
Effects of Fermentation on Bioactivity and the Composition of Polyphenols Contained in Polyphenol-Rich Foods: A Review

 ,
, 

Abstract
: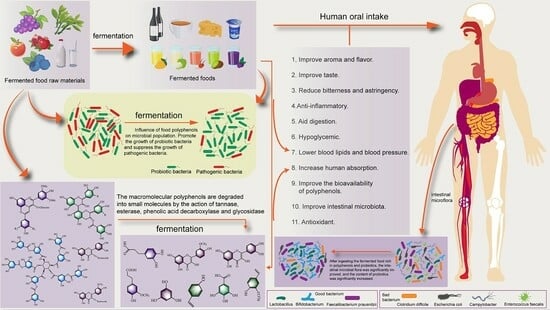
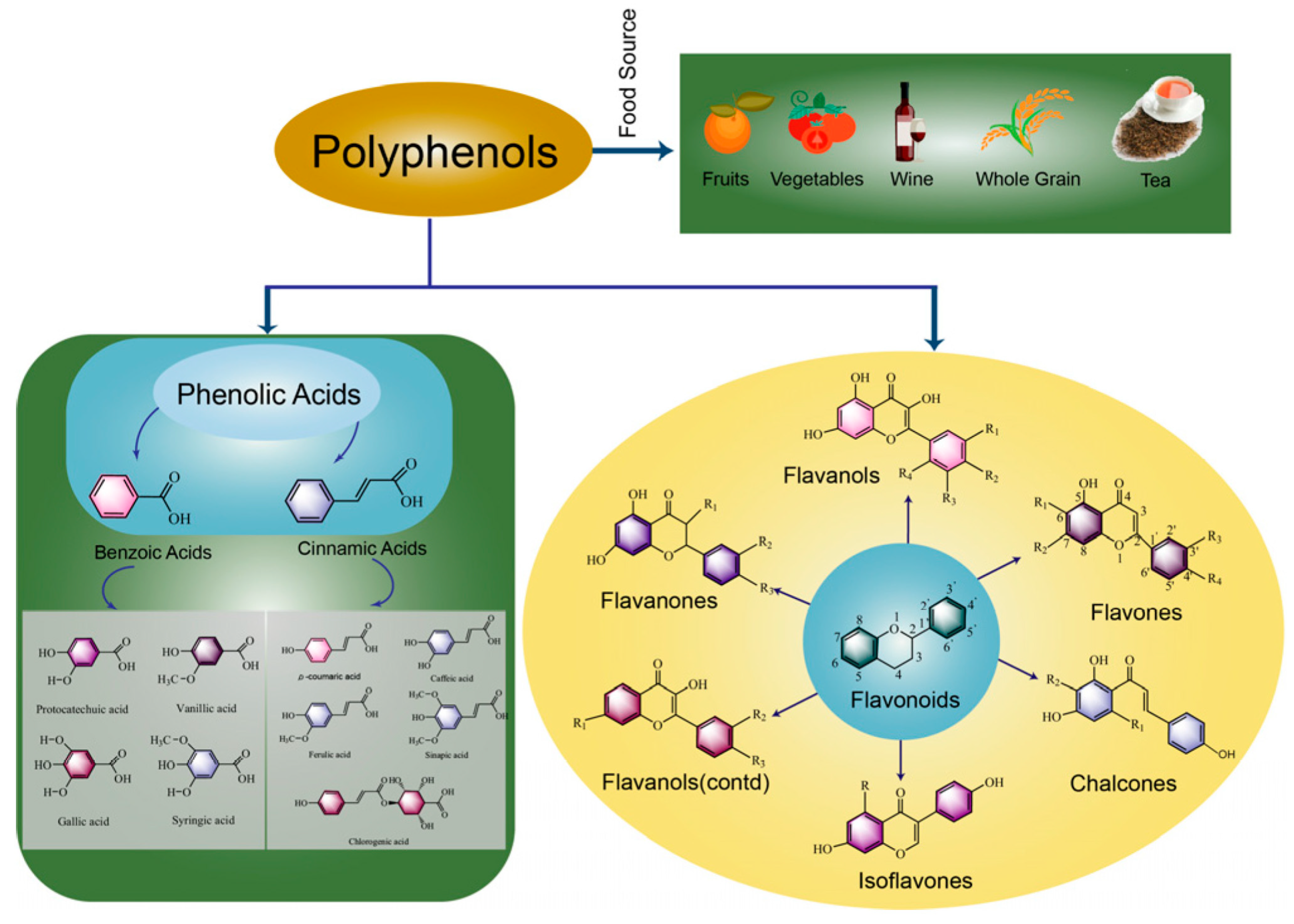
1. Introduction
2. Effect of Microorganisms on Polyphenol Metabolism during PFF Fermentation
2.1. Changes of Functional Activity and Polyphenols in PFFs after Fermentation
2.2. PAEs Secreted by Microorganisms during Fermentation
2.2.1. Tannase
2.2.2. Esterase
2.2.3. Phenolic acid Decarboxylase
2.2.4. Glucosidase
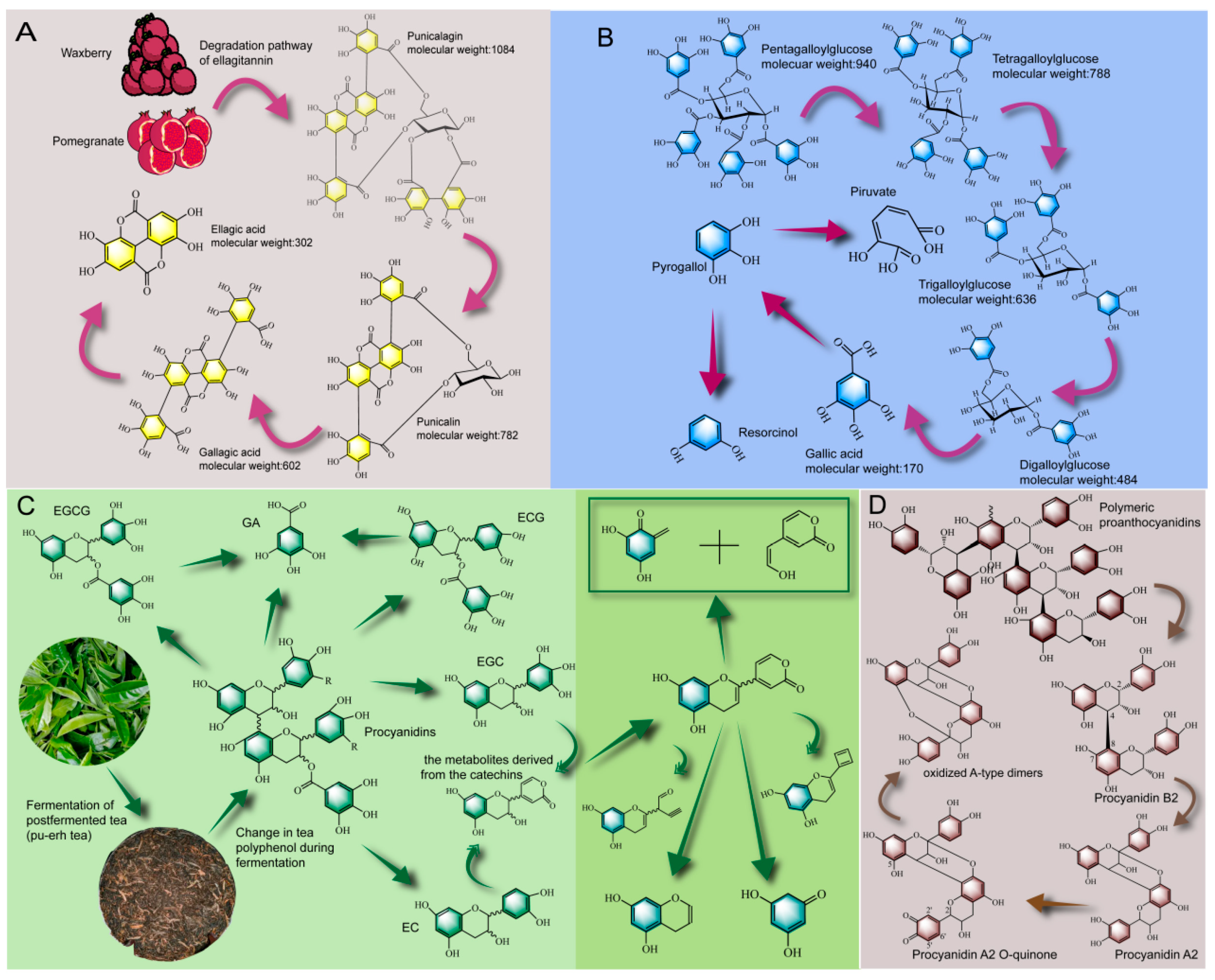
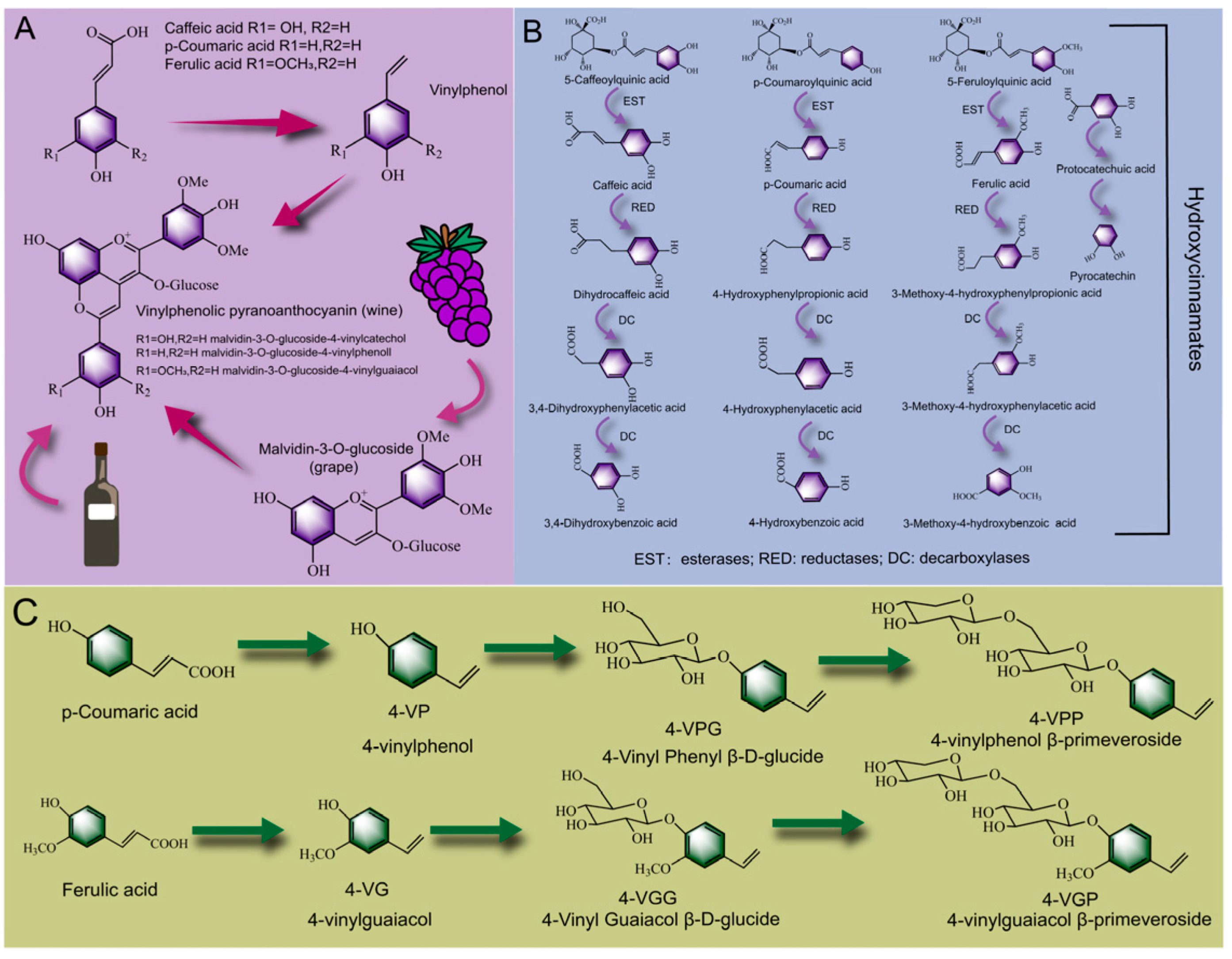
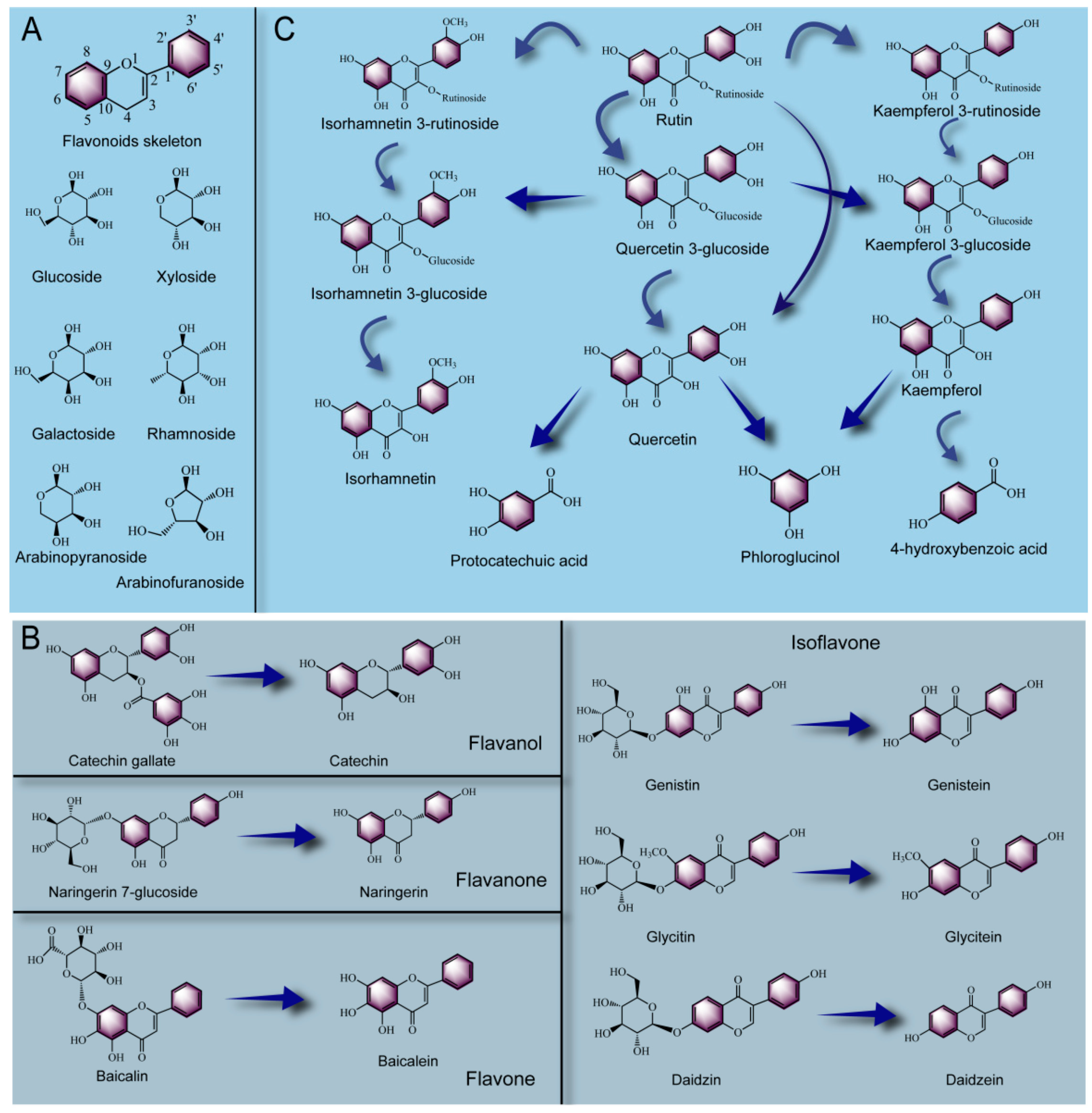
2.3. Biotransformation Pathways of Polyphenols during PFF Fermentation
2.3.1. Biotransformation Pathway of Tannin
2.3.2. Biotransformation Pathway of Phenolic Acid
2.3.3. Biotransformation Pathway of Flavonoids
3. Effects of Polyphenols Contained in PFFs on Microorganism
3.1. Effects of Polyphenols on Microbial Growth in PFF Fermentation Process
3.2. Effects of PFFs on Human Intestinal Microorganisms
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| PFFs | Polyphenol-rich fermented foods |
| PAEs | Polyphenol-associated enzymes |
| 4-VP | 4-Vinylphenol |
| 4-VPG | 4-Vinyl Phenyl β-D-glucide |
| 4-VPP | 4-Vinylphenol β-primeveroside |
| 4-VG | 4-Vinylguaiacol |
| 4-VGG | 4-Vinyl Guaiacol β-D-glucide |
| 4-VGP | 4-Vinylguaiacol β-primeveroside |
| PAD | Phenolic acid decarboxylase |
| PPE | Pomegranate peel extracts |
| FE | Feruloyl esterase |
| PACs | Proanthocyanidins |
References
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health benefits of fermented foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M. The periodic table of fermented foods: Limitations and opportunities. Appl. Microbiol. Biotechnol. 2022, 106, 2815–2826. [Google Scholar] [CrossRef] [PubMed]
- Marti-Quijal, F.J.; Remize, F.; Meca, G.; Ferrer, E.; Ruiz, M.-J.; Barba, F.J. Fermentation in fish and by-products processing: An overview of current research and future prospects. Curr. Opin. Food Sci. 2020, 31, 9–16. [Google Scholar] [CrossRef]
- Long, X.-S.; Liao, S.-T.; Li, E.-N.; Pang, D.-R.; Li, Q.; Liu, S.-C.; Hu, T.-G.; Zou, Y.-X. The hypoglycemic effect of freeze-dried fermented mulberry mixed with soybean on type 2 diabetes mellitus. Food Sci. Nutr. 2021, 9, 3641–3654. [Google Scholar] [CrossRef] [PubMed]
- Tonolo, F.; Moretto, L.; Folda, A.; Scalcon, V.; Bindoli, A.; Bellamio, M.; Feller, E.; Rigobello, M.P. Antioxidant Properties of Fermented Soy during Shelf Life. Plant Foods Hum. Nutr. 2019, 74, 287–292. [Google Scholar] [CrossRef]
- Yue, Q.; Wang, Z.; Tang, X.; Zhao, C.; Li, K.; Su, L.; Zhang, S.; Sun, X.; Liu, X.; Zhao, L. Hypolipidemic Effects of Fermented Seaweed Extracts by Saccharomyces cerevisiae and Lactiplantibacillus plantarum. Front. Microbiol. 2021, 12, 772585. [Google Scholar] [CrossRef]
- Chen, L.; Wang, L.; Shu, G.; Yuan, J.; Zhang, J.; Qin, S.; Li, J. Enhanced antihypertensive potential of fermented pomegranate juice: The contribution of phenolic compounds biotransformation and the resultant angiotensin-I-converting enzyme inhibition mechanism. Food Chem. 2023, 404, 134745. [Google Scholar] [CrossRef]
- Yu, C.; Liu, Y.; Xuemei, Z.; Ma, A.; Jianxin, T.; Yiling, T. Fermented Carrot Pulp Regulates the Dysfunction of Murine Intestinal Microbiota. Oxidative Med. Cell. Longev. 2022, 2022, 2479956. [Google Scholar] [CrossRef]
- Cai, W.; Tang, F.; Zhao, X.; Guo, Z.; Zhang, Z.; Dong, Y.; Shan, C. Different lactic acid bacteria strains affecting the flavor profile of fermented jujube juice. J. Food Process. Preserv. 2019, 43, e14095. [Google Scholar] [CrossRef]
- Ashokbhai, J.K.; Basaiawmoit, B.; Sakure, A.; Das, S.; Patil, G.B.; Mankad, M.; Hati, S. Purification and characterization of antioxidative and antimicrobial peptides from lactic-fermented sheep milk. J. Food Sci. Technol. 2022, 59, 4262–4272. [Google Scholar] [CrossRef]
- Taha, S.; El Abd, M.; De Gobba, C.; Abdel-Hamid, M.; Khalil, E.; Hassan, D. Antioxidant and antibacterial activities of bioactive peptides in buffalo’s yoghurt fermented with different starter cultures. Food Sci. Biotechnol. 2017, 26, 1325–1332. [Google Scholar] [CrossRef]
- Yu, Y.; Wu, J.; Xu, Y.; Xiao, G.; Zou, B. The effect of litchi juice on exopolysaccharide production in milk fermented by Lactobacillus casei. Int. J. Food Sci. Technol. 2018, 53, 2730–2737. [Google Scholar] [CrossRef]
- Li, J.; Wu, H.; Liu, Y.; Nan, J.; Park, H.J.; Chen, Y.; Yang, L. The chemical structure and immunomodulatory activity of an exopolysaccharide produced by Morchella esculenta under submerged fermentation. Food Funct. 2021, 12, 9327–9338. [Google Scholar] [CrossRef] [PubMed]
- Shiferaw Terefe, N.; Augustin, M.A. Fermentation for tailoring the technological and health related functionality of food products. Crit. Rev. Food Sci. Nutr. 2020, 60, 2887–2913. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2017, 20, 1689–1699. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Alves-Santos, A.M.; Sugizaki, C.S.A.; Lima, G.C.; Naves, M.M.V. Prebiotic effect of dietary polyphenols: A systematic review. J. Funct. Foods 2020, 74, 104169. [Google Scholar] [CrossRef]
- Rodríguez-Daza, M.C.; Pulido-Mateos, E.C.; Lupien-Meilleur, J.; Guyonnet, D.; Desjardins, Y.; Roy, D. Polyphenol-Mediated Gut Microbiota Modulation: Toward Prebiotics and Further. Front. Nutr. 2021, 8, 689456. [Google Scholar] [CrossRef]
- Garcia-Alonso, A.; Sánchez-Paniagua López, M.; Manzanares-Palenzuela, C.L.; Redondo-Cuenca, A.; López-Ruíz, B. Edible plant by-products as source of polyphenols: Prebiotic effect and analytical methods. Crit. Rev. Food Sci. Nutr. 2022, 1–22. [Google Scholar] [CrossRef]
- Estrela, J.M.; Mena, S.; Obrador, E.; Benlloch, M.; Castellano, G.; Salvador, R.; Dellinger, R.W. Polyphenolic Phytochemicals in Cancer Prevention and Therapy: Bioavailability versus Bioefficacy. J. Med. Chem. 2017, 60, 9413–9436. [Google Scholar] [CrossRef]
- Siemińska-Kuczer, A.; Szymańska-Chargot, M.; Zdunek, A. Recent advances in interactions between polyphenols and plant cell wall polysaccharides as studied using an adsorption technique. Food Chem. 2022, 373, 131487. [Google Scholar] [CrossRef] [PubMed]
- Castellone, V.; Bancalari, E.; Rubert, J.; Gatti, M.; Neviani, E.; Bottari, B. Eating Fermented: Health Benefits of LAB-Fermented Foods. Foods 2021, 10, 2639. [Google Scholar] [CrossRef]
- Díaz-Mula, H.M.; Tomás-Barberán, F.A.; García-Villalba, R. Pomegranate Fruit and Juice (cv. Mollar), Rich in Ellagitannins and Anthocyanins, Also Provide a Significant Content of a Wide Range of Proanthocyanidins. J. Agric. Food Chem. 2019, 67, 9160–9167. [Google Scholar] [CrossRef] [PubMed]
- Lukšič, L.; Bonafaccia, G.; Timoracka, M.; Vollmannova, A.; Trček, J.; Nyambe, T.K.; Melini, V.; Acquistucci, R.; Germ, M.; Kreft, I. Rutin and quercetin transformation during preparation of buckwheat sourdough bread. J. Cereal Sci. 2016, 69, 71–76. [Google Scholar] [CrossRef]
- Li, S.; Jin, Z.; Hu, D.; Yang, W.; Yan, Y.; Nie, X.; Lin, J.; Zhang, Q.; Gai, D.; Ji, Y.; et al. Effect of solid-state fermentation with Lactobacillus casei on the nutritional value, isoflavones, phenolic acids and antioxidant activity of whole soybean flour. LWT 2020, 125, 109264. [Google Scholar] [CrossRef]
- Nie, Q.; Feng, L.; Hu, J.; Wang, S.; Chen, H.; Huang, X.; Nie, S.; Xiong, T.; Xie, M. Effect of fermentation and sterilization on anthocyanins in blueberry. J. Sci. Food Agric. 2017, 97, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Kim, Y.; Lee, J. Comparison of antioxidant and anti-inflammatory activity of quercetin, isoquercitrin and rutin against alcohol-induced liver injury in HepG2 Cells. FASEB J. 2018, 32, 670.60. [Google Scholar] [CrossRef]
