

Antarctic Soil Yeasts with Fermentative Capacity and Potential for the Wine Industry

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
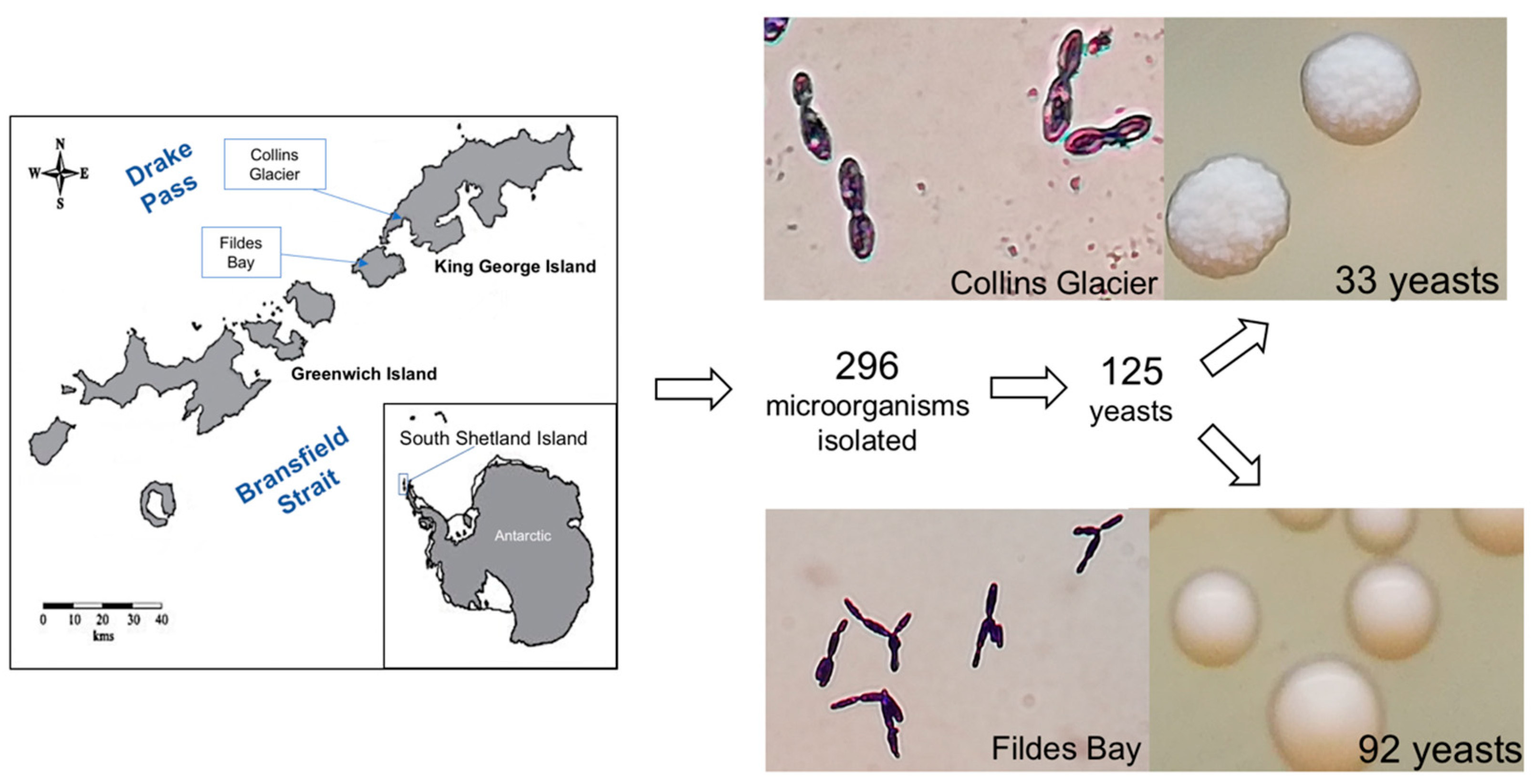
2.1. Yeast Isolation
2.2. Macroscopic and Microscopic Analyses
2.3. Fermentative Capacity Test
2.4. Sugar and Alcohol Tolerance Test
2.5. DNA Isolation
2.6. Molecular Characterization of Antarctic Fermentative Yeasts
2.7. Optimal Growth Temperature
3. Results
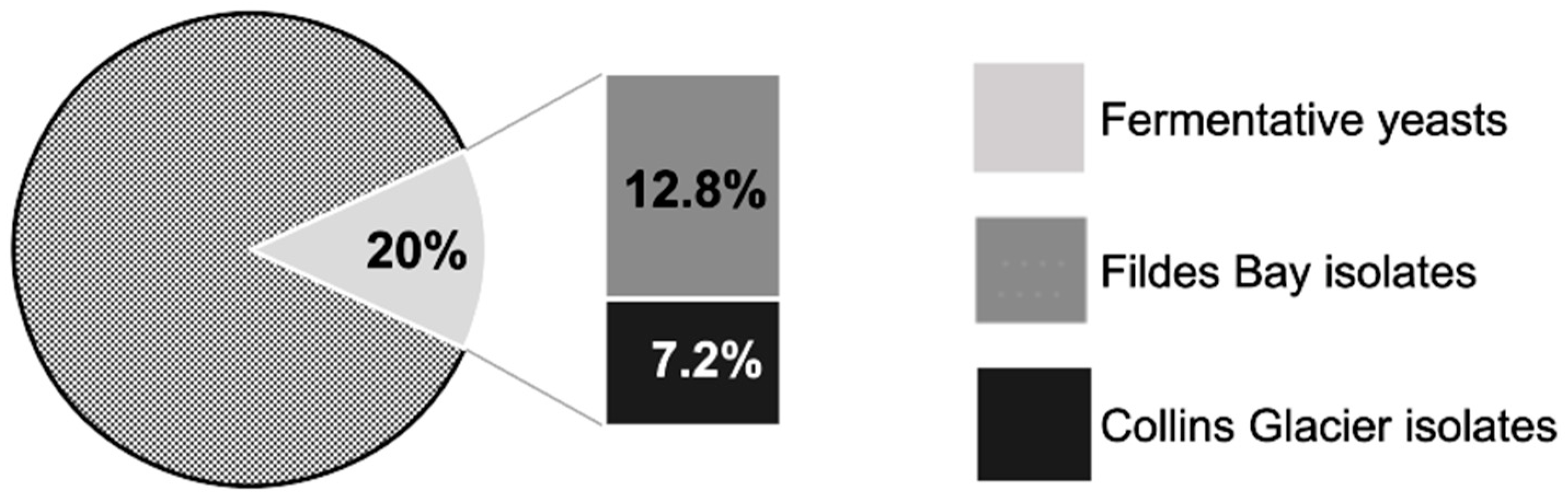
3.1. Yeast Isolation
3.2. Fermentative Capacity Test
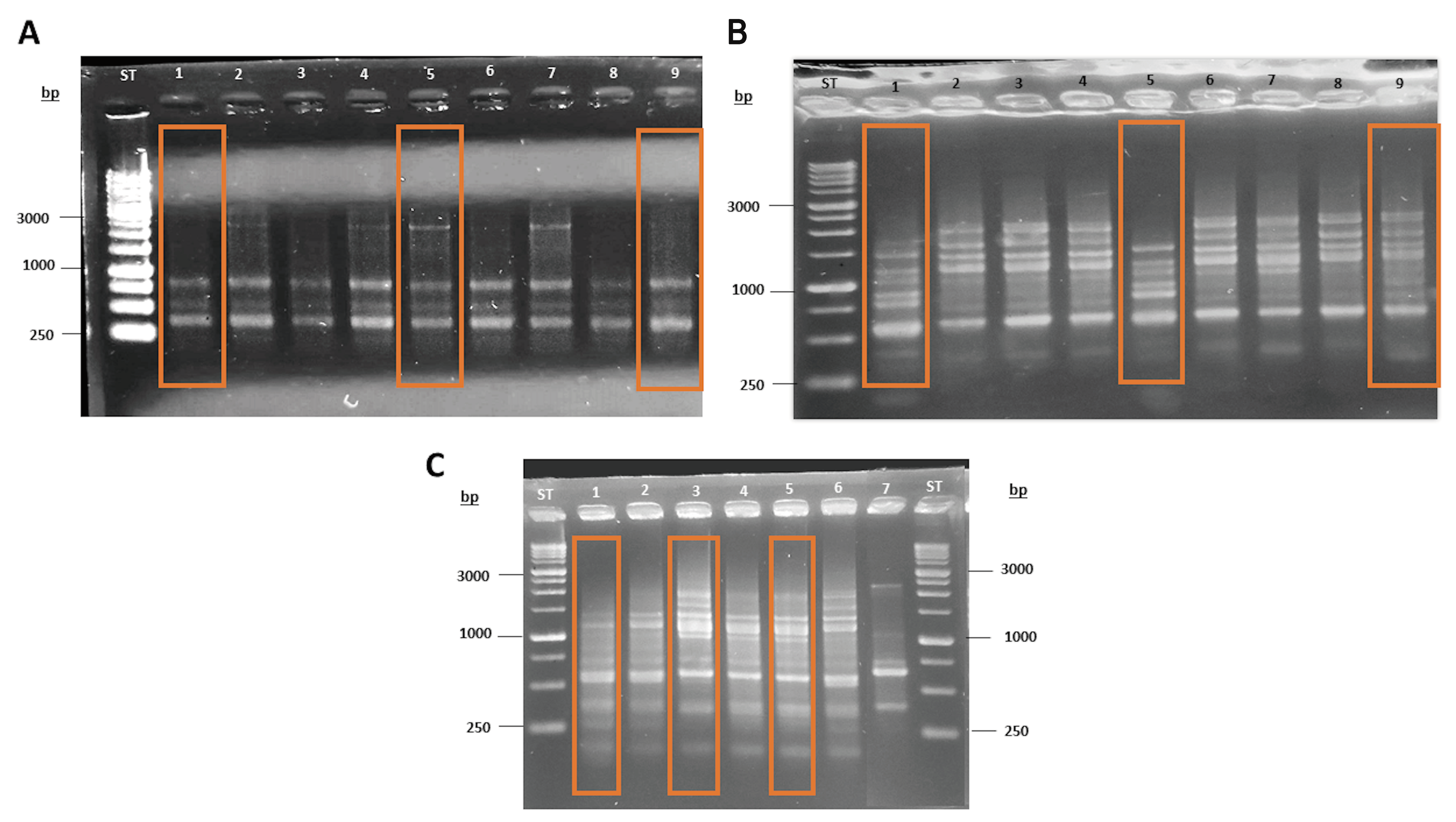
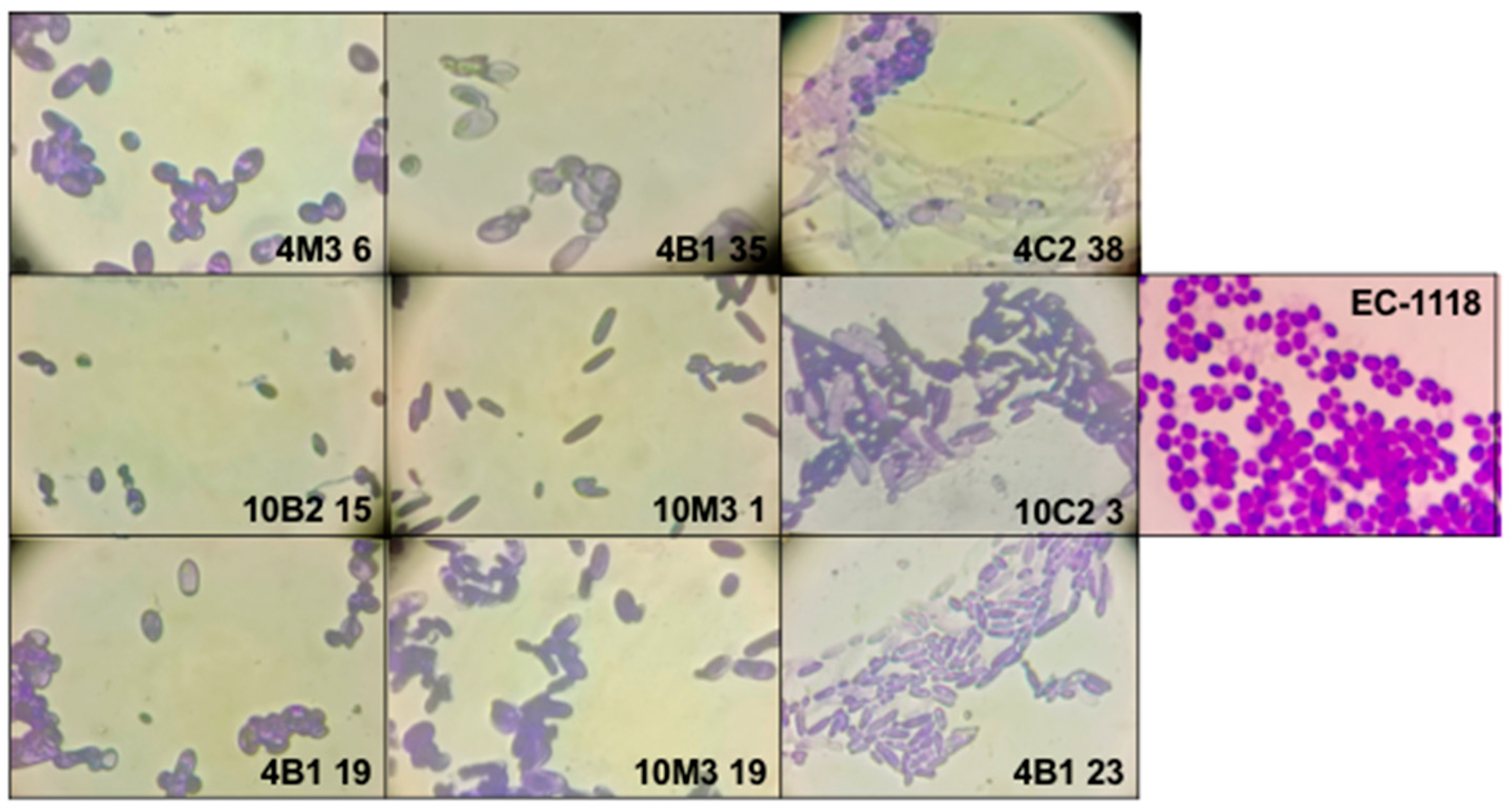
3.3. Molecular Characterization of Antarctic Fermentative Yeasts
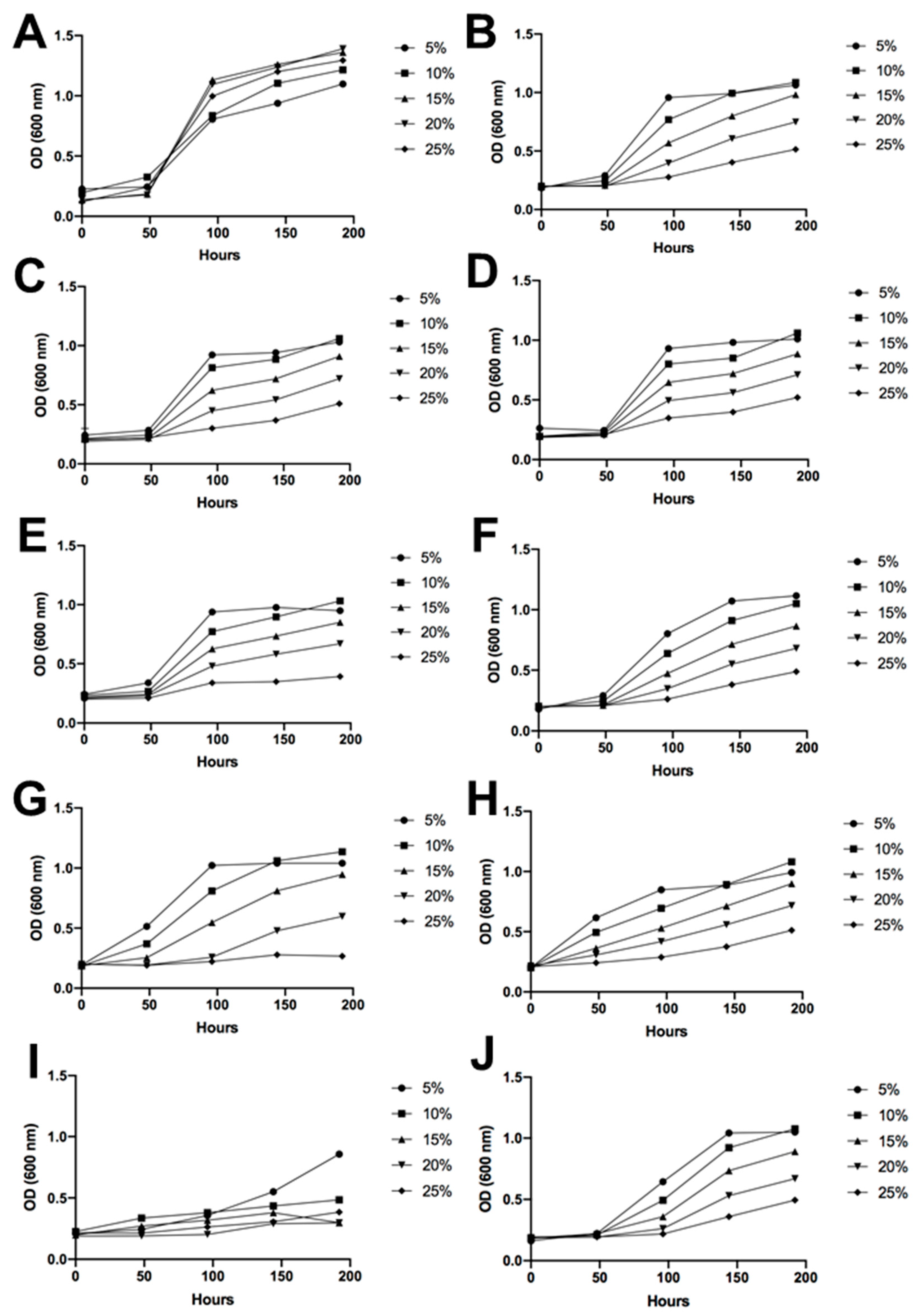
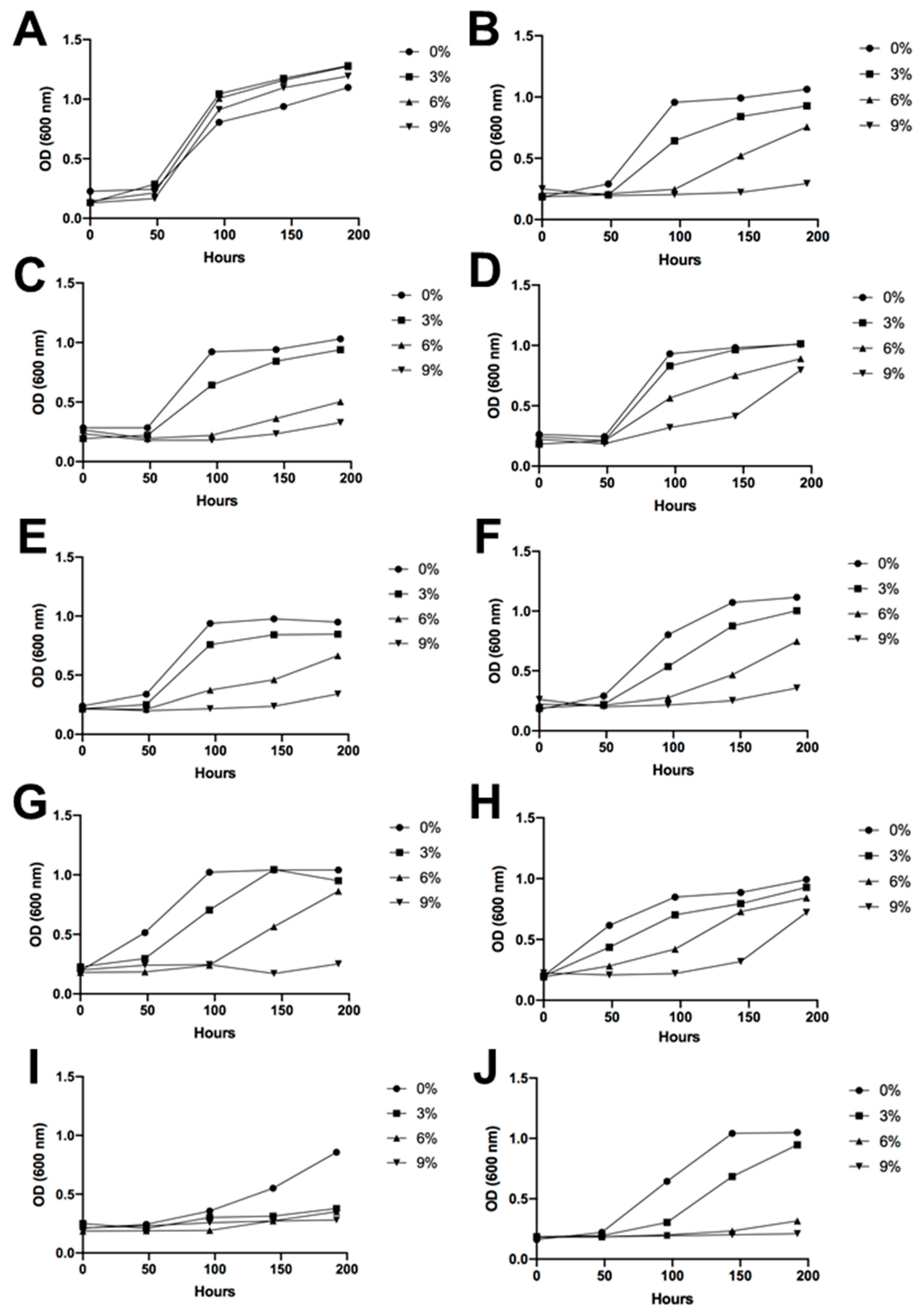
3.4. Sugar and Alcohol Tolerance of Antarctic Fermentative Yeasts
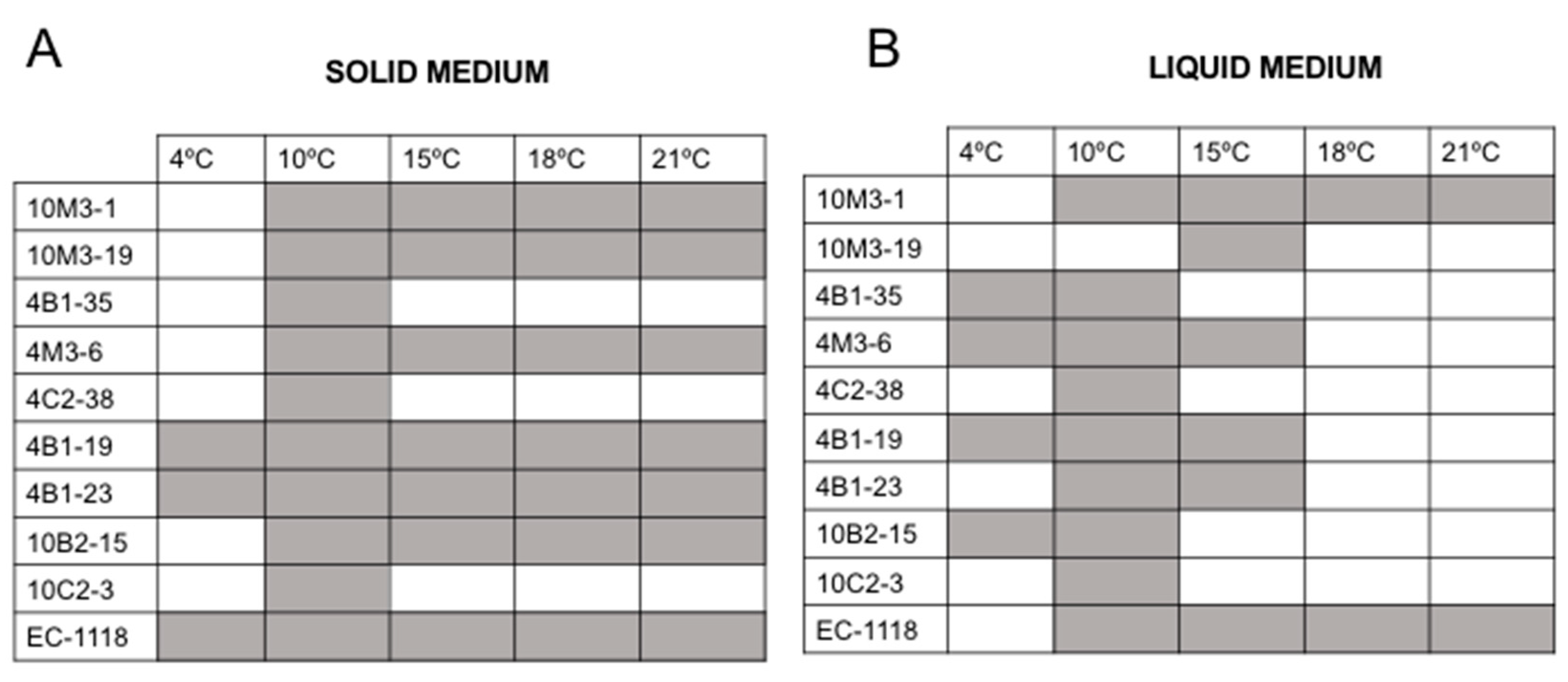
3.5. Optimal Growth Temperature
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Leeuwen, C.; Seguin, G. The concept of terroir in viticulture. J. Wine Res. 2006, 17, 1–10. [Google Scholar] [CrossRef]
- Hernandez-Orte, P.; Concejero, B.; Astrain, J.; Lacau, B.; Cacho, J.; Ferreira, V. Influence of viticulture practices on grape aroma precursors and their relation with wine aroma. J. Sci. Food Agric. 2014, 95, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Curiel, J.A.; Morales, P.; Gonzalez, R.; Tronchoni, J. Different Non-Saccharomyces Yeast Species Stimulate Nutrient Consumption in S. cerevisiae Mixed Cultures. Front. Microbiol. 2017, 8, 2121. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aroma: A review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef]
- Chambers, P.; Borneman, A.; Varela, C.; Cordente, A.; Bellon, J.; Tran, T.; Henschke, P.; Curtin, C. Ongoing domestication of wine yeast: Past, present and future. Aust. J. Grape Wine Res. 2015, 21, 642–650. [Google Scholar] [CrossRef]
- Boulton, R.B.; Singleton, V.L.; Bisson, L.F.; Kunkee, E.R. Principles and Practices of Winemaking, 1st ed.; Chapman & Hall: New York, NY, USA, 1996; pp. 146–150. [Google Scholar]
