
Pre-Treatment of Starter Cultures with Mild Pulsed Electric Fields Influences the Characteristics of Set Yogurt
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Pulsed Electric Field (PEF) Treatment
2.3. Yogurt Fermentation
2.4. Monitoring the pH Development of Yogurt
2.5. Microbiological Analysis
2.6. Syneresis and Texture Analysis
2.7. Proteolytic Activity
3. Results
3.1. Factorial Experiments
3.2. Effect of PEF Treatment on the Performance of the Starter Culture after a Freeze–Thaw Cycle
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosa, M.C.; Carmo, M.R.S.; Balthazar, C.F.; Guimarães, J.T.; Esmerino, E.A.; Freitas, M.Q.; Silva, M.C.; Pimentel, T.C.; Cruz, A.G. Dairy Products with Prebiotics: An Overview of the Health Benefits, Technological and Sensory Properties. Int. Dairy J. 2021, 117, 105009. [Google Scholar] [CrossRef]
- Arioli, S.; Della Scala, G.; Remagni, M.C.; Stuknyte, M.; Colombo, S.; Guglielmetti, S.; De Noni, I.; Ragg, E.; Mora, D. Streptococcus Thermophilus Urease Activity Boosts Lactobacillus Delbrueckii Subsp. Bulgaricus Homolactic Fermentation. Int. J. Food Microbiol. 2017, 247, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Siemer, C.; Aganovic, K.; Toepfl, S.; Heinz, V. Application of Pulsed Electric Fields in Food. In Conventional and Advanced Food Processing Technologies; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 645–672. [Google Scholar]
- Timmermans, R.; Nierop Groot, M.; Matser, A. Liquid Food Pasteurization by Pulsed Electric Fields. In Pulsed Electric Fields Technology for the Food Industry. Food Engineering Series; Raso, J., Heinz, V., Alvarez, I., Toepfl, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 299–323. [Google Scholar]
- Vorobiev, E.; Lebovka, N.I. Cell Membrane Permeabilization by Pulsed Electric Fields for Efficient Extraction of Intercellular Components from Foods. In Pulsed Electric Fields Technology for the Food Industry. Food Engineering Series; Raso, J., Heinz, V., Alvarez, I.T.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 209–269. [Google Scholar]
- Middendorf, D.; Bindrich, U.; Siemer, C.; Töpfl, S.; Heinz, V. Affecting Casein Micelles by Pulsed Electrical Field (PEF) for Inclusion of Lipophilic Organic Compounds. Appl. Sci. 2021, 11, 4611. [Google Scholar] [CrossRef]
- Ewe, J.A.; Wan-Abdullah, W.N.; Alias, A.K.; Liong, M.T. Enhanced Growth of Lactobacilli and Bioconversion of Isoflavones in Biotin-Supplemented Soymilk by Electroporation. Int. J. Food Sci. Nutr. 2012, 63, 580–596. [Google Scholar] [CrossRef]
- Fologea, D.; Vassu-Dimov, T.; Stoica, I.; Csutak, O.; Radu, M. Increase of Saccharomyces Cerevisiae Plating Efficiency after Treatment with Bipolar Electric Pulses. Bioelectrochemistry Bioenerg. 1998, 46, 285–287. [Google Scholar] [CrossRef]
- Vassu, T. Secondary Effects of Electroporation with Bipolar Electric Pulses: Electrostimulation. Period. Mineral. 2021, 90, 45–48. [Google Scholar]
