
Evidence for the Beneficial Effects of Brazilian Native Fruits and Their By-Products on Human Intestinal Microbiota and Repercussions on Non-Communicable Chronic Diseases—A Review
 , and
, and 
Abstract
:1. Introduction
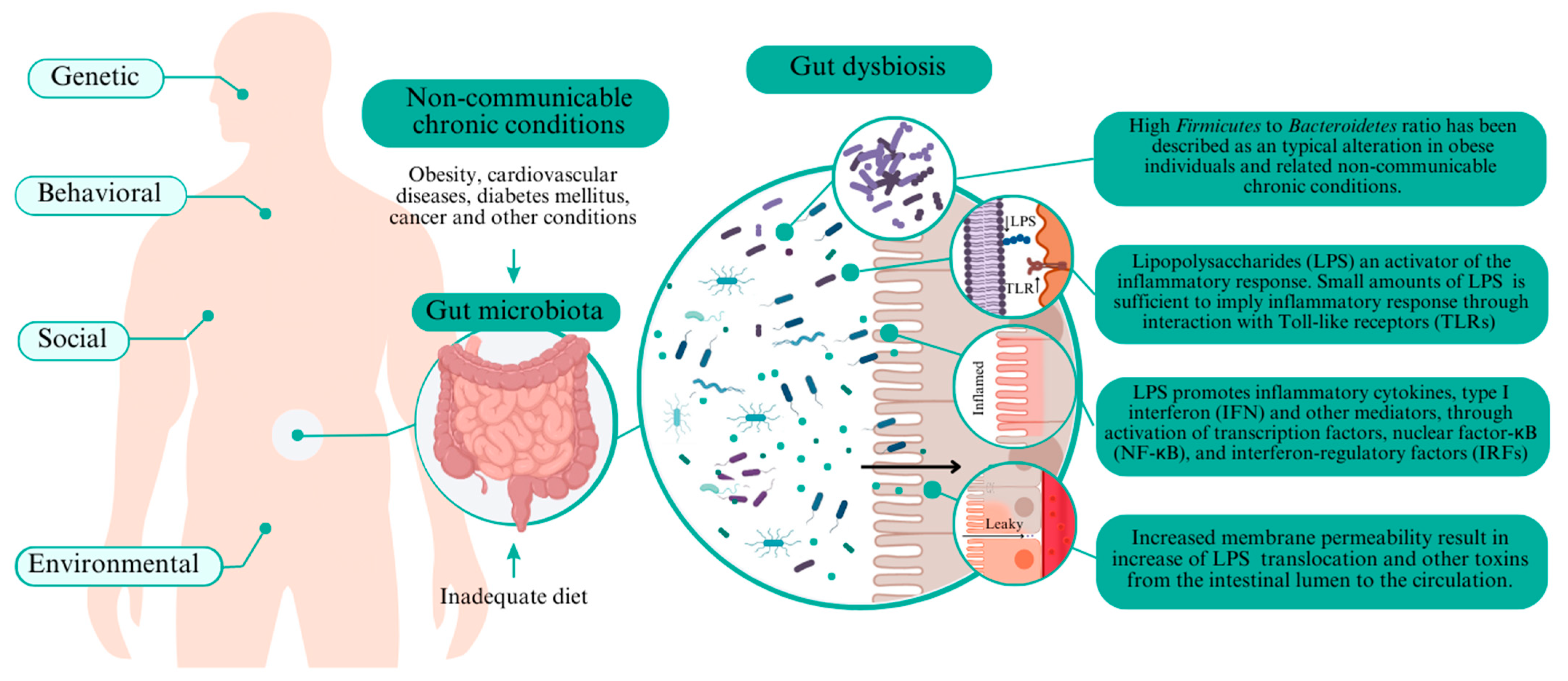
2. The Potential Role of Intestinal Microbiota in NCD
3. Bioactive Compounds in Brazilian Native Fruits
4. Potential Effects of Brazilian Native Fruits and Their By-Products on Intestinal Microbiota and Clinical Outcomes
4.1. Acerola (Malpighia emarginata D.C.)

{kind=link}
| In Vitro Studies | |||
|---|---|---|---|
| Brazilian Fruit | Bioactive Compounds | Main Results | Reference |
| Acerola (Malpighia emarginata D.C.) | Insoluble and soluble dietary fibers, cyanidin3-rhamnoside, (+)-catechin, isorhamnetin, 2,5-dihydroxybenzoic acid, catechin, myricetin, salicylic acid, and rutin. | Acerola and guava by-products previously submitted to simulated gastrointestinal digestion increased the abundance of Bifidobacterium spp., Lactobacillus spp./Enterococcus spp., Bacteroides/Prevotella, and E. rectale/C. coccoides, and decreased the abundance of C. histolyticum during colonic fermentation, besides increasing SCFA concentration and decreasing pH. | [15,16] |
| Acerola (Malpighia emarginata D.C.) | Insoluble and soluble dietary fibers, cyanidin3-rhamnoside, (+)-catechin, isorhamnetin, 2,5-dihydroxybenzoic acid, catechin, myricetin, salicylic acid, and rutin. | ||
| Guava (Psidium guajava L.) | Insoluble and soluble dietary fibers, 3,4 dihydroxybenzoic acid, salicylic acid, 2,5-dihidorxybenzoic acid, myricetin, synapic acid. | ||
| Açaí (Euterpe oleracea Mart.) | Gallic acid, protocatechuic acid, p-hydroxybenzoic acid, gentisic acid, chlorogenic acid, caffeic acid, syringic acid, ferulic acid, trains-cinnamic acid, quercetin, vanillic acid, cyanidin-3-O-glucoside, cyanidin-3-O-rutinoside, pelargonidin-3-O-glucoside, and peonidin-3-O-rutinoside. | Açaí pulp digestion resulted in the degradation of phenolic compounds, decreased abundance of Bacteroides/Prevotella spp. and Clostridium-histolyticum groups, and increased SCFA. Açaí caused no cytotoxic effects in HT29 cells and reduced DNA damage, indicating an anti-genotoxicity effect. | [87] |
| Buriti (Mauritia flexuosa L.) | Insoluble and soluble fibers, isoquercetin, ferulic acid, vanillic acid, caffeic acid, and quercetin. | Fermented milk with passion and buriti fruit into SHIME® resulted in an increased abundance of Bacteroidetes and Actinobacteria, reduced abundance of Proteobacteria phyla, decreased ammonia amounts, and increased the contents of propionic and butyric acids. | [48,88] |
| Passion (Passiflora capsularis L.) | Insoluble and soluble fibers, phenolic compounds, and carotenoids. | Low-fat goat milk fermented by Lactobacillus casei Lc-1 and supplemented with passion fruit by-product decreased the abundance of Prevotella, Megamonas, and Succinivibrio genera, and increased the abundance of Lactobacillus and Bifidobacterium genera. | [44] |
| Jabuticaba (Myrciaria jaboticaba (Vell.) Berg) | Castalagin, vescalagin, procyanidin a2, ellagic acid, gallic acid, cyanidin-3-O-glucoside, delphinidin-3-O-glucoside, malvidin-3-O-glucoside, pelargonidin-3-O-glucoside, peonidin, and hesperidin. | Simulated gastrointestinal digestion and colonic fermentation of jabuticaba by-product increased the contents of caftaric acid, gallic acid, catechin, epicatechin gallate, procyanidin A1, procyanidin B1, and procyanidin B2, and decreased the contents of cyanidin 3-glucoside, delphinidin 3-glucoside, epicatechin, hesperidin, kaempferol 3-glucoside, and cis-resveratrol. Jabuticaba by-product decreased pH, increased SCFA, increased the population of Lactobacillus spp./Enterococcus spp., Bifidobacterium spp., and E. rectale/C. coccoides, and decreased the population of Bacteroides spp./Prevotella spp. during colonic fermentation. | [17,55] |
| Consumption of the yogurt supplemented with lyophilized jabuticaba seed extract increased alpha diversity and equalized the bacterial biodiversity in feces, as well as promoted cytotoxic effects on cancer cells and exhibited antioxidant activity by reducing reactive oxygen species generation. | [55,90] | ||
| Juçara (Euterpe edulis Mart.) | Cyanidin-3-rutinoside, cyanidin-3-glucoside, malvidin-3-glucoside, peonidin-3-rutinoside, pelargonidin-3-glucoside, rutin, quercetin, and p-coumaric acid. | Juçara pulp colonic fermentation increased the abundance of Bifidobacterium spp., Eubacterium rectale/Clostridium coccoides group and Bacteroides spp./Prevotella, reduced the abundance of Clostridium histolyticum, and caused no alteration in populations of Lactobacillus/Enterococcus spp. | [91] |
| Preclinical studies | |||
| Acerola (Malpighia emarginata D.C.) | Insoluble and soluble dietary fibers, cyanidin3-rhamnoside, (+)-catechin, isorhamnetin, 2,5-dihydroxybenzoic acid, catechin, myricetin, salicylic acid, and rutin. | Consumption of guava and acerola by-products improved lipid metabolism and weight loss, decreased fecal pH, increased fat excretion, decreased liver fat accumulation, maintained the integrity of crypts, goblet cells, and epithelial cells at the colon, decreased the fecal viable cell counts of Enterobacteriaceae and increased Bifidobacterium spp. and Lactobacillus spp. viable cell counts. Acerola by-product improved fecal moisture and total SCFA contents in the feces. | [15,16] |
| Guava (Psidium guajava L.) | Insoluble and soluble dietary fibers, 3,4 dihydroxybenzoic acid, salicylic acid, 2,5-dihidorxybenzoic acid, myricetin, and synapic acid. | ||
