
Antibacterial Effects of Ramulus mori Oligosaccharides against Streptococcus mutans

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of RMOS
2.2. Bacterial Strain and Culture Conditions
2.3. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) Assays
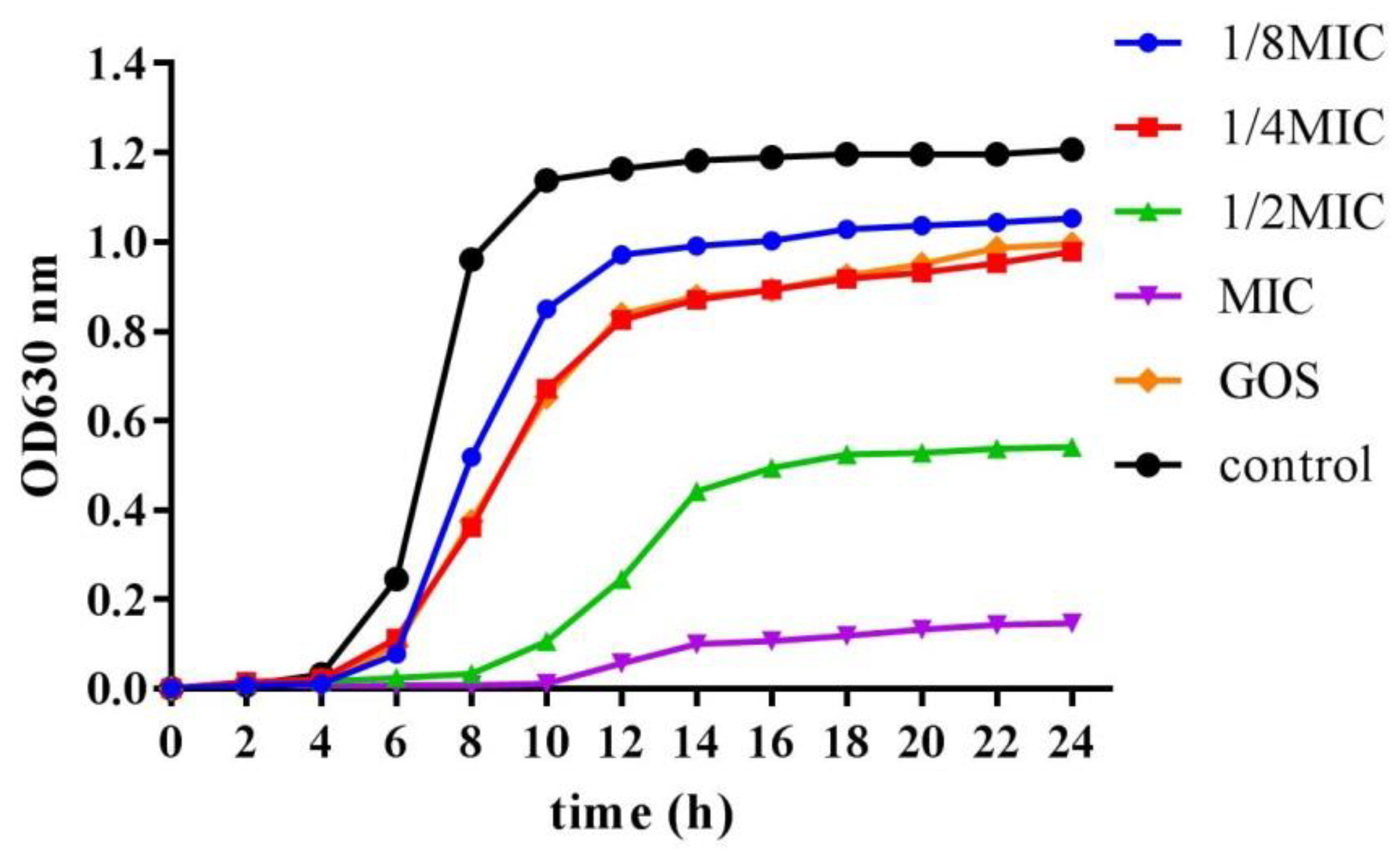
2.4. Growth Curve Assay
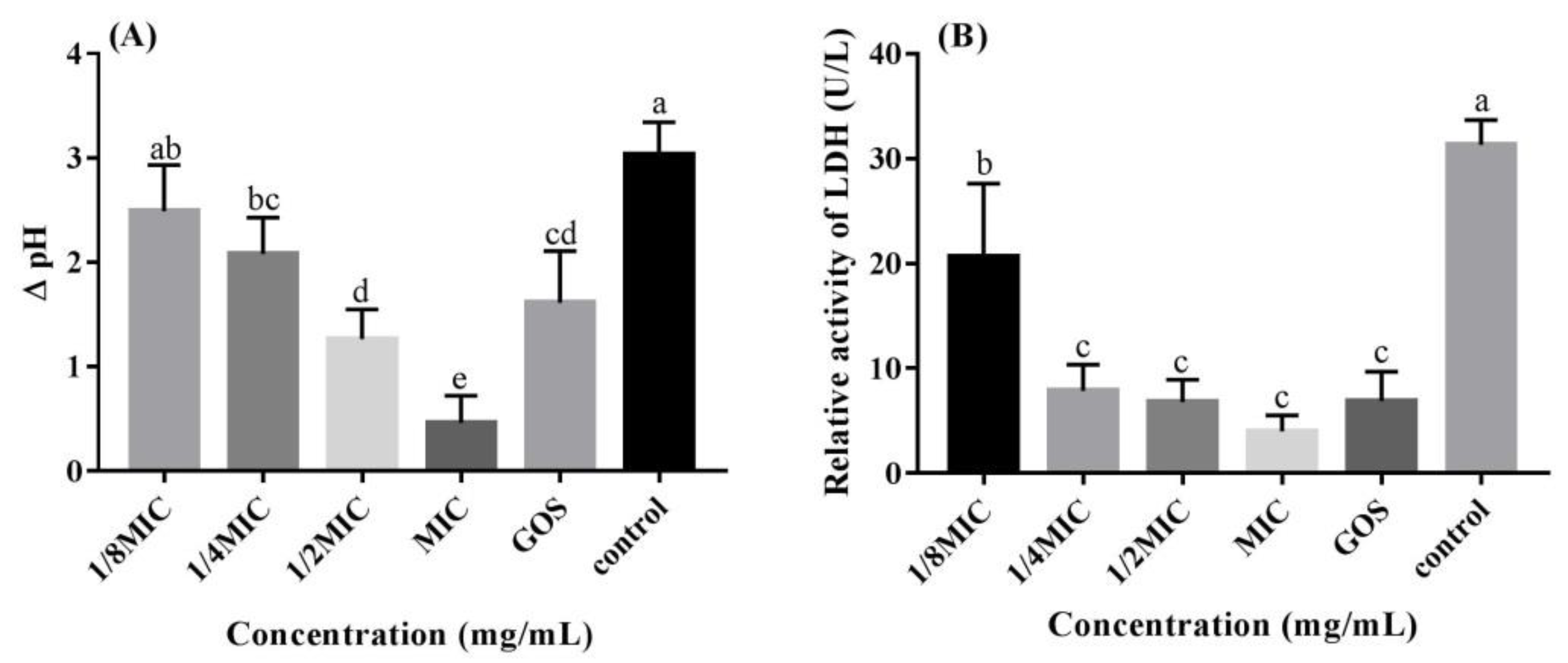
2.5. Glycolytic pH Drop Assay
2.6. Lactate Dehydrogenase (LDH) Measurement
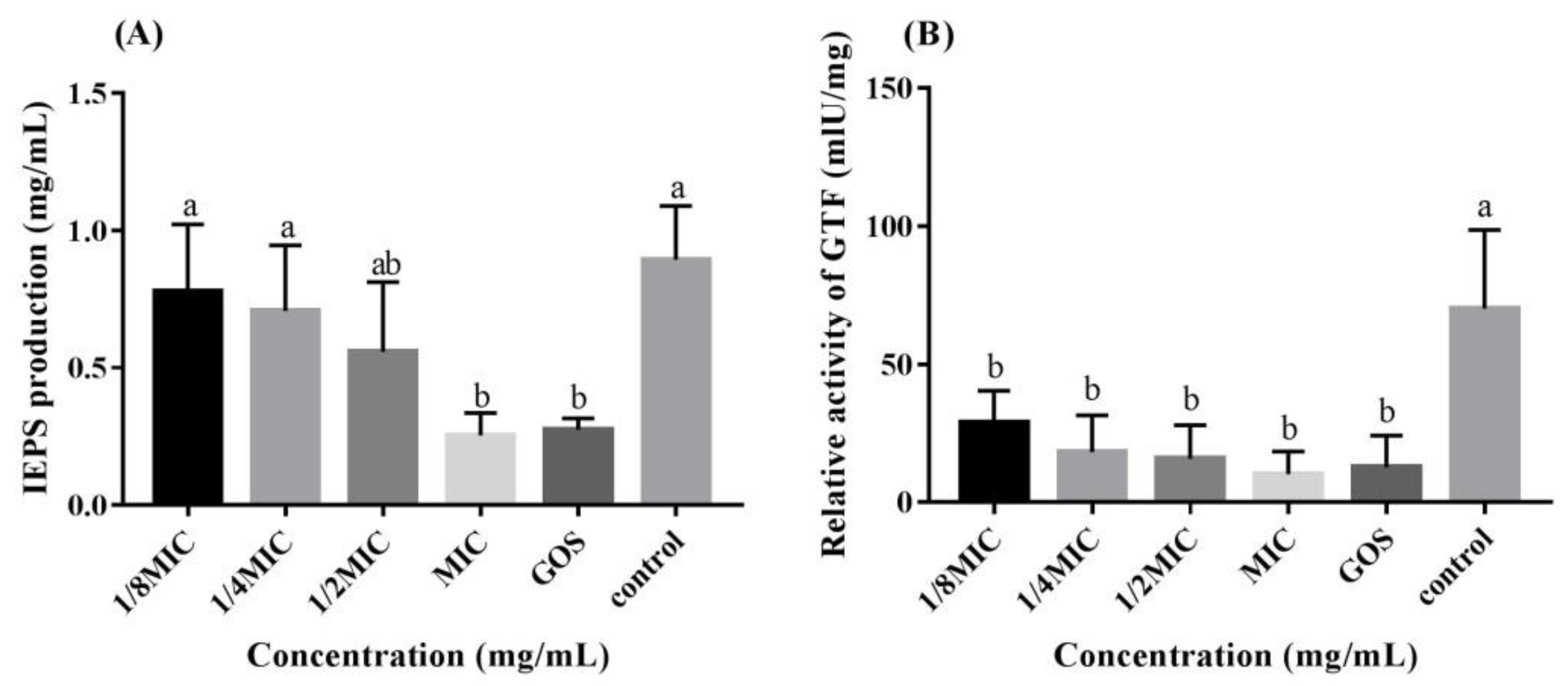
2.7. IEPS Measurement
2.8. GTF Measurement
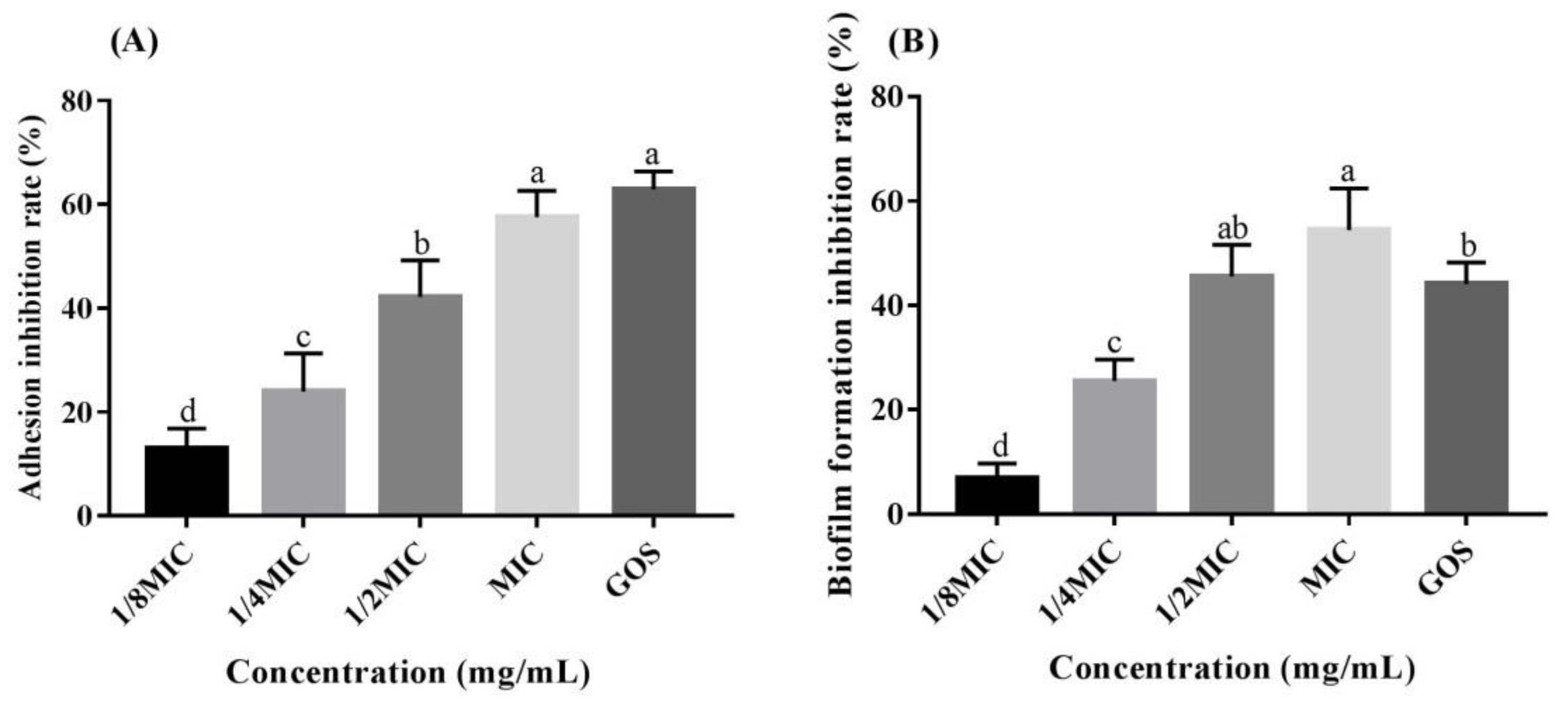
2.9. Adhesion Assay
2.10. Biofilm Formation Assay
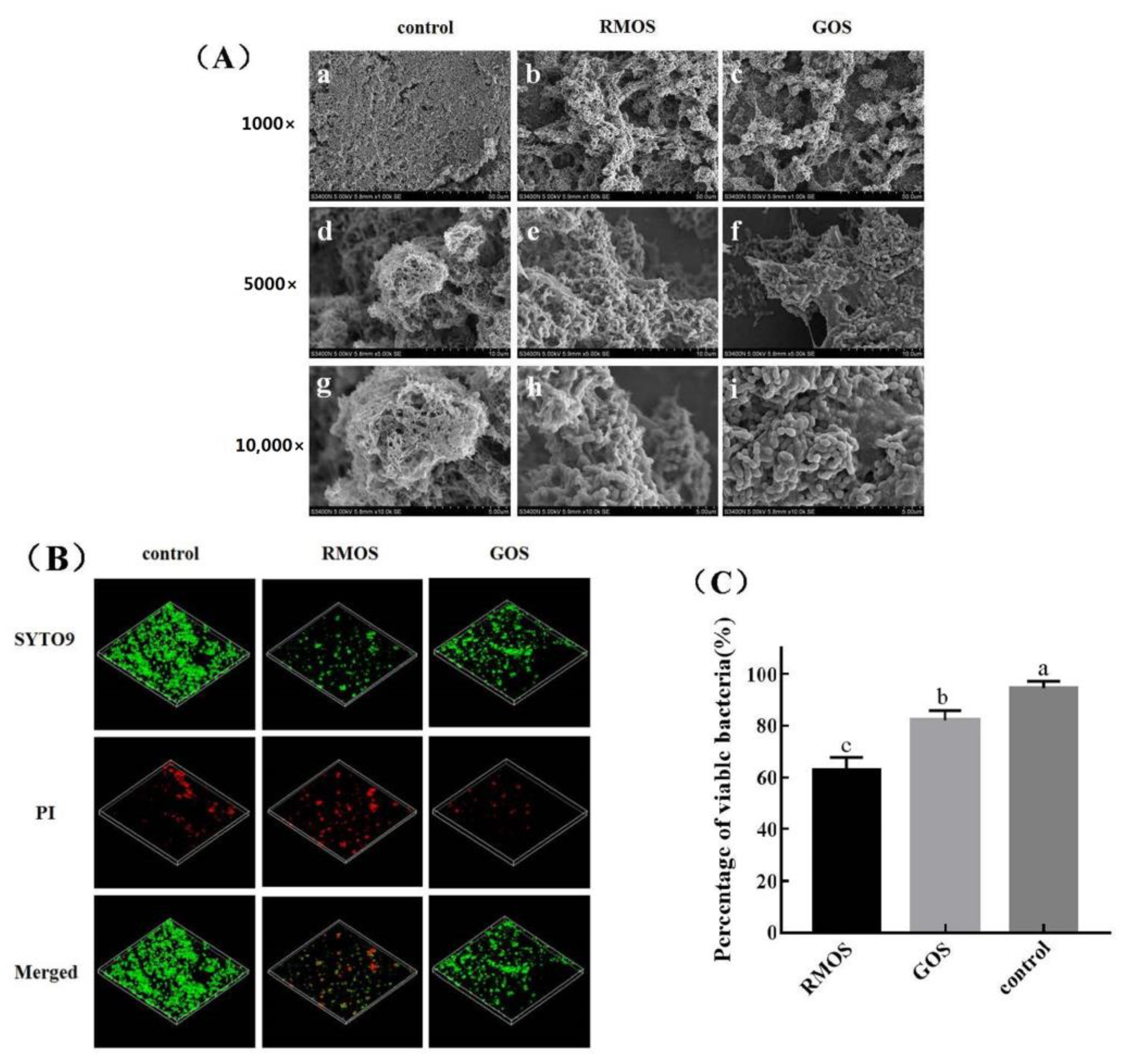
2.11. Scanning Electron Microscopy (SEM)
2.12. Confocal Laser Scanning Microscopy (CLSM)
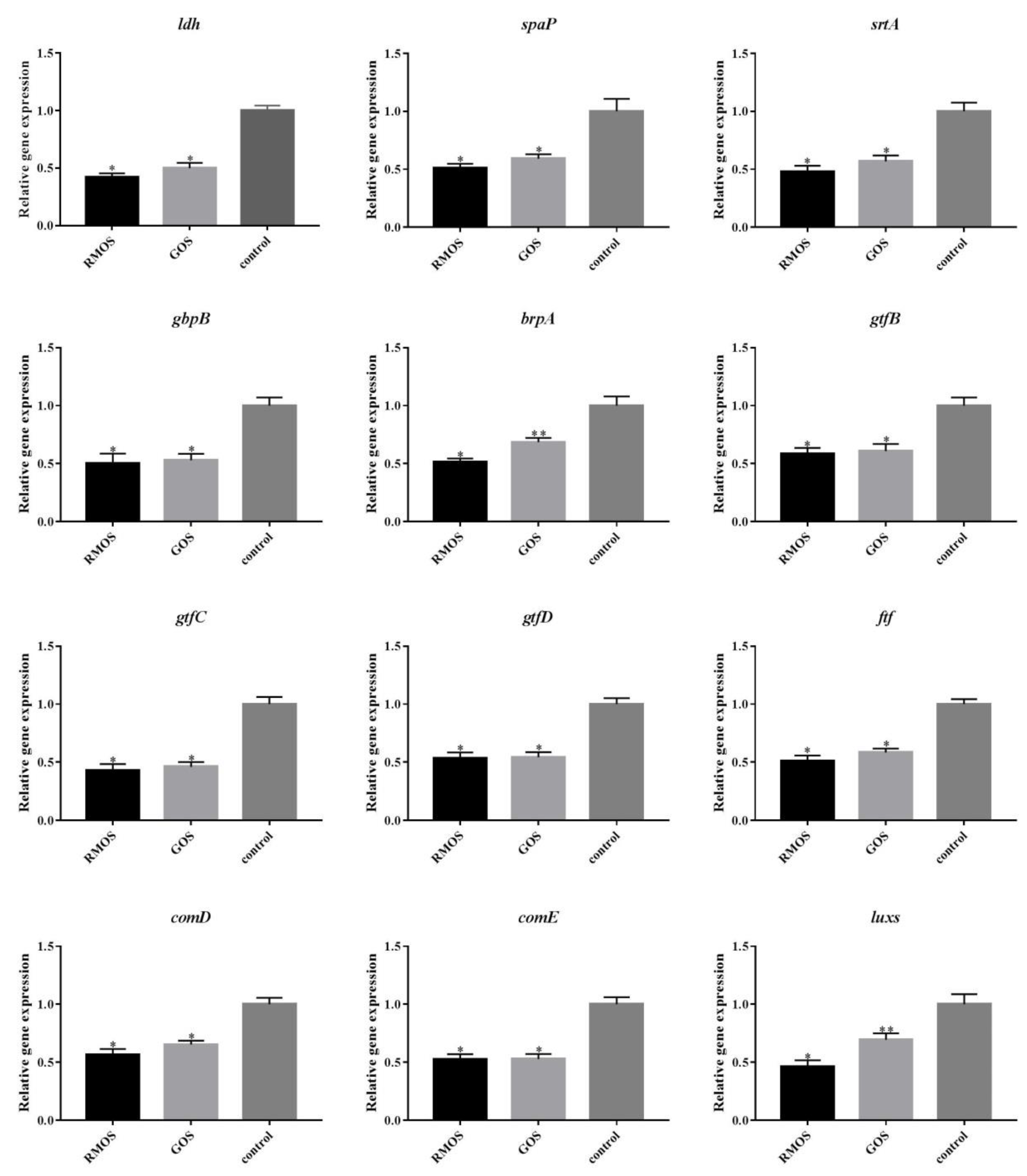
2.13. qRT-PCR Assay
2.14. Analysis and Statistics
3. Results and Discussion
3.1. Antibacterial Activity of RMOS on S. mutans
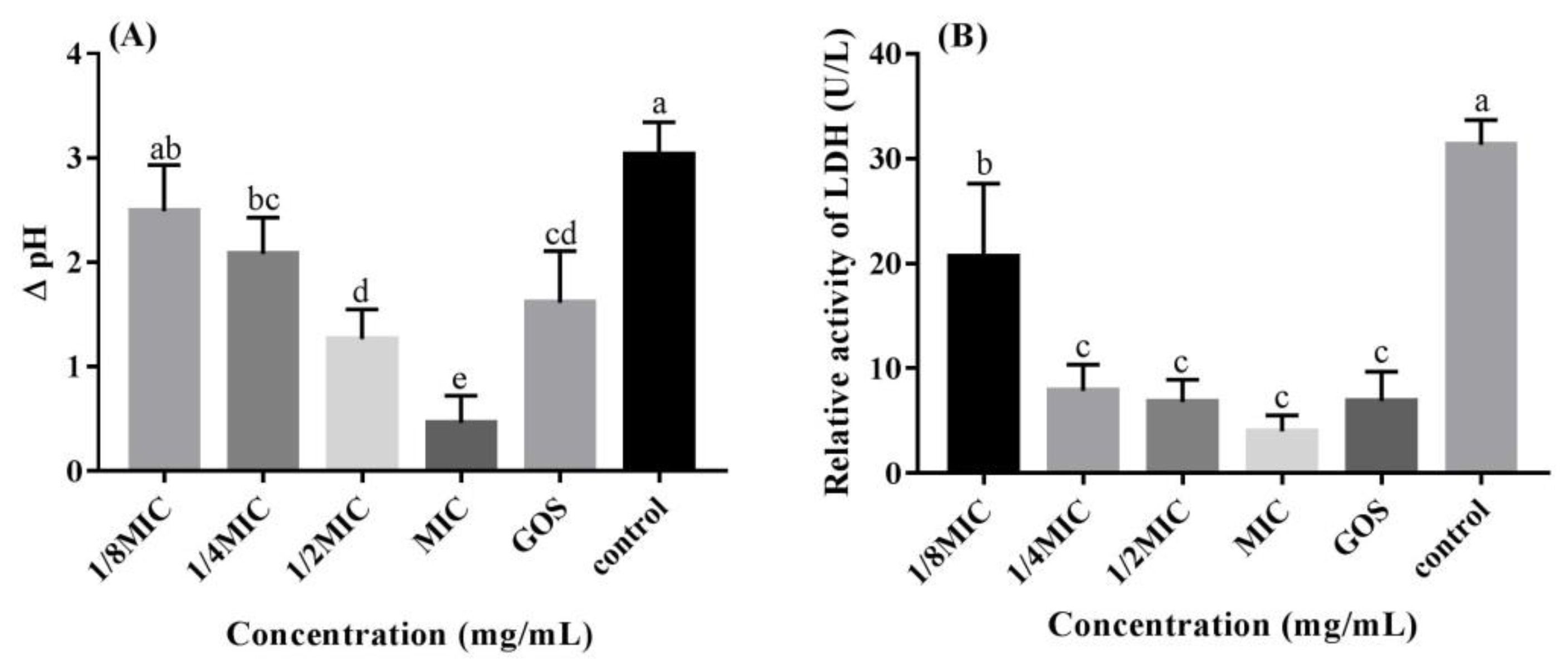
3.2. RMOS Inhibits the Acidogenicity of S. mutans
3.3. RMOS Inhibits IEPS and the GTF Activity of S. mutans
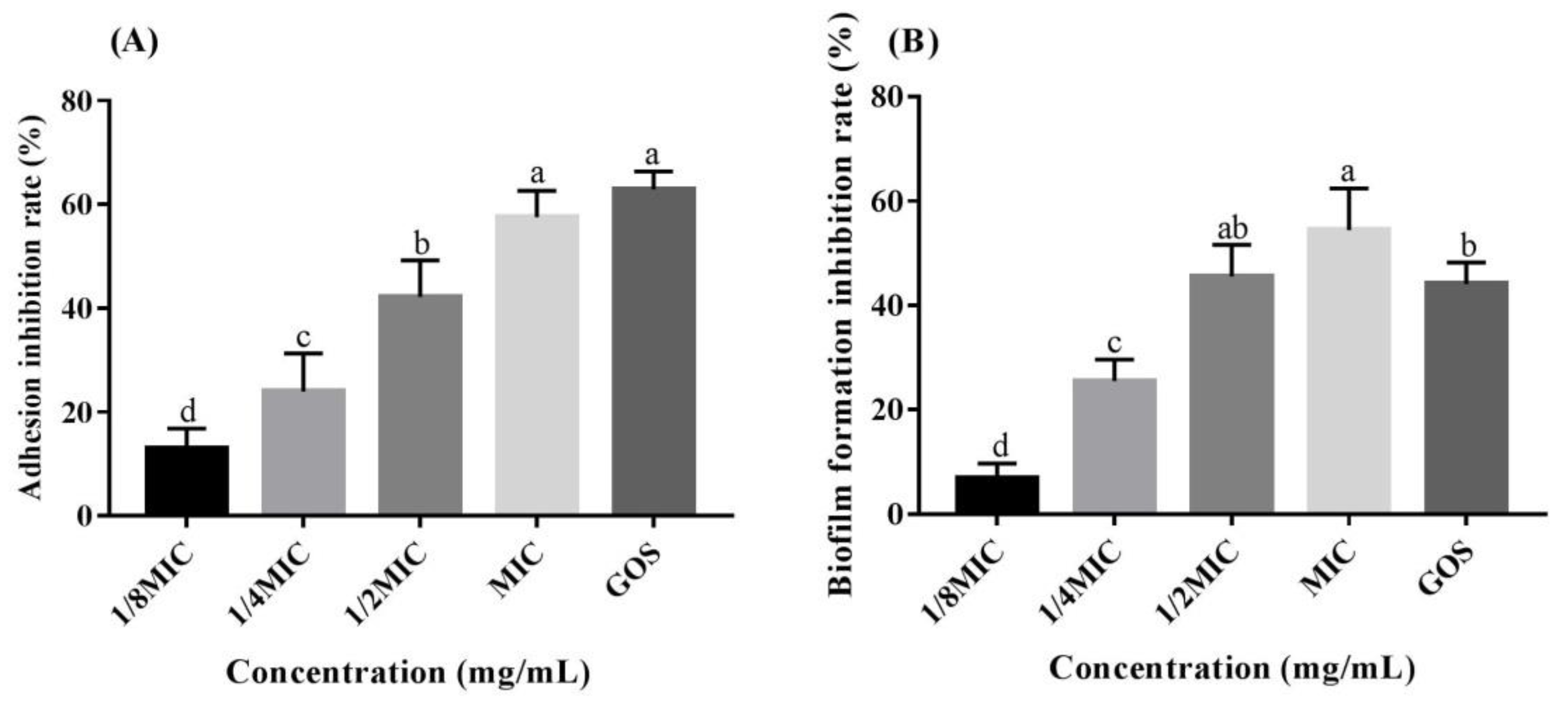
3.4. RMOS Suppresses Biofilm Formation by S. mutans
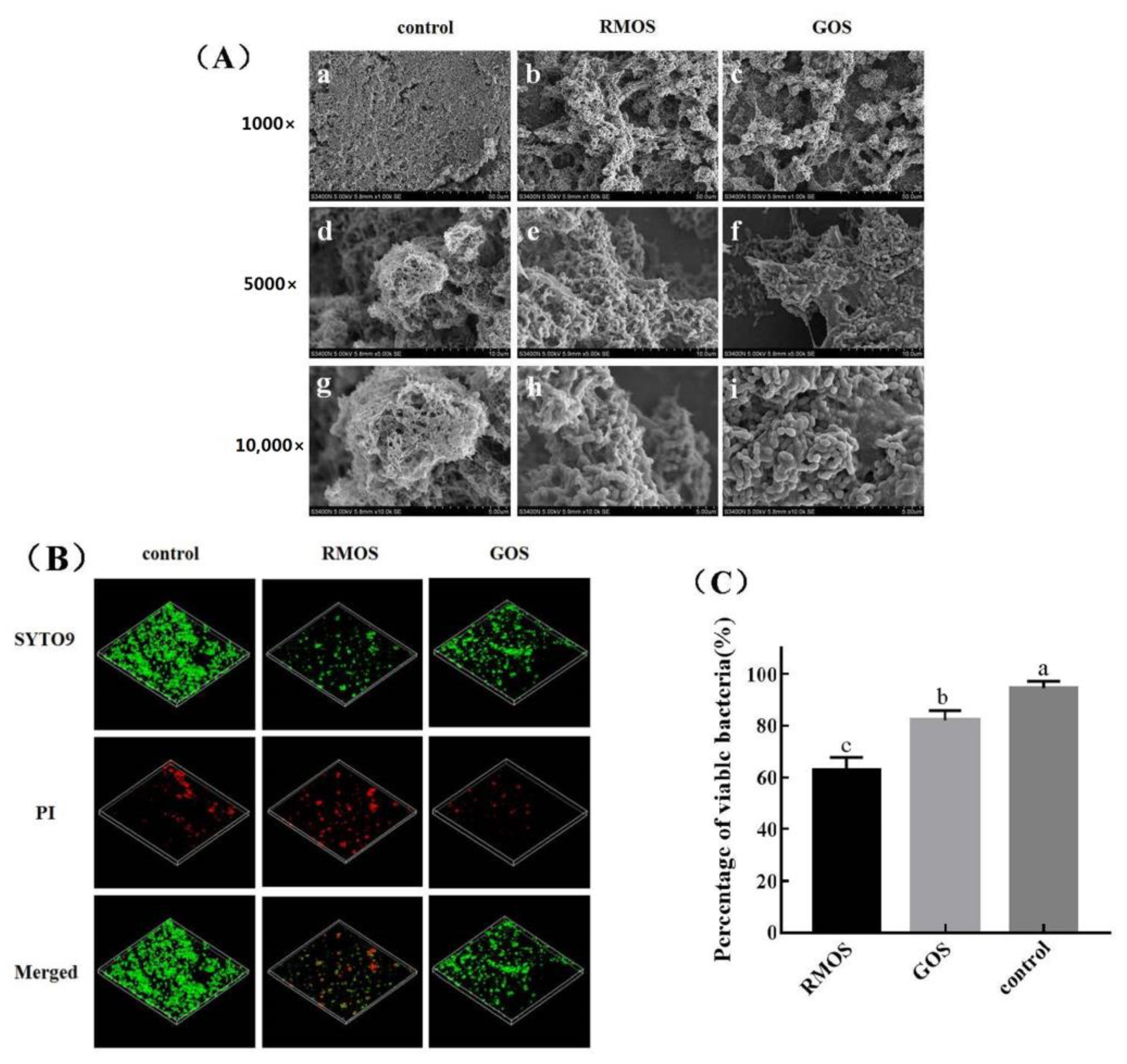
3.5. RMOS Destroys the Biofilm Structures of S. mutans
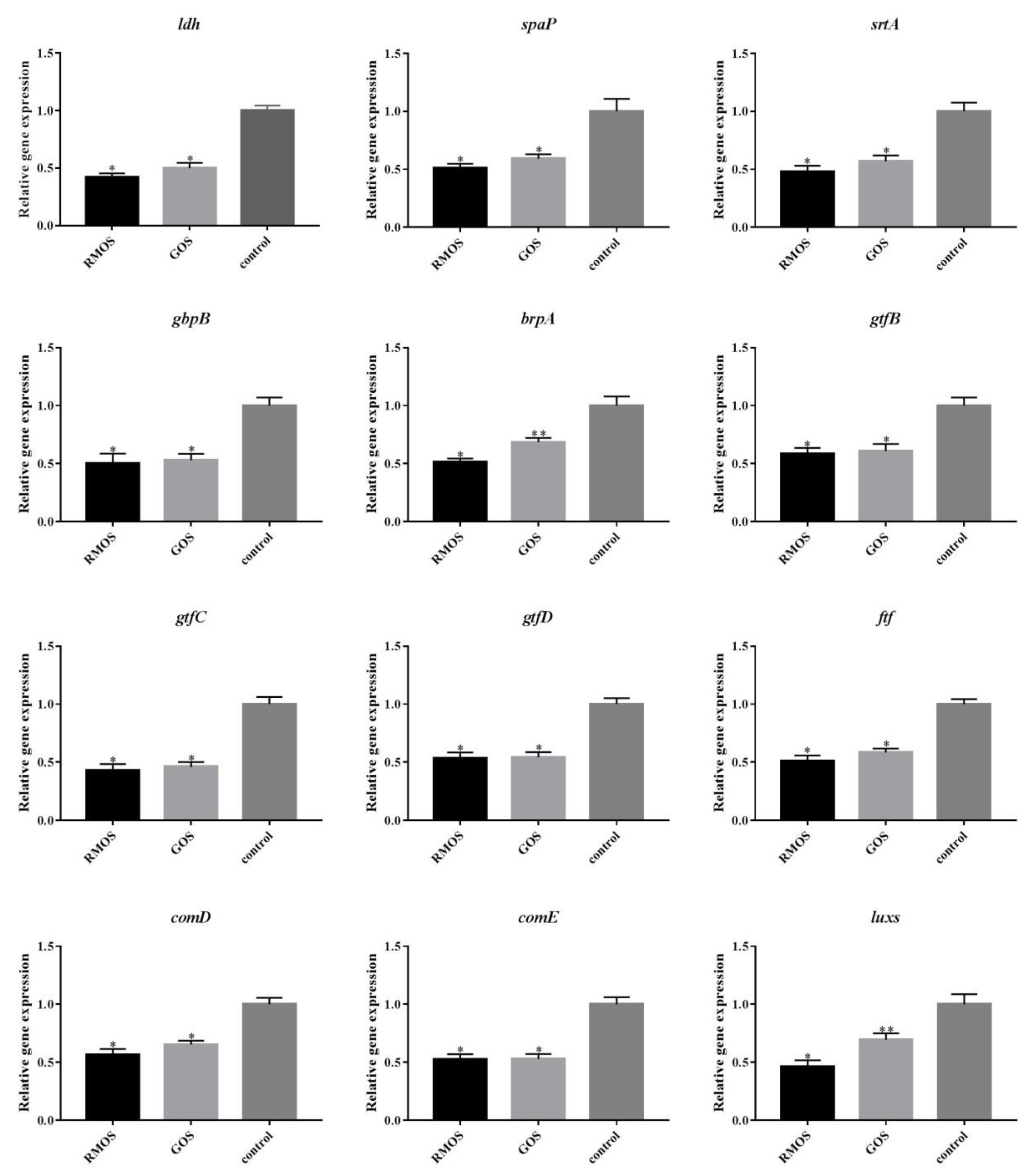
3.6. RMOS Downregulates the Expression of Virulence Genes of S. mutans
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Krzyściak, W.; Jurczak, A.; Kościelniak, D.; Bystrowska, B.; Skalniak, A. The virulence of Streptococcus mutans and the ability to form biofilms. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Marquis, R. Oxygen metabolism, oxidative stress and acid-base physiology of dental plaque biofilms. J. Ind. Microbiol. Biotechnol. 1995, 15, 198–207. [Google Scholar] [CrossRef]
