
Dietary Polysaccharides Exert Anti-Fatigue Functions via the Gut-Muscle Axis: Advances and Prospectives

 , , and
, , and
Abstract
:1. Introduction
2. Pathogenesis of Exercise-Induced Fatigue
2.1. Pathogenesis of Peripheral Fatigue
2.2. Pathogenesis of Central Fatigue
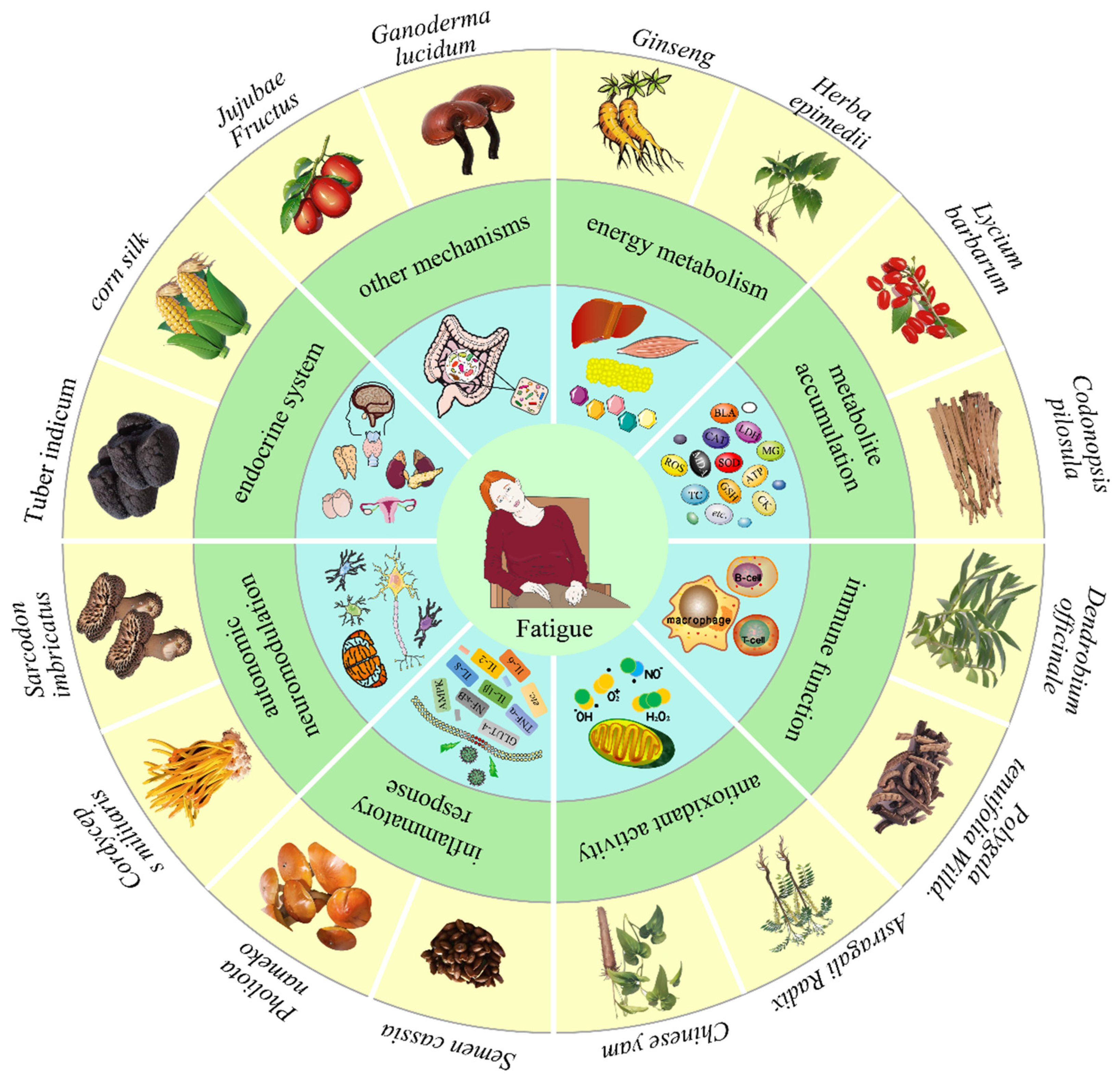
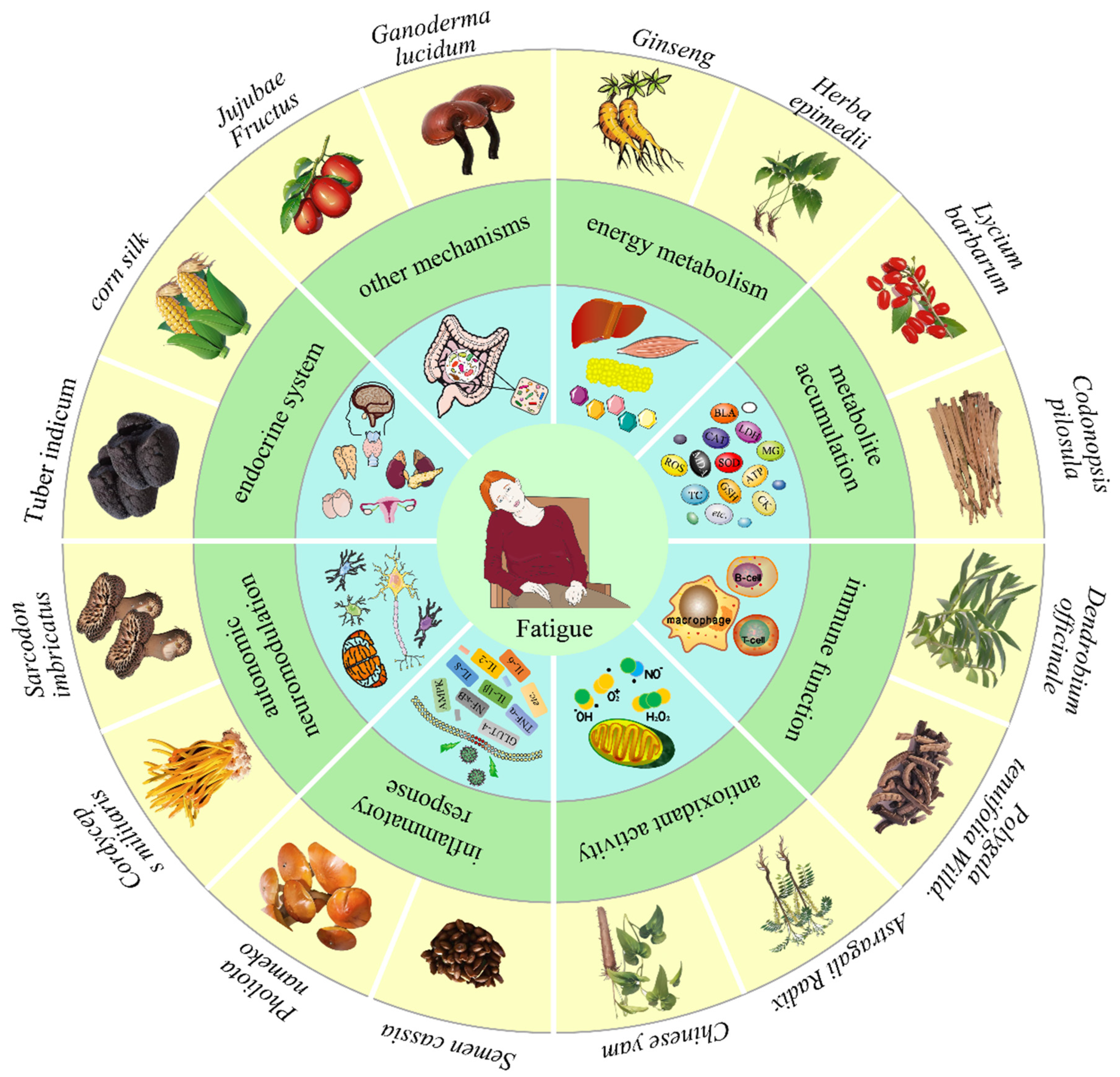
3. Polysaccharides Ameliorate Fatigue
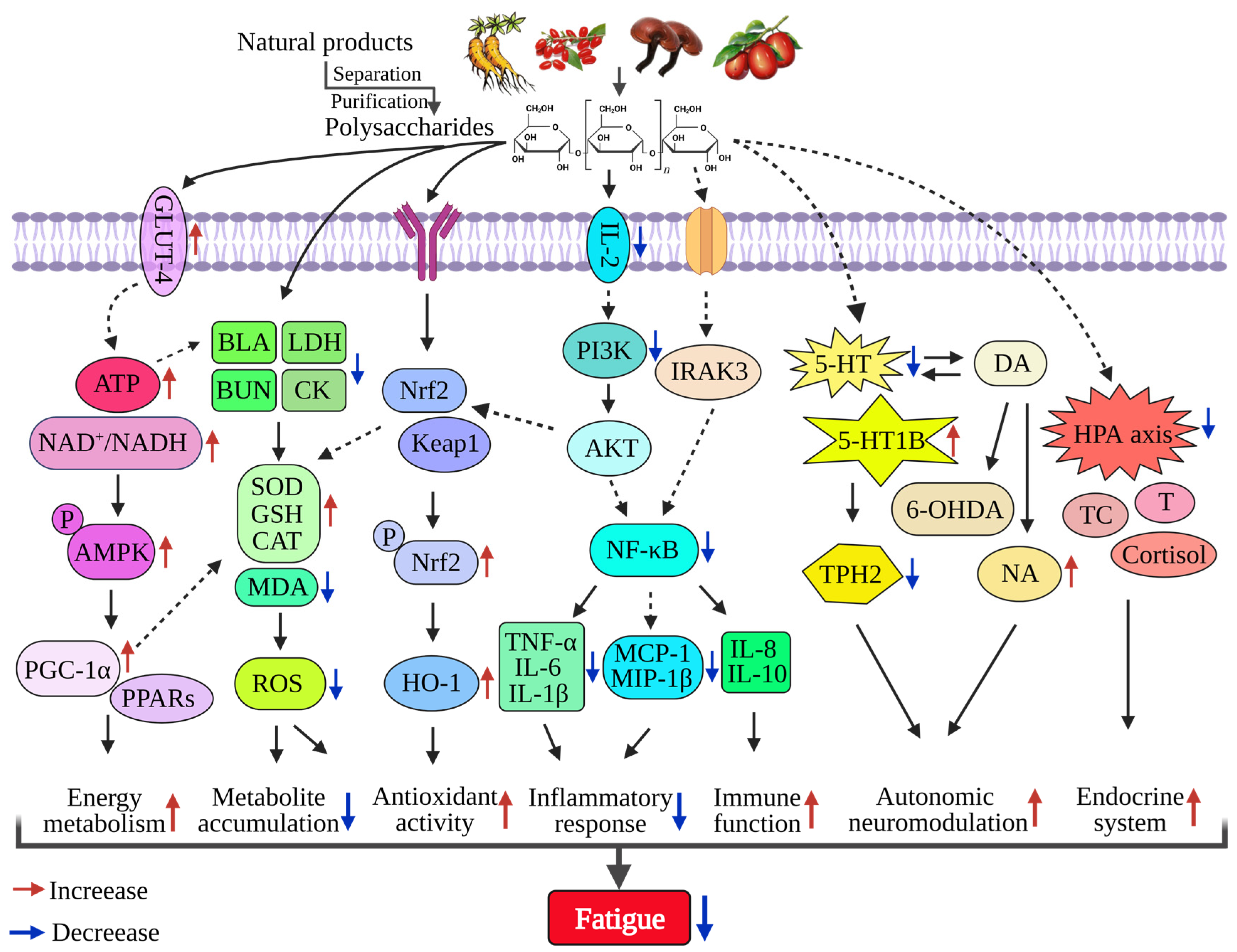
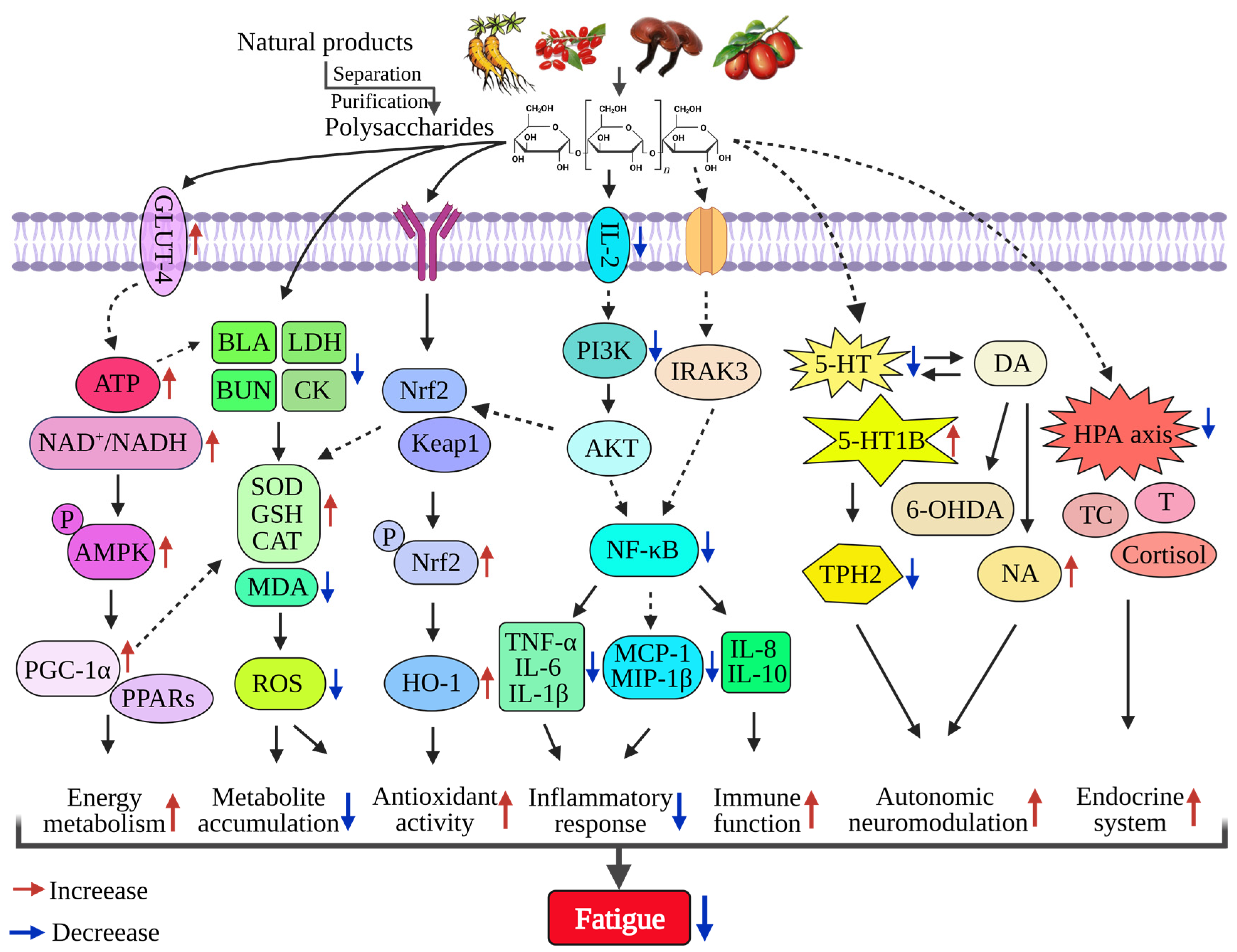
3.1. Impacts on Energy Metabolism
3.2. Reduce the Accumulation of Metabolites
3.3. Improve Immune Function
3.4. Enhance Antioxidant Activity
3.5. Inhibit Inflammatory Response
3.6. Interfere with Autonomic Neuromodulation
3.7. Regulate Endocrine System
3.8. Other Anti-Fatigue Mechanisms
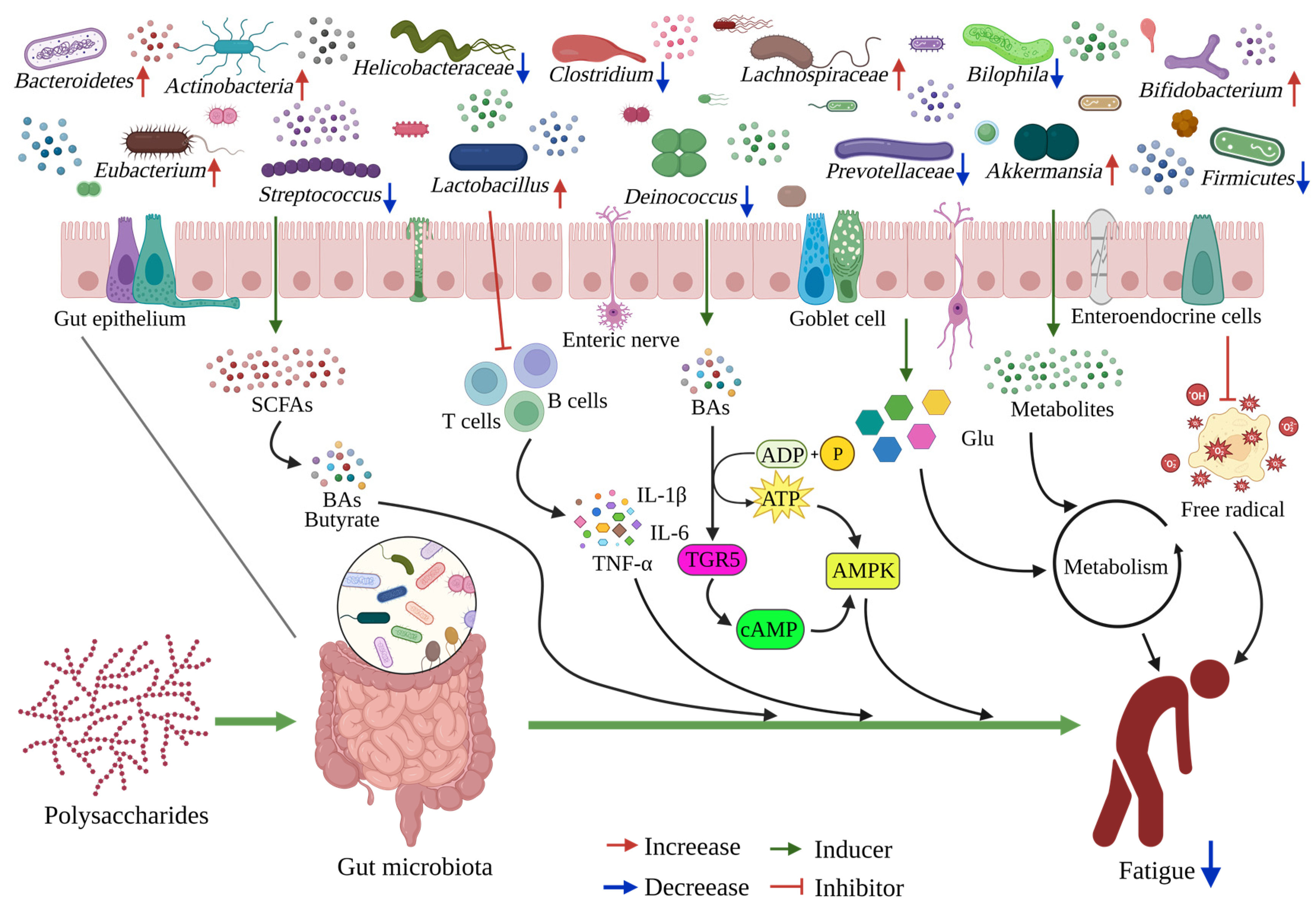
4. Polysaccharides Ameliorate Fatigue via the Gut-Muscle Axis
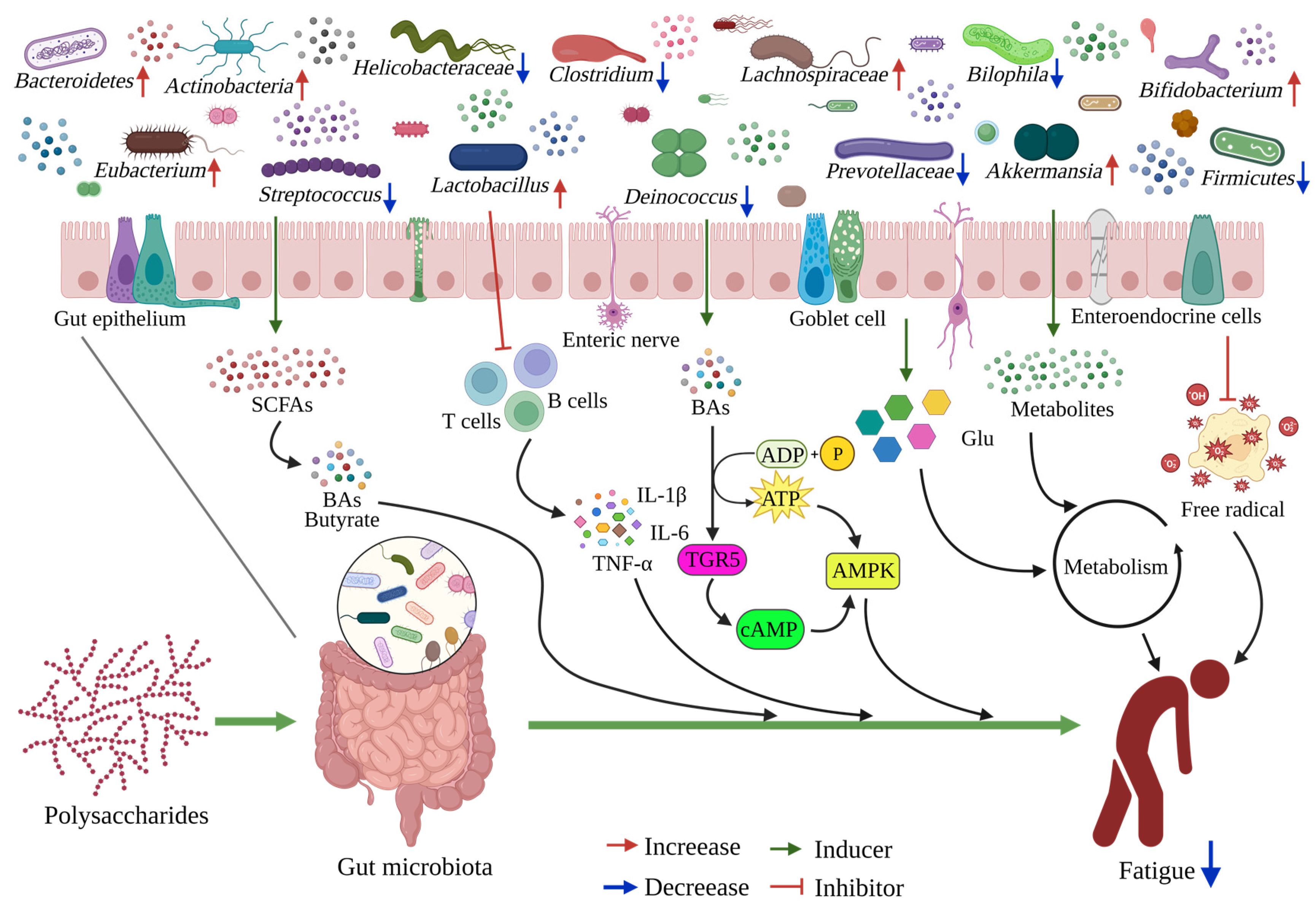
4.1. Polysaccharide Interferes with Gut Microbiome
4.2. Polysaccharides Ameliorate Fatigue via Gut Microbiota
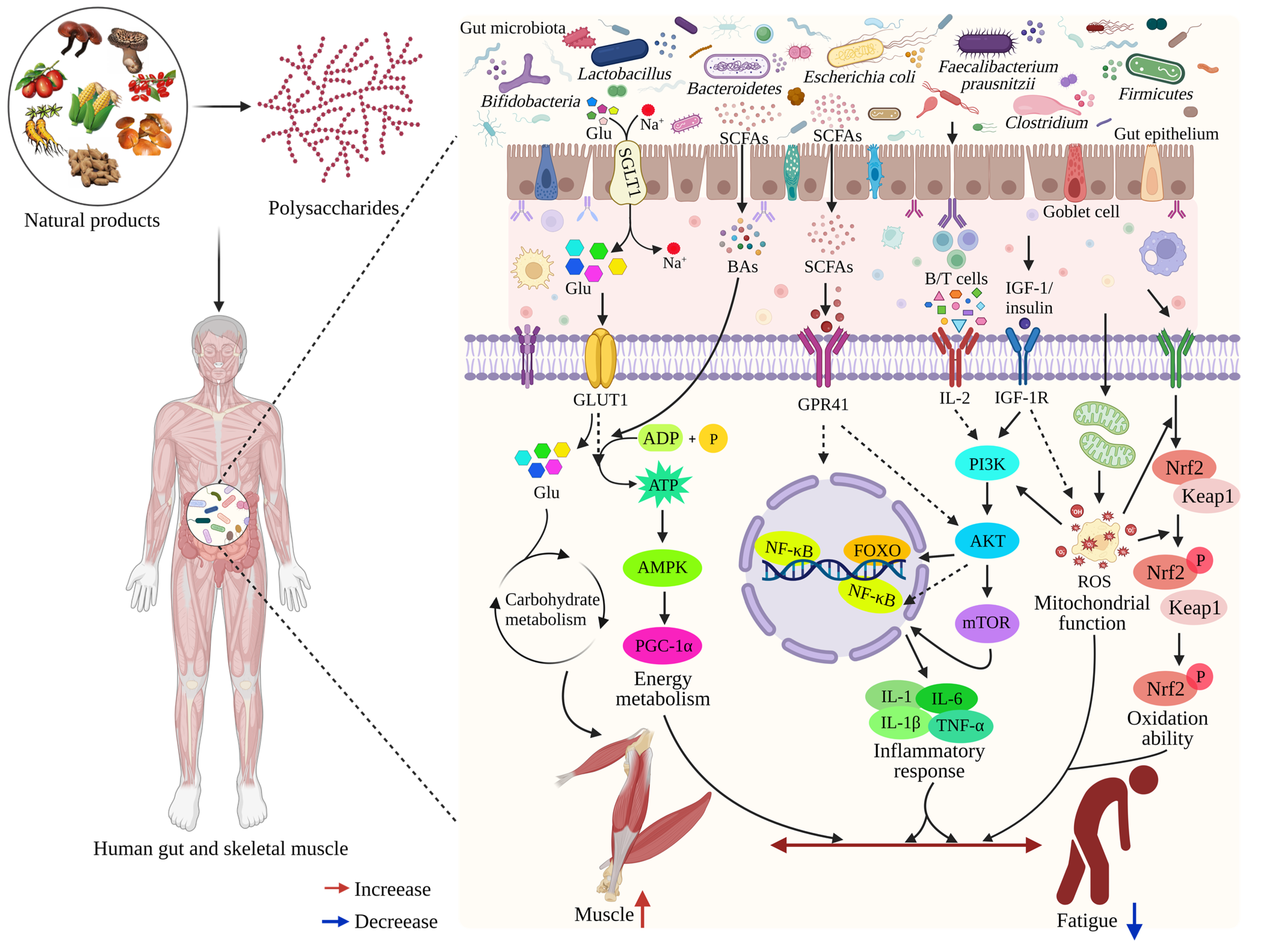
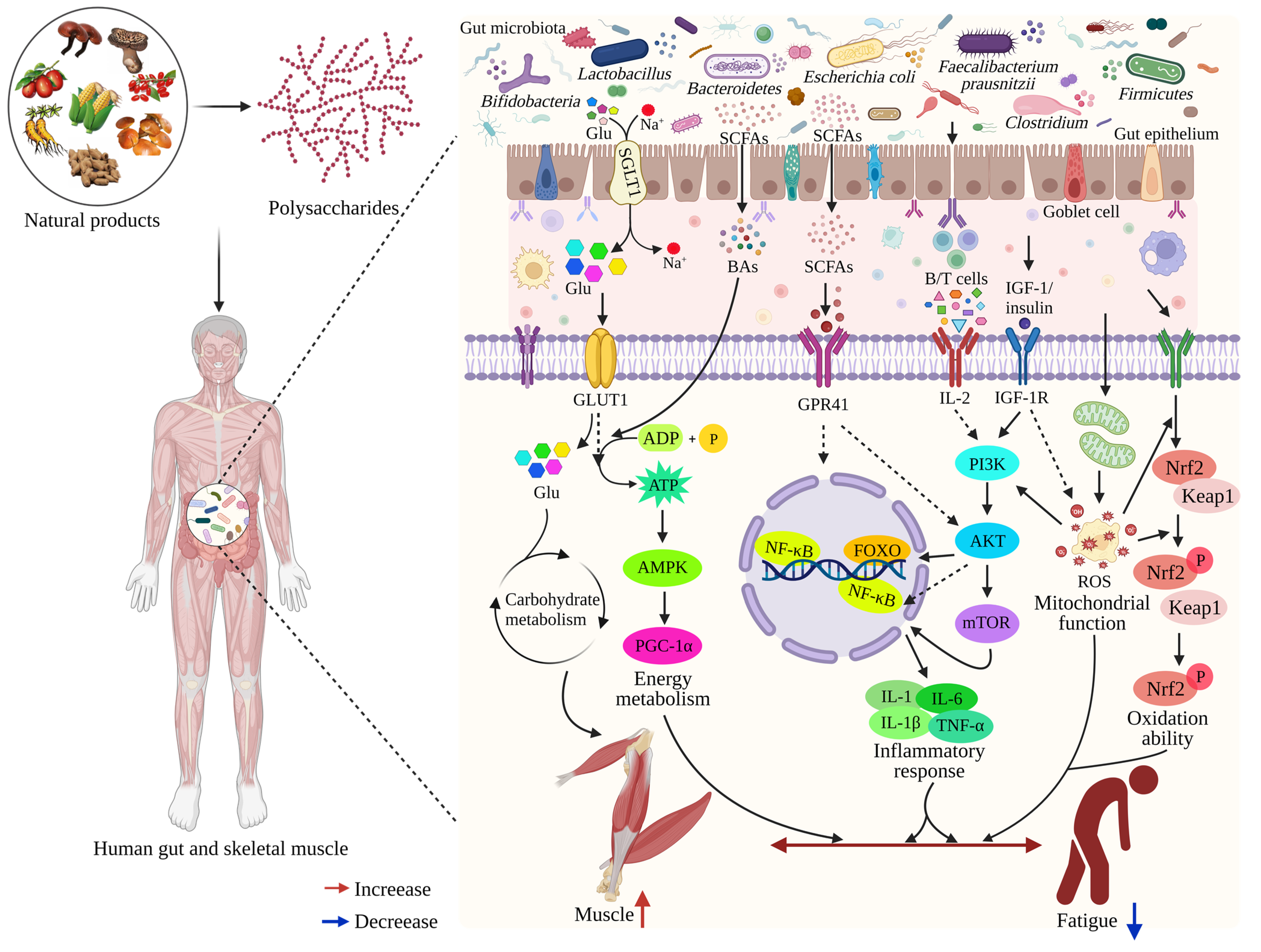
4.3. Relationship between Gut Microbiota and Muscle
4.4. Impact of the Gut-Muscle Axis on Muscle Movement Ability
4.5. Polysaccharide Interactions with the Gut-Muscle Axis
4.6. Polysaccharides Relieve Fatigue by Gut-Muscle Axis
5. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Finsterer, J.; Mahjoub, S.Z. Fatigue in healthy and diseased individuals. Am. J. Hosp. Palliative Care 2014, 31, 562–575. [Google Scholar] [CrossRef]
- Matura, L.A.; Malone, S.; Jaime-Lara, R.; Riegel, B. A systematic review of biological mechanisms of fatigue in chronic illness. Biol. Res. Nurs. 2018, 20, 410–421. [Google Scholar] [CrossRef]