| Açaí | Cyanidin-3-rutinoside, cyanidin-3-glucoside, and delphinidin-3-glucoside. | Administration of anthocyanin-rich extract of açaí to C57BL/6J obese high-fat-fed mice reduced body weight and lipid accumulation in adipose tissue, improved lipid metabolism, reduced liver damage and steatosis, improved liver function, glucose intolerance, glucose and insulin levels, HOMA-IR index, and reduced lipogenesis-related genes expression. It changed OTU abundance clustering of gut microbiota, decreased the abundance of Firmicutes and Proteobacteria, and increased the abundance of Verrucomicrobia and Akkermansia muciniphila in the feces. | [54] |
| Guava (Psidium guajava L.) | Galic acid, epicatchin, cathechin, caffeic acid, kaempferol, chorogenic acid, quercetin, guavinosides A and B, guavinoside C, and psiguadials A and B. | Administration of guava leaves extract improved glucose tolerance and insulin sensitivity and reduced inflammatory cell infiltration, fibrosis in the kidney, fatty liver accumulation, total cholesterol concentration in plasma, expression of hepatic gluconeogenesis-related genes. It increased microbial diversity and species number, Bacteroidetes and Verrucomicrobia phylum abundance, and decreased Firmicutes, Actinobacteria, and Firmicutes to Bacteroidetes ratio in db/db mice. | [33,109] |
| Uronic acid, galacturonic acid, galactose, and arabinose. | Administration of guava polysaccharides in C57BL/6 mice reduced weight gain and energy intake, restored lipid metabolism, reduced blood glucose and glucose intolerance, improved liver function, reduced hepatic steatosis and TNF-α level, restored AMPKα phosphorylation, prevented abnormal lipogenesis, and decreased adipose tissue inflammation. It decreased Firmicutes to Bacteroidetes ratio and abundance of Mucispirillum genus, and restored the total SCFA concentration in the feces. | [56] | |
| Galic acid, epicatchin, cathechin, caffeic acid, kaempferol, chorogenic acid, quercetin, guavinosides A and B, guavinoside C, and psiguadials A and B. | KM mice with induced diarrhea treated with guava extracts reduced stools and diarrhea rate, diarrhea index, and intestinal propulsion rate, increased the α-diversity of the intestinal microbiota, decreased Deferribacteraceae family, changed OTU abundance cluster, and restored microbial Alpha diversity in the feces. | [108,109] | |
| Jabuticaba (Myrciaria jaboticaba (Vell.) Berg) | Cyanidin-3-O-glucoside, delphinidin-3-O-glucoside, gallic acid, rutin, myricetin, and quercetin. | Administration of aqueous extract of jabuticaba peel treatment in colitis reduced body weight loss, improved stool consistency score, and spleen enlargement. It increased the viable counts of Bifidobacteria spp. and Lactobacillus spp., reduced the viable counts of enterobacteria, restored total bacterial aerobic count, and increased butyric and acetic acid production in the feces. | [79] |
| Proanthocyanidins, ellagitannins, castalagin, vescalagin, procyanidin A2, and ellagic acid. | Lyophilized jabuticaba seed extract protected against DNA damage, inhibited LDL oxidation, and enhanced the effect of cisplatin (chemotherapy), thereby reducing chromosomal aberrations in A549 cells. Wistar rats receiving yogurt supplemented with jabuticaba extract had decreased aberrant crypt foci in the colon, was protected against colon pathological remodeling, had reduced leukocyte infiltration of the epithelium and lamina propria, COX-2 and TNF-α expression in the colon, and expression of RNA from antiapoptotic cytokines. The seed extract increased bacterial abundance in the feces and restored phylum abundance. | [106] | |
| Gallic acid, cyanidin-3-O-glucoside, delphinidin- 3-O-glucoside, malvidin-3-O-glucoside, pelargonidin-3-O-glucoside, and peonidin. | Freeze-dried jabuticaba peel supplementation reduced weight gain and inflammatory markers expression, and increased cholesterol excretion, SCFA contents, and the abundance of Bacteroidota and Patescibacteria phylum, besides reducing the relative abundance of Actinobacteriota and Firmicutes in the feces of black mice (C57BL/6). | [55] | |
| Ellagic acid, gallic acid, 3,4-dihydroxybenzoic acid, quercetin-3-O-rutinoside, myricetin-3-O-rhamnoside, quercetin, cyanidin-3-O-glucoside, and delphinidin-3-O-glucoside. | Freeze-dried jabuticaba peel and seed to C57BL/6 J mice with NAFLD reduced weight gain, glucose intolerance, glucose levels, insulin levels, HOMA-IR index, and total cholesterol, LDL-c level, IL-6 and TNF-α levels, improved liver function, reduced steatosis and fibrosis, improved gut barrier (mucin and claudin), and reduced LPS plasma concentration. It reduced Firmicutes to Bacteroidetes ratio and the abundance of Bifidobacteriaceae, Mogibacteriaceae, Christensenellaceae, Clostridiaceae, Dehalobacteriaceae, Peptococcaceae, Peptostreptococcaceae, and Ruminococcaceae in the feces. | [29] | |
| Clinical trial | |||
| Juçara (Euterpe edulis Mart.) | Anthocyanins, cyanidin 3-rutinoside, and cyanidin 3-glucoside. | Ingestion of freeze-dried juçara for 6 weeks enhanced fecal acetic acid content and the relative abundance of A. muciniphila, Bifidobacterium spp., and C. coccoides. | [10] |
4.2. Açaí (Euterpe oleracea Mart.)
4.3. Baru (Mauritia flexuosa L.)
4.4. Buriti (Mauritia flexuosa L.)
4.5. Guava (Psidium guajava L.)
4.6. Juçara (Euterpe edulis Mart.)
4.7. Jabuticaba (Myrciaria jaboticaba (Vell.) Berg)
4.8. Passion Fruit (Passiflora capsularis L.)
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 14 August 2023).
