bio. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Altissimi, S.; Mercuri, M.L.; Framboas, M.; Tommasino, M.; Pelli, S.; Benedetti, F.; Bella, S.D.; Haouet, N. Indicators of protein spoilage in fresh and defrosted crustaceans and cephalopods stored in domestic condition. Italian J. Food Saf. 2018, 6, 6921. [Google Scholar] [CrossRef]
- Jeong, S.H.; Shin, S.B.; Oh, E.G.; Jo, M.R.; Yoon, M.C.; Lee, H.J.; Son, K.T. Sanitary evaluation for seawater and laver Pyropia sp. in the major laver growing areas, Korea. Korean J. Fish Aquat. Sci. 2017, 50, 343–351. [Google Scholar]
- Ye, K.; Ding, D.; Zhu, X.; Wu, Z.; Hu, Q.; Li, R. Modified atmosphere packaging with a small change in gas ratio cloud maintain pork quality during −3 °C storage. Food Control 2020, 109, 1069453. [Google Scholar] [CrossRef]
- Dumay, J.; Morançais, M. Proteins and pigments. In Seaweed in Health and Disease Prevention, 1st ed.; Fleurence, J., Levine, I., Eds.; N. Levy: San Diego, CA, USA, 2016; pp. 275–318. [Google Scholar]
- Ge, M.; Shen, J.; Liu, C.; Xia, W.; Xu, Y. Effect of acidification and thermal treatment on quality characteristics of high-moisture laver (Porphyra spp.). J. Food Process. Preserv. 2022, 46, e16762. [Google Scholar] [CrossRef]
- Isaka, S.; Cho, K.; Nakazono, S.; Abu, R.; Ueno, M.; Kim, D.; Oda, T. Antioxidant and anti-inflammatory activities of porphyran isolated from discolored nori (Porphyra yezoensis). Int. J. Biol. Macromol. 2015, 74, 68–75. [Google Scholar] [CrossRef]
- Takahashi, M.; Kumari, P.; Li, C.; Mikami, K. Low temperature causes discoloration by repressing growth and nitrogen transporter gene expression in the edible red alga Pyropia yezoensis. Mar. Environ. Res. 2020, 159, 105004. [Google Scholar] [CrossRef]
- Ronceros, B.; Díaz, D.; Miguel Bastías-Montes, J.; Muñoz-Fariña, O.; Lespinard, A.R.; Espinoza-Tellez, T.; Quevedo-León, R. Shelf life of barley shoots chlorophyll under four frozen-storage conditions. J. Food Process. Preserv. 2021, 45, e15972. [Google Scholar] [CrossRef]
- Kwartiningsih, E.; Ramadhani, A.N.; Putri, N.G.A.; Damara, V.C.J. Chlorophyll extraction methods review and chlorophyll stability of katuk leaves (Sauropus androgynous). J. Phys. Conf. Ser. 2021, 1858, 012015. [Google Scholar] [CrossRef]