- Yeo, S.K.; Liong, M.T. Effect of Electroporation on Viability and Bioconversion of Isoflavones in Mannitol-Soymilk Fermented by Lactobacilli and Bifidobacteria. J. Sci. Food Agric. 2013, 93, 396–409. [Google Scholar] [CrossRef]
- Najim, N.; Aryana, K.J. A Mild Pulsed Electric Field Condition That Improves Acid Tolerance, Growth, and Protease Activity of Lactobacillus Acidophilus LA-K and Lactobacillus Delbrueckii Subspecies Bulgaricus LB-12. J. Dairy Sci. 2013, 96, 3424–3434. [Google Scholar] [CrossRef]
- Ohba, T.; Uemura, K.; Nabetani, H. Moderate Pulsed Electric Field Treatment Enhances Exopolysaccharide Production by Lactococcus Lactis Subspecies Cremoris. Process Biochem. 2016, 51, 1120–1128. [Google Scholar] [CrossRef]
- Ohba, T.; Uemura, K.; Nabetani, H. Changes in Biosynthesis of Exopolysaccharide in Lactococcus Lactis Subspecies Cremoris Treated by Moderate Pulsed Electric Field Treatment. Biosci. Biotechnol. Biochem. 2017, 81, 724–734. [Google Scholar] [CrossRef][Green Version]
- Kanafusa, S.; Uhlig, E.; Uemura, K.; Gómez Galindo, F.; Håkansson, Å. The Effect of Nanosecond Pulsed Electric Field on the Production of Metabolites from Lactic Acid Bacteria in Fermented Watermelon Juice. Innov. Food Sci. Emerg. Technol. 2021, 72, 102749. [Google Scholar] [CrossRef]
- Mattar, J.R.; Turk, M.F.; Nonus, M.; Lebovka, N.I.; El Zakhem, H.; Vorobiev, E. Stimulation of Saccharomyces Cerevisiae Cultures by Pulsed Electric Fields. Food Bioproc. Tech. 2014, 7, 3328–3335. [Google Scholar] [CrossRef]
- Mattar, J.R.; Turk, M.F.; Nonus, M.; Lebovka, N.I.; El Zakhem, H.; Vorobiev, E.S. Cerevisiae Fermentation Activity after Moderate Pulsed Electric Field Pre-Treatments. Bioelectrochemistry 2015, 103, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Koubaa, M.; Bals, O.; Vorobiev, E. Effect of Pulsed Electric Fields on the Growth and Acidification Kinetics of Lactobacillus Delbrueckii Subsp. Bulgaricus. Foods 2020, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Raso, J.; Frey, W.; Ferrari, G.; Pataro, G.; Knorr, D.; Teissie, J.; Miklavčič, D. Recommendations Guidelines on the Key Information to Be Reported in Studies of Application of PEF Technology in Food and Biotechnological Processes. Innov. Food Sci. Emerg. Technol. 2016, 37, 312–321. [Google Scholar] [CrossRef]
- Meroth, C.B.; Hammes, W.P.; Hertel, C. Identification and Population Dynamics of Yeasts in Sourdough Fermentation Processes by PCR-Denaturing Gradient Gel Electrophoresis. Appl. Environ. Microbiol. 2003, 69, 7453–7461. [Google Scholar] [CrossRef]
- Bauer, S.A.W.; Schneider, S.; Behr, J.; Kulozik, U.; Foerst, P. Combined Influence of Fermentation and Drying Conditions on Survival and Metabolic Activity of Starter and Probiotic Cultures after Low-Temperature Vacuum Drying. J. Biotechnol. 2012, 159, 351–357. [Google Scholar] [CrossRef]