- Takahashi, N.; Nyvad, B. Caries ecology revisited: Microbial dynamics and the caries process. Caries Res. 2008, 42, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Yu, L.-C. In-situ, time-lapse study of extracellular polymeric substance discharge in Streptococcus mutans biofilm. Colloids Surf. B Biointerfaces 2017, 150, 98–105. [Google Scholar] [CrossRef]
- Rossoni, R.D.; dos Santos Velloso, M.; de Barros, P.P.; de Alvarenga, J.A.; Dos Santos, J.D.; dos Santos Prado, A.C.C.; de Camargo Ribeiro, F.; Anbinder, A.L.; Junqueira, J.C. Inhibitory effect of probiotic Lactobacillus supernatants from the oral cavity on Streptococcus mutans biofilms. Microb. Pathog. 2018, 123, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Saleem, H.G.M.; Seers, C.A.; Sabri, A.N.; Reynolds, E.C. Dental plaque bacteria with reduced susceptibility to chlorhexidine are multidrug resistant. BMC Microbiol. 2016, 16, 214. [Google Scholar] [CrossRef]
- Mejàre, I.; Lingström, P.; Petersson, L.G.; Holm, A.K.; Twetman, S.; Källestål, C.; Nordenram, G.; Lagerlöf, F.; Söder, B.; Norlund, A. Caries-preventive effect of fissure sealants: A systematic review. Acta Odontol. Scand. 2003, 61, 321–330. [Google Scholar] [CrossRef]
- Zhang, G.; Tan, Y.; Yu, T.; Wang, S.; Liu, L.; Li, C. Synergistic antibacterial effects of reuterin and catechin against Streptococcus mutans. LWT 2021, 139, 110527. [Google Scholar] [CrossRef]
- Xiao, J.; Zuo, Y.; Liu, Y.; Li, J.; Hao, Y.; Zhou, X. Effects of Nidus Vespae extract and chemical fractions on glucosyltransferases, adherence and biofilm formation of Streptococcus mutans. Arch. Oral Biol. 2007, 52, 869–875. [Google Scholar] [CrossRef]