- Ren, Y.; Wu, S.; Xia, Y.; Huang, J.; Ye, J.; Xuan, Z.; Li, P.; Du, B. Probiotic-fermented black tartary buckwheat alleviates hyperlipidemia and gut microbiota dysbiosis in rats fed with a high-fat diet. Food Funct. 2021, 12, 6045–6057. [Google Scholar] [CrossRef]
- Ren, W.; Ma, Y.; Liu, D.; Liang, P.; Du, J.; Yang, S.; Tang, L.; Wu, Y. Chemical composition analysis, antioxidant activity, and target cell-based screening of the potential active components in jujube and its fermented product. J. Food Sci. 2022, 87, 664–685. [Google Scholar] [CrossRef]
- Xiao, Y.; Fan, J.; Chen, Y.; Rui, X.; Zhang, Q.; Dong, M. Enhanced total phenolic and isoflavone aglycone content, antioxidant activity and DNA damage protection of soybeans processed by solid state fermentation with Rhizopus oligosporus RT-3. RSC Adv. 2016, 6, 29741–29756. [Google Scholar] [CrossRef]
- Liu, M.; Xie, H.; Ma, Y.; Li, H.; Li, C.; Chen, L.; Jiang, B.; Nian, B.; Guo, T.; Zhang, Z.; et al. High Performance Liquid Chromatography and Metabolomics Analysis of Tannase Metabolism of Gallic Acid and Gallates in Tea Leaves. J. Agric. Food Chem. 2020, 68, 4946–4954. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, S.; Yu, Q.; Wang, J.; Deng, Y.; Hua, J.; Zhou, Q.; Yuan, H.; Jiang, Y. Chemical profile of a novel ripened Pu-erh tea and its metabolic conversion during pile fermentation. Food Chem. 2022, 378, 132126. [Google Scholar] [CrossRef] [PubMed]
- D’Antuono, I.; Bruno, A.; Linsalata, V.; Minervini, F.; Garbetta, A.; Tufariello, M.; Mita, G.; Logrieco, A.F.; Bleve, G.; Cardinali, A. Fermented Apulian table olives: Effect of selected microbial starters on polyphenols composition, antioxidant activities and bioaccessibility. Food Chem. 2018, 248, 137–145. [Google Scholar] [CrossRef]
- Zulkawi, N.; Ng, K.H.; Zamberi, R.; Yeap, S.K.; Jaganath, I.B.; Satharasinghe, D.; Yeah Yong, C.; Jamaluddin, A.B.; Tan, S.W.; Ho, W.Y.; et al. The in vivo hepato-recovery effects of the polyphenol-rich fermented food Xeniji™ on ethanol-induced liver damage. RSC Adv. 2017, 7, 38287–38299. [Google Scholar] [CrossRef]
- Chan, C.-L.; Gan, R.-y.; Shah, N.P.; Corke, H. Enhancing antioxidant capacity of Lactobacillus acidophilus-fermented milk fortified with pomegranate peel extracts. Food Biosci. 2018, 26, 185–192. [Google Scholar] [CrossRef]
- Ma, S.; Zhang, H.; Xu, J. Characterization, Antioxidant and Anti-Inflammation Capacities of Fermented Flammulina velutipes Polyphenols. Molecules 2021, 26, 6205. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Yu, H.; Qi, M.; Yuan, X.; Ruan, Z.; Hu, C.; Xiao, M.; Xue, Y.; Yao, Y.; Liu, Q. Biotransformation of citrus fruits phenolic profiles by mixed probiotics in vitro anaerobic fermentation. LWT 2022, 160, 113087. [Google Scholar] [CrossRef]
- Gao, Q.; Song, Y.; Liang, Y.; Li, Y.; Chang, Y.; Ma, R.; Cao, X.; Wang, S. Dynamics of Physicochemical Properties, Functional Compounds and Antioxidant Capacity during Spontaneous Fermentation of Lycium ruthenicum Murr. (Qinghai–Tibet Plateau) Natural Vinegar. Foods 2022, 11, 1344. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Chen, J.; Li, X.-L.; Yi, K.; Ye, Y.; Liu, G.; Wang, S.-F.; Hu, H.-L.; Zou, L.; Wang, Z.-G. Dynamic changes in antioxidant activity and biochemical composition of tartary buckwheat leaves during Aspergillus niger fermentation. J. Funct. Foods 2017, 32, 375–381. [Google Scholar] [CrossRef]
- Huang, Z.; Zhao, M.; Cui, C.; Huang, M.; Lin, L.; Feng, Y. A new sight on soy isoflavones during the whole soy sauce fermentation process by UPLC-MS/MS. LWT 2021, 152, 112249. [Google Scholar] [CrossRef]
- Piao, Y.-Z.; Eun, J.-B. Physicochemical characteristics and isoflavones content during manufacture of short-time fermented soybean product (cheonggukjang). J. Food Sci. Technol. 2020, 57, 2190–2197. [Google Scholar] [CrossRef]
- Choi, I.; Kim, Y.; Park, Y.; Seog, H.; Choi, H. Anti-obesity activities of fermented soygerm isoflavones by Bif idobacterium breve. BioFactors 2007, 29, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Pan, M.; El-Nezami, H.S.; Wan, J.M.F.; Wang, M.F.; Habimana, O.; Lee, J.C.Y.; Louie, J.C.Y.; Shah, N.P. Effects of Lactic Acid Bacteria-Fermented Soymilk on Isoflavone Metabolites and Short-Chain Fatty Acids Excretion and Their Modulating Effects on Gut Microbiota. J. Food Sci. 2019, 84, 1854–1863. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tao, Y.; Li, D.; Wen, G.; Zhou, J.; Manickam, S.; Han, Y.; Chai, W.S. Fermentation of blueberry juices using autochthonous lactic acid bacteria isolated from fruit environment: Fermentation characteristics and evolution of phenolic profiles. Chemosphere 2021, 276, 130090. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Wang, J.; Wang, Y.; Xu, Z.; Li, B.; Meng, X.; Sun, X.; Zhu, J. Influence of fermentation by lactic acid bacteria and in vitro digestion on the biotransformations of blueberry juice phenolics. Food Control. 2022, 133, 108603. [Google Scholar] [CrossRef]
- Jia, B.; Wei, Z.; Kong, X.; Xia, S.; Gan, L.; Han, S. Antioxidant Properties of Larch Tannins with Different Mean Polymerization Degrees: Controlled Degradation Based on Hydroxyl Radical Degradation. J. Agric. Food Chem. 2022, 70, 9367–9376. [Google Scholar] [CrossRef]
- Zhong, H.; Xue, Y.; Lu, X.; Shao, Q.; Cao, Y.; Wu, Z.; Chen, G. The Effects of Different Degrees of Procyanidin Polymerization on the Nutrient Absorption and Digestive Enzyme Activity in Mice. Molecules 2018, 23, 2916. [Google Scholar] [CrossRef]
- Wu, M.; Wang, Q.; McKinstry, W.J.; Ren, B. Characterization of a tannin acyl hydrolase from Streptomyces sviceus with substrate preference for digalloyl ester bonds. Appl. Microbiol. Biotechnol. 2015, 99, 2663–2672. [Google Scholar] [CrossRef]
- Aguilar, C.N.; Rodríguez, R.; Gutiérrez-Sánchez, G.; Augur, C.; Favela-Torres, E.; Prado-Barragan, L.A.; Ramírez-Coronel, A.; Contreras-Esquivel, J.C. Microbial tannases: Advances and perspectives. Appl. Microbiol. Biotechnol. 2007, 76, 47–59. [Google Scholar] [CrossRef]
- Lekshmi, R.; Arif Nisha, S.; Thirumalai Vasan, P.; Kaleeswaran, B. A comprehensive review on tannase: Microbes associated production of tannase exploiting tannin rich agro-industrial wastes with special reference to its potential environmental and industrial applications. Environ. Res. 2021, 201, 111625. [Google Scholar] [CrossRef]
- de la Iglesia, R.; Milagro, F.I.; Campión, J.; Boqué, N.; Martínez, J.A. Healthy properties of proanthocyanidins. BioFactors 2010, 36, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-X.; Feng, H.-L.; Ding, Y.-M.; Chai, W.-M.; Xiang, Z.-H.; Shi, Y.; Chen, Q.-X. Structure characterization of proanthocyanidins from Caryota ochlandra Hance and their bioactivities. Food Chem. 2014, 155, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Mo, M.; Zhang, K.; Bi, Y.; Kong, F. Preparation, characterization and biological activity of proanthocyanidin-chitosan nanoparticles. Int. J. Biol. Macromol. 2021, 188, 43–51. [Google Scholar] [CrossRef]