- Yuan, R.; Yang, B.; Liu, Y.; Huang, L. Modified Gompertz Sigmoidal Model Removing Fine-Ending of Grain-Size Distribution. Open Geosci. 2019, 11, 29–36. [Google Scholar] [CrossRef]
- Amani, E.; Eskandari, M.H.; Shekarforoush, S. The Effect of Proteolytic Activity of Starter Cultures on Technologically Important Properties of Yogurt. Food Sci. Nutr. 2017, 5, 525–537. [Google Scholar] [CrossRef]
- Matumoto-Pintro, P.T.; Rabiey, L.; Robitaille, G.; Britten, M. Use of Modified Whey Protein in Yoghurt Formulations. Int. Dairy J. 2011, 21, 21–26. [Google Scholar] [CrossRef]
- de Giori, G.S.; Hébert, E.M. Methods to Determine Proteolytic Activity of Lactic Acid Bacteria. In Food Microbiology Protocols; Spencer, J.F., Ragout de Spencer, A.L., Eds.; Humana Press: Totowa, NJ, USA, 2001; pp. 197–202. ISBN 0-89603-867-X. [Google Scholar]
- Chanos, P.; Warncke, M.C.; Ehrmann, M.A.; Hertel, C. Application of Mild Pulsed Electric Fields on Starter Culture Accelerates Yogurt Fermentation. Eur. Food Res. Technol. 2020, 246, 621–630. [Google Scholar] [CrossRef]
- Torriani, S.; Gardini, F.; Guerzoni, M.E.; Dellaglio, F. Use of Response Surface Methodology to Evaluate Some Variables Affecting the Growth and Acidification Characteristics of Yoghurt Cultures. Int. Dairy J. 1996, 6, 625–636. [Google Scholar] [CrossRef]
- Loghavi, L.; Sastry, S.K.; Yousef, A.E. Effect of Moderate Electric Field Frequency and Growth Stage on the Cell Membrane Permeability of Lactobacillus acidophilus. Biotechnol. Prog. 2009, 25, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Tryfona, T.; Bustard, M.T. Impact of Pulsed Electric Fields on Corynebacterium Glutamicum Cell Membrane Permeabilization. J. Biosci. Bioeng. 2008, 105, 375–382. [Google Scholar] [CrossRef]
- Hunt, R.W.; Zavalin, A.; Bhatnagar, A.; Chinnasamy, S.; Das, K.C. Electromagnetic Biostimulation of Living Cultures for Biotechnology, Biofuel and Bioenergy Applications. Int. J. Mol. Sci. 2009, 10, 4515–4558. [Google Scholar] [CrossRef] [PubMed]
- Loghavi, L.; Sastry, S.K.; Yousef, A.E. Effect of Moderate Electric Field Frequency on Growth Kinetics and Metabolic Activity of Lactobacillus Acidophilus. Biotechnol. Prog. 2008, 24, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Pakhomova, O.N.; Khorokhorina, V.A.; Bowman, A.M.; Rodaite, R. Oxidative effects of nanosecond pulsed electric field exposure in cells and cell-free media. Arch. Biochem. Biophys. 2012, 527, 55–64. [Google Scholar] [CrossRef]
- Teraguchi, S.; Ono, J.; Kiyosawa, I.; Okonogi, S. Oxygen Uptake Activity and Aerobic Metabolism of Streptococcus Thermophilus STH450. J. Dairy Sci. 1987, 70, 514–523. [Google Scholar] [CrossRef]
- Sasaki, Y.; Horiuchi, H.; Kawashima, H.; Mukai, T.; Yamamoto, Y. NADH Oxidase of Streptococcus Thermophilus 1131 Is Required for the Effective Yogurt Fermentation with Lactobacillus Delbrueckii Subsp. Bulgaricus 2038. Biosci. Microbiota Food Health 2014, 33, 31–40. [Google Scholar] [CrossRef]