- GIL, J.; Mateo, J.; Jiménez, M.; Pastor, A.; Huerta, T. Aroma Compounds in Wine as Influenced by Apiculate Yeasts. J. Food Sci. 1996, 61, 1247–1250. [Google Scholar] [CrossRef]
- Manzanares, P.; Ramon, D.; Querol, A. Screening of non-Saccharomyces wine yeasts for the production of β-D-xylosidase activity. Int. J. Food Microbiol. 1999, 46, 105–112. [Google Scholar] [CrossRef]
- Strauss, M.L.A.; Jolly, N.P.; Lambrechts, M.G.; Van Rensburg, P. Screening for the production of extracellular hydrolytic enzymes by non-Saccharomyces wine yeasts. J. Appl. Microbiol. 2001, 91, 182–190. [Google Scholar] [CrossRef]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aroma evolution throughout alcoholic fermentation sequentially inoculated with non- Saccharomyces/Saccharomyces yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef]
- Quirós, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of non-Saccharomyces yeast strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Rojas, V.; Quirós, M.; Gonzalez, R. The impact of oxygen on the final alcohol content of wine fermented by a mixed starter culture. Appl. Microbiol. Biotechnol. 2015, 99, 3993–4003. [Google Scholar] [CrossRef]
- Molina, A.M.; Swiegers, J.H.; Varela, C.; Pretorius, I.S.; Agosin, E. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. Biotechnol. 2007, 77, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Scrimgeour, N.; Nordestgaard, S.; Lloyd, N.; Wilkes, E. Exploring the effect of elevated storage temperature on wine composition. Aust. J. Grape Wine Res. 2015, 21, 713–722. [Google Scholar] [CrossRef]
- Beltran, G.; Novo, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effect of fermentation temperature and culture media on the yeast lipid composition and wine volatile compounds. Int. J. Food Microbiol. 2008, 121, 169–177. [Google Scholar] [CrossRef] [PubMed]