- Pimentel, F.B.; Cermeño, M.; Kleekayai, T.; Harnedy, P.A.; FitzGerald, R.J.; Alves, R.C.; Oliveira, M. Effect of in vitro simulated gastrointestinal digestion on the antioxidant activity of the red seaweed Porphyra dioica. Food Res. Int. 2020, 136, 109309. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.G.; Jeong, M.C.; Park, S.K.; Lee, J.W.; Cho, J.H.; Eom, S.H.; Huh, M.K.; Kim, Y.M. Analysis of seasonal and regional changes in major food components of raw laver Pyropia sp. Korean J. Fish Aquat. Sci. 2018, 51, 510–517. [Google Scholar]
- Kawashima, T.; Shirai, T.; Matsuda, H.; Osako, K.; Okazaki, E. Identification and roles of the taste-active components of dried Nori. Jpn. J. Food Eng. 2018, 19, 121–127. [Google Scholar] [CrossRef]
- Xu, K.; Xu, Y.; Ji, D.; Xie, J.; Chen, C.; Xie, C. Proteomic analysis of the economic seaweed Pyropia haitanensis in response to desiccation. Algal Res. 2016, 19, 198–206. [Google Scholar] [CrossRef]
- Utrera, M.; Morcuende, D.; Estévez, M. Temperature of frozen storage affects the nature and consequences of protein oxidation in beef patties. Meat Sci. 2014, 96, 1250–1257. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Free Amino Acid Content (mg/g) | ||||||
|---|---|---|---|---|---|---|
| Free Amino Acid | Storage Method | Storage Period (Day) | ||||
| 0 | 3 | 6 | 9 | 15 | ||
| Tau | R | 12.37 ± 0.01 | 12.92 ± 0.24 | 12.57 ± 0.10 | 12.41 ± 0.23 | 10.24 ± 0.09 |
| SS | 12.96 ± 0.11 | 12.25 ± 0.10 | 12.54 ± 0.10 | 10.01 ± 0.09 | ||
| CS | 12.18 ± 0.20 | 12.23 ± 0.23 | 12.23 ± 0.21 | 10.90 ± 0.13 | ||
| F | 13.73 ± 0.20 | 13.31 ± 0.12 | 12.94 ± 0.20 | 13.06 ± 0.18 | ||
| Ala | R | 6.86 ± 0.01 | 8.44 ± 0.06 | 8.79 ± 0.02 | 8.59 ± 0.03 | 5.65 ± 0.02 |
| SS | 8.35 ± 0.05 | 8.51 ± 0.05 | 8.71 ± 0.05 | 4.72 ± 0.03 | ||
| CS | 7.40 ± 0.05 | 8.99 ± 0.08 | 9.01 ± 0.04 | 7.22 ± 0.06 | ||
| F | 8.60 ± 0.03 | 8.69 ± 0.06 | 8.45 ± 0.05 | 9.18 ± 0.02 | ||
| Glu | R | 3.73 ± 0.01 | 2.92 ± 0.01 | 2.74 ± 0.01 | 1.93 ± 0.01 | 0.34 ± 0.01 |
| SS | 3.31 ± 0.02 | 2.99 ± 0.02 | 1.74 ± 0.01 | 0.46 ± 0.01 | ||
| CS | 2.99 ± 0.02 | 3.49 ± 0.02 | 2.25 ± 0.02 | 1.02 ± 0.01 | ||
| F | 3.97 ± 0.01 | 3.17 ± 0.03 | 4.17 ± 0.02 | 3.23 ± 0.03 | ||
| Asp | R | 1.07 ± 0.01 | 0.90 ± 0.01 | 0.70 ± 0.01 | 0.49 ± 0.02 | 0.05 ± 0.01 |
| SS | 1.01 ± 0.01 | 0.78 ± 0.03 | 0.53 ± 0.01 | 0.12 ± 0.03 | ||
| CS | 0.84 ± 0.01 | 0.83 ± 0.04 | 0.62 ± 0.01 | 0.17 ± 0.03 | ||