- Marty-Teysset, C.; de la Torre, F.; Garel, J.-R. Increased Production of Hydrogen Peroxide by Lactobacillus Delbrueckii Subsp. Bulgaricus upon Aeration: Involvement of an NADH Oxidase in Oxidative Stress. Appl. Environ. Microbiol. 2000, 66, 262–267. [Google Scholar] [CrossRef]
- Tiwari, S.; Kavitake, D.; Devi, P.B.; Halady Shetty, P. Bacterial Exopolysaccharides for Improvement of Technological, Functional and Rheological Properties of Yoghurt. Int. J. Biol. Macromol. 2021, 183, 1585–1595. [Google Scholar] [CrossRef] [PubMed]
- Podolska, V.I.; Voitenko, O.Y.; Ulberg, Z.R.; Yakubenko, L.M.; Grishchenko, N.I.; Ermakov, V.N. Influence of Pulse Electric Field on the Surface Properties of Lactic Acid Bacteria Lactobacillus Plantarum and Biogenic Formation of Ultradisperse Silver. Him. Fiz. Tehnol. Poverhni 2017, 8, 143–154. [Google Scholar] [CrossRef]
- Nguyen, P.T.; Nguyen, T.T.; Bui, D.C.; Hong, P.T.; Hoang, Q.K.; Nguyen, H.T. Exopolysaccharide Production by Lactic Acid Bacteria: The Manipulation of Environmental Stresses for Industrial Applications. AIMS Microbiol. 2020, 6, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, C.; Li, D.; Zhao, Y.; Zhang, X.; Zeng, X.; Yang, Z.; Li, S. Antioxidant Activity of an Exopolysaccharide Isolated from Lactobacillus Plantarum C88. Int. J. Biol. Macromol. 2013, 54, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Wang, B.H.; Xu, X.; der Meister, T.; Tabač, H.T.; Hwang, F.F.; Liu, Z. Extrusion of Dissolved Oxygen by Exopolysaccharide from Leuconostoc Mesenteroides and Its Implications in Relief of the Oxygen Stress. Front. Microbiol. 2018, 9, 2467. [Google Scholar] [CrossRef]
- Jang, I.A.; Kim, J.; Park, W. Endogenous Hydrogen Peroxide Increases Biofilm Formation by Inducing Exopolysaccharide Production in Acinetobacter Oleivorans DR1. Sci. Rep. 2016, 6, 21121. [Google Scholar] [CrossRef]
- Li, N.; Wang, Y.; Zhu, P.; Liu, Z.; Guo, B.; Ren, J. Improvement of Exopolysaccharide Production in Lactobacillus Casei LC2W by Overexpression of NADH Oxidase Gene. Microbiol. Res. 2015, 171, 73–77. [Google Scholar] [CrossRef]
- López-Gámez, G.; Elez-Martínez, P.; Martín-Belloso, O.; Soliva-Fortuny, R. Pulsed Electric Fields Affect Endogenous Enzyme Activities, Respiration and Biosynthesis of Phenolic Compounds in Carrots. Postharvest. Biol. Technol. 2020, 168, 111284. [Google Scholar] [CrossRef]
- Góral, M.; Pankiewicz, U. Effect of Pulsed Electric Fields (PEF) on Accumulation of Magnesium in Lactobacillus Rhamnosus B 442 Cells. J. Membr. Biol. 2017, 250, 565–572. [Google Scholar] [CrossRef]
- Boyaval, P. Lactic Acid Bacteria and Metal Ions. Lait 1989, 69, 87–113. [Google Scholar] [CrossRef]