- Chen, Y.; Zhou, J.; Wang, L. Role and mechanism of gut microbiota in human disease. Front. Cell. Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef]
- Soliman, G.A. Dietary fiber, atherosclerosis, and cardiovascular disease. Nutrients 2019, 11, 1155. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: Mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef]
- De Brito Alves, J.L.; Alves Brasil, J.M.; Maia, L.A.; Lima, M.d.C.; Sampaio, K.B.; de Souza, E.L. Phenolic compounds in hypertension: Targeting gut-brain interactions and endothelial dysfunction. J. Funct. Foods 2023, 104, 105531. [Google Scholar] [CrossRef]
- Jiang, Z.; Sun, T.; He, Y.; Gou, W.; Zuo, L.-S.; Fu, Y.; Miao, Z.; Shuai, M.; Xu, F.; Xiao, C.; et al. Dietary fruit and vegetable intake, gut microbiota, and type 2 Diabetes: Results from two large human cohort studies. BMC Med. 2020, 18, 371. [Google Scholar] [CrossRef]
- Matta, F.V.; Xiong, J.; Lila, M.A.; Ward, N.I.; Felipe-Sotelo, M.; Esposito, D. Chemical composition and bioactive properties of commercial and non-commercial purple and white açaí berries. Foods 2020, 9, 1481. [Google Scholar] [CrossRef]
- Ferreira, I.C.; da Silva, V.P.; Vilvert, J.C.; Souza, F.d.F.; de Freitas, S.T.; Lima, M.d.S. Brazilian varieties of acerola (Malpighia emarginata DC.) produced under tropical semi-arid conditions: Bioactive phenolic compounds, sugars, organic acids, and antioxidant capacity. J. Food Biochem. 2021, 45, e13829. [Google Scholar]
- Jamar, G.; Santamarina, A.B.; Casagrande, B.P.; Estadella, D.; de Rosso, V.V.; Wagner, R.; Fagundes, M.B.; Pisani, L.P. Prebiotic potential of juçara berry on changes in gut bacteria and acetate of individuals with obesity. Eur. J. Nutr. 2020, 59, 3767–3778. [Google Scholar] [CrossRef]
- Schiassi, M.C.E.V.; de Souza, V.R.; Lago, A.M.T.; Campos, L.G.; Queiroz, F. Fruits from the Brazilian Cerrado region: Physico-chemical characterization, bioactive compounds, antioxidant activities, and sensory evaluation. Food Chem. 2018, 245, 305–311. [Google Scholar] [CrossRef]
- Stafussa, A.P.; Maciel, G.M.; Rampazzo, V.; Bona, E.; Makara, C.N.; Demczuk Junior, B.; Haminiuk, C.W.I. Bioactive compounds of 44 traditional and exotic Brazilian fruit pulps: Phenolic compounds and antioxidant activity. Int. J. Food Prop. 2018, 21, 106–118. [Google Scholar] [CrossRef]
- Rabelo, T.; Farias, B.; Sanches, N.B.; Petrus, R.R.; Rodrigues Petrus, R.; Farias, T.R.B.; Sanches, N.B.; Petrus, R.R. The amazing native Brazilian fruits. Crit. Rev. Food Sci. Nut. 2023, 17, 1–18. [Google Scholar]
- Campos, D.A.; Gómez-García, R.; Vilas-Boas, A.A.; Madureira, A.R.; Pintado, M.M. Management of fruit industrial by-products—A case study on circular economy approach. Molecules 2020, 25, 320. [Google Scholar] [CrossRef] [PubMed]
- Batista, K.S.; Alves, A.F.; Lima, M.d.S.; da Silva, L.A.; Lins, P.P.; de Sousa Gomes, J.A.; Silva, A.S.; Toscano, L.T.; de Albuquerque Meireles, B.R.L.; de Magalhães Cordeiro, A.M.T.; et al. Beneficial effects of consumption of acerola, cashew or guava processing by-products on intestinal health and lipid metabolism in dyslipidaemic female Wistar rats. Br. J. Nutr. 2018, 119, 30–41. [Google Scholar] [CrossRef]
- Menezes, F.N.D.D.; de Melo, F.H.C.; Vieira, A.R.S.; Almeida, T.C.; Lima, M.S.; Aquino, J.S.; Gomez-Zavaglia, A.; Magnani, M.; de Souza, E.L. Acerola (Malpighia glabra L.) and guava (Psidium guayaba L.) industrial processing by-products stimulate probiotic Lactobacillus and Bifidobacterium growth and induce beneficial changes in colonic microbiota. J. Appl. Microbiol. 2021, 130, 1323–1336. [Google Scholar] [CrossRef]
- Massa, N.M.L.; de Oliveira, S.P.A.; Rodrigues, N.P.A.; Menezes, F.N.D.D.; dos Santos Lima, M.; Magnani, M.; de Souza, E.L. In vitro colonic fermentation and potential prebiotic properties of pre-digested jabuticaba (Myrciaria jaboticaba (Vell.) Berg) by-products. Food Chem. 2022, 15, 133003. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.d.C.L.; Magnani, M.; Lima, M.S.; de Sousa, C.P.; Dubreuil, J.D.; de Souza, E.L. Phenolic-rich extracts from acerola, cashew apple and mango by-products cause diverse inhibitory effects and cell damages on enterotoxigenic Escherichia coli. Lett. Appl. Microbiol. 2021, 75, 565–577. [Google Scholar] [CrossRef]
- De Souza, E.L.; de Albuquerque, T.M.R.; dos Santos, A.S.; Massa, N.M.L.; de Brito Alves, J.L. Potential interactions among phenolic compounds and probiotics for mutual boosting of their health-promoting properties and food functionalities—A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1645–1659. [Google Scholar] [CrossRef] [PubMed]
- Dyson, P.A.; Twenefour, D.; Breen, C.; Duncan, A.; Elvin, E.; Goff, L.; Hill, A.; Kalsi, P.; Marsland, N.; Mcardle, P.; et al. Diabetes UK Position Statements Diabetes UK Evidence-Based Nutrition Guidelines for the Prevention and Management of Diabetes. Diabet. Med. 2018, 35, 541–547. [Google Scholar] [CrossRef]
- Srour, B.; Fezeu, L.K.; Kesse-Guyot, E.; Allès, B.; Méjean, C.; Andrianasolo, R.M.; Chazelas, E.; Deschasaux, M.; Hercberg, S.; Galan, P.; et al. Ultra-processed food intake and risk of cardiovascular disease: Prospective cohort study (NutriNet-Santé). BMJ 2019, 365, 1451. [Google Scholar] [CrossRef]
- Wali, J.A.; Jarzebska, N.; Raubenheimer, D.; Simpson, S.J.; Rodionov, R.N.; O’Sullivan, J.F. Cardio-metabolic effects of high-fat diets and their underlying mechanisms—A narrative review. Nutrients 2020, 12, 1505. [Google Scholar] [CrossRef] [PubMed]
- Baidoun, F.; Elshiwy, K.; Elkeraie, Y.; Merjaneh, Z.; Khoudari, G.; Sarmini, M.T.; Gad, M.; Al-Husseini, M.; Saad, A. colorectal cancer epidemiology: Recent trends and impact on outcomes. Curr. Drug Targets 2021, 22, 998–1009. [Google Scholar] [PubMed]
- Malik, V.S.; Hu, F.B. The role of sugar-sweetened beverages in the global epidemics of obesity and chronic diseases. Nat. Rev. Endocrinol. 2022, 18, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Kliemann, N.; Rauber, F.; Bertazzi Levy, R.; Viallon, V.; Vamos, E.P.; Cordova, R.; Freisling, H.; Casagrande, C.; Nicolas, G.; Aune, D.; et al. Food processing and cancer risk in Europe: Results from the prospective EPIC cohort study. Lancet Planet. Health 2023, 7, e219–e232. [Google Scholar] [CrossRef] [PubMed]
- Wharton, S.; Lau, D.C.W.; Vallis, M.; Sharma, A.M.; Biertho, L.; Campbell-Scherer, D.; Adamo, K.; Alberga, A.; Bell, R.; Boulé, N.; et al. Obesity in adults: A clinical practice guideline. Can. Med. Assoc. J. 2020, 192, E875–E891. [Google Scholar] [CrossRef]
- Piché, M.E.; Tchernof, A.; Després, J.P. Obesity phenotypes, diabetes, and cardiovascular diseases. Circ. Res. 2020, 126, 1477–1500. [Google Scholar] [CrossRef]
- Green, M.; Arora, K.; Prakash, S. Microbial medicine: Prebiotic and probiotic functional foods to target obesity and metabolic syndrome. Int. J. Mol. Sci. 2020, 21, 2890. [Google Scholar] [CrossRef]