- López-Malo, M.; Chiva, R.; Rozes, N.; Guillamon, J.M. Phenotypic analysis of mutant and overexpressing strains of lipid metabolism genes in Saccharomyces cerevisiae: Implication in growth at low temperatures. Int. J. Food Microbiol. 2013, 162, 26–36. [Google Scholar] [CrossRef]
- Torija, M.J.; Beltran, G.; Novo, M.; Poblet, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effects of fermentation temperature and Saccharomyces species on the cell wall fatty acid composition and presence of volatile compounds in wine. Int. J. Food Microbiol. 2003, 85, 127–136. [Google Scholar] [CrossRef]
- Massera, A.; Assof, M.; Sari, S.; Ciklic, I.; Mercado, L.; Jofré, V.; Combina, M. Effect of low temperature fermentation on the yeast-derived volatile aroma composition and sensory profile in Merlot wines. LWT 2021, 142, 111069. [Google Scholar] [CrossRef]
- Li, H.; Guo, A.; Wang, H. Mechanisms of oxidative browning of wine. Food Chem. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Buxaderas, S.; López-Tamames, E. Wines|production of sparkling wines. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Trugo, L., Finglas, P., Eds.; Elsevier Science: London, UK, 2010; Volume 10, pp. 6203–6209. [Google Scholar]
- Ferreira, V.; Escudero, A.; Fernández, P.; Cacho, J.F. Changes in the profile of volatile compounds in wines stored under oxygen and their relationship with the browning process. Z. Lebensm. Unters. Forsch. 1997, 205, 392–396. [Google Scholar] [CrossRef]
- Salvadó, Z.; Arroyo-López, F.N.; Guillamón, J.M.; Salazar, G.; Querol, A.; Barrio, E. Temperature adaptation markedly determines evolution within the genus Saccharomyces. Appl. Environ. Microbiol. 2011, 77, 2292–2302. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.R.; Fell, J.W. Molecular analyses of the IGS & ITS regions of rDNA of the psychrophilic yeasts in the genus Mrakia. Antonie Van Leeuwenhoek 2000, 77, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, J.; Randez-Gil, F.; Prieto, J.A. Cold response in Saccharomyces cerevisiae: New functions for old mechanisms. FEMS Microbiol. Rev. 2007, 31, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Margesin, R.; Miteva, V. Diversity and ecology of psychrophilic microorganisms. Res. Microbiol. 