- Fan, W.; Zong, H.; Zhao, T.; Deng, J.; Yang, H. Bioactivities and mechanisms of dietary proanthocyanidins on blood pressure lowering: A critical review of in vivo and clinical studies. Crit. Rev. Food Sci. Nutr. 2022, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, Y.; Li, D.; Ho, C.-T.; Li, J.; Wan, X. The absorption, distribution, metabolism and excretion of procyanidins. Food Funct. 2016, 7, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Saura-Calixto, F.; Pérez-Jiménez, J.; Touriño, S.; Serrano, J.; Fuguet, E.; Torres, J.L.; Goñi, I. Proanthocyanidin metabolites associated with dietary fibre from in vitro colonic fermentation and proanthocyanidin metabolites in human plasma. Mol. Nutr. Food Res. 2010, 54, 939–946. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef]
- Higdon, J.V.; Frei, B. Tea Catechins and Polyphenols: Health Effects, Metabolism, and Antioxidant Functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef]
- Bertram, B.; Bollow, U.; Rajaee-Behbahani, N.; Bürkle, A.; Schmezer, P. Induction of poly(ADP-ribosyl)ation and DNA damage in human peripheral lymphocytes after treatment with (−)-epigallocatechin-gallate. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2003, 534, 77–84. [Google Scholar] [CrossRef]
- Malik, A.; Azam, S.; Hadi, N.; Hadi, S.M. DNA degradation by water extract of green tea in the presence of copper ions: Implications for anticancer properties. Phytother. Res. 2003, 17, 358–363. [Google Scholar] [CrossRef]
- Macedo, J.A.; Ferreira, L.R.; Camara, L.E.; Santos, J.C.; Gambero, A.; Macedo, G.A.; Ribeiro, M.L. Chemopreventive potential of the tannase-mediated biotransformation of green tea. Food Chem. 2012, 133, 358–365. [Google Scholar] [CrossRef]
- Govindarajan, R.K.; Khanongnuch, C.; Mathivanan, K.; Shyu, D.J.H.; Sharma, K.P.; De Mandal, S. In-vitro biotransformation of tea using tannase produced by Enterobacter cloacae 41. J. Food Sci. Technol. 2021, 58, 3235–3242. [Google Scholar] [CrossRef] [PubMed]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent Research on the Health Benefits of Blueberries and Their Anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Ju, Y.; Ma, T.; Zhang, J.; Fang, Y.; Sun, X. New perspectives on the biosynthesis, transportation, astringency perception and detection methods of grape proanthocyanidins. Crit. Rev. Food Sci. Nutr. 2021, 61, 2372–2398. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Wollmann, N.; Schieberle, P.; Hofmann, T. Reconstitution of the Flavor Signature of Dornfelder Red Wine on the Basis of the Natural Concentrations of Its Key Aroma and Taste Compounds. J. Agric. Food Chem. 2011, 59, 8866–8874. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Brandão, E.; Mateus, N.; de Freitas, V. Sensorial properties of red wine polyphenols: Astringency and bitterness. Crit. Rev. Food Sci. Nutr. 2017, 57, 937–948. [Google Scholar] [CrossRef]
- Ramos-Pineda, A.M.; García-Estévez, I.; Brás, N.F.; Martín del Valle, E.M.; Dueñas, M.; Escribano Bailón, M.T. Molecular Approach to the Synergistic Effect on Astringency Elicited by Mixtures of Flavanols. J. Agric. Food Chem. 2017, 65, 6425–6433. [Google Scholar] [CrossRef]
- Morzel, M.; Canon, F.; Guyot, S. Interactions between Salivary Proteins and Dietary Polyphenols: Potential Consequences on Gastrointestinal Digestive Events. J. Agric. Food Chem. 2022, 70, 6317–6327. [Google Scholar] [CrossRef]
- Kyraleou, M.; Kotseridis, Y.; Koundouras, S.; Chira, K.; Teissedre, P.-L.; Kallithraka, S. Effect of irrigation regime on perceived astringency and proanthocyanidin composition of skins and seeds of Vitis vinifera L. cv. Syrah grapes under semiarid conditions. Food Chem. 2016, 203, 292–300. [Google Scholar] [CrossRef]
- Sun, B.; Sá, M.d.; Leandro, C.; Caldeira, I.; Duarte, F.L.; Spranger, I. Reactivity of Polymeric Proanthocyanidins toward Salivary Proteins and Their Contribution to Young Red Wine Astringency. J. Agric. Food Chem. 2013, 61, 939–946. [Google Scholar] [CrossRef]
- McRae, J.M.; Schulkin, A.; Kassara, S.; Holt, H.E.; Smith, P.A. Sensory Properties of Wine Tannin Fractions: Implications for In-Mouth Sensory Properties. J. Agric. Food Chem. 2013, 61, 719–727. [Google Scholar] [CrossRef] [PubMed]
- González-Royo, E.; Esteruelas, M.; Kontoudakis, N.; Fort, F.; Canals, J.M.; Zamora, F. The effect of supplementation with three commercial inactive dry yeasts on the colour, phenolic compounds, polysaccharides and astringency of a model wine solution and red wine. J. Sci. Food Agric. 2017, 97, 172–181. [Google Scholar] [CrossRef]
- Gao, Y.; Guo, X.; Liu, Y.; Zhang, M.; Zhang, R.; Abbasi, A.M.; You, L.; Li, T.; Liu, R.H. Comparative assessment of phytochemical profile, antioxidant capacity and anti-proliferative activity in different varieties of brown rice (Oryza sativa L.). LWT 2018, 96, 19–25. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J. Insoluble-Bound Phenolics in Food. Molecules 2016, 21, 1216. [Google Scholar] [CrossRef] [PubMed]
- Schär, A.; Sprecher, I.; Topakas, E.; Faulds, C.B.; Nyström, L. Hydrolysis of Nonpolar n-Alkyl Ferulates by Feruloyl Esterases. J. Agric. Food Chem. 2016, 64, 8549–8554. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Soomro, L.; Wei, X.; Yuan, X.; Gu, T.; Li, Z.; Wang, Y.; Bao, Y.; Wang, F.; Wen, B.; et al. Directed evolution of feruloyl esterase from Lactobacillus acidophilus and its application for ferulic acid production. Bioresour. Technol. 2021, 332, 124967. [Google Scholar] [CrossRef]
- Mancuso, C.; Santangelo, R. Ferulic acid: Pharmacological and toxicological aspects. Food Chem. Toxicol. 2014, 65, 185–195. [Google Scholar] [CrossRef]
- Smit, A.; Cordero Otero, R.R.; Lambrechts, M.G.; Pretorius, I.S.; van Rensburg, P. Enhancing Volatile Phenol Concentrations in Wine by Expressing Various Phenolic Acid Decarboxylase Genes in Saccharomyces cerevisiae. J. Agric. Food Chem. 2003, 51, 4909–4915. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, Z.; Li, Y.; Feng, Z.; Zhang, X.; Zhou, R.; Liu, C.; Yang, L. The Combined Cultivation of Feruloyl Esterase-Producing Strains with CMCase and Xylanase-Producing Strains Increases the Release of Ferulic Acid. Microorganisms 2022, 10, 1889. [Google Scholar] [CrossRef]
- Kongor, J.E.; Hinneh, M.; de Walle, D.V.; Afoakwa, E.O.; Boeckx, P.; Dewettinck, K. Factors influencing quality variation in cocoa (Theobroma cacao) bean flavour profile—A review. Food Res. Int. 2016, 82, 44–52. [Google Scholar] [CrossRef]
- Melo, T.S.; Pires, T.C.; Engelmann, J.V.P.; Monteiro, A.L.O.; Maciel, L.F.; Bispo, E.d.S. Evaluation of the content of bioactive compounds in cocoa beans during the fermentation process. J. Food Sci. Technol. 2021, 58, 1947–1957. [Google Scholar] [CrossRef] [PubMed]
- de Vries, R.P.; vanKuyk, P.A.; Kester, H.C.M.; Visser, J. The Aspergillus niger faeB gene encodes a second feruloyl esterase involved in pectin and xylan degradation and is specifically induced in the presence of aromatic compounds. Biochem. J. 2002, 363, 377–386. [Google Scholar] [CrossRef]
- Crepin, V.F.; Faulds, C.B.; Connerton, I.F. A non-modular type B feruloyl esterase from Neurospora crassa exhibits concentration-dependent substrate inhibition. Biochem. J. 2003, 370, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Conesa, M.-T.; Crepin, V.F.; Goldson, A.J.; Williamson, G.; Cummings, N.J.; Connerton, I.F.; Faulds, C.B.; Kroon, P.A. The feruloyl esterase system of Talaromyces stipitatus: Production of three discrete feruloyl esterases, including a novel enzyme, TsFaeC, with a broad substrate specificity. J. Biotechnol. 2004, 108, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Moukouli, M.; Topakas, E.; Christakopoulos, P. Cloning, characterization and functional expression of an alkalitolerant type C feruloyl esterase from Fusarium oxysporum. Appl. Microbiol. Biotechnol. 2008, 79, 245–254. [Google Scholar] [CrossRef]