- Montel, M.-C.; Seronie, M.-P.; Gine Talon, R.É.; Braud, M.H. Purification and Characterization of a Dipeptidase from Lactobacillus Sake. Appl. Environ. Microbiol. 1995, 61, 837–839. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, T.; Tamura, T.; Sato, M. Influence of Pulsed Electric Field on Various Enzyme Activities. J. Electrostat. 2007, 65, 156–161. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, Z.; Gao, Y.; Bao, Z.; Sun, N.; Lin, S. Mechanism of Trypsin Activation by Pulsed Electric Field Treatment Revealed Based on Chemical Experiments and Molecular Dynamics Simulations. Food Chem. 2022, 394, 133477. [Google Scholar] [CrossRef] [PubMed]
- Erkmen, O.; Bozoglu, T.F. (Eds.) Fermented Dairy Products. In Food Microbiology: Principles into Practice; Wiley: Hoboken, NJ, USA, 2016; pp. 253–287. [Google Scholar]
- Kesenkas, H.; Dinkci, N.; Seckin, K.; Gursoy, O.; Kinik, O. Physicochemical, Biochemical, Textural and Sensory Properties of Telli Cheese—A Traditional Turkish Cheese Made from Cow Milk. Bulg. J. Agric. Sci. 2012, 18, 763–770. [Google Scholar]
- Ruas-Madiedo, P.; Alting, A.C.; Zoon, P. Effect of Exopolysaccharides and Proteolytic Activity of Lactococcus Lactis Subsp. Cremoris Strains on the Viscosity and Structure of Fermented Milks. Int. Dairy J. 2005, 15, 155–164. [Google Scholar] [CrossRef]
- Li, C.; Song, J.; Kwok, L.; Wang, J.; Dong, Y.; Yu, H.; Hou, Q.; Zhang, H.; Chen, Y. Influence of Lactobacillus Plantarum on Yogurt Fermentation Properties and Subsequent Changes during Postfermentation Storage. J. Dairy Sci. 2017, 100, 2512–2525. [Google Scholar] [CrossRef]
- Yang, H.; He, M.; Wu, C. Cross Protection of Lactic Acid Bacteria during Environmental Stresses: Stress Responses and Underlying Mechanisms. LWT 2021, 144, 111203. [Google Scholar] [CrossRef]
- Rangel, D.E.N. Stress Induced Cross-Protection against Environmental Challenges on Prokaryotic and Eukaryotic Microbes. World J. Microbiol. Biotechnol. 2011, 27, 1281–1296. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
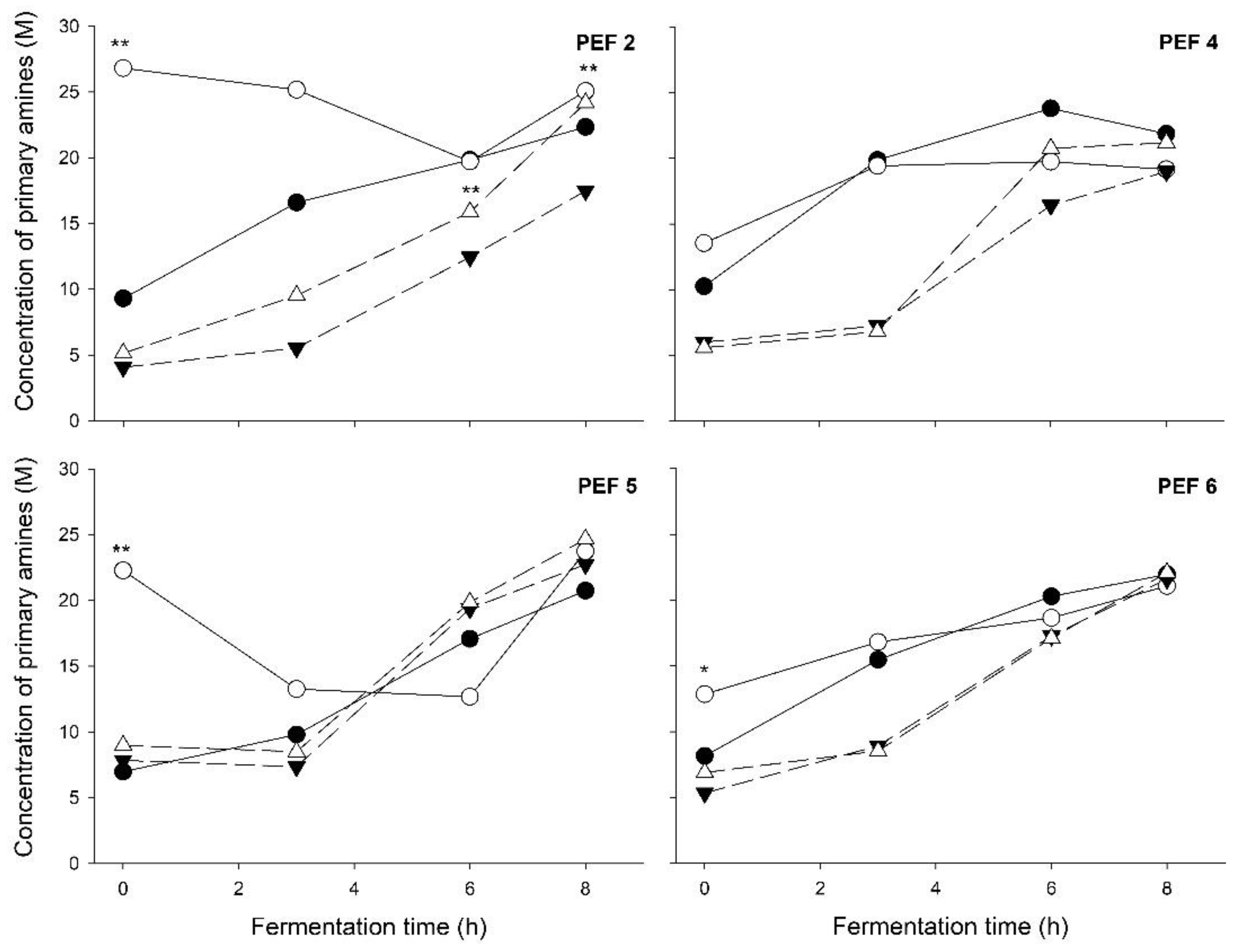{kind=link}
| PEF Treatment | PEF Factors | Mean λpH ± SD (min) | Mean µmax ± SD (pH/min) | Log10 Cell Counts (CFU/g) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S. thermophilus ± SD | L. bulgaricus ± SD | |||||||||
| Voltage (V) | Pulse Number (n) | Frequency (Hz) | Pulse Width (µs) | SC | NI | SC | NI | |||
| 1 | −1 | −1 | −1 | −1 | 78.66 ± 22.26 | 0.0128 ± 0.0008 | 8.20 ± 8.01 | 8.02 ± 7.76 | 8.22 ± 8.06 | 8.11 ± 7.93 |
| 2 | −1 | −1 | −1 | 1 | 73.53 ± 13.47 | 0.0143 ± 0.0018 | 7.79 ± 7.68 | 7.68 ± 7.73 | 8.14 ± 7.97 | 8.12 ± 7.98 |
| 3 | −1 | −1 | 1 | −1 | 64.36 ± 21.37 | 0.0144 ± 0.0017 | 8.13 ± 7.88 | 8.02 ± 7.78 | 8.19 ± 7.99 | 8.11 ± 8.00 |
| 4 | −1 | −1 | 1 | 1 | 65.68 ± 11.98 | 0.0152 ± 0.0004 | 8.26 ± 8.10 | 8.03 ± 7.73 | 8.25 ± 8.09 | 8.20 ± 8.10 |
| 5 | −1 | 1 | −1 | −1 | 58.40 ± 8.31 | 0.0140 ± 0.0015 | 8.65 ± 8.60 | 8.11. ± 7.99 | 8.28 ± 8.02 | 8.10 ± 7.92 |
| 6 | −1 | 1 | −1 | 1 | 72.92 ± 9.10 | 0.0144 ± 0.0015 | 8.45 ± 8.36 | 7.96 ± 7.84 | 8.11 ± 8.05 | 8.08 ± 8.03 |
| 7 | −1 | 1 | 1 | −1 | 68.94 ± 12.28 | 0.0135 ± 0.0012 | 8.48 ± 8.50 | 7.83 ± 7.73 | 7.91 ± 7.84 | 7.94 ± 7.87 |
| 8 | −1 | 1 | 1 | 1 | 69.43 ± 1.48 | 0.0139 ± 0.0009 | 8.48 ± 8.46 | 7.97 ± 7.86 | 8.71 ± 8.77 | 7.94 ± 7.63 |