2011, 162, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Córcoles-Sáez, I.; Ballester-Tomas, L.; de la Torre-Ruiz, M.A.; Prieto, J.A.; Randez-Gil, F. Low temperature highlights the functional role of the cell wall integrity pathway in the regulation of growth in Saccharomyces cerevisiae. Biochem. J. 2012, 446, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Turchetti, B.; Hall, S.R.T.; Connell, L.B.; Branda, E.; Buzzini, P.; Theelen, B.; Müller, W.H.; Boekhout, T. Psychrophilic yeasts from Antarctica and European glaciers: Description of Glaciozyma gen. nov., Glaciozyma martinii sp. nov. and Glaciozyma watsonii sp. nov. Extremophiles 2011, 15, 573. [Google Scholar] [CrossRef] [PubMed]
- Bergauer, P.; Fonteyne, P.-A.; Nolard, N.; Schinner, F.; Margesin, R. Biodegradation of phenol and phenol-related compounds by psychrophilic and cold-tolerant alpine yeasts. Chemosphere 2005, 59, 909–918. [Google Scholar] [CrossRef]
- Buzzini, P.; Branda, E.; Goretti, M.; Turchetti, B. Psychrophilic yeasts from worldwide glacial habitats: Diversity, adaptation strategies and biotechnological potential. FEMS Microbiol. Ecol. 2012, 82, 217–241. [Google Scholar] [CrossRef]
- De Garcãa, V.; Brizzio, S.; Libkind, D.; Buzzini, P.; Van Broock, M. Biodiversity of cold-adapted yeasts from glacial meltwater rivers in Patagonia, Argentina. FEMS Microbiol. Ecol. 2007, 59, 331–341. [Google Scholar] [CrossRef]
- Vaz, A.B.M.; Rosa, L.H.; Vieira, M.L.A.; De Garcia, V.; Brandão, L.R.; Teixeira, L.C.R.S.; Moliné, M.; Libkind, D.; Van Broock, M.; Rosa, C.A. The diversity, extracellular enzymatic activities and photoprotective compounds of yeasts isolated in Antarctica. Braz. J. Microbiol. 2011, 42, 937–947. [Google Scholar] [CrossRef]
- Carrasco, M.; Rozas, J.M.; Barahona, S.; Alcaíno, J.; Cifuentes, V.; Baeza, M. Diversity and extracellular enzymatic activities of yeasts isolated from King George Island, the sub-Antarctic region. BMC Microbiol. 2012, 12, 251. [Google Scholar] [CrossRef]