- Luo, C.; Xu, X.; Wei, X.; Feng, W.; Huang, H.; Liu, H.; Xu, R.; Lin, J.; Han, L.; Zhang, D. Natural medicines for the treatment of fatigue: Bioactive components, pharmacology, and mechanisms. Pharmacol. Res. 2019, 148, 104409. [Google Scholar] [CrossRef]
- Kizina, K.; Stolte, B.; Totzeck, A.; Bolz, S.; Schlag, M.; Ose, C.; von Velsen, O.; Kleinschnitz, C.; Hagenacker, T. Fatigue in adults with spinal muscular atrophy under treatment with nusinersen. Sci. Rep. 2020, 10, 11069. [Google Scholar] [CrossRef]
- Bower, J.E. Cancer-related fatigue—Mechanisms, risk factors, and treatments. Nat. Rev. Clin. Oncol. 2014, 11, 597–609. [Google Scholar] [CrossRef]
- Knoop, V.; Cloots, B.; Costenoble, A.; Debain, A.; Vella Azzopardi, R.; Vermeiren, S.; Jansen, B.; Scafoglieri, A.; Bautmans, I.; Gerontopole Brussels Study Group. Fatigue and the prediction of negative health outcomes: A systematic review with meta-analysis. Ageing Res. Rev. 2021, 67, 101261. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Zhang, L.; Li, X.; Kan, Y.; Li, S.; Zou, Y.; Liu, L.; Yuan, Y.; Gong, W.; Zhang, Y. Contributing factors of fatigue in patients with type 2 diabetes: A systematic review. Psychoneuroendocrinology 2021, 130, 105280. [Google Scholar] [CrossRef] [PubMed]
- Penner, I.K.; Paul, F. Fatigue as a symptom or comorbidity of neurological diseases. Nat. Rev. Neurol. 2017, 13, 662–675. [Google Scholar] [CrossRef]
- Liu, Y.; Li, C.; Shen, X.; Liu, Y. The use of traditional Chinese medicines in relieving exercise-induced fatigue. Front. Pharmacol. 2022, 13, 969827. [Google Scholar] [CrossRef]