- Soares, E.; Soares, A.C.; Trindade, P.L.; Monteiro, E.B.; Martins, F.F.; Forgie, A.J.; Inada, K.O.P.; de Bem, G.F.; Resende, A.; Perrone, D.; et al. Jaboticaba (Myrciaria Jaboticaba) powder consumption improves the metabolic profile and regulates gut microbiome composition in high-fat diet-fed mice. Biomed. Pharmacother. 2021, 144, 112314. [Google Scholar] [CrossRef]
- Iqbal, J.; Al Qarni, A.; Hawwari, A.; Alghanem, A.F.; Ahmed, G. Metabolic syndrome, dyslipidemia and regulation of lipoprotein metabolism. Curr. Diabetes Rev. 2017, 14, 427–433. [Google Scholar] [CrossRef]
- Torres, N.; Guevara-Cruz, M.; Velázquez-Villegas, L.A.; Tovar, A.R. Nutrition and atherosclerosis. Arch. Med. Res. 2015, 46, 408–426. [Google Scholar] [CrossRef]
- Fan, J.; Watanabe, T. Atherosclerosis: Known and unknown. Pathol. Int. 2022, 72, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Zhang, F.; Wang, H.; Xie, L.; Chen, Z.; Zeng, W.; Zhou, Z.; Hu, F. Aqueous extract of guava (Psidium Guajava L.) leaf ameliorates hyperglycemia by promoting hepatic glycogen synthesis and modulating gut microbiota. Front. Pharmacol. 2022, 13, 907702. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.M. Highlighting Diabetes mellitus. Arterioscler. Thromb. Vasc. Biol. 2018, 38, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Przegla̜d Gastroenterol. 2019, 14, 89. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- Muralidharan, J.; Moreno-Indias, I.; Bulló, M.; Lopez, J.V.; Corella, D.; Castañer, O.; Vidal, J.; Atzeni, A.; Fernandez-García, J.C.; Torres-Collado, L.; et al. Effect on gut microbiota of a 1-y lifestyle intervention with Mediterranean diet compared with energy-reduced Mediterranean diet and physical activity promotion: PREDIMED-Plus Study. Am. J. Clin. Nutr. 2021, 114, 1148–1158. [Google Scholar] [CrossRef]
- Mayer, E.A.; Nance, K.; Chen, S. The gut–brain axis. Annu. Rev. Med. 2022, 73, 439–453. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2020, 19, 55–71. [Google Scholar] [CrossRef]
- O’Donnell, J.A.; Zheng, T.; Meric, G.; Marques, F.Z. The gut microbiome and hypertension. Nat. Rev. Nephrol. 2023, 19, 153–157. [Google Scholar] [CrossRef]
- Ottosson, F.; Brunkwall, L.; Ericson, U.; Nilsson, P.M.; Almgren, P.; Fernandez, C.; Melander, O.; Orho-Melander, M. Connection between BMI-related plasma metabolite profile and gut microbiota. J. Clin. Endocrinol. Metab. 2018, 103, 1491–1501. [Google Scholar] [CrossRef]
- Stanislawski, M.A.; Dabelea, D.; Lange, L.A.; Wagner, B.D.; Lozupone, C.A. Gut microbiota phenotypes of obesity. NPJ Biofilms Microbiomes 2019, 5, 18. [Google Scholar] [CrossRef]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef]
- Neves Casarotti, S.; Fernanda Borgonovi, T.; de Mello Tieghi, T.; Sivieri, K.; Lúcia Barretto Penna, A. Probiotic low-fat fermented goat milk with passion fruit by-product: In vitro effect on obese individuals’ microbiota and on metabolites production. Food Res. Int. 2020, 136, 109453. [Google Scholar] [CrossRef]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between Body Mass Index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17, 120. [Google Scholar] [CrossRef] [PubMed]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.Y.; Jang, H.M.; Kim, D.H. High-fat diet causes psychiatric disorders in mice by increasing Proteobacteria population. Neurosci. Lett. 2019, 698, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Borgonovi, T.F.; Salgaço, M.K.; de Oliveira, G.L.V.; de Carvalho, L.A.L.; Pinheiro, D.G.; Todorov, S.D.; Sivieri, K.; Casarotti, S.N.; Penna, A.L.B. Functional fermented milk with fruit pulp modulates the in vitro intestinal microbiota. Foods 2022, 11, 4113. [Google Scholar] [CrossRef]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A common factor in human diseases. BioMed Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef]
- Mohammad, S.; Thiemermann, C. Role of metabolic endotoxemia in systemic inflammation and potential interventions. Front. Immunol. 2021, 11, 3379. [Google Scholar] [CrossRef]
- Salguero, M.V.; Al-Obaide, M.A.I.; Singh, R.; Siepmann, T.; Vasylyeva, T.L. Dysbiosis of Gram-negative gut microbiota and the associated serum lipopolysaccharide exacerbates inflammation in type 2 diabetic patients with chronic kidney disease. Exp. Ther. Med. 2019, 18, 3461–3469. [Google Scholar] [CrossRef]
- Hudson, B.I.; Lippman, M.E. Targeting RAGE signaling in inflammatory disease. Annu. Rev. Med. 2018, 69, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.E.; Gillis, E.E.; Musall, J.B.; Baban, B.; Sullivan, J.C. High-fat diet-induced hypertension is associated with a proinflammatory T cell profile in male and female dahl salt-sensitive rats. Am. J. Physiol.-Heart Circ. Physiol. 2018, 315, H1713–H1723. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Shen, X.; Deng, R.; Zhang, Y.; Zheng, X. Dietary anthocyanin-rich extract of açai protects from diet-induced obesity, liver steatosis, and insulin resistance with modulation of gut microbiota in mice. Nutrition 2021, 86, 111176. [Google Scholar] [CrossRef] [PubMed]
- Loubet Filho, P.S.; Baseggio, A.M.; Vuolo, M.M.; Reguengo, L.M.; Telles Biasoto, A.C.; Correa, L.C.; Junior, S.B.; Alves Cagnon, V.H.; Betim Cazarin, C.B.; Maróstica Júnior, M.R. Gut microbiota modulation by jabuticaba peel and its effect on glucose metabolism via inflammatory signaling. Curr. Res. Food Sci. 2022, 5, 382–391. [Google Scholar] [CrossRef]
- Li, Y.; Bai, D.; Lu, Y.; Chen, J.; Yang, H.; Mu, Y.; Xu, J.; Huang, X.; Li, L. The crude guava polysaccharides ameliorate high-fat diet-induced obesity in mice via reshaping gut microbiota. Int. J. Biol. Macromol. 2022, 213, 234–246. [Google Scholar] [CrossRef]
- Zhu, L.; Han, J.; Li, L.; Wang, Y.; Li, Y.; Zhang, S. Claudin family participates in the pathogenesis of inflammatory bowel diseases and colitis-associated colorectal cancer. Front. Immunol. 2019, 10, 1441. [Google Scholar] [CrossRef]
- Ahmad, A.; Yang, W.; Chen, G.; Shafiq, M.; Javed, S.; Zaidi, S.S.A.; Shahid, R.; Liu, C.; Bokhari, H. Analysis of Gut Microbiota of Obese Individuals with type 2 Diabetes and healthy individuals. PLoS ONE 2019, 14, e0226372. [Google Scholar] [CrossRef]
- Zhang, Z.; Tian, T.; Chen, Z.; Liu, L.; Luo, T.; Dai, J. Characteristics of the gut microbiome in patients with prediabetes and type 2 Diabetes. PeerJ 2021, 9, e10952. [Google Scholar] [CrossRef]
- Rebersek, M. Gut microbiome and its role in colorectal cancer. BMC Cancer 2021, 21, 1325. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, D.Y.; Kang, J.H.; Kim, J.H.; Jeong, J.W.; Kim, H.W.; Oh, D.H.; Yoon, S.H.; Hur, S.J. Relationship between gut microbiota and colorectal cancer: Probiotics as a potential strategy for prevention. Food Res. Int. 2022, 156, 111327. [Google Scholar] [CrossRef]