- Touchette, D.; Altshuler, I.; Gostinčar, C.; Zalar, P.; Raymond-Bouchard, I.; Zajc, J.; McKay, C.P.; Gunde-Cimerman, N.; Whyte, L.G. Novel Antarctic yeast adapts to cold by switching energy metabolism and increasing small RNA synthesis. ISME J. 2022, 16, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Baeza, M.; Alcaíno, J.; Cifuentes, V.; Turchetti, B.; Buzzini, P. Cold-Active Enzymes from Cold-Adapted Yeasts. In Biotechnology of Yeasts and Filamentous Fungi, 1st ed.; Sibirny, A.A., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 297–324. [Google Scholar]
- Yusof, N.A.; Hashim, N.H.F.; Bharudin, I. Cold adaptation strategies and the potential of psychrophilic enzymes from the antarctic yeast, Glaciozyma antarctica PI12. J. Fungi 2021, 7, 528. [Google Scholar] [CrossRef] [PubMed]
- Ballester-Tomás, L.; Prieto, J.A.; Gil, J.V.; Baeza, M.; Randez-Gil, F. The Antarctic yeast Candida sake: Understanding cold metabolism impact on wine. Int. J. Food Microbiol. 2017, 245, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Henrici, A.T. The Staining of Yeasts by Gram’s Method. J. Med. Res. 1914, 30, 409–416. [Google Scholar]
- Catrileo, D.; Acuña-Fontecilla, A.; Godoy, L. Adaptive laboratory evolution of native Torulaspora delbrueckii YCPUC10 with enhanced ethanol resistance and evaluation in co-inoculated fermentation. Front. Microbiol. 2020, 11, 595023. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F. Non-Saccharomyces wine yeasts have a promising role in biotechnological approaches to winemaking. Ann. Microbiol. 2011, 61, 25–32. [Google Scholar] [CrossRef]
- Dessi-Fulgheri, F. Antarctic ecosystems: An extreme environment in a changing world. Ethol. Ecol. Evol. 2015, 27, 335–344. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Güzel, N.; Güzel, M.; Savaş Bahçei, K. Nonalcoholic Beer. In Trends in Non-Alcoholic Beverages, 1st ed.; Galanakis, C.M., Ed.; Elsevier Academic Press: London, UK, 2020; pp. 167–200. ISBN 9780128169384. [Google Scholar]