| F | 0.77 ± 0.01 | 0.69 ± 0.01 | 0.67 ± 0.01 | 1.34 ± 0.07 | ||
| Cit | R | 0.54 | 0.81 ± 0.08 | 0.67 ± 0.04 | 0.36 ± 0.04 | 0.18 ± 0.02 |
| SS | 0.72 ± 0.08 | 0.73 ± 0.02 | 0.60 ± 0.04 | 0.05 | ||
| CS | 0.72 ± 0.04 | 0.71 ± 0.07 | 0.62 ± 0.03 | 0.27 ± 0.02 | ||
| F | 0.76 ± 0.06 | 0.57 ± 0.07 | 0.62 ± 0.03 | 0.81 ± 0.05 | ||
| Thr | R | 0.25 ± 0.01 | 0.48 ± 0.03 | 0.47 ± 0.03 | 0.36 ± 0.01 | 0.02 ± 0.01 |
| SS | 0.36 ± 0.03 | 0.34 ± 0.03 | 0.30 ± 0.02 | 0.12 ± 0.04 | ||
| CS | 0.30 ± 0.02 | 0.34 ± 0.03 | 0.28 ± 0.06 | 0.14 ± 0.02 | ||
| F | 0.32 ± 0.02 | 0.27 ± 0.04 | 0.29 ± 0.02 | 0.38 ± 0.06 | ||
| g-ABA | R | 0.25 ± 0.01 | 0.35 ± 0.04 | 0.41 ± 0.02 | 0.39 ± 0.03 | 0.10 ± 0.02 |
| SS | 0.40 ± 0.03 | 0.41 ± 0.05 | 0.40 ± 0.03 | 0.13 ± 0.06 | ||
| CS | 0.38 ± 0.03 | 0.37 ± 0.03 | 0.47 ± 0.02 | 0.29 ± 0.04 | ||
| F | 0.35 ± 0.03 | 0.29 ± 0.05 | 0.32 ± 0.02 | 0.39 ± 0.04 | ||
| Ser | R | 0.24 ± 0.01 | 0.18 ± 0.03 | 0.19 ± 0.03 | 0.09 ± 0.01 | 0.01 |
| SS | 0.16 ± 0.02 | 0.17 ± 0.02 | 0.11 ± 0.03 | 0.07 | ||
| CS | 0.14 ± 0.02 | 0.16 ± 0.03 | 0.11 ± 0.03 | 0.06 | ||
| F | 0.14 ± 0.02 | 0.11 ± 0.01 | 0.13 ± 0.02 | 0.15 ± 0.01 | ||
| Val | R | 0.22 ± 0.01 | 0.47 ± 0.05 | 0.47 ± 0.02 | 0.50 ± 0.04 | 1.15 ± 0.09 |
| SS | 0.39 ± 0.06 | 0.44 ± 0.04 | 0.39 ± 0.06 | 0.40 ± 0.02 | ||
| CS | 0.35 ± 0.08 | 0.46 ± 0.03 | 0.41 ± 0.02 | 0.34 ± 0.02 | ||
| F | 0.42 ± 0.02 | 0.33 ± 0.04 | 0.32 ± 0.07 | 0.50 ± 0.02 | ||
| Leu | R | 0.17 ± 0.01 | 0.50 ± 0.02 | 0.50 ± 0.01 | 0.48 ± 0.03 | 0.88 ± 0.01 |
| SS | 0.35 ± 0.01 | 0.41 ± 0.02 | 0.37 ± 0.01 | 0.32 ± 0.02 | ||
| CS | 0.31 ± 0.01 | 0.42 ± 0.01 | 0.34 ± 0.01 | 0.26 ± 0.02 | ||
| F | 0.35 ± 0.01 | 0.25 ± 0.02 | 0.26 ± 0.03 | 0.43 ± 0.01 | ||
| P—Ser | R | 0.17 ± 0.01 | 0.27 ± 0.02 | 0.32 ± 0.03 | 0.33 ± 0.02 | 0.88 ± 0.04 |
| SS | 0.20 ± 0.01 | 0.19 ± 0.01 | 0.23 ± 0.01 | 0.28 ± 0.01 | ||
| CS | 0.16 ± 0.02 | 0.20 ± 0.01 | 0.24 ± 0.01 | 0.31 ± 0.01 | ||
| F | 0.18 ± 0.01 | 0.20 ± 0.03 | 0.20 ± 0.01 | 0.21 ± 0.01 | ||
| b—Ala | R | 0.13 ± 0.01 | 0.14 ± 0.01 | 0.15 ± 0.01 | 0.15 ± 0.02 | 0.12 ± 0.01 |
| SS | 0.14 ± 0.01 | 0.14 ± 0.01 | 0.15 ± 0.01 | 0.10 ± 0.01 | ||
| CS | 0.14 ± 0.01 | 0.13 ± 0.01 | 0.14 ± 0.01 | 0.10 ± 0.01 | ||
| F | 0.13 ± 0.01 | 0.12 ± 0.01 | 0.13 ± 0.01 | 0.14 ± 0.01 | ||