- Guo, T.; Akan, O.D.; Luo, F.; Lin, Q. Dietary polysaccharides exert biological functions via epigenetic regulations: Advance and prospectives. Crit. Rev. Food Sci. Nutr. 2023, 63, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Dedhia, N.; Marathe, S.J.; Singhal, R.S. Food polysaccharides: A review on emerging microbial sources, bioactivities, nanoformulations and safety considerations. Carbohydr. Polym. 2022, 287, 119355. [Google Scholar] [CrossRef]
- Álvarez-Mercado, A.I.; Plaza-Diaz, J. Dietary polysaccharides as modulators of the gut microbiota ecosystem: An update on their impact on health. Nutrients 2022, 14, 4116. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.S.; Jiang, J.G. Anti-fatigue effects of active ingredients from traditional Chinese medicine: A review. Curr. Med. Chem. 2019, 26, 1833–1848. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Liu, Y.; Chi, A.; Zhang, Z. The anti-fatigue potential of water-soluble polysaccharides of Semen cassiae on BALB/c mice. Cell. Mol. Biol. (Noisy-le-grand) 2021, 67, 148–154. [Google Scholar] [CrossRef]
- Cai, M.; Xing, H.; Tian, B.; Xu, J.; Li, Z.; Zhu, H.; Yang, K.; Sun, P. Characteristics and antifatigue activity of graded polysaccharides from Ganoderma lucidum separated by cascade membrane technology. Carbohydr. Polym. 2021, 269, 118329. [Google Scholar] [CrossRef]
- Zhu, H.; Xu, W.; Wang, N.; Jiang, W.; Cheng, Y.; Guo, Y.; Yao, W.; Hu, B.; Du, P.; Qian, H. Anti-fatigue effect of Lepidium meyenii Walp. (Maca) on preventing mitochondria-mediated muscle damage and oxidative stress in vivo and vitro. Food Funct. 2021, 12, 3132–3141. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Li, J.; Liu, F.; Wang, J.; Jiang, P.; Li, B.; Li, H.; Chen, C.; Wu, W. Characterisation, chain conformation and antifatigue effect of steamed ginseng polysaccharides with different molecular weight. Front. Pharmacol. 