- Tsuji, M.; Kudoh, S.; Hoshino, T. Ethanol productivity of cryophilic basidiomycetous yeast Mrakia spp. correlates with ethanol tolerance. Mycoscience 2016, 57, 42–50. [Google Scholar] [CrossRef]
- Tsuji, M.; Tanabe, Y.; Vincent, W.F.; Uchida, M. Mrakia arctica sp. nov., a new psychrophilic yeast isolated from an ice island in the Canadian High Arctic. Mycoscience 2018, 59, 54–58. [Google Scholar] [CrossRef]
- Linnakoski, R.; Jyske, T.; Eerikäinen, R.; Veteli, P.; Cortina-Escribano, M.; Magalhães, F.; Järvenpää, E.; Heikkilä, L.; Hutzler, M.; Gibson, B. Brewing potential of strains of the boreal wild yeast Mrakia gelida. Front. Microbiol. 2023, 14, 1108961. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, G.; Sannino, C.; Sileoni, V.; Marconi, O.; Filippucci, S.; Tasselli, G.; Turchetti, B. Mrakia gelida in brewing process: An innovative production of low alcohol beer using a psychrophilic yeast strain. Food Microbiol. 2018, 76, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Singh, R.P.; Khare, R. Influence of climate change on Antarctic flora. Polar Sci. 2018, 18, 94–101. [Google Scholar] [CrossRef]
- Penacho, V.; Valero, E.; Gonzalez, R. Transcription profiling of sparkling wine second fermentation. Int. J. Food Microbiol. 2012, 153, 176–182. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isotype | Yeasts | Selected Yeast |
|---|---|---|
| I1 | 4B1-26, 4B1-25, 4B1-24, 4B1-22, 4B1-21, and 4B1-20 | 4B1-35 |
| I2 | 4B1-23 | 4B1-23 |
| I3 | 4B1-19 | 4B1-19 |
| I4 | 4C2-38, 4C2-39, and 4C2-4 | 4C2-38 |
| I5 | 10B2-13 and 10C2-3 | 10C2-3 |
| I6 | 10B2-15 | 10B2-15 |
| I7 | 10M3-1, 10M3-4, 10M3-9, 10M3-12, 10M3-21, and 10M3-22 | 10M3-1 |
| I8 | 10M3-19 | 10M3-19 |
| I9 | 4M3-6 and 10M3-20 | 4M3-6 |
| Yeast (Isotype) | Identification | Isotype ITS Length (bp)/GenBank Sequence Length (bp) | Identity (%)/Covered (%) | GenBank Accession Number |
|---|---|---|---|---|
| 4B1-35 (I1) | Mrakia blollopis CPCC 300333 | 550/639 | 100/100 | MZ683240.1 |