- Hashemi Goradel, N.; Heidarzadeh, S.; Jahangiri, S.; Farhood, B.; Mortezaee, K.; Khanlarkhani, N.; Negahdari, B. Fusobacterium nucleatum and colorectal cancer: A mechanistic overview. J. Cell. Physiol. 2019, 234, 2337–2344. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, Q.; Fu, X. Fusobacterium nucleatum contributes to the carcinogenesis of colorectal cancer by inducing inflammation and suppressing host immunity. Transl. Oncol. 2019, 12, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Iyadorai, T.; Mariappan, V.; Vellasamy, K.M.; Wanyiri, J.W.; Roslani, A.C.; Lee, G.K.; Sears, C.; Vadivelu, J. Prevalence and association of Pks+ Escherichia coli with colorectal cancer in patients at the University Malaya Medical Centre, Malaysia. PLoS ONE 2020, 15, e0228217. [Google Scholar] [CrossRef] [PubMed]
- Pleguezuelos-Manzano, C.; Puschhof, J.; Rosendahl Huber, A.; van Hoeck, A.; Wood, H.M.; Nomburg, J.; Gurjao, C.; Manders, F.; Dalmasso, G.; Stege, P.B.; et al. Mutational signature in colorectal cancer caused by genotoxic Pks+ E. coli. Nature 2020, 580, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Martín, R.; Langella, P. Emerging Health Concepts in the probiotics field: Streamlining the definitions. Front. Microbiol. 2019, 10, 457799. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From Biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Santacroce, L.; Charitos, I.A.; Bottalico, L. A Successful history: Probiotics and their potential as antimicrobials. Exp. Rev. Anti-Infect. Ther. 2019, 17, 635–645. [Google Scholar] [CrossRef]
- Childs, C.E.; Röytiö, H.; Alhoniemi, E.; Fekete, A.A.; Forssten, S.D.; Hudjec, N.; Lim, Y.N.; Steger, C.J.; Yaqoob, P.; Tuohy, K.M.; et al. Xylo-oligosaccharides alone or in synbiotic combination with Bifidobacterium animalis subsp. lactis induce bifidogenesis and modulate markers of immune function in healthy adults: A double-blind, placebo-controlled, randomised, factorial cross-over study. Br. J. Nutr. 2014, 111, 1945–1956. [Google Scholar] [CrossRef]
- Korpela, K.; Salonen, A.; Vepsäläinen, O.; Suomalainen, M.; Kolmeder, C.; Varjosalo, M.; Miettinen, S.; Kukkonen, K.; Savilahti, E.; Kuitunen, M.; et al. Probiotic supplementation restores normal microbiota composition and function in antibiotic-treated and in caesarean-born infants. Microbiome 2018, 182, 6. [Google Scholar] [CrossRef]
- Van Zyl, W.F.; Deane, S.M.; Dicks, L.M.T. Molecular insights into probiotic mechanisms of action employed against intestinal pathogenic bacteria. Gut Microbes 2020, 12, 1831339. [Google Scholar] [CrossRef] [PubMed]
- Khalili, L.; Alipour, B.; Jafar-Abadi, M.A.; Faraji, I.; Hassanalilou, T.; Abbasi, M.M.; Vaghef-Mehrabany, E.; Sani, M.A. The effects of Lactobacillus casei on glycemic response, serum sirtuin1 and fetuin-a levels in patients with type 2 Diabetes mellitus: A randomized controlled trial. Iran. Biomed. J. 2019, 23, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Rittiphairoj, T.; Pongpirul, K.; Janchot, K.; Mueller, N.T.; Li, T. Probiotics contribute to glycemic control in patients with type 2 Diabetes mellitus: A systematic review and meta-analysis. Adv. Nutr. 2021, 12, 722–734. [Google Scholar] [CrossRef]
- Romão Da Silva, L.D.F.; De Oliveira, Y.; De Souza, E.L.; De Luna Freire, M.O.; Braga, V.D.A.; Magnani, M.; De Brito Alves, J.L. Effects of probiotic therapy on cardio-metabolic parameters and autonomic modulation in hypertensive women: A randomized, triple-blind, placebo-controlled trial. Food Funct. 2020, 11, 7152–7163. [Google Scholar] [CrossRef] [PubMed]
- O’Morain, V.L.; Ramji, D.P. The potential of probiotics in the prevention and treatment of atherosclerosis. Mol. Nutr. Food Res. 2020, 64, 1900797. [Google Scholar] [CrossRef]
- Cavalcante, R.G.S.; de Albuquerque, T.M.R.; de Luna Freire, M.O.; Ferreira, G.A.H.; Carneiro dos Santos, L.A.; Magnani, M.; Cruz, J.C.; Braga, V.A.; de Souza, E.L.; de Brito Alves, J.L. The probiotic Lactobacillus fermentum 296 attenuates cardiometabolic disorders in high fat diet-treated rats. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 1408–1417. [Google Scholar] [CrossRef]
- Duarte, F.N.D.; Rodrigues, J.B.; da Costa Lima, M.; Lima, M.d.S.; Pacheco, M.T.B.; Pintado, M.M.E.; de Souza Aquino, J.; de Souza, E.L. Potential prebiotic properties of cashew apple (Anacardium occidentale L.) agro-industrial byproduct on Lactobacillus species. J. Sci. Food Agric. 2017, 97, 3712–3719. [Google Scholar] [CrossRef]
- Da Silva-Maia, J.K.; Batisita, A.G.; Correa, L.C.; Lima, G.C.; Bogusz Junior, S.; Marostica Junior, M.R. Aqueous extract of berry (Plinia jaboticaba) byproduct modulates gut microbiota and maintains the balance on antioxidant defense system in rats. J. Food Biochem. 2019, 43, e12705. [Google Scholar] [CrossRef]
- Serra, D.; Almeida, L.M.; Dinis, T.C.P. Polyphenols in the management of brain disorders: Modulation of the microbiota-gut-brain axis. Adv. Food Nutr. Res. 2020, 91, 1–27. [Google Scholar] [PubMed]
- Alves-Santos, A.M.; Sampaio, K.B.; Lima, M.d.S.; Coelho, A.S.G.; de Souza, E.L.; Naves, M.M.V. Chemical composition and prebiotic activity of baru (Dipteryx alata Vog.) pulp on probiotic strains and human colonic microbiota. Food Res. Int. 2023, 164, 112366. [Google Scholar] [CrossRef]
- Brito Sampaio, K.; Luiz de Brito Alves, J.; Mangueira do Nascimento, Y.; Fechine Tavares, J.; Sobral da Silva, M.; dos Santos Nascimento, D.; dos Santos Lima, M.; Priscila de Araújo Rodrigues, N.; Fernandes Garcia, E.; Leite de Souza, E. Nutraceutical formulations combining Limosilactobacillus fermentum, quercetin, and or resveratrol with beneficial impacts on the abundance of intestinal bacterial populations, metabolite production, and antioxidant capacity during colonic fermentation. Food Res. Int. 2022, 161, 111800. [Google Scholar] [CrossRef] [PubMed]
- Van der Hee, B.; Wells, J.M. Microbial regulation of host physiology by short-chain fatty acids. Trends Microbiol. 2021, 29, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cai, X.; Fei, W.; Ye, Y.; Zhao, M.; Zheng, C. The role of short-chain fatty acids in immunity, inflammation and metabolism. Crit. Rev. Food Sci. Nutr. 2020, 62, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Finan, B.; Bloom, S.R.; D’Alessio, D.; Drucker, D.J.; Flatt, P.R.; Fritsche, A.; Gribble, F.; Grill, H.J.; Habener, J.F.; et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Alqurashi, R.M.; Alarifi, S.N.; Walton, G.E.; Costabile, A.F.; Rowland, I.R.; Commane, D.M. In vitro approaches to assess the effects of açai (Euterpe oleracea) digestion on polyphenol availability and the subsequent impact on the faecal microbiota. Food Chem. 2017, 234, 190–198. [Google Scholar] [CrossRef]
- Pereira-Freire, J.A.; Aquino, J.d.S.; Campos, A.R.N.; Viana, V.G.F.; Júnior, J.S.d.C.; Silva, J.D.N.; Moura, A.K.S.; Citó, A.M.D.G.L.; Moreira-Araújo, R.S.D.R.; Frota, K.d.M.G.; et al. Nutritional, physicochemical and structural parameters of Mauritia Flexuosa fruits and by-products for biotechnological exploration of sustainable goods. Food Technol. Biotechnol. 2022, 60, 155–165. [Google Scholar] [CrossRef]