- Shui, Y.; Jiang, Q.; Lyu, X.; Wang, L.; Ma, Q.; Gong, T.; Zeng, J.; Yang, R.; Li, Y. Inhibitory effects of sodium new houttuyfonate on growth and biofilm formation of Streptococcus mutans. Microb. Pathog. 2021, 157, 104957. [Google Scholar] [CrossRef]
- Wen, P.; Hu, T.-G.; Linhardt, R.J.; Liao, S.-T.; Wu, H.; Zou, Y.-X. Mulberry: A review of bioactive compounds and advanced processing technology. Trends Food Sci. Technol. 2019, 83, 138–158. [Google Scholar] [CrossRef]
- He, N.; Zhang, C.; Qi, X.; Zhao, S.; Tao, Y.; Yang, G.; Lee, T.-H.; Wang, X.; Cai, Q.; Li, D. Draft genome sequence of the mulberry tree Morus notabilis. Nat. Commun. 2013, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Chen, H.; Xiang, Z.; He, N. Preparation of polysaccharides from Ramulus mori, and their antioxidant, anti-inflammatory and antibacterial activities. Molecules 2019, 24, 856. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; He, T.-Z.; Zhang, Y.-Q. The isolation and the characterization of two polysaccharides from the branch bark of mulberry (Morus alba L.). Arch. Pharm. Res. 2016, 39, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ji, H.; Zhang, C.; Yu, J.; Liu, A. Structural characterization and antitumor activity of a novel polysaccharide from Grifola frondosa. J. Food Meas. Charact. 2020, 14, 272–282. [Google Scholar] [CrossRef]
- He, X.; Fang, J.; Ruan, Y.; Wang, X.; Sun, Y.; Wu, N.; Zhao, Z.; Chang, Y.; Ning, N.; Guo, H.; et al. Structures, bioactivities and future prospective of polysaccharides from Morus alba (white mulberry): A review. Food Chem. 2018, 245, 899–910. [Google Scholar] [CrossRef]