2021, 12, 712836. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Zhang, Y.; Huo, Z.; Wang, G.; He, Y.; Man, S.; Gao, W. Effects of the polysaccharides extracted from Chinese yam (Dioscorea opposita Thunb.) on cancer-related fatigue in mice. Food Funct. 2021, 12, 10602–10614. [Google Scholar] [CrossRef]
- Zhang, C.J.; Guo, J.Y.; Cheng, H.; Li, L.; Liu, Y.; Shi, Y.; Xu, J.; Yu, H.T. Spatial structure and anti-fatigue of polysaccharide from Inonotus obliquus. Int. J. Biol. Macromol. 2020, 151, 855–860. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, M.; Wang, X.; Ren, Y.; Yue, T.; Wang, Z.; Gao, Z. Edible fungal polysaccharides, the gut microbiota, and host health. Carbohydr. Polym. 2021, 273, 118558. [Google Scholar]
- Zhou, S.S.; Zhou, J.; Xu, J.D.; Shen, H.; Kong, M.; Yip, K.M.; Han, Q.B.; Zhao, Z.Z.; Xu, J.; Chen, H.B.; et al. Ginseng ameliorates exercise-induced fatigue potentially by regulating the gut microbiota. Food Funct. 2021, 12, 3954–3964. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, R.; Hua, H.; Cheng, Y.; Guo, Y.; Qian, H.; Du, P. Network pharmacology exploration reveals gut microbiota modulation as a common therapeutic mechanism for anti-fatigue effect treated with maca compounds prescription. Nutrients 2022, 14, 1533. [Google Scholar] [CrossRef] [PubMed]
- Giron, M.; Thomas, M.; Dardevet, D.; Chassard, C.; Savary-Auzeloux, I. Gut microbes and muscle function: Can probiotics make our muscles stronger? J. Cachexia Sarcopenia Muscle 2022, 13, 1460–1476. [Google Scholar] [CrossRef]