| Mrakia psychrophilia T5Mp | 550/595 | 100/100 | JQ857018.1 | |
| Mrakia psychrophilia AS 2.1971 | 550/601 | 100/100 | EU224267.1 | |
| Mrakia psychrophilia AB16-13 | 550/610 | 99.81/100 | LC125322.1 | |
| Mrakia psychrophilia BF-OTU180 | 550/813 | 99.81/100 | AM901861.1 | |
| 4B1-23 (I2) | Mrakia robertii YH18 | 504/ 635 | 100/100 | MT048630.1 |
| Mrakia robertii YH01 | 504/631 | 100/100 | MT048621.1 | |
| Mrakia robertii GR2-2-4-9 | 504/1207 | 100/100 | LC514985.1 | |
| Mrakia robertii A2(b) | 504/ 1210 | 100/100 | MH481688.1 | |
| Mrakia robertii AI | 504/1212 | 100/100 | MH481684.1 | |
| 4B1-19 (I3) | Mrakia robertii YH18 | 513/ 635 | 100/100 | MT048630.1 |
| Mrakia robertii YH01 | 513/631 | 100/100 | MT048621.1 | |
| Mrakia robertii GR2-2-4-9 | 513/1207 | 100/100 | LC514985.1 | |
| Mrakia robertii A2(b) | 513/1210 | 100/100 | MH481688.1 | |
| Mrakia robertii AI | 513/1212 | 100/100 | MH481684.1 | |
| 4C2-38 (I4) | Mrakia blollopis CPCC 300333 | 491/639 | 100/100 | MZ683240.1 |
| Mrakia sp. I12F-02259 | 491/619 | 100/100 | JX852329.1 | |
| Mrakia psychrophilia T5Mp | 491/595 | 100/100 | JQ857018.1 | |
| Mrakia psychrophilia AS 2.1971 | 491/601 | 100/100 | EU224267.1 | |
| Mrakia psychrophilia AB16-13 | 491/610 | 99.80/100 | LC125322.1 | |
| 10C2-3 (I5) | Mrakia blollopis CPCC 300333 | 513/639 | 100/100 | MZ683240.1 |
| Mrakia psychrophilia AB16-13 | 513/610 | 99.81/100 | LC125322.1 | |
| Mrakia blollopis MKOTU31 | 513/606 | 99.60/100 | KP714624.1 | |
| Mrakia psychrophilia AB16-12 | 513/606 | 99.61/100 | LC125311.1 | |
| Mrakia psychrophilia AS 2.1971 | 513/601 | 100/98 | EU224267.1 | |
| 10B2-15 (I6) | Mrakia robertii YH18 | 554/ 635 | 100/100 | MT048630.1 |
| Mrakia robertii YH01 | 554/631 | 100/100 | MT048621.1 | |
| Mrakia robertii GR2-2-4-9 | 554/1207 | 100/100 | LC514985.1 | |
| Mrakia robertii A2(b) | 554/ 1210 | 100/100 | MH481688.1 | |
| Mrakia sp. isolate J-39 | 554/1212 | 100/100 | KY782313.1 | |
| 10M3-1 (I7) | Mrakia blollopis CPCC 300333 | 528/639 | 100/100 | MZ683240.1 |