| Ile | R | 0.11 ± 0.01 | 0.31 ± 0.03 | 0.30 ± 0.01 | 0.27 ± 0.01 | 0.47 ± 0.01 |
| SS | 0.22 ± 0.01 | 0.24 ± 0.02 | 0.21 ± 0.01 | 0.15 ± 0.01 | ||
| CS | 0.18 ± 0.01 | 0.25 ± 0.01 | 0.21 ± 0.01 | 0.16 ± 0.01 | ||
| F | 0.21 ± 0.02 | 0.15 ± 0.01 | 0.16 ± 0.01 | 0.25 ± 0.01 | ||
| Lys | R | 0.11 ± 0.01 | 0.30 ± 0.02 | 0.29 ± 0.01 | 0.29 ± 0.04 | 0.01 ± 0.01 |
| SS | 0.23 ± 0.01 | 0.27 ± 0.01 | 0.23 ± 0.01 | 0.29 ± 0.01 | ||
| CS | 0.20 ± 0.01 | 0.30 ± 0.02 | 0.24 ± 0.02 | 0.19 ± 0.02 | ||
| F | 0.24 ± 0.01 | 0.17 ± 0.01 | 0.17 ± 0.01 | 0.30 ± 0.03 | ||
| Tyr | R | 0.09 ± 0.01 | 0.18 ± 0.03 | 0.19 ± 0.02 | 0.19 ± 0.03 | 0.57 ± 0.04 |
| SS | 0.15 ± 0.01 | 0.18 ± 0.02 | 0.14 ± 0.01 | 0.12 ± 0.01 | ||
| CS | 0.13 ± 0.01 | 0.18 ± 0.02 | 0.14 ± 0.01 | 0.10 ± 0.01 | ||
| F | 0.17 ± 0.01 | 0.13 ± 0.01 | 0.13 ± 0.01 | 0.21 ± 0.02 | ||
| Gly | R | 0.09 ± 0.01 | 0.12 ± 0.01 | 0.19 ± 0.01 | 0.54 ± 0.05 | 1.15 ± 0.05 |
| SS | 0.14 ± 0.01 | 0.18 ± 0.01 | 0.32 ± 0.01 | 0.92 ± 0.03 | ||
| CS | 0.11 ± 0.01 | 0.19 ± 0.01 | 0.25 ± 0.01 | 0.44 ± 0.02 | ||
| F | 0.17 ± 0.01 | 0.13 ± 0.01 | 0.14 ± 0.01 | 0.16 ± 0.02 | ||
| Pro | R | 0.06 ± 0.01 | 0.21 ± 0.02 | 0.18 ± 0.03 | 0.14 ± 0.01 | ND |
| SS | 0.12 ± 0.01 | 0.17 ± 0.03 | 0.15 ± 0.02 | ND | ||
| CS | 0.10 ± 0.01 | 0.23 ± 0.04 | 0.12 ± 0.01 | ND | ||
| F | 0.19 ± 0.02 | 0.14 ± 0.01 | 0.11 ± 0.01 | 0.17 ± 0.04 | ||
| Phe | R | 0.06 ± 0.01 | 0.17 ± 0.01 | 0.15 ± 0.01 | 0.16 ± 0.01 | 0.53 ± 0.01 |
| SS | 0.13 ± 0.01 | 0.16 ± 0.01 | 0.13 ± 0.01 | 0.13 ± 0.01 | ||
| CS | 0.12 ± 0.01 | 0.17 ± 0.01 | 0.13 ± 0.01 | 0.14 ± 0.01 | ||
| F | 0.16 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.19 ± 0.01 | ||
| Arg | R | 0.04 ± 0.01 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.01 ± 0.01 |
| SS | 0.06 ± 0.01 | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.03 ± 0.01 | ||
| CS | 0.07 ± 0.01 | 0.09 ± 0.01 | 0.06 ± 0.01 | 0.15 ± 0.01 | ||
| F | 0.09 ± 0.01 | 0.10 ± 0.01 | 0.10 ± 0.02 | 0.09 ± 0.01 | ||
| NH3 | R | 0.03 ± 0.01 | 0.12 ± 0.01 | 0.70 ± 0.05 | 1.28 ± 0.05 | 2.80 ± 0.10 |
| SS | 0.04 ± 0.01 | 0.06 ± 0.01 | 0.15 ± 0.01 | 1.10 ± 0.03 | ||
| CS | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.07 ± 0.01 | 0.56 ± 0.01 | ||
| F | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.05 ± 0.01 | ||
| His | R | 0.03 ± 0.01 | 0.06 ± 0.02 | 0.06 ± 0.01 | 0.05 ± 0.01 | 0.02 ± 0.01 |