- Qin, Y.; Xie, J.; Xue, B.; Li, X.; Gan, J.; Zhu, T.; Sun, T. Effect of acid and oxidative degradation on the structural, rheological, and physiological properties of oat β-glucan. Food Hydrocoll. 2021, 112, 106284. [Google Scholar] [CrossRef]
- Silva, N.S.d.; Araújo, N.K.; Daniele-Silva, A.; Oliveira, J.W.d.F.; Medeiros, J.M.d.; Araújo, R.M.; Ferreira, L.D.S.; Rocha, H.A.O.; Silva-Junior, A.A.; Silva, M.S. Antimicrobial activity of chitosan oligosaccharides with special attention to antiparasitic potential. Mar. Drugs 2021, 19, 110. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Mancilha, I.M. Non-digestible oligosaccharides: A review. Carbohydr. Polym. 2007, 68, 587–597. [Google Scholar] [CrossRef]
- Salli, K.; Söderling, E.; Hirvonen, J.; Gürsoy, U.; Ouwehand, A. Influence of 2′-fucosyllactose and galacto-oligosaccharides on the growth and adhesion of Streptococcus mutans. Br. J. Nutr. 2020, 124, 824–831. [Google Scholar] [CrossRef]
- Li, E.; Yang, H.; Zou, Y.; Wang, H.; Hu, T.; Li, Q.; Liao, S. In-vitro digestion by simulated gastrointestinal juices of Lactobacillus rhamnosus cultured with mulberry oligosaccharides and subsequent fermentation with human fecal inocula. LWT 2019, 101, 61–68. [Google Scholar] [CrossRef]
- Cai, S.; Meng, K.; Liu, P.; Cao, X.; Wang, G. Suppressive effects of gecko cathelicidin on biofilm formation and cariogenic virulence factors of Streptococcus mutans. Arch. Oral Biol. 2021, 129, 105205. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Fan, Y.; Wang, X.; Jiang, X.; Zou, J.; Huang, R. Effects of the natural compound, oxyresveratrol, on the growth of Streptococcus mutans, and on biofilm formation, acid production, and virulence gene expression. Eur. J. Oral Sci. 2020, 128, 18–26. [Google Scholar] [CrossRef]