| Mrakia psychrophilia BF-OTU180 | 528/813 | 99.82/100 | AM901861.1 | |
| Mrakia psychrophilia AB16-13 | 528/610 | 99.82/100 | LC125322.1 | |
| Mrakia psychrophilia AB16-2 | 528/610 | 99.64/100 | LC125311.1 | |
| Mrakia psychrophilia T5Mp | 528/595 | 100/98 | JQ857018.1 | |
| 10M3-19 (I8) | Mrakia blollopis CPCC 300333 | 502/639 | 100/100 | MZ683240.1 |
| Mrakia psychrophilia T5Mp | 502/595 | 100/100 | JQ857018.1 | |
| Mrakia psychrophilia AS 2.1971 | 502/601 | 100/100 | EU224267.1 | |
| Mrakia psychrophilia AB16-13 | 502/610 | 99.80/100 | LC125322.1 | |
| Mrakia psychrophilia BF-OTU180 | 502/813 | 99.80/100 | AM901861.1 | |
| 4M3-6 (I9) | Mrakia blollopis CPCC 300333 | 492/639 | 100/100 | MZ683240.1 |
| Mrakia sp. I12F-02259 | 492/619 | 100/100 | JX852329.1 | |
| Mrakia psychrophilia T5Mp | 492/595 | 100/100 | JQ857018.1 | |
| Mrakia psychrophilia AS 2.1971 | 492/601 | 100/100 | EU224267.1 | |
| Mrakia blollopis MKOTU31 | 492/606 | 99.50/100 | KP714624.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro, L.; Gil i Cortiella, M.; Gutiérrez-Moraga, A.; Calisto, N.; Ubeda, C.; Corsini, G. Antarctic Soil Yeasts with Fermentative Capacity and Potential for the Wine Industry. Foods 2023, 12, 4496. https://doi.org/10.3390/foods12244496
Navarro L, Gil i Cortiella M, Gutiérrez-Moraga A, Calisto N, Ubeda C, Corsini G. Antarctic Soil Yeasts with Fermentative Capacity and Potential for the Wine Industry. Foods. 2023; 12(24):4496. https://doi.org/10.3390/foods12244496
Chicago/Turabian StyleNavarro, Laura, Mariona Gil i Cortiella, Ana Gutiérrez-Moraga, Nancy Calisto, Cristina Ubeda, and Gino Corsini. 2023. "Antarctic Soil Yeasts with Fermentative Capacity and Potential for the Wine Industry" Foods 12, no. 24: 4496. https://doi.org/10.3390/foods12244496
APA StyleNavarro, L., Gil i Cortiella, M., Gutiérrez-Moraga, A., Calisto, N., Ubeda, C., & Corsini, G. (2023). Antarctic Soil Yeasts with Fermentative Capacity and Potential for the Wine Industry. Foods, 12(24), 4496. https://doi.org/10.3390/foods12244496