| 9 | 1 | −1 | −1 | −1 | 66.77 ± 11.96 | 0.0139 ± 0.0021 | 8.26 ± 8.08 | 8.03 ± 7.85 | 8.26 ± 8.10 | 8.21 ± 8.08 |
| 10 | 1 | −1 | −1 | 1 | 72.08 ± 6.83 | 0.0126 ± 0.0009 | 8.93 ± 9.00 | 7.99 ± 7.92 | 8.80 ± 8.85 | 8.10 ± 8.07 |
| 11 | 1 | −1 | 1 | −1 | 82.92 ± 17.89 | 0.0129 ± 0.0005 | 8.87 ± 8.92 | 7.96 ± 7.81 | 8.92 ± 9.02 | 7.98 ± 7.75 |
| 12 | 1 | −1 | 1 | 1 | 71.60 ± 15.35 | 0.0132 ± 0.0022 | 8.21 ± 7.97 | 8.02 ± 7.76 | 8.23 ± 8.07 | 8.09 ± 7.96 |
| 13 | 1 | 1 | −1 | −1 | 68.29 ± 18.14 | 0.0158 ± 0.0027 | 8.93 ± 9.00 | 7.88 ± 7.78 | 8.15 ± 8.12 | 8.10 ± 8.07 |
| 14 | 1 | 1 | −1 | 1 | 78.00 ± 9.18 | 0.0126 ± 0.0013 | 8.71 ± 8.74 | 8.05 ± 7.99 | 8.73 ± 8.76 | 8.07 ± 8.04 |
| 15 | 1 | 1 | 1 | −1 | 70.66 ± 23.57 | 0.0135 ± 0.0017 | 8.45 ± 8.30 | 8.13 ± 8.03 | 8.21 ± 8.09 | 8.01 ± 7.85 |
| 16 | 1 | 1 | 1 | 1 | 65.61 ± 8.66 | 0.0127 ± 0.0008 | 8.57 ± 8.58 | 7.79 ± 7.46 | 7.96 ± 7.79 | 7.91 ± 7.83 |
| Center Point | 0 | 0 | 0 | 0 | 70.54 ± 14.29 | 0.0134 ± 0.0015 | 8.19 ± 8.16 | 7.90 ± 7.87 | 8.15 ± 8.11 | 8.27 ± 8.27 |
| PEF Treatment | Syneresis (% Serum Released ± SD) | Texture Analysis Force (N ± SD) | Proteolysis Concentration in Primary Amines (M ± SD) | ||
|---|---|---|---|---|---|
| t = 3 h | t = 6 h | t = 8 h | |||
| 1 | 44.57 ± 7.54 | 0.85 ± 0.48 | 20.66 ± 18.14 | 19.18 ± 12.66 | 18.62 ± 11.68 |
| 2 | 41.35 ± 7.36 | 1.37 ± 0.25 | 13.64 ± 11.47 | 13.00 ± 9.41 | 15.02 ± 6.27 |
| 3 | 41.80 ± 11.87 | 0.81 ± 0.38 | 26.00 ± 19.95 | 20.60 ± 15.55 | 22.52 ± 7.05 |
| 4 | 42.32 ± 9.82 | 0.83 ± 0.45 | 28.30 ± 15.66 | 26.78 ± 16.13 | 29.32 ± 16.27 |
| 5 | 46.70 ± 18.30 | 1.48 ± 0.44 | 20.38 ± 9.59 | 23.70 ± 8.10 | 22.42 ± 16.36 |
| 6 | 38.93 ± 9.92 | 1.26 ± 0.06 | 13.60 ± 11.58 | 21.94 ± 7.15 | 14.62 ± 10.62 |
| 7 | 47.73 ± 16.55 | 1.02 ± 0.65 | 13.84 ± 13.86 | 11.16 ± 6.26 | 13.86 ± 9.41 |
| 8 | 43.58 ± 12.32 | 1.14 ± 0.06 | 39.86 ± 9.07 | 21.68 ± 14.99 | 26.42 ± 12.00 |
| 9 | 43.87 ± 6.63 | 1.21 ± 0.57 | 40.12 ± 12.37 | 29.44 ± 17.96 | 27.56 ± 13.19 |
| 10 | 39.39 ± 10.38 | 0.93 ± 0.18 | 32.06 ± 13.37 | 23.38 ± 12.99 | 45.26 ± 12.06 |
| 11 | 40.42 ± 15.86 | 1.50 ± 0.44 | 24.18 ± 20.51 | 22.08 ± 17.79 | 20.74 ± 15.43 |
| 12 | 49.40 ± 20.20 | 1.26 ± 0.36 | 23.92 ± 21.30 | 18.54 ± 13.83 | 20.82 ± 16.66 |
| 13 | 44.03 ± 9.11 | 1.05 ± 0.39 | 22.98 ± 17.23 | 20.86 ± 8.50 | 30.82 ± 11.67 |
| 14 | 46.55 ± 12.25 | 1.08 ± 0.30 | 30.56 ± 3.35 | 24.92 ± 10.62 | 36.32 ± 10.80 |
| 15 | 42.82 ± 11.68 | 1.12 ± 0.05 | 15.36 ± 16.80 | 22.04 ± 18.11 | 18.33 ± 16.23 |
| 16 | 36.78 ± 9.01 | 1.42 ± 0.47 | 21.88 ± 13.34 | 25.22 ± 17.61 | 26.32 ± 18.43 |
| Center Point | 43.67 ± 10.57 | 1.05 ± 0.39 | 23.26 ± 16.39 | 22.07 ± 14.43 | 25.61 ± 17.54 |
| Starter Culture Treatment | Mean λpH ± SD (min) | Mean µmax ± SD (pH/h) | Log10 CFU/g S. thermophilus ± SD | Log10 CFU/g L. bulgaricus ± SD | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| t = 0 h | t = 3 h | t = 6 h | t = 8 h | t = 0 h | t = 3 h | t = 6 h | t = 8 h | |||