- Senpuku, H.; Tuna, E.B.; Nagasawa, R.; Nakao, R.; Ohnishi, M. The inhibitory effects of polypyrrole on the biofilm formation of Streptococcus mutans. PLoS ONE 2019, 14, e0225584. [Google Scholar] [CrossRef] [PubMed]
- LeBel, G.; Haas, B.; Adam, A.-A.; Veilleux, M.-P.; Lagha, A.B.; Grenier, D. Effect of cinnamon (Cinnamomum verum) bark essential oil on the halitosis-associated bacterium Solobacterium moorei and in vitro cytotoxicity. Arch. Oral Biol. 2017, 83, 97–104. [Google Scholar] [CrossRef]
- Söderling, E.M.; Ekman, T.C.; Taipale, T.J. Growth inhibition of Streptococcus mutans with low xylitol concentrations. Curr. Microbiol. 2008, 56, 382–385. [Google Scholar] [CrossRef]
- Ferrazzano, G.F.; Amato, I.; Ingenito, A.; Zarrelli, A.; Pinto, G.; Pollio, A. Plant polyphenols and their anti-cariogenic properties: A review. Molecules 2011, 16, 1486–1507. [Google Scholar] [CrossRef]
- Hasibul, K.; Nakayama-Imaohji, H.; Hashimoto, M.; Yamasaki, H.; Ogawa, T.; Waki, J.; Tada, A.; Yoneda, S.; Tokuda, M.; Miyake, M. D-Tagatose inhibits the growth and biofilm formation of Streptococcus mutans. Mol. Med. Rep. 2018, 17, 843–851. [Google Scholar]
- Nakahara, K.; Kawabata, S.; Ono, H.; Ogura, K.; Tanaka, T.; Ooshima, T.; Hamada, S. Inhibitory effect of oolong tea polyphenols on glycosyltransferases of mutans Streptococci. Appl. Environ. Microbiol. 1993, 59, 968–973. [Google Scholar] [CrossRef]
- Yabuta, Y.; Sato, Y.; Miki, A.; Nagata, R.; Bito, T.; Ishihara, A.; Watanabe, F. Lemon myrtle extract inhibits lactate production by Streptococcus mutans. Biosci. Biotechnol. Biochem. 2021, 85, 2185–2190. [Google Scholar] [CrossRef]