- Grosicki, G.J.; Fielding, R.A.; Lustgarten, M.S. Gut microbiota contribute to age-related changes in skeletal muscle size, composition, and function: Biological basis for a gut-muscle axis. Calcif. Tissue Int. 2018, 102, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Lustgarten, M.S. The role of the gut microbiome on skeletal muscle mass and physical function: 2019 update. Front. Physiol. 2019, 10, 1435. [Google Scholar] [CrossRef]
- Ticinesi, A.; Mancabelli, L.; Tagliaferri, S.; Nouvenne, A.; Milani, C.; Del Rio, D.; Lauretani, F.; Maggio, M.G.; Ventura, M.; Meschi, T. The gut-muscle axis in older subjects with low muscle mass and performance: A proof of concept study exploring fecal microbiota composition and function with shotgun metagenomics sequencing. Int. J. Mol. Sci. 2020, 21, 8946. [Google Scholar] [CrossRef] [PubMed]
- Jamshidi, S.; Masoumi, S.J.; Abiri, B.; Vafa, M. The effects of synbiotic and/or vitamin D supplementation on gut-muscle axis in overweight and obese women: A study protocol for a double-blind, randomized, placebo-controlled trial. Trials 2022, 23, 631. [Google Scholar] [CrossRef]
- Zhao, J.; Huang, Y.; Yu, X. A narrative review of gut-muscle axis and sarcopenia: The potential role of gut microbiota. Int. J. Gen. Med. 2021, 14, 1263–1273. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, R.; Hua, H.; Qian, H.; Du, P. Deciphering the potential role of Maca compounds prescription influencing gut microbiota in the management of exercise-induced fatigue by integrative genomic analysis. Front. Nutr. 2022, 9, 1004174. [Google Scholar] [CrossRef]
- Cai, M.; Zhu, H.; Xu, L.; Wang, J.; Xu, J.; Li, Z.; Yang, K.; Wu, J.; Sun, P. Structure, anti-fatigue activity and regulation on gut microflora in vivo of ethanol-fractional polysaccharides from Dendrobium officinale. Int. J. Biol. Macromol. 2023, 234, 123572. [Google Scholar] [CrossRef]