| PEF 5 | ||||||||||
| control | 94 ± 18.00 | 0.546 ± 0.025 | 5.98 ± 0.18 | 8.42 ± 0.17 | 8.59 ± 0.23 | 8.89 ± 0.16 | 5.91 ± 0.10 | 8.01 ± 0.29 | 8.49 ± 0.07 | 8.77 ± 0.19 |
| PEF-treated | 73 ± 11.00 ** | 0.597 ± 0.052 | 5.88 ± 0.12 | 8.38 ± 0.15 | 8.43 ± 0.08 | 8.86 ± 0.11 | 5.78 ± 0.09 *** | 7.99 ± 0.37 | 8.36 ± 0.18 | 8.69 ± 0.24 |
| control + frozen | 188 ± 22.80 | 0.628 ± 0.140 | 5.04 ± 0.14 | 5.89 ± 0.36 | 8.28 ± 0.29 | 8.92 ± 0.06 | 5.02 ± 0.16 | 6.99 ± 0.26 | 8.45 ± 0.03 | 8.98 ± 0.21 |
| PEF-treated + frozen | 202 ± 30.60 | 0.915 ± 0.082 | 5.2 ± 0.17 *** | 5.92 ± 0.39 | 8.32 ± 0.22 | 8.94 ± 0.14 | 5.03 ± 0.23 | 6.93 ± 0.31 | 8.43 ± 0.03 | 8.86 ± 0.03 |
| PEF 2 | ||||||||||
| control | 115 ± 23.90 | 0.536 ± 0.044 | 6.07 ± 0.12 | 7.72 ± 0.89 | 8.18 ± 0.11 | 8.83 ± 0.38 | 5.38 ± 0.75 | 8.23 ± 0.10 | 8.28 ± 0.14 | 8.52 ± 0.00 |
| PEF-treated | 109 ± 13.80 | 0.702 ± 0.101 | 5.88 ± 0.32 | 8.02 ± 0.10 | 8.19 ± 0.17 | 8.69 ± 0.41 | 5.33 ± 0.65 | 8.23 ± 0.06 | 8.15 ± 0.12 | 8.54 ± 0.44 |
| control + frozen | 185 (162–251) † | 0.703 ± 0.235 | 5.01 ± 0.06 | 6.51 ± 0.62 | 8.20 ± 0.38 | 8.60 ± 0.34 | 4.75 ± 0.26 | 7.08 ± 0.56 | 8.58 ± 0.09 | 9.08 ± 0.33 |
| PEF-treated + frozen | 196 (174–214) † | 0.718 ± 0.213 | 5.01 ± 0.06 | 6.79 ± 0.04 | 8.18 ± 0.36 | 8.74 ± 0.56 | 4.79 ± 0.22 | 7.20 ± 0.43 | 8.47 ± 0.35 | 9.10 ± 0.21 |
| PEF 4 | ||||||||||
| control | 95 ± 15.50 | 0.386 ± 0.031 | 6.06 ± 0.10 | 8.17 ± 0.33 | 8.35 ± 0.34 | 8.93 ± 0.16 | 6.09 ± 0.18 | 8.47 ± 0.32 | 8.73 ± 0.39 | 9.03 ± 0.13 |
| PEF-treated | 106 ± 6.70 | 0.447 ± 0.036 | 6.04 ± 0.10 | 8.18 ± 0.21 | 8.49 ± 0.22 | 8.91 ± 0.20 | 5.99 ± 0.06 | 8.47 ± 0.22 | 7.72 ± 1.56 | 8.81 ± 0.22 |
| control + frozen | 203 ± 6.90 | 0.601 ± 0.194 | 5.03 ± 0.11 | 6.29 ± 0.47 | 8.67 ± 0.32 | 9.06 ± 0.33 | 5.05 ± 0.14 | 6.87 ± 0.14 | 8.38 ± 0.57 | 9.06 ± 0.20 |
| PEF-treated + frozen | 182 ± 28.70 | 0.637 ± 0.142 | 4.93 ± 0.05 | 6.31 ± 0.52 | 8.54 ± 0.41 | 8.87 ± 0.18 | 5.01 ± 0.15 | 6.90 ± 0.19 | 8.06 ± 0.10 | 8.91 ± 0.08 |
| PEF 6 | ||||||||||
| control | 70 ± 9.97 | 0.544 ± 0.031 | 6.14 ± 0.16 | 7.99 ± 0.34 | 8.15 ± 0.23 | 8.83 ± 0.27 | 7.47 ± 1.27 | 8.27 ± 0.21 | 8.43 ± 0.26 | 9.01 ± 0.18 |
| PEF-treated | 69 ± 8.40 | 0.583 ± 0.055 | 6.10 ± 0.14 | 7.90 ± 0.29 | 8.12 ± 0.22 | 8.86 ± 0.31 | 6.05 ± 0.15 | 8.12 ± 0.13 | 8.47 ± 0.27 | 8.66 ± 0.42 |
| control + frozen | 146 ± 10.82 | 0.583 ± 0.139 | 5.12 ± 0.08 | 6.29 ± 0.18 | 8.17 ± 0.19 | 9.10 ± 0.18 | 5.13 ± 0.16 | 6.73 ± 0.24 | 8.46 ± 0.32 | 9.11 ± 0.21 |
| PEF-treated + frozen | 135 ± 6.38 * | 0.614 ± 0.162 | 5.02 ± 0.07 | 6.27 ± 0.25 | 8.21 ± 0.10 | 8.96 ± 0.16 | 5.03 ± 0.12 | 6.76 ± 0.23 | 8.49 ± 0.33 | 9.13 ± 0.19 |
| Starter Culture Treatment | Syneresis (% Serum Released ± SD) | Texture Analysis Force (N ± SD) | Proteolysis Concentration in Primary Amines (M ± SD) | |||
|---|---|---|---|---|---|---|