- Jeong, S.-I.; Kim, B.-S.; Keum, K.-S.; Lee, K.-H.; Kang, S.-Y.; Park, B.-I.; Lee, Y.-R.; You, Y.-O. Kaurenoic acid from Aralia continentalis inhibits biofilm formation of Streptococcus mutans. Evid.-Based Complement. Altern. Med. 2013, 2013, 160592. [Google Scholar] [CrossRef] [PubMed]
- Bowen, W.; Koo, H. Biology of Streptococcus mutans-derived glucosyltransferases: Role in extracellular matrix formation of cariogenic biofilms. Caries Res. 2011, 45, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Katsura, H.; Tsukiyama, R.-I.; Suzuki, A.; Kobayashi, M. In vitro antimicrobial activities of bakuchiol against oral microorganisms. Antimicrob. Agents Chemother. 2001, 45, 3009–3013. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Xia, K.; Su, X.; Zheng, X.; Diao, C.; Yang, X.; Zuo, X.; Xu, J.; Liang, X. Mechanistic insights into the inhibitory effect of theaflavins on virulence factors production in Streptococcus mutans. AMB Express 2021, 11, 102. [Google Scholar] [CrossRef] [PubMed]
- Abranches, J.; Zeng, L.; Kajfasz, J.K.; Palmer, S.; Chakraborty, B.; Wen, Z.; Richards, V.P.; Brady, L.J.; Lemos, J.A. Biology of oral streptococci. Microbiol. Spectr. 2018, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, A.; Sarveswari, H.B.; Vasudevan, S.; Shanmugam, K.; Solomon, A.P.; Neelakantan, P. Baicalein inhibits Streptococcus mutans biofilms and dental caries-related virulence phenotypes. Antibiotics 2021, 10, 215. [Google Scholar] [CrossRef]
- Vaillancourt, K.; LeBel, G.; Pellerin, G.; Ben Lagha, A.; Grenier, D. Effects of the licorice isoflavans licoricidin and glabridin on the growth, adherence properties, and acid production of Streptococcus mutans, and assessment of their biocompatibility. Antibiotics 2021, 10, 163. [Google Scholar] [CrossRef]
- Wang, Y.; Lam, A.T. Epigallocatechin gallate and gallic acid affect colonization of abiotic surfaces by oral bacteria. Arch. Oral Biol. 2020, 120, 104922. [Google Scholar] [CrossRef]
- Yang, R.-Q.; Zhao, G.-P. Inhibitory Effects of Glycyrrhiza uralensis Fisch Extract on Cariogenic Virulence Factors of Streptococcus mutans. Indian J. Microbiol. 2022, 62, 473. [Google Scholar] [CrossRef]
- Higham, S.; Edgar, M. Effects of lactate dehydrogenase and nicotinamide adenine dinucleotide on human dental plaque pH and acid anion concentrations. Arch. Oral Biol. 1995, 40, 55–59. [Google Scholar] [CrossRef]
- Fujiwara, T.; Hoshino, T.; Ooshima, T.; Hamada, S. Differential and quantitative analyses of mRNA expression of glucosyltransferases from Streptococcus mutans MT8148. J. Dent. Res. 2002, 81, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Avilés-Reyes, A.; Miller, J.H.; Lemos, J.A.; Abranches, J. Collagen-binding proteins of Streptococcus mutans and related streptococci. Mol. Oral Microbiol. 2017, 32, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Duque, C.; Stipp, R.N.; Wang, B.; Smith, D.J.; Höfling, J.F.; Kuramitsu, H.K.; Duncan, M.J.; Mattos-Graner, R.O. Downregulation of GbpB, a component of the VicRK regulon, affects biofilm formation and cell surface characteristics of Streptococcus mutans. Infect. Immun. 2011, 79, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Ling, J. Streptococcus gordonii LuxS/autoinducer-2 quorum-sensing system modulates the dual-species biofilm formation with Streptococcus mutans. J. Basic Microbiol. 2017, 57, 605–616. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) |
|---|---|
| ldh | F:ACTTCACTTGATACTGCTCGTT |
| R:ACACCAGCTACATTGGCATGA | |
| gtfB | F:ACGAACTTTGCCGTTATTGTCA |
| R:AGCAATGCAGCCAATCTACAA | |
| gtfC | F:CTCAACCAACCGCCACTGTT |
| R:GGTTTAACGTCAAAATTAGCTGTATTAG | |
| gtfD | F:TGTCTTGGTGGCCAGATAAAC |
| R:GAACGGTTTGTGCAGCAAGG | |
| ftf | F:CCTGCGACTTCATTACGATTGGTC |
| R:ATTGGCGAACGGCGACTTACTC | |
| spaP | F;TGGGATAGTTCAGATGCGCC |
| R;AGGATCAGCAGGCACAACAA | |
| srtA | F:TGGCAATTCCGCCAATTACAG |
| R;AAGGCTGCCCATTCTTCCTT | |
| luxs | F:GCTTTGATGACTGTGGCTATTTG |
| R:ACTGCAGGCCTTCATACTATTG | |
| comD | F:CGCGATTGGAGCCTTTAG |
| R:CCTGAAATTCAGTTAGCCTTT | |
| Come | F:GGCTACTTCCAGTCCTTTCTTT |
| R:AACCACCATTGCAGCTATCA | |
| gbpB | F:ATGGCGGTTATGGACACGTT |
| R:TTTGGCCACCTTGAACACCT | |
| brpA | F:GGAGGAGCTGCATCAGGATTC |
| R:AACTCCAGCACATCCAGCAAG | |
| 16s RNA | F:CCTACGGGAGGCAGCAGTAG |
| R:CAACAGAGCTTTACGATCCGAAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, E.; Li, S.; Wang, S.; Li, Q.; Pang, D.; Yang, Q.; Zhu, Q.; Zou, Y. Antibacterial Effects of Ramulus mori Oligosaccharides against Streptococcus mutans. Foods 2023, 12, 3182. https://doi.org/10.3390/foods12173182
Li E, Li S, Wang S, Li Q, Pang D, Yang Q, Zhu Q, Zou Y. Antibacterial Effects of Ramulus mori Oligosaccharides against Streptococcus mutans. Foods. 2023; 12(17):3182. https://doi.org/10.3390/foods12173182
Chicago/Turabian StyleLi, Erna, Shipei Li, Siyuan Wang, Qian Li, Daorui Pang, Qiong Yang, Qiaoling Zhu, and Yuxiao Zou. 2023. "Antibacterial Effects of Ramulus mori Oligosaccharides against Streptococcus mutans" Foods 12, no. 17: 3182. https://doi.org/10.3390/foods12173182
APA StyleLi, E., Li, S., Wang, S., Li, Q., Pang, D., Yang, Q., Zhu, Q., & Zou, Y. (2023). Antibacterial Effects of Ramulus mori Oligosaccharides against Streptococcus mutans. Foods, 12(17), 3182. https://doi.org/10.3390/foods12173182