- Rudke, A.R.; Mazzutti, S.; Andrade, K.S.; Vitali, L.; Ferreira, S.R.S. Optimization of green PLE method applied for the recovery of antioxidant compounds from buriti (Mauritia flexuosa L.) shell. Food Chem. 2019, 298, 125061. [Google Scholar] [CrossRef]
- Fidelis, M.; Santos, J.S.; Escher, G.B.; Rocha, R.S.; Cruz, A.G.; Cruz, T.M.; Marques, M.B.; Nunes, J.B.; do Carmo, M.A.V.; de Almeida, L.A.; et al. Polyphenols of jabuticaba [Myrciaria jaboticaba (Vell.) O.Berg] seeds incorporated in a yogurt model exert antioxidant activity and modulate gut microbiota of 1,2-dimethylhydrazine-induced colon cancer in rats. Food Chem. 2021, 334, 127565. [Google Scholar] [CrossRef]
- Guergoletto, K.B.; Costabile, A.; Flores, G.; Garcia, S.; Gibson, G.R. In vitro fermentation of juçara pulp (Euterpe edulis) by human colonic microbiota. Food Chem. 2016, 196, 251–258. [Google Scholar] [CrossRef]
- Carpiné, D.; Dagostin, J.L.A.; Mazon, E.; Barbi, R.C.T.; Alves, F.E.d.S.B.; Chaimsohn, F.P.; Ribani, R.H. Valorization of Euterpe edulis Mart. agroindustrial residues (pomace and seeds) as sources of unconventional starch and bioactive compounds. J. Food Sci. 2020, 85, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Casarotti, S.N.; Borgonovi, T.F.; Batista, C.L.F.M.; Penna, A.L.B. Guava, orange and passion fruit by-products: Characterization and its impacts on kinetics of acidification and properties of probiotic fermented products. LWT Food Sci. Technol. 2018, 98, 69–76. [Google Scholar] [CrossRef]
- Cui, J.; Lian, Y.; Zhao, C.; Du, H.; Han, Y.; Gao, W.; Xiao, H.; Zheng, J. Dietary fibers from fruits and vegetables and their health benefits via modulation of gut microbiota. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1514–1532. [Google Scholar] [CrossRef] [PubMed]
- Massa, N.M.L.; Menezes, F.N.D.D.; de Albuquerque, T.M.R.; de Oliveira, S.P.A.; Lima, M.d.S.; Magnani, M.; de Souza, E.L. Effects of Digested jabuticaba (Myrciaria jaboticaba (Vell.) Berg) by-product on growth and metabolism of Lactobacillus and Bifidobacterium indicate prebiotic properties. LWT Food Sci. Technol. 2020, 131, 109766. [Google Scholar] [CrossRef]
- Albuquerque, B.R.; Pereira, C.; Calhelha, R.C.; José Alves, M.; Abreu, R.M.V.; Barros, L.; Oliveira, M.B.P.P.; Ferreira, I.C.F.R. Jabuticaba residues (Myrciaria jaboticaba (Vell.) Berg) are rich sources of valuable compounds with bioactive properties. Food Chem. 2020, 309, 125735. [Google Scholar] [CrossRef]
- Inada, K.O.P.; Leite, I.B.; Martins, A.B.N.; Fialho, E.; Tomás-Barberán, F.A.; Perrone, D.; Monteiro, M. Jaboticaba berry: A comprehensive review on its polyphenol composition, health effects, metabolism, and the development of food products. Food Res. Int. 2021, 147, 110518. [Google Scholar] [CrossRef]
- Baptista, A.; Gonçalves, R.V.; Bressan, J.; do Carmo Gouveia Pelúzio, M. Antioxidant and antimicrobial activities of crude extracts and fractions of cashew (Anacardium occidentale L.), cajui (Anacardium microcarpum), and pequi (Caryocar brasiliense C.): A systematic review. Oxid. Med. Cell. Longev. 2018, 2018, 3753562. [Google Scholar] [CrossRef]
- Haminiuk, C.W.I.; Maciel, G.M.; Plata-Oviedo, M.S.V.; Peralta, R.M. Phenolic compounds in fruits-An overview. Int. J. Food Sci. Technol. 2012, 47, 2023–2044. [Google Scholar] [CrossRef]
- Lima, M.C.; Paiva de Sousa, C.; Fernandez-Prada, C.; Harel, J.; Dubreuil, J.D.; de Souza, E.L. A review of the current evidence of fruit phenolic compounds as potential antimicrobials against pathogenic bacteria. Microb. Pathog. 2019, 130, 259–270. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Barrow, C.J.; Dunshea, F.R. Screening and characterization of phenolic compounds and their antioxidant capacity in different fruit peels. Foods 2020, 10, 1206. [Google Scholar] [CrossRef]
- Yalcin, H.; Çapar, T.D. Bioactive compounds of fruits and vegetables. In Minimally Processed Fruits and Vegetables; Yildz, F., Wiley, R., Eds.; Food Engineering Series; Springer: Boston, MA, USA, 2017; pp. 723–745. [Google Scholar]
- Man, S.; Ye, S.; Ma, L.; Yao, Y.; Liu, T.; Yin, L. Polyphenol-rich extract from litchi chinensis seeds alleviates hypertension-induced renal damage in rats. J. Agric. Food Chem. 2021, 69, 2138–2148. [Google Scholar]
- Gowd, V.; Karim, N.; Shishir, M.R.I.; Xie, L.; Chen, W. Dietary polyphenols to combat the metabolic diseases via altering gut microbiota. Trends Food Sci. Technol. 2019, 93, 81–93. [Google Scholar] [CrossRef]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent research on the health benefits of blueberries and their anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [CrossRef]
- Do Carmo, M.A.V.; Fidelis, M.; de Oliveira, P.F.; Feitoza, L.Q.; Marques, M.J.; Ferreira, E.B.; Oh, W.Y.; Shahidi, F.; Hellström, J.; Almeida, L.A.; et al. Ellagitannins from jabuticaba (Myrciaria jboticaba) seeds attenuated inflammation, oxidative stress, aberrant crypt foci, and modulated gut microbiota in rats with 1,2 dimethyl hydrazine-induced colon carcinogenesis. Food Chem. Toxicol. 2021, 154, 112287. [Google Scholar] [CrossRef]
- Bianchi, F.; Lopes, N.P.; Adorno, M.A.T.; Sakamoto, I.K.; Genovese, M.I.; Saad, S.M.I.; Sivieri, K. Impact of combining acerola by-product with a probiotic strain on a gut microbiome model. Int. J. Food Sci. Nutr. 2018, 70, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Mao, D.; Li, X.; Ma, Y.; Luan, Y.; Cao, Y.; Luan, Y. Changes of intestinal microflora diversity in diarrhea model of km mice and effects of Psidium guajava L. as the treatment agent for diarrhea. J. Infect. Public Health 2020, 13, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Tomar, M.; Amarowicz, R.; Saurabh, V.; Nair, M.S.; Maheshwari, C.; Sasi, M.; Prajapati, U.; Hasan, M.; Singh, S.; et al. Guava (Psidium guajava L.) leaves: Nutritional composition, phytochemical profile, and health-promoting bioactivities. Foods 2021, 10, 752. [Google Scholar] [CrossRef]
- Crobach, M.J.T.; Vernon, J.J.; Loo, V.G.; Kong, L.Y.; Péchiné, S.; Wilcox, M.H.; Kuijper, E.J. Understanding Clostridium difficile colonization. Clin. Microbiol. Rev. 2018, 31, e-00021–e-00027. [Google Scholar] [CrossRef]
- Abuqwider, J.N.; Mauriello, G.; Altamimi, M. Akkermansia muciniphila, a new generation of beneficial microbiota in modulating obesity: A systematic review. Microorganisms 2021, 9, 1098. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia muciniphila is a promising probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [Google Scholar] [CrossRef]