| t = 0 h | t = 3 h | t = 6 h | t = 8 h | |||
| PEF 5 | ||||||
| control | 47.57 ± 0.40 | 0.65 ± 0.19 | 6.98 ± 3.12 | 9.80 ± 7.61 | 17.08 ± 2.00 | 20.74 ± 3.70 |
| PEF-treated | 46.05 ± 0.91 | 1.05 ± 0.11 ** | 22.28 ± 5.35 ** | 13.26 ± 9.24 | 12.68 ± 8.60 | 23.74 ± 3.96 |
| control + frozen | 47.70 ± 0.41 | 0.57 ± 0.20 | 7.84 ± 5.16 | 7.36 ± 0.78 | 19.37 ± 2.19 | 22.74 ± 2.64 |
| PEF-treated + frozen | 45.15 ± 0.66 * | 0.68 ± 0.11 | 8.98 ± 4.43 | 8.50 ± 0.81 | 19.86 ± 2.97 | 24.66 ± 3.19 |
| PEF 2 | ||||||
| control | 46.21 ± 1.13 | 0.77 ± 0.22 | 9.30 ± 4.58 | 16.60 ± 6.04 | 19.84 ± 9.74 | 22.34 ± 5.14 |
| PEF-treated | 42.35 ± 0.68 ** | 1.20 ± 0.16 | 26.82 ± 4.88 * | 25.18 ± 12.61 | 19.72 ± 8.35 | 25.08 ± 5.87 ** |
| control + frozen | 44.52 ± 1.31 | 0.66 ± 0.03 | 4.08 ± 1.37 | 5.54 ± 2.71 | 12.46 ± 1.91 | 17.50 ± 5.23 |
| PEF-treated + frozen | 43.29 ± 0.71 | 0.82 ± 0.07 ** | 5.14 ± 1.08 | 9.54 ± 4.34 | 15.88 ± 0.92 ** | 24.18 ± 5.16 |
| PEF 4 | ||||||
| control | 50.48 ± 2.77 | 0.82 ± 0.10 | 10.26 ± 5.23 | 19.87 ± 11.13 | 23.79 ± 6.68 | 21.86 ± 5.24 |
| PEF-treated | 40.54 ± 0.95 | 1.25 ± 0.04 ** | 13.54 ± 3.33 | 19.42 ± 3.00 | 19.74 ± 0.57 | 19.16 ± 1.98 |
| control + frozen | 48.69 ± 1.09 | 0.66 ± 0.13 | 6.02 ± 0.52 | 7.26 ± 0.42 | 16.42 ± 2.34 | 19.00 ± 1.37 |
| PEF-treated + frozen | 41.55 ± 0.70 ** | 0.86 ± 0.10 ** | 5.60 ± 1.47 | 6.82 ± 0.71 | 20.76 ± 1.93 | 21.18 ± 3.58 |
| PEF 6 | ||||||
| control | 46.58 ± 0.70 | 0.82 ± 0.80 † | 8.14 ± 1.38 | 15.46 ± 3.17 | 20.28 ± 6.41 † | 21.94 ± 3.55 |
| PEF-treated | 41.10 ± 1.54 | 1.06 ± 0.07 † | 12.84 ± 2.59 * | 16.8 ± 1.02 | 18.64 ± 2.64 † | 21.06 ± 4.67 |
| control + frozen | 46.79 ± 0.57 | 0.56 ± 0.06 | 5.32 ± 1.43 | 8.90 ± 1.18 | 17.26 ± 1.27 | 21.60 ± 1.75 |
| PEF-treated + frozen | 41.11 ± 0.47 ** | 0.73 ± 0.11 | 6.88 ± 1.04 | 8.52 ± 0.44 | 17.10 ± 1.48 | 22.08 ± 1.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stühmeier-Niehe, C.; Lass, L.; Brocksieper, M.; Chanos, P.; Hertel, C. Pre-Treatment of Starter Cultures with Mild Pulsed Electric Fields Influences the Characteristics of Set Yogurt. Foods 2023, 12, 442. https://doi.org/10.3390/foods12030442
Stühmeier-Niehe C, Lass L, Brocksieper M, Chanos P, Hertel C. Pre-Treatment of Starter Cultures with Mild Pulsed Electric Fields Influences the Characteristics of Set Yogurt. Foods. 2023; 12(3):442. https://doi.org/10.3390/foods12030442
Chicago/Turabian StyleStühmeier-Niehe, Corinna, Luca Lass, Miriam Brocksieper, Panagiotis Chanos, and Christian Hertel. 2023. "Pre-Treatment of Starter Cultures with Mild Pulsed Electric Fields Influences the Characteristics of Set Yogurt" Foods 12, no. 3: 442. https://doi.org/10.3390/foods12030442
APA StyleStühmeier-Niehe, C., Lass, L., Brocksieper, M., Chanos, P., & Hertel, C. (2023). Pre-Treatment of Starter Cultures with Mild Pulsed Electric Fields Influences the Characteristics of Set Yogurt. Foods, 12(3), 442. https://doi.org/10.3390/foods12030442