| SS | 0.05 ± 0.01 | 0.06 ± 0.01 | 0.04 ± 0.01 | 0.04 ± 0.01 | ||
| CS | 0.04 ± 0.01 | 0.06 ± 0.01 | 0.05 ± 0.01 | 0.03 ± 0.01 | ||
| F | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.04 ± 0.01 | 0.07 ± 0.01 | ||
| a—ABA | R | 0.02 ± 0.01 | 0.07 ± 0.01 | 0.08 ± 0.02 | 0.10 ± 0.02 | 0.59 ± 0.01 |
| SS | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.04 ± 0.01 | ND | ||
| CS | 0.04 ± 0.01 | 0.05 ± 0.01 | 0.04 ± 0.01 | ND | ||
| F | 0.04 ± 0.01 | 0.00 ± 0.01 | 0.04 ± 0.01 | 0.05 ± 0.01 | ||
| Orn | R | 0.01 | 0.07 ± 0.02 | 0.19 ± 0.01 | 0.27 ± 0.03 | 0.45 ± 0.04 |
| SS | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.27 ± 0.01 | ||
| CS | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.15 ± 0.01 | ||
| F | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | ||
| Met | R | 0.01 ± 0.01 | 0.01 | 0.01 | 0.01 | 0.02 |
| SS | 0.01 | 0.01 | 0.01 | 0.01 | ||
| CS | 0.01 | 0.02 | 0.01 | 0.04 | ||
| F | 0.02 | 0.02 | 0.02 | 0.01 | ||
| a—AAA | R | ND | ND | ND | ND | 0.08 ± 0.01 |
| SS | ND | ND | ND | 0.04 ± 0.01 | ||
| CS | ND | ND | ND | ND | ||
| F | ND | ND | ND | ND | ||
| Total content | R | 26.67 ± 0.26 | 30.05 ± 0.77 | 30.34 ± 1.12 | 29.40 ± 0.78 | 26.31 ± 0.55 |
| SS | 29.60 ± 1.23 | 28.81 ± 0.64 | 27.56 ± 0.66 | 19.87 ± 0.23 | ||
| CS | 26.98 ± 0.54 | 29.93 ± 0.40 | 28.07 ± 0.32 | 23.03 ± 0.43 | ||
| F | 31.14 ± 0.35 | 29.06 ± 0.22 | 29.57 ± 1.14 | 31.37 ± 1.01 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Park, D.H.; Kim, E.J.; Choi, M.-J. Freshness Analysis of Raw Laver (Pyropia yenzoensis) Conserved under Supercooling Conditions. Foods 2023, 12, 510. https://doi.org/10.3390/foods12030510
Lee H, Park DH, Kim EJ, Choi M-J. Freshness Analysis of Raw Laver (Pyropia yenzoensis) Conserved under Supercooling Conditions. Foods. 2023; 12(3):510. https://doi.org/10.3390/foods12030510
Chicago/Turabian StyleLee, Hyeonbo, Dong Hyeon Park, Eun Jeong Kim, and Mi-Jung Choi. 2023. "Freshness Analysis of Raw Laver (Pyropia yenzoensis) Conserved under Supercooling Conditions" Foods 12, no. 3: 510. https://doi.org/10.3390/foods12030510
APA StyleLee, H., Park, D. H., Kim, E. J., & Choi, M.-J. (2023). Freshness Analysis of Raw Laver (Pyropia yenzoensis) Conserved under Supercooling Conditions. Foods, 12(3), 510. https://doi.org/10.3390/foods12030510