- Guo, Q.; Xu, L.; Chen, Y.; Ma, Q.; Santhanam, R.K.; Xue, Z.; Gao, X.; Chen, H. Structural characterization of corn silk polysaccharides and its effect in H2O2 induced oxidative damage in L6 skeletal muscle cells. Carbohydr. Polym. 2019, 208, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.L.; Li, X.P.; Zhu, Y.L.; Yi, G.Q.; Wang, W.; Chen, X.Y.; Deng, G.M.; Yang, L.; Cai, H.Z.; Tong, Q.Z.; et al. Polygonatum sibiricum polysaccharides (PSP) improve the palmitic acid (PA)-induced inhibition of survival, inflammation, and glucose uptake in skeletal muscle cells. Bioengineered 2021, 12, 10147–10159. [Google Scholar] [CrossRef]
- Meeusen, R.; Van Cutsem, J.; Roelands, B. Endurance exercise-induced and mental fatigue and the brain. Exp. Physiol. 2021, 106, 2294–2298. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; Gao, H.; Liu, X.; Zhao, Z.; Gao, B.; Zhang, L. Establishment and identification of an animal model of long-term exercise-induced fatigue. Front. Endocrinol. 2022, 13, 915937. [Google Scholar] [CrossRef] [PubMed]
- Tornero-Aguilera, J.F.; Jimenez-Morcillo, J.; Rubio-Zarapuz, A.; Clemente-Suárez, V.J. Central and peripheral fatigue in physical exercise explained: A narrative review. Int. J. Environ. Res. Public Health 2022, 19, 3909. [Google Scholar] [CrossRef]
- Filippi, M.; Krähenmann, R.; Fissler, P. The link between energy-related sensations and metabolism: Implications for treating fatigue. Front. Psychol. 2022, 13, 920556. [Google Scholar] [CrossRef]
- Giuriato, G.; Venturelli, M.; Matias, A.; Soares, E.; Gaetgens, J.; Frederick, K.A.; Ives, S.J. Capsaicin and its effect on exercise performance, fatigue and inflammation after exercise. Nutrients 2022, 14, 232. [Google Scholar] [CrossRef]