- Ferreira, L.M.; Wood, T.M.; Williamson, G.; Faulds, C.; Hazlewood, G.P.; Black, G.W.; Gilbert, H.J. A modular esterase from Pseudomonas fluorescens subsp. cellulosa contains a non-catalytic cellulose-binding domain. Biochem. J. 1993, 294, 349–355. [Google Scholar] [CrossRef]
- Sheng, X.; Lind, M.E.S.; Himo, F. Theoretical study of the reaction mechanism of phenolic acid decarboxylase. FEBS J. 2015, 282, 4703–4713. [Google Scholar] [CrossRef]
- Nyachoti, C.M.; Atkinson, J.L.; Leeson, S. Sorghum tannins: A review. World’s Poult. Sci. J. 1997, 53, 5–21. [Google Scholar] [CrossRef]
- Svensson, L.; Sekwati-Monang, B.; Lutz, D.L.; Schieber, A.; Gänzle, M.G. Phenolic Acids and Flavonoids in Nonfermented and Fermented Red Sorghum (Sorghum bicolor (L.) Moench). J. Agric. Food Chem. 2010, 58, 9214–9220. [Google Scholar] [CrossRef]
- Ripari, V.; Bai, Y.; Gänzle, M.G. Metabolism of phenolic acids in whole wheat and rye malt sourdoughs. Food Microbiol. 2019, 77, 43–51. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, H.; Cui, Y.; Zeng, D.; Bao, X. Increasing the level of 4-vinylguaiacol in top-fermented wheat beer by secretory expression of ferulic acid decarboxylase from Bacillus pumilus in brewer’s yeast. Biotechnol. Lett. 2020, 42, 2711–2720. [Google Scholar] [CrossRef] [PubMed]
- Rosimin, A.A.; Kim, K.-S. Production of volatile phenols by kimchi Lactobacillus plantarum isolates and factors influencing their phenolic acid decarboxylase gene expression profiles. Food Res. Int. 2015, 78, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Linke, D.; Riemer, S.J.L.; Schimanski, S.; Nieter, A.; Krings, U.; Berger, R.G. Cold generation of smoke flavour by the first phenolic acid decarboxylase from a filamentous ascomycete—Isaria farinosa. Fungal Biol. 2017, 121, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Kytidou, K.; Artola, M.; Overkleeft, H.S.; Aerts, J.M.F.G. Plant Glycosides and Glycosidases: A Treasure-Trove for Therapeutics. Front. Plant Sci. 2020, 11, 357. [Google Scholar] [CrossRef]
- Xiao, J.; Muzashvili, T.S.; Georgiev, M.I. Advances in the biotechnological glycosylation of valuable flavonoids. Biotechnol. Adv. 2014, 32, 1145–1156. [Google Scholar] [CrossRef]
- Johnson, J.B.; Mani, J.S.; Broszczak, D.; Prasad, S.S.; Ekanayake, C.P.; Strappe, P.; Valeris, P.; Naiker, M. Hitting the sweet spot: A systematic review of the bioactivity and health benefits of phenolic glycosides from medicinally used plants. Phytother. Res. 2021, 35, 3484–3508. [Google Scholar] [CrossRef]
- Goh, L.M.L.; Barlow, P.J. Flavonoid recovery and stability from Ginkgo biloba subjected to a simulated digestion process. Food Chem. 2004, 86, 195–202. [Google Scholar] [CrossRef]
- Zhang, X.; Thuong, P.T.; Min, B.-S.; Ngoc, T.M.; Hung, T.M.; Lee, I.S.; Na, M.; Seong, Y.-H.; Song, K.-S.; Bae, K. Phenolic Glycosides with Antioxidant Activity from the Stem Bark of Populus davidiana. J. Nat. Prod. 2006, 69, 1370–1373. [Google Scholar] [CrossRef]
- Plumb, G.W.; De Pascual-Teresa, S.; Santos-Buelga, C.; Cheynier, V.; Williamson, G. Antioxidant properties of catechins and proanthocyanidins: Effect of polymerisation, galloylation and glycosylation. Free. Radic. Res. 1998, 29, 351–358. [Google Scholar] [CrossRef]
- Xiao, J.; Capanoglu, E.; Jassbi, A.R.; Miron, A. Advance on the Flavonoid C-glycosides and Health Benefits. Crit. Rev. Food Sci. Nutr. 2016, 56, S29–S45. [Google Scholar] [CrossRef]
- Hostetler, G.; Riedl, K.; Cardenas, H.; Diosa-Toro, M.; Arango, D.; Schwartz, S.; Doseff, A.I. Flavone deglycosylation increases their anti-inflammatory activity and absorption. Mol. Nutr. Food Res. 2012, 56, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Li, R.; Ni, H.; Li, L.J.; Li, Q.B. The effects of α-L-rhamnosidase, β-D-glucosidase, and their combination on the quality of orange juice. J. Food Process. Preserv. 2021, 45, e15604. [Google Scholar] [CrossRef]
- Gu, Q.; Duan, G.; Yu, X. Bioconversion of Flavonoid Glycosides from Hippophae rhamnoides Leaves into Flavonoid Aglycones by Eurotium amstelodami. Microorganisms 2019, 7, 122. [Google Scholar] [CrossRef] [PubMed]
- Baffi, M.A.; Martin, N.; Tobal, T.M.; Ferrarezi, A.L.; Lago, J.H.G.; Boscolo, M.; Gomes, E.; Da-Silva, R. Purification and Characterization of an Ethanol-Tolerant β-Glucosidase from Sporidiobolus pararoseus and Its Potential for Hydrolysis of Wine Aroma Precursors. Appl. Biochem. Biotechnol. 2013, 171, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Zhu, Y.-W.; Jiang, Y.-W.; Li, H.-K.; Liu, Z.-M.; Wang, W.; Shan, C.-H.; Fu, Y.-J. Improvement of flavonoid aglycone and biological activity of mulberry leaves by solid-state fermentation. Ind. Crops Prod. 2020, 148, 112287. [Google Scholar] [CrossRef]
- Wang, L.; Luo, Y.; Wu, Y.; Liu, Y.; Wu, Z. Fermentation and complex enzyme hydrolysis for improving the total soluble phenolic contents, flavonoid aglycones contents and bio-activities of guava leaves tea. Food Chem. 2018, 264, 189–198. [Google Scholar] [CrossRef]
- Santos, A.C.d.; Biluca, F.C.; Braghini, F.; Gonzaga, L.V.; Costa, A.C.O.; Fett, R. Phenolic composition and biological activities of stingless bee honey: An overview based on its aglycone and glycoside compounds. Food Res. Int. 2021, 147, 110553. [Google Scholar] [CrossRef]
- Khosravi, A.; Razavi, S.H. Therapeutic effects of polyphenols in fermented soybean and black soybean products. J. Funct. Foods 2021, 81, 104467. [Google Scholar] [CrossRef]
- Lodha, D.; Das, S.; Hati, S. Antioxidant activity, total phenolic content and biotransformation of isoflavones during soy lactic-fermentations. J. Food Process. Preserv. 2021, 45, e15583. [Google Scholar] [CrossRef]
- Jian, X.; Zhang, J. Component and Structure of Aspergillus flavipes sp.-Biodegraded Bayberry Tannins: A Potential Routine for Condensed Tannin Cleaner Degradation and Disposal. ACS Omega 2022, 7, 5809–5816. [Google Scholar] [CrossRef]
- Ascacio-Valdés, J.A.; Aguilera-Carbó, A.F.; Buenrostro, J.J.; Prado-Barragán, A.; Rodríguez-Herrera, R.; Aguilar, C.N. The complete biodegradation pathway of ellagitannins by Aspergillus niger in solid-state fermentation. J. Basic Microbiol. 2016, 56, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.; Lee, R.; Hardy, M.; Heber, D. Rapid large scale purification of ellagitannins from pomegranate husk, a by-product of the commercial juice industry. Sep. Purif. Technol. 2005, 41, 49–55. [Google Scholar] [CrossRef]
- Valero-Cases, E.; Nuncio-Jáuregui, N.; Frutos, M.J. Influence of Fermentation with Different Lactic Acid Bacteria and in Vitro Digestion on the Biotransformation of Phenolic Compounds in Fermented Pomegranate Juices. J. Agric. Food Chem. 2017, 65, 6488–6496. [Google Scholar] [CrossRef] [PubMed]
- Chávez-González, M.L.; Guyot, S.; Rodríguez-Herrera, R.; Prado-Barragán, A.; Aguilar, C.N. Production profiles of phenolics from fungal tannic acid biodegradation in submerged and solid-state fermentation. Process Biochem. 2014, 49, 541–546. [Google Scholar] [CrossRef]
- Li, C.; Leverence, R.; Trombley, J.D.; Xu, S.; Yang, J.; Tian, Y.; Reed, J.D.; Hagerman, A.E. High Molecular Weight Persimmon (Diospyros kaki L.) Proanthocyanidin: A Highly Galloylated, A-Linked Tannin with an Unusual Flavonol Terminal Unit, Myricetin. J. Agric. Food Chem. 2010, 58, 9033–9042. [Google Scholar] [CrossRef]
- Poupard, P.; Sanoner, P.; Baron, A.; Renard, C.M.G.C.; Guyot, S. Characterization of procyanidin B2 oxidation products in an apple juice model solution and confirmation of their presence in apple juice by high-performance liquid chromatography coupled to electrospray ion trap mass spectrometry. J. Mass Spectrom. 2011, 46, 1186–1197. [Google Scholar] [CrossRef]
- Wen, K.-S.; Ruan, X.; Wang, J.; Yang, L.; Wei, F.; Zhao, Y.-X.; Wang, Q. Optimizing Nucleophilic Depolymerization of Proanthocyanidins in Grape Seeds to Dimeric Proanthocyanidin B1 or B2. J. Agric. Food Chem. 2019, 67, 5978–5988. [Google Scholar] [CrossRef]
- De Taeye, C.; Caullet, G.; Eyamo Evina, V.J.; Collin, S. Procyanidin A2 and Its Degradation Products in Raw, Fermented, and Roasted Cocoa. J. Agric. Food Chem. 2017, 65, 1715–1723. [Google Scholar] [CrossRef]
- Zuo, Y.; Chen, H.; Deng, Y. Simultaneous determination of catechins, caffeine and gallic acids in green, Oolong, black and pu-erh teas using HPLC with a photodiode array detector. Talanta 2002, 57, 307–316. [Google Scholar] [CrossRef]
- Zhao, Z.J.; Tong, H.R.; Zhou, L.; Wang, E.X.; Liu, Q.J. Fungal colonization of Pu-Erh tea in yunnan. J. Food Saf. 2010, 30, 769–784. [Google Scholar] [CrossRef]
- Qin, J.-H.; Li, N.; Tu, P.-F.; Ma, Z.-Z.; Zhang, L. Change in Tea Polyphenol and Purine Alkaloid Composition during Solid-State Fungal Fermentation of Postfermented Tea. J. Agric. Food Chem. 2012, 60, 1213–1217. [Google Scholar] [CrossRef] [PubMed]
- An, T.; Chen, M.; Zu, Z.; Chen, Q.; Lu, H.; Yue, P.; Gao, X. Untargeted and targeted metabolomics reveal changes in the chemical constituents of instant dark tea during liquid-state fermentation by Eurotium cristatum. Food Res. Int. 2021, 148, 110623. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; González, C.; Suárez-Lepe, J.A. Formation of vinylphenolic pyranoanthocyanins by selected yeasts fermenting red grape musts supplemented with hydroxycinnamic acids. Int. J. Food Microbiol. 2007, 116, 144–152. [Google Scholar] [CrossRef]
- Morata, A.; Vejarano, R.; Ridolfi, G.; Benito, S.; Palomero, F.; Uthurry, C.; Tesfaye, W.; González, C.; Suárez-Lepe, J.A. Reduction of 4-ethylphenol production in red wines using HCDC+ yeasts and cinnamyl esterases. Enzym. Microb. Technol. 2013, 52, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Vanbeneden, N.; Gils, F.; Delvaux, F.; Delvaux, F.R. Formation of 4-vinyl and 4-ethyl derivatives from hydroxycinnamic acids: Occurrence of volatile phenolic flavour compounds in beer and distribution of Pad1-activity among brewing yeasts. Food Chem. 2008, 107, 221–230. [Google Scholar] [CrossRef]
- Ricci, A.; Cirlini, M.; Calani, L.; Bernini, V.; Neviani, E.; Del Rio, D.; Galaverna, G.; Lazzi, C. In vitro metabolism of elderberry juice polyphenols by lactic acid bacteria. Food Chem. 2019, 276, 692–699. [Google Scholar] [CrossRef]
- Huang, J.; de Paulis, T.; May, J.M. Antioxidant effects of dihydrocaffeic acid in human EA.hy926 endothelial cells. J. Nutr. Biochem. 2004, 15, 722–729. [Google Scholar] [CrossRef]
- Sáyago-Ayerdi, S.G.; Venema, K.; Tabernero, M.; Sarriá, B.; Bravo, L.; Mateos, R. Bioconversion of polyphenols and organic acids by gut microbiota of predigested Hibiscus sabdariffa L. calyces and Agave (A. tequilana Weber) fructans assessed in a dynamic in vitro model (TIM-2) of the human colon. Food Res. Int. 2021, 143, 110301. [Google Scholar] [CrossRef]
- Cárdenas-Castro, A.P.; Venema, K.; Sarriá, B.; Bravo, L.; Sáyago-Ayerdi, S.G.; Mateos, R. Study of the impact of a dynamic in vitro model of the colon (TIM-2) in the phenolic composition of two Mexican sauces. Food Res. Int. 2021, 139, 109917. [Google Scholar] [CrossRef]
- Rodriguez, A.; Meadows, J.A.; Sun, N.; Simmons, B.A.; Gladden, J.M. Evaluation of bacterial hosts for conversion of lignin-derived p-coumaric acid to 4-vinylphenol. Microb. Cell Factories 2021, 20, 181. [Google Scholar] [CrossRef]
- Sun, L.-H.; Lv, S.-W.; Yu, F.; Li, S.-N.; He, L.-Y. Biosynthesis of 4-vinylguaiacol from crude ferulic acid by Bacillus licheniformis DLF-17056. J. Biotechnol. 2018, 281, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Kitaoka, N.; Nomura, T.; Ogita, S.; Kato, Y. Bioproduction of 4-Vinylphenol and 4-Vinylguaiacol β-Primeverosides Using Transformed Bamboo Cells Expressing Bacterial Phenolic Acid Decarboxylase. Appl. Biochem. Biotechnol. 2021, 193, 2061–2075. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, F.; De Paepe, B.; Maertens, J.; Beauprez, J.; De Cocker, P.; Mincke, S.; Stevens, C.; De Mey, M. Development of an in vivo glucosylation platform by coupling production to growth: Production of phenolic glucosides by a glycosyltransferase of Vitis vinifera. Biotechnol. Bioeng. 2015, 112, 1594–1603. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Fang, Z. Hydroxycinnamic acids on gut microbiota and health. Compr. Rev. Food Sci. Food Saf. 2021, 20, 710–737. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, N.; Mandal, A.K.A. Pharmacokinetic, toxicokinetic, and bioavailability studies of epigallocatechin-3-gallate loaded solid lipid nanoparticle in rat model. Drug Dev. Ind. Pharm. 2019, 45, 1506–1514. [Google Scholar] [CrossRef]
- Kim, H.-S.; Jeon, D.Y.; Javaid, H.M.A.; Sahar, N.E.; Lee, H.-N.; Hong, S.-J.; Huh, J.Y.; Kim, Y.-M. Bio-transformation of green tea infusion with tannase and its improvement on adipocyte metabolism. Enzym. Microb. Technol. 2020, 135, 109496. [Google Scholar] [CrossRef]
- Roberto, B.S.; Macedo, G.A.; Macedo, J.A.; Martins, I.M.; Nakajima, V.M.; Allwood, J.W.; Stewart, D.; McDougall, G.J. Immobilized tannase treatment alters polyphenolic composition in teas and their potential anti-obesity and hypoglycemic activities in vitro. Food Funct. 2016, 7, 3920–3932. [Google Scholar] [CrossRef]
- María Landete, J.; Hernández, T.; Robredo, S.; Dueñas, M.; de las Rivas, B.; Estrella, I.; Muñoz, R. Effect of soaking and fermentation on content of phenolic compounds of soybean (Glycine max cv. Merit) and mung beans (Vigna radiata [L] Wilczek). Int. J. Food Sci. Nutr. 2015, 66, 203–209. [Google Scholar] [CrossRef]
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi. (Lamiaceae): A review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J. Pharm. Pharmacol. 2019, 71, 1353–1369. [Google Scholar] [CrossRef]
- Xu, C.; Ji, G.E. Bioconversion of Flavones During Fermentation in Milk Containing Scutellaria baicalensis Extract by Lactobacillus brevis. J. Microbiol. Biotechnol. 2013, 23, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Izumi, T.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kikuchi, M.; Piskula, M.K.; Kubota, Y. Soy Isoflavone Aglycones Are Absorbed Faster and in Higher Amounts than Their Glucosides in Humans. J. Nutr. 2000, 130, 1695–1699. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.J.; Lim, B.; Kim, H.Y.; Kwon, S.-J.; Eom, S.H. Deglycosylation patterns of isoflavones in soybean extracts inoculated with two enzymatically different strains of lactobacillus species. Enzym. Microb. Technol. 2020, 132, 109394. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Chen, J.; Tang, H.; Wang, C.; Li, Z.; Xiao, Y. Bioprocessing of soybeans (Glycine max L.) by solid-state fermentation with Eurotium cristatum YL-1 improves total phenolic content, isoflavone aglycones, and antioxidant activity. RSC Adv. 2020, 10, 16928–16941. [Google Scholar] [CrossRef] [PubMed]
- Di Gioia, D.; Strahsburger, E.; Lopez de Lacey, A.M.; Bregola, V.; Marotti, I.; Aloisio, I.; Biavati, B.; Dinelli, G. Flavonoid bioconversion in Bifidobacterium pseudocatenulatum B7003: A potential probiotic strain for functional food development. J. Funct. Foods 2014, 7, 671–679. [Google Scholar] [CrossRef]
- Lin, S.; Zhu, Q.; Wen, L.; Yang, B.; Jiang, G.; Gao, H.; Chen, F.; Jiang, Y. Production of quercetin, kaempferol and their glycosidic derivatives from the aqueous-organic extracted residue of litchi pericarp with Aspergillus awamori. Food Chem. 2014, 145, 220–227. [Google Scholar] [CrossRef]
- Song, W.; Lagmay, V.; Jeong, B.-G.; Jung, J.; Chun, J. Changes in physicochemical and functional properties of Opuntia humifusa during fermentation with cellulolytic enzyme and lactic acid bacteria. LWT 2022, 159, 113192. [Google Scholar] [CrossRef]
- Duckstein, S.M.; Lorenz, P.; Stintzing, F.C. Conversion of Phenolic Constituents in Aqueous Hamamelis virginiana Leaf Extracts During Fermentation. Phytochem. Anal. 2012, 23, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, P.; Zhang, B.; Yu, X.; Li, X.; Han, G.; Ren, Y.; Zhang, J. Combining Transcriptomics and Polyphenol Profiling to Provide Insights into Phenolics Transformation of the Fermented Chinese Jujube. Foods 2022, 11, 2546. [Google Scholar] [CrossRef]
- Jia, R.; Ge, S.; Ren, S.; Luo, Y.; Xiu, L.; Sanabil; Liu, H.; Cai, D. Antibacterial mechanism of adzuki bean seed coat polyphenols and their potential application in preservation of fresh raw beef. Int. J. Food Sci. Technol. 2021, 56, 5025–5039. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Jiang, M.; Rui, X.; Li, W.; Chen, X.H.; Dong, M.S. Effect of rose polyphenols on oxidation, biogenic amines and microbial diversity in naturally dry fermented sausages. Food Control. 2017, 78, 324–330. [Google Scholar] [CrossRef]
- Ahmed, I.A.M.; Babiker, E.E.; Al-Juhaimi, F.Y.; Bekhit, A.E. Clove Polyphenolic Compounds Improve the Microbiological Status, Lipid Stability, and Sensory Attributes of Beef Burgers during Cold Storage. Antioxidants 2022, 11, 1354. [Google Scholar] [CrossRef] [PubMed]
- Budryn, G.; Klewicka, E.; Grzelczyk, J.; Gałązka-Czarnecka, I.; Mostowski, R. Lactic acid fermentation of legume seed sprouts as a method of increasing the content of isoflavones and reducing microbial contamination. Food Chem. 2019, 285, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Abbasi-Parizad, P.; De Nisi, P.; Adani, F.; Pepé Sciarria, T.; Squillace, P.; Scarafoni, A.; Iametti, S.; Scaglia, B. Antioxidant and Anti-Inflammatory Activities of the Crude Extracts of Raw and Fermented Tomato Pomace and Their Correlations with Aglycate-Polyphenols. Antioxidants 2020, 9, 179. [Google Scholar] [CrossRef] [PubMed]
- Milutinović, M.; Dimitrijević-Branković, S.; Rajilić-Stojanović, M. Plant Extracts Rich in Polyphenols as Potent Modulators in the Growth of Probiotic and Pathogenic Intestinal Microorganisms. Front. Nutr. 2021, 8, 688843. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, S.; Tao, Y.; Li, D.; Han, Y.; Show, P.L.; Wen, G.; Zhou, J. Fermentation of blueberry and blackberry juices using Lactobacillus plantarum, Streptococcus thermophilus and Bifidobacterium bifidum: Growth of probiotics, metabolism of phenolics, antioxidant capacity in vitro and sensory evaluation. Food Chem. 2021, 348, 129083. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Sun, Y.; Chen, J.; Cheng, Y.; Zhang, H.; Gao, T.; Xu, F.; Pan, S.; Tao, Y.; Lu, J. Fermentation of ginkgo biloba kernel juice using Lactobacillus plantarum Y2 from the ginkgo peel: Fermentation characteristics and evolution of phenolic profiles, antioxidant activities in vitro, and volatile flavor compounds. Front. Nutr. 2022, 9, 1025080. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, Z.; Zhang, L. Optimization of lactic acid fermentation conditions for fermented tofu whey beverage with high-isoflavone aglycones. LWT 2019, 111, 211–217. [Google Scholar] [CrossRef]
- Lalouckova, K.; Mala, L.; Marsik, P.; Skrivanova, E. In Vitro Antibacterial Effect of the Methanolic Extract of the Korean Soybean Fermented Product Doenjang against Staphylococcus aureus. Animals 2021, 11, 2319. [Google Scholar] [CrossRef]
- Kuziel, G.A.; Rakoff-Nahoum, S. The gut microbiome. Curr. Biol. 2022, 32, R257–R264. [Google Scholar] [CrossRef]
- Linares, D.M.; Ross, P.; Stanton, C. Beneficial Microbes: The pharmacy in the gut. Bioengineered 2016, 7, 11–20. [Google Scholar] [CrossRef]
- Gou, H.-Z.; Zhang, Y.-L.; Ren, L.-F.; Li, Z.-J.; Zhang, L. How do intestinal probiotics restore the intestinal barrier? Front. Microbiol. 2022, 13, 929346. [Google Scholar] [CrossRef]
- Zhou, F.; Li, Y.-L.; Zhang, X.; Wang, K.-B.; Huang, J.-A.; Liu, Z.-H.; Zhu, M.-Z. Polyphenols from Fu Brick Tea Reduce Obesity via Modulation of Gut Microbiota and Gut Microbiota-Related Intestinal Oxidative Stress and Barrier Function. J. Agric. Food Chem. 2021, 69, 14530–14543. [Google Scholar] [CrossRef]
- Zhao, D.; Cao, J.; Jin, H.; Shan, Y.; Fang, J.; Liu, F. Beneficial impacts of fermented celery (Apium graveolens L.) juice on obesity prevention and gut microbiota modulation in high-fat diet fed mice. Food Funct. 2021, 12, 9151–9164. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PFFs | Strains Used for Fermentation | Total Phenolic Content | Changes in Polyphenol Content in PFFs after Fermentation | Parameters of Fermentation | Functional Activity Test Doses of PFFs and Corresponding Experimental Models | Changes in Functional Activity | References |
|---|---|---|---|---|---|---|---|
| Fermented Apulian table olives (Cellina di Nardò, Bella di Cerignola, Termite di Bitetto) | Yeast, lactic acid bacteria | - | Hydroxytyrosol and its derivatives ↑ | Healthy olives (150 kg) were collected, washed and placed in plastic vessels of 200 kg capacity filled with 50 L of 12% NaCI (w/v) for Cellina di Nardò and of 10% NaCI (w/v) for Bella di Cerignola and Termite di Bitetto cultivars. Olives were co-inoculated with yeast and lactic acid bacteria. Fermentations were carried out at room temperature. | Models: at present, the mainstream antioxidant capacity detection model (DPPH radical scavenging rates) Test dose: DPPH (6~42 μg/mL/Trolox) | Antioxidant activity and bioaccessibility ↑ | [33] |
| Xeniji | Lactic acid bacteria | - | (5-O-caffeoylquinic acid, 3-O-caffeoylquinic acid, sakuranetin) ↑ | - | Models: mice with alcoholic liver injury Test doses: 0.1 g/kg−1 BW, 1.0 g/kg−1 BW and 2.0 g/kg−1 BW | (1) Antioxidant activity ↑, the levels of MDA and ROS in the livers of the Xeniji-treated mice were significantly reduced as compared to the untreated mice. (2) Anti-inflammatory activity ↑, the serum levels of 1 L-1βand IL-6 were significantly lower than in untreated mice. (3) Alcoholic liver injury was significantly improved compared to the untreated mice ↑ | [34] |
| Pomegranate peel extracts (PPE) | L. acidophilus | +29.17% | (valoneic acid bilactone, punicalagin, granatin A/lagerstannin A, punigluconin, galloyl-HHDP-hexoside and pedunculagin II) ↑ | Sterilized milk (8.6 mL), 0.5–4% (w/v) PPE, membrane filtered sterilized sucrose solution (0.4 mL) and L. acidophilus (1 × 108 CFU/mL, 1 mL) were mixed together and fermented at 37 °C for 0, 4, 8, 12, and 24 h. Milk mixtures without PPE and milk mixtures without adding L. acidophilus were used as positive and negative controls, respectively | Models: Trolox-equivalent antioxidant capacity assay Test doses: ABTS value (μmol trolox/mL) | Antioxidant activity ↑, fermented: 27 μmol trolox/mL, non-fermented: 23 μmol trolox/mL. | [35] |
| Flammulina velutipes | Bacillus subtilis, Bifidobacterium longum, Saccharomyces cerevisiae (Chuangbo microorganism) | +10.11% | (syringic acid, quercetin) ↑ | 0.10% Chuangbo microorganism, molasses (3%), 28 °C, moisture contents (40%), culture time (10 days) | Models: (DPPH radical scavenging rates, hydroxyl radical scavenging rate, superoxide anion radical scavenging rate and overall reducing power), RAW264.7 Cells (anti-inflammation capacities) | (1) Antioxidant activity ↑, the DPPH, hydroxyl and superoxide anion radical scavenging rate of fermented foods were significantly higher than in non-fermented foods. (2) Anti-inflammatory activity ↑, the cell viability of the fermented foods group was higher than that of the non-fermented foods group (25, 50, and 100 µg/mL total polyphenols). | [36] |
| Citrus | Streptococcus thermophilus, Lactobacillus bulgaricus, Bifidobacterium longum, Bifidobacterium breve, Bifidobacterium lactis, Bifidobacterium infantis, Bifidobacterium adolescentis, Lactobacillus acidophilus, Lactobacillus reuteri, Bifidobacterium bifidum | - | (Naringin, vanillic acid, gallic acid, dihydrocaffeic acid, phloretic acid, 3-(3-hydroxyphenyl) propanoic acid, 4-hydroxybenzoic acid) ↑ (Hesperidin, neohesperidin, ferulic acid, p-coumaric acid) ↓ | 2 mL samples (0.5 g grapefruit or kumquat or navel orange freeze-dried powder dissolved in 4 mL PBS), 9 mL MRS liquid culture, 9 mL 1% mixed lactic acid bacteria solution, 37 °C, culture time (48 h) | Models: the free radical scavenging of DPPH, ABTS and FRAP and Caco-2 cells Test dose: 5, 10, 20, 25, 30 μg/mL. | Antioxidant activity ↑, the supernatant of fermented citrus had a significant increase in scavenging DPPH, ABTS and FRAP free radical ability. | [37] |
| Black wolfberry | Spontaneous fermentation | +42.91% | Total anthocyanins ↓, (total flavonoid and polyphenol contents) ↑ | Fresh black wolfberry was used to prepare the vinegar, crushed berries and sterile water were used at a ratio of 0.425:1 (w/w), black wolfberry was fermented in glass jars (28 °C, 60 days), the jars were sealed and placed in a dark temperature-controlled incubator. | Models: (DPPH radical scavenging rates) | Antioxidant activity ↑, the DPPH free radical scavenging ability increased gradually with fermentation time and tended to stabilize from the 20th day, until the 60th day; DPPH free radical scavenging ability reached 63.93%. | [38] |
| Tartary buckwheat leaves | Aspergillus niger | - | Rutin ↓, quercetin ↑ | A 680 mL bottle with air-vent capping was used in fermentation, 110 g tartary buckwheat leaves, inoculated with spore suspension (5%, v:m), sterile water (1:1, v:m), 28 °C, culture time: 24 days. | Models: (DPPH radical scavenging rates, Ferricyanide reduction assay) | Antioxidant activity ↑, both the 2 antioxidant activities of fermented foods were promoted, a 150% increase in DPPH scavenging activity and a two-fold higher reducing power. | [39] |
| Soy sauce (production with soybean and wheat flour) | Aspergillus oryzae 3.042 | - | (Glucosides, malonylglucosides and acetylglucosides) ↓, (daidzein, glycitein and genistein) ↑ | Fermentation process according to China national standard (GB 18186-2000), soybean and wheat flour were mixed at the mass ratio of 4:1 with Aspergillus oryzae 3.042 as koji for 44 h at 28–30 °C, then koji was immersed in a brine containing 17 g/100 g NaCl as moroml and fermented for 90 days. | Models: (DPPH radical scavenging rates) | Antioxidant activity ↑, the antioxidant capacity of soy sauce was tripled after fermentation, the maximum value was detected in soy sauce-90 d sample (45.29 ± 0.71 TE μmol/100 mL) | [40] |
| Cheonggukjang (production with soybean) | Spontaneous fermentation | +30.26% | (Daidzin, genisin and glycitin) ↓, (daidzein, glycitein and genistein) ↑ | Raw soybeans were washed and soaked for 12 h at 25 °C, then steamed for 45 min at 110 °C and left to stand for 1 h at 25 °C; afterwards, the cooled soybeans were moved to the fermentation room for 72 h at 46 °C, and lastly they were mixed with 4% salt. | Models: at present, the mainstream antioxidant capacity detection model (DPPH radical scavenging rates, ferric-reducing antioxidant power) | Antioxidant activity ↑, the DPPH radical scavenging activity: 99.80 to 104.24 μM TE/g; FRAP values: 38.00 to 63.49 μM TE/g | [41] |
| Soybean | Bifidobacterium breve | - | Glycosides almost disappeared after 24 h of fermentation, while aglycogens increased significantly up to 24 h of fermentation. Aglycones have a better effect than glycosides | Isoflavones were extracted from soybean and added to the medium (400 ppm), the culture was incubated at 37 °C under anaerobic conditions, and samples were removed from each reaction vessel at 0, 10, and 24 h | Models: (1) lipase; (2) mice; (3) 3T3-L1 cells. Test dose: (1) 100 ppm glycosides and 50 ppm aglycones; (2) 1 mg/mL glycosides and aglycones; (3) 0.1, 0.25 mg/mL glycosides and aglycones. | (1) Inhibition of lipase activity ↑, non-fermented 30%→fermented 63.6%; (2) inhibition of adipocyte differentiation and reduction of triglyceride content) ↑ | [42] |
| Fermented soymilk | Lactobacillus rhamnosus strain ASCC 1520 | - | - | Strain ASCC 1520 was activated by two successive transfers in MRS media, then the activated cultures were transferred into 50 mL of sterile soymilk at an inoculum level of 1% (v/v) and incubated in a CO2 incubator at 37 °C without agitation to obtain fermented soymilk | Models: mice. Test dose: Mice drink freely | (Maintain the balance of intestinal flora, bioavailability) ↑ | [43] |
| Fermented blueberry juices | Lacto- bacillus plantarum, Lactobacillus fer- mentum | +41.8% | (Delphinium-3-glucoside, peonidin-3-glucoside, malvidin-3-glucoside, and malvidin-3- Arabinoside) ↓ (gallic acid, chlorogenic acid, rutin, myricetin and quercetin-3- Rhamnoside) ↑ | Blueberries were washed, homogenized and filtered, sterilized to obtain the juices, pH value was adjusted to 4.0 with 1 M Na2CO3. After pasteurization, juices were inoculated with 1.0% (v/v) inoculum (7.0 log CFU/mL), 37 °C, 48 h, in darkness. | Models: the free radical scavenging of ABTS and FRAP | Antioxidant activity ↑, non-fermented foods: ABTS and FRAP 68.08 ± 0.25 mmol Trolox/L and 7.22 ± 0.25 mmol Fe2+/L; fermented foods: ABTS and FRAP 108.94 ± 0.19 mmol Trolox/L and 12.32 ± 1.77 mmol Fe2+/L | [44] |
| Fermented blueberry juices | Streptococcus lactis, Pediococcus pentosaceus | - | Malvidin anthocyanins ↓ (catechins, syringic acid, and p-coumaric acid, epicatechin, ferulic acid and caffeic acid) ↑ | Blueberry pulp was placed into glass bottles (250 mL), fermented blueberry pulp was prepared by inoculating each bottle with 3.2 vol% Streptococcus lactis followed by 49 h incubation at 37 °C. (3.7 vol% Pediococcus pentosaceus followed by 47 h incubation at 40 °C). | Models: (DPPH radical scavenging rates, ABTS radical scavenging rates). | (1) Antioxidant activity ↑, the ABTS antioxidant capacity of free phenols increased by 8.47%, the DPPH radical scavenging capacity increased by 18.38%; (2) Bioavailability ↑ | [45] |
| FE Type | Strain Source | Substrate of Action | Reference |
|---|---|---|---|
| Type A | A. Tubingensis, A. awamori | Ferulic acid and erucic acid (substrate with methoxy substituents) | [82] |
| Type B | N. crassa Fae-1, Penicillium funiculosum | p-Coumaric acid, caffeic acid (substrates containing 1 or 2 hydroxyl substituents) | [83] |
| Type C | A. oryzae, A. niger, Talaromyces stipitatus, F. oxysporum | Caffeic acid, erucic acid, ferulic acid, p-coumaric acid (four methyl esters of p-hydroxycinnamic acid have extensive activity and are more active against n-propyl ferulate) | [84,85] |
| Type D | Psuedomonas fluorescens Subsp. | Caffeic acid, erucic acid, ferulic acid, p-coumaric acid (broad activity against the four methyl esters of hydroxycinnamic acid and strong hydrolytic activity against acetyl residues) | [86] |
| Glycosidase Name | PFFs | Target Phenolic Glycosides | Aglycones by Action of Glycosidase | Changes in PFFs Following Glycosidase Action | Microorganisms That Secrete Glycosidase | References |
|---|---|---|---|---|---|---|
| α-L-rhamnosidase, β-D-glucosidase | Citrus Juice | Naringin (356.33 mg/L→10.52 mg/L) | Naringenin | (1) Significantly reduced bitterness, (2) Increased antioxidant activity (DPPH, ABTS radical scavenging ability) | - | [102] |
| β-glucosidase | Hippophae rhamnoides leaves | Rutin (4.61 mg/L→0.92 mg/L) | Quercetin, kaempferol, and isorhamnetin | (1) Increased antioxidant activity (DPPH, ABTS radical scavenging ability and ferric reducing/antioxidant power (FRAP) assay. DPPH: fermented 166.62, non-fermented 124.11; ABTS: fermented 188.32, non-fermented 135.67; FRAP: fermented 212.45, non-fermented 135.67 (mg Trolox equivalents/g dry leaf)) (2) Increased total phenolic content (55.97 ± 1.72 mg GAE/g DW→100.16 ± 3.25 mg GAE/g DW) | Eurotium amstelodami BSX001 | [103] |
| Cellulase, β-glucosidase, xylanase, α-amylase | Mulberry leaves | Flavonoid Glycosides were consumed, | Quercetin and kaempferol | (1) Increased antioxidant activity (DPPH and ABTS radical scavenging ability, DPPH: fermented IC50 = 95.75 μg/mL, non-fermented IC50 = 106.99 μg/mL; ABTS: fermented IC50 = 82.19 μg/mL, non-fermented: IC50 = 161.58 μg/mL). (2) Increased inhibitory activities of α-glucosidase (fermented: IC50 = 35.02 μg/mL, non-fermented: IC50 = 95.75 μg/mL). (3) Antibacterial activity (Candida albicans, Staphylococcus aureuswine) | Monascus | [105] |
| Cellulase, β-glucosidase, xylanase, hemicellulase | Guava leaf tea | Isoquercitrin, quercetin-3-O-β-D-xylopyranoside, Avicularin, kaempferol-3-O-glucose | Quercetin, kaempferol, gallic acid | (1) Increase in free phenol content (56.1%→88.4%) (2) Increased antioxidant activity (DPPH, ABTS radical scavenging ability, DPPH:fermented IC50 = 14.7 μg/mL, non-fermented IC50 = 39.5 μg/mL; ABTS: fermented IC50 = 4.5 μg/mL, non-fermented: IC50 = 9.4 μg/mL). (3) Increased α-glucosidase inhibitory activity (fermented: IC50 = 11.8 μg/mL, non-fermented: IC50 = 19.2 μg/mL). | Monascus anka, Saccharomyces cerevisiae | [106] |
| α-L-rhamnosidase, β-glucosidase | Stingless bee honey | Quercetin-3-rutinoside, Quercetrin-3-rhamnoside | Quercetin and kaempferol | Increased antioxidant ability, antibacterial ability, anti-inflammatory ability and anti-cancer activity | Penicillium sp. Monascus anka, Bacillus spp. | [107] |
| β-glucosidase | Black soybeans | Anthocyanins (cyanidin-3-o-β-glucoside), soy isoflavones (isoflavones 7-O-β-d-glucoside) | Genistein, daidzein and cyanidin | Increased antioxidant activity, anti-proliferative activity, and antidiabetic activity | Rhizoctonia, Bacillus | [108] |
| β-glucosidase | Soy milk | Daidzin, genistin | Daidzein, genistein | Increased antioxidant activity (the result of antioxidant activity of soymilks is expressed as a percentage of inhibition; fermented: 83.3%, non-fermented: 20%) | Lactobacillus | [109] |
| Polyphenol Composition | Inhibit Pathogenic Bacteria Growth | Promote Probiotic Growth | PFFs | Parameters of Fermentation | Methods for Determining Microbiological Counts | Reference |
|---|---|---|---|---|---|---|
| Hydrolyzable tannin, flavonol | Eudomonas, Psychrobacter, Acinetobacter, Staphylococcus and Kocuria | Lactobacillus | Fermented sausages with rose polyphenol extract | 9.6 kg of lean pork and 2.4 kg of backfat were diced into pieces, and supplemented with 1, 2 and 3 mg rose polyphenols per gm meat. Ground meat: 80% lean pork, 20% pork backfat, 2% salt (w/w), 0.5% sodium glutamate (w/w), 2% sugar (w/w), 1% rice wine (w/w), 0.05% sodium erythorbate, 0.2% sodium polyphosphate(w/w), 0.012% sodium nitrite (w/w); blended for 3 min, then stuffed into hog casing; the sausages were fermented, dried in an incubator (20 °C, 90% relative humidity) for 3 days, then 10 °C, 80% relative humidity for 5 days, and 10 °C, 70% relative humidity for 16 days. | Standard plate count method, 16s rDNA sequencing. | [151] |
| Isoflavonoids | E.coli, Klebsiella sp., Salmonella sp. and Shigella sp. | Lactic acid bacteria | Legume sprouts | 50 g of sprouts were flooded with 50 mL of lactic acid bacteria inoculum suspension, fermented condition (30 °C, 48~96 h, depending on the characteristics of the fermented sprouts) | Standard plate count method | [153] |
| Naringin, kaempferol, gallic acid, hydroxycinnamic acid | - | Lactic acid bacteria | Tomato | Approximately 300 g of tomato (wet weight) were packed into airtight glass containers of 500 mL and pressed to favor the air exit, fluxed with N2 before being closed, stored at 20 °C, dark conditions for 240 days. | - | [154] |
| Flavonoid (quercetin, rutin, quercetin, hyperoside, epicatechin, catechin, etc.), phenolic acids (chlorogenic acid, caffeic acid, p-coumaric acid, etc.) | S.aureus, L.monocytogenes, E.faecalis, E.coli, P.aeruginosa, C.albicans | Yeast (S. boulardii) Lactobacilli (L. plantarum) L.rhamnosus | St. John’s wort, winter savory, yarrow, willow gentian | - | Broth microdilution method (results were determined with the minimum inhibitory concentration) | [155] |
| Syringic acid, ferulic acid, gallic acid | - | Lactobacillus plantarum, Streptococcus thermophilus and Bifidobacterium | Blueberry, blackberry | 2% (v/v) activated potential probiotics were allotted into 100 mL flasks containing 50 mL of pasteurized blueberry or blackberry juice (fermentation 37 °C, 48 h) | Standard plate count method | [156] |
| Catechin, proanthocyanidin B2, gallic acid, phloretin | - | Lactobacillus plantarum Y2 | Ginkgo peel | Ginkgo peel juice was pasteurized in a water bath at 90 °C for 20 min; then 1% (v/v) inoculum of ginko peel juice was inoculated to ensure an initial viable count of approximately 5.0 Log CFU/mL, fermented in an incubator at 37 °C for 48 h. | Standard plate count method | [157] |
| Daidzein, genistein | - | Lactobacillus paracasei, Leuconostoc mesenteroides, Lactobacillus rhamnosus GG and Lactobacillus plantarum | Fermented tofu whey beverage | pH was adjusted to 6 after centrifugation, sterilization at 115 °C for 10 min (tofu whey), then about 500 mL of tofu whey was inoculated with screened strains, then fermented at optimal temperature and pH for 36 h. | Standard plate count method | [158] |
| Soybean isoflavone | Staphylococcus aureuswine | - | Doenjang (the Korean soybean fermented product) | - | Broth microdilution method (results determined with the minimum inhibitory concentration) | [159] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Chen, C.; Ni, D.; Yang, Y.; Tian, J.; Li, Y.; Chen, S.; Ye, X.; Wang, L. Effects of Fermentation on Bioactivity and the Composition of Polyphenols Contained in Polyphenol-Rich Foods: A Review. Foods 2023, 12, 3315. https://doi.org/10.3390/foods12173315
Yang F, Chen C, Ni D, Yang Y, Tian J, Li Y, Chen S, Ye X, Wang L. Effects of Fermentation on Bioactivity and the Composition of Polyphenols Contained in Polyphenol-Rich Foods: A Review. Foods. 2023; 12(17):3315. https://doi.org/10.3390/foods12173315
Chicago/Turabian StyleYang, Fan, Chao Chen, Derang Ni, Yubo Yang, Jinhu Tian, Yuanyi Li, Shiguo Chen, Xingqian Ye, and Li Wang. 2023. "Effects of Fermentation on Bioactivity and the Composition of Polyphenols Contained in Polyphenol-Rich Foods: A Review" Foods 12, no. 17: 3315. https://doi.org/10.3390/foods12173315
APA StyleYang, F., Chen, C., Ni, D., Yang, Y., Tian, J., Li, Y., Chen, S., Ye, X., & Wang, L. (2023). Effects of Fermentation on Bioactivity and the Composition of Polyphenols Contained in Polyphenol-Rich Foods: A Review. Foods, 12(17), 3315. https://doi.org/10.3390/foods12173315