- Aranha, L.N.; Silva, M.G.; Uehara, S.K.; Luiz, R.R.; Nogueira Neto, J.F.; Rosa, G.; Moraes de Oliveira, G.M. Effects of a hypoenergetic diet associated with açaí (Euterpe oleracea Mart.) pulp consumption on antioxidant status, oxidative stress and inflammatory biomarkers in overweight, dyslipidemic individuals. Clin. Nutr. 2020, 39, 1464–1469. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.J.; Draper, S.L.; Centanni, M.; Carnachan, S.M.; Tannock, G.W.; Sims, I.M. Characterization of polysaccharides from feijoa fruits (Acca sellowiana Berg.) and their utilization as growth substrates by gut commensal bacteroides species. J. Agric. Food Chem. 2018, 66, 13277–13284. [Google Scholar] [CrossRef] [PubMed]
- Rajan, K.; D’Souza, D.H.; Kim, K.; Choi, J.M.; Elder, T.; Carrier, D.J.; Labbé, N. Production and characterization of high value prebiotics from biorefinery-relevant feedstocks. Front. Microbiol. 2021, 12, 675314. [Google Scholar] [CrossRef] [PubMed]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-chain fatty acid propionate protects from hypertensive cardiovascular damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef]
- Kong, D.; Schipper, L.; van Dijk, G. Distinct effects of short chain fatty acids on host energy balance and fuel homeostasis with focus on route of administration and host species. Front. Neurosci. 2021, 15, 755845. [Google Scholar] [CrossRef]
- Silva-Luis, C.C.; de Brito Alves, J.L.; de Oliveira, J.C.P.L.; de Sousa Luis, J.A.; Araújo, I.G.A.; Tavares, J.F.; Do Nascimento, Y.M.; Bezerra, L.S.; Araújo de Azevedo, F.d.L.A.; Sobral, M.V.; et al. Effects of baru almond oil (Dipteryx alata Vog.) treatment on thrombotic processes, platelet aggregation, and vascular function in aorta arteries. Nutrients 2022, 14, 2098. [Google Scholar] [CrossRef]
- De Souza, R.G.M.; Gomes, A.C.; Navarro, A.M.; da Cunha, L.C.; Silva, M.A.C.; Barbosa Junior, F.; Mota, J.F. Baru almonds increase the activity of glutathione peroxidase in overweight and obese women: A randomized, placebo-controlled trial. Nutrients 2019, 11, 1750. [Google Scholar] [CrossRef]
- De Souza Aquino, J.; Batista, K.S.; Araujo-Silva, G.; dos Santos, D.C.; de Brito, N.J.N.; López, J.A.; da Silva, J.A.; das Graças Almeida, M.; Pincheira, C.G.; Magnani, M.; et al. Antioxidant and lipid-lowering effects of buriti oil (Mauritia flexuosa L.) administered to iron-overloaded rats. Molecules 2023, 28, 2585. [Google Scholar] [CrossRef]
- Chen, Z.; Radjabzadeh, D.; Chen, L.; Kurilshikov, A.; Kavousi, M.; Ahmadizar, F.; Ikram, M.A.; Uitterlinden, A.G.; Zhernakova, A.; Fu, J.; et al. Association of insulin resistance and type 2 Diabetes with gut microbial diversity: A microbiome-wide analysis from population studies. JAMA Netw. Open 2021, 4, e2118811. [Google Scholar] [CrossRef]
- Pasmans, K.; Meex, R.C.R.; Trommelen, J.; Senden, J.M.G.; Vaughan, E.E.; Van Loon, L.J.C.; Blaak, E.E. L-Arabinose co-ingestion delays glucose absorption derived from sucrose in healthy men and women: A double-blind, randomised crossover trial. Br. J. Nutr. 2022, 128, 1072–1081. [Google Scholar] [CrossRef]
- Wu, T.; Gao, Y.; Guo, X.; Zhang, M.; Gong, L. Blackberry and blueberry anthocyanin supplementation counteract high-fat-diet-induced obesity by alleviating oxidative stress and inflammation and accelerating energy expenditure. Oxid. Med. Cell. Longev. 2018, 2018, 4051232. [Google Scholar] [CrossRef] [PubMed]
- Barthichoto, M.; Moretto, T.L.; de Carvalho, F.P.; Benfato, I.D.; de Rosso, V.V.; Ribeiro, D.A.; Estadella, D.; Le Sueur-Maluf, L.; Pisani, L.P.; de Oliveira, C.A.M. Juçara (Euterpe edulis Mart.) supplementation reduces body weight gain and protects mice from metabolic complications induced by high-fat diet. Nutrire 2021, 46, 6. [Google Scholar] [CrossRef]
- Bueno, L.R.; Da Silva Soley, B.; Abboud, K.Y.; França, I.W.; Da Silva, K.S.; De Oliveira, N.M.T.; Barros, J.S.; Gois, M.B.; Cordeiro, L.M.C.; Maria-Ferreira, D. Protective effect of dietary polysaccharides from yellow passion fruit peel on dss-induced colitis in mice. Oxid. Med. Cell. Longev. 2022, 2022, 6298662. [Google Scholar] [CrossRef] [PubMed]

| Fruits | Main Bioactive Compounds in Brazilian Native Fruits Related to Intestinal Microbiota Modulation | Reference |
|---|---|---|
| Acerola (Malpighia emarginata D.C.) | ||
| Pulp | Phenolic compounds: cyanidin3-rhamnoside: 68.22 ± 0.71 mg/100 g; (+)-catechin: 2.50 ± 0.25 mg/100 g; isorhamnetin: 4.53 ± 0.25 mg/100 g. | [9] |
| By-product | Total dietary fibers: 48.57 ± 0.39 g/100 g; insoluble dietary fibers: 34.35 ± 0.29 g/100 g; soluble dietary fibers: 14.22 ± 0.10 g/100 g; phenolic compounds: 2,5-dihydroxybenzoic acid: 737.20 ± 0.4 mg/100 g; catechin: 125.20 ± 0.09 mg/100 g; myricetin: 94.20 ± 0.01 mg/100 g; salicylic acid: 348.64 ± 0.51 mg/100 g; rutin: 68.60 ± 0.05 mg/100 g. | [15] |
| Açaí (Euterpe oleracea Mart.) | ||
| Whole fruit | Total phenolic compound concentrations: 32.00 ± 1.03 mg GAE/100 g; total flavonoids: 6.39 ± 1.23 mg quercetin equivalent/g; total anthocyanins: 10.20 ± 0.24 mg cyanidin-3-glucoside equivalent/g; total proanthocyanidins: 6.10 ± 2.09 mg procyanidin B1 equivalent/g. | [8] |
| Baru (Dipteryx alata Vog.) | ||
| Pulp | Total dietary fibers: 32.90 ± 0.42 g/100 g; insoluble dietary fibers: 32.90 ± 0.42 g/100 g; soluble dietary fibers: <0.10 g/100 g; fructose: 7.76 ± 0.82 g/100 g; glucose: 6.32 ± 0.70 g/100 g; phenolic compounds: hesperidin: 19.34 ± 0.10; procyanidin B2: 0.53 ± 0.00 mg/100 g; epicatechin gallate: 0.49 ± 0.01 mg/100 g; chlorogenic acid: 0.46 ± 0.01 mg/100 g; catechin: 0.43 ± 0.00 mg/100 g; total phenolic compounds: 507.01 ± 15.41 mg GAE/100 g; total tannins: 142.91 ± 7.16 mg TAE/100 g (trolox equivalent). | [81] |
| Buriti (Mauritia flexuosa L.) | ||
| Fresh pulp | Total dietary fibers: 38.0 ± 0.3 g/100 g; insoluble dietary fibers: 27.3 ± 0.4 g/100 g; soluble dietary fibers: 10.6 ± 0.3 g/100 g. | [88] |
| Freeze-dried pulp | Total dietary fibers: 38.9 ± 0.6 g/100 g; insoluble dietary fibers: 28.8 ± 1.0 g/100 g; soluble dietary fibers: 10.1 ± 0.2 g/100 g. | |
| Freeze-dried peel | Total dietary fibers: 50.5 ± 0.6 g/100 g; insoluble dietary fibers: 50.0 ± 1.0 g/100 g; soluble dietary fibers: 0.55 ± 0.03 g/100 g. | |
| Freeze-dried endocarp | Total dietary fibers: 28.14 ± 0.05 g/100 g; insoluble dietary fibers: 24.66 ± 0.01 g/100 g; soluble dietary fibers: 3.48 ± 0.08 g/100 g. | |
| Fruit | Total pectin: 0.59 ± 0.02 g/100 g; soluble pectin: 0.49 ± 0.01 g/100 g; total phenolic compounds: 110.72 ± 0.26 mg GAE/100 g (gallic acid equivalents). | [11] |
| Fruit | Isoquercetin: 156.07; ferulic acid: 135.22 mg GAE/100 g; vanillic acid: 46.94 mg GAE/100 g; caffeic acid: 15.88 mg GAE/100 g; quercetin: 14.61 mg GAE/100 g. | [89] |
| Guava (Psidium guajava L.) | ||
| By-product | Total dietary fibers: 44.36 ± 0.23 g/100 g; insoluble dietary fibers: 40.62 ± 0.24 g/100 g; soluble dietary fibers: 7.98 ± 0.20 g/100 g; phenolic compounds: 3,4-dihydroxybenzoic acid 97.80 ± 0.11 mg/100 g; salicylic acid: 310.60 ± 0.89 mg/100 g; 2,5-dihidorxybenzoic acid: 89.40 ± 0.78 mg/100 g; myricetin: 44.60 ± 0.01 mg/100 g; synapic acid: 18.80 ± 0.0 mg/100 g. | [15] |
| Jaboticaba (Myrciaria jaboticaba (Vell.) Berg) | ||
| Seed | Total phenolic compound concentrations: 53,944 ± 773 mg GAE/100 g; phenolic compounds: castalagin: 13,349 ± 1494 mg/100 g; vescalagin: 6875 ± 1240 mg/100 g; procyanidin A2: 451 ± 81 mg/100 g; ellagic acid: 388 ± 49 mg/100 g; gallic acid: 230 ± 14 mg/100 g. | [90] |
| Freeze-dried peel | Total dietary fiber: 33.77 ± 1.20 g/100 g; insoluble dietary fiber: 25.34 ± 0.33 g/100 g; soluble dietary fiber (g/100 g): 8.49 ± 0.48 g/100 g; total phenolic compounds: 8219.94 ± 99.56 mg GAE/100 g; phenolic compounds: gallic acid: 11.39 ± 1.58 mg/100 g; cyanidin-3-O-glucoside: 675.74 ± 51.57 mg/100 g; delphinidin- 3-O-glucoside: 63.67 ± 2.67 mg/100 g; malvidin-3-O-glucoside: 17.27 ± 1.57 mg/100 g; pelargonidin-3-O-glucoside: 10.3 ± 0.17 mg/100 g; peonidin: 2.79 ± 0.84 mg/100 g. | [55] |
| By-product | Total dietary fiber: 84.75 ± 1.22 g/100 g; insoluble dietary fiber: 28.49 ± 0.82 g/100 g; soluble dietary fiber: 56.26 ± 1.42 g/100 g; total dietary fiber: 84.75 ± 1.22 g/100 g; fructose: 0.45 ± 0.08 g/100 g; glucose: 0.38 ± 0.05 g/100 g; Total phenolic compounds: 14.44 ± 0.152 mg/100 g; hesperidin: 12.07 ± 0.18 mg/100 g; gallic acid: 2.80 ± 0.05 mg/100 g. | [17] |
| Juçara (Euterpe edulis Mart.) | ||
| Freeze-dried pulp | Phenolic profile: cyanidin-3-rutinoside, cyanidin-3-glucoside, malvidin-3-glucoside, peonidin-3-rutinoside, pelargonidin-3-glucoside, rutin, quercetin, and p-coumaric acid (no quantification of a specific phenolic compound); total phenolic compound concentrations: 3474 ± 98.0 mg. | [91] |
| Pomace | Monomeric anthocyanins: 7230.0 mg cyanidin 3-O-glycoside/100 g; total dietary fibers: 72.7 g/100 g; insoluble dietary fiber: 68.62 ± 0.23 g/100 g; total phenolic compounds: 1380.0 ± 170.0 mg GAE/100 g. | [92] |
| Passion (Passiflora capsularis L.) | ||
| By-product | Total dietary fiber: 64.20 ± 0.28 g/100 g; insoluble dietary fiber: 44.80 ± 0.14 g/100 g; soluble dietary fiber: 19.40 ± 0.14 g/100 g; total phenolic compounds: 384.44 ± 22.50 mg GAE/100 g. | [93] |
| Type of Study | Intestinal Microbiota Condition | Fruit Evaluated | Dose | Reference |
|---|---|---|---|---|
| In vitro studies | ||||
| In vitro: simulated digestion and fluorescence in situ hybridization (FISH) | Healthy volunteers | Acerola by-product (M. emarginata D.C.) | ND | [107] |
| Simulated digestion and fluorescence in situ hybridization (FISH) | Healthy volunteers | Açai fruit pulp (Euterpe oleracea). | ND | [87] |
| The Simulator of the Human Intestinal Microbial Ecosystem (SHIME®) | Healthy male volunteers | Milk fermented by Lacticaseibacillus casei SJRP38 (2%), Lactiplantibacillus plantarum ST8ShStr (2%), and Streptococcus thermophilus TA 080 (0.1%) with 1% (w/v) of passion fruit (Passiflora edulis F.) or buriti fruit (Mauritia flexuosa) pulp | 80 g for 5 days | [48] |
| The Simulator of the Human Intestinal Microbial Ecosystem (SHIME®) | Obese volunteer (body mass index between 30 and 39.9 kg/m2 and waist circumference > 80 cm) | Low-fat goat milk fermented by L. casei Lc-1 and S. thermophilus TA040 with passion fruit by-product (1%, w/v, seeds, peels and pomace) | 400 mL | [44] |
| Simulated digestion and fluorescence in situ hybridization (FISH) | Healthy volunteers | Freeze-dried juçara fruit (Euterpe edulis) pulp | 1% | [91] |
| Simulated digestion and fluorescence in situ hybridization (FISH) | Healthy volunteers | Jabuticaba (Myrciaria jaboticaba) by-product | 20% | [17] |
| Simulated digestion and fluorescence in situ hybridization (FISH) | Healthy volunteers | Guava (Psidium guajava L.) and acerola (Malpighia glabra L.) by-products | 20% | [16] |
| In vitro: Cell-based cytotoxicity evaluation, erythrocyte protection In vivo: Preclinical study | 1,2 dimethyl hydrazine-induced colon carcinogenesis in male Wistar rats | Yogurt with (0.4%) or without jabuticaba (Myrciaria jaboticaba) seed extract. | 10 mL/kg | [90] |
| Preclinical studies | ||||
| Animal model | Dyslipidaemic diet in female Wistar rats | Guava (Psidium guajava L.) by-product | 800 mg/kg/day for 28 days | [15] |
| Animal model | Male db/db mice (diabetes and obesity) | Aqueous extract of guava (Psidium guajava L.) leaf | 7.0 g/kg for 12 weeks | [33] |
| Animal model | 2,4,6-trinitrobenzene sulfonic acid-induced colitis in Wistar rats | Extract of jabuticaba (Myrciaria jaboticaba) peel Extract was offered instead of water | From 141.1 ± 16.0 to 215.1 ± 31 mg GAE/kg | [79] |
| Animal model | 1,2-dimethyl hydrazine-inducedcolon carcinogenesis in Wistar rats | Yogurt with (0.4%) or without jabuticaba (Myrciaria jaboticaba) seed extract | 10 mL/kg | [106] |
| Animal model | Male C57BL/6 mice | Freeze-dried jabuticaba (Plinia cauliflora (Mart.) Kausel) peel | 4% (3.28 g/kg of experimental diet) | [55] |
| Animal model | C57BL/6 mice | Aqueous polysaccharides guava (Psidium guajava) extract | 100 mg/kg for 11 weeks | [56] |
| Animal model | Mice diarrhea induced by Folium Sennae | Aqueous Psidium guajava extract | 0.05 mL/10 g | [108] |
| Animal model | Male C57BL/6 mice | Jabuticaba (Myrciaria jaboticaba) seed and peel | 5%, 10%, and 15% of diet | [29] |
| Animal model | Male C57BL/6J mice | Anthocyanin-rich extract of açaí (Euterpe oleracea) | 150 mg/kg for 4 weeks | [54] |
| Clinical study | ||||
| Double-blind randomized controlled trial | Adults (31 to 59 years) with obesity [body mass index (BMI) ≥ 30 ≤ 39.9 kg/m2] | Freeze-dried juçara fruit pulp (Euterpe edulis) | 5 g for 42 days | [10] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, M.d.C.; do Nascimento, H.M.A.; da Silva, J.Y.P.; de Brito Alves, J.L.; de Souza, E.L. Evidence for the Beneficial Effects of Brazilian Native Fruits and Their By-Products on Human Intestinal Microbiota and Repercussions on Non-Communicable Chronic Diseases—A Review. Foods 2023, 12, 3491. https://doi.org/10.3390/foods12183491
Lima MdC, do Nascimento HMA, da Silva JYP, de Brito Alves JL, de Souza EL. Evidence for the Beneficial Effects of Brazilian Native Fruits and Their By-Products on Human Intestinal Microbiota and Repercussions on Non-Communicable Chronic Diseases—A Review. Foods. 2023; 12(18):3491. https://doi.org/10.3390/foods12183491
Chicago/Turabian StyleLima, Maiara da Costa, Heloísa Maria Almeida do Nascimento, Jaielison Yandro Pereira da Silva, José Luiz de Brito Alves, and Evandro Leite de Souza. 2023. "Evidence for the Beneficial Effects of Brazilian Native Fruits and Their By-Products on Human Intestinal Microbiota and Repercussions on Non-Communicable Chronic Diseases—A Review" Foods 12, no. 18: 3491. https://doi.org/10.3390/foods12183491
APA StyleLima, M. d. C., do Nascimento, H. M. A., da Silva, J. Y. P., de Brito Alves, J. L., & de Souza, E. L. (2023). Evidence for the Beneficial Effects of Brazilian Native Fruits and Their By-Products on Human Intestinal Microbiota and Repercussions on Non-Communicable Chronic Diseases—A Review. Foods, 12(18), 3491. https://doi.org/10.3390/foods12183491