- Wang, P.; Wang, D.; Hu, J.; Tan, B.K.; Zhang, Y.; Lin, S. Natural bioactive peptides to beat exercise-induced fatigue: A review. Food Biosci. 2021, 43, 101298. [Google Scholar] [CrossRef]
- Medrado de Barcellos, L.A.; Gonçalves, W.A.; Esteves de Oliveira, M.P.; Guimarães, J.B.; Queiroz-Junior, C.M.; de Resende, C.B.; Russo, R.C.; Coimbra, C.C.; Silva, A.N.; Teixeira, M.M.; et al. Effect of physical training on exercise-induced inflammation and performance in mice. Front. Cell Dev. Biol. 2021, 9, 625680. [Google Scholar] [CrossRef]
- Davies, K.; Dures, E.; Ng, W.F. Fatigue in inflammatory rheumatic diseases: Current knowledge and areas for future research. Nat. Rev. Rheumatol. 2021, 17, 651–664. [Google Scholar] [CrossRef]
- Wei, W.; Li, Z.P.; Zhu, T.; Fung, H.Y.; Wong, T.L.; Wen, X.; Ma, D.L.; Leung, C.H.; Han, Q.B. Anti-fatigue effects of the unique polysaccharide marker of Dendrobium officinale on BALB/c Mice. Molecules 2017, 22, 155. [Google Scholar] [CrossRef]
- Magherini, F.; Fiaschi, T.; Marzocchini, R.; Mannelli, M.; Gamberi, T.; Modesti, P.A.; Modesti, A. Oxidative stress in exercise training: The involvement of inflammation and peripheral signals. Free Radic. Res. 2019, 53, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Deminice, R.; Ozdemir, M.; Yoshihara, T.; Bomkamp, M.P.; Hyatt, H. Exercise-induced oxidative stress: Friend or foe? J. Sport Health Sci. 2020, 9, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qu, Y.; Zhang, Y.; Li, S.; Sun, Y.; Chen, Z.; Teng, L.; Wang, D. Antifatigue potential activity of Sarcodon imbricatus in acute excise-treated and chronic fatigue syndrome in mice via regulation of Nrf2-mediated oxidative stress. Oxid. Med. Cell. Longev. 2018, 2018, 9140896. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Liu, Z.; Wang, X.; Wang, C. Recent advances in Panax ginseng C.A. Meyer as a herb for anti-fatigue: An effects and mechanisms review. Foods 2021, 10, 1030. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.; Liu, Y.; Zhang, Z. Effects of exercise-induced ROS on the pathophysiological functions of skeletal muscle. Oxid. Med. Cell. Longev. 2021, 2021, 3846122. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, B. Mechanism of hyperbaric oxygen combined with Astaxanthin mediating Keap1/Nrf2/HO-1 pathway to improve exercise fatigue in mice. Comput. Intell. Neurosci. 2022, 2022, 6444747. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, R.; Hua, H.; Cheng, Y.; Guo, Y.; Qian, H.; Du, P. The macamide relieves fatigue by acting as inhibitor of inflammatory response in exercising mice: From central to peripheral. Eur. J. Pharmacol. 2022, 917, 174758. [Google Scholar] [CrossRef]
- Constantin-Teodosiu, D.; Constantin, D. Molecular mechanisms of muscle fatigue. Int. J. Mol. Sci. 2021, 22, 11587. [Google Scholar] [CrossRef]
- Amann, M.; Sidhu, S.K.; McNeil, C.J.; Gandevia, S.C. Critical considerations of the contribution of the corticomotoneuronal pathway to central fatigue. J. Physiol. 2022, 600, 5203–5214. [Google Scholar] [CrossRef]
- Thorstensen, J.R.; Taylor, J.L.; Tucker, M.G.; Kavanagh, J.J. Enhanced serotonin availability amplifies fatigue perception and modulates the TMS-induced silent period during sustained low-intensity elbow flexions. J. Physiol. 2020, 598, 2685–2701. [Google Scholar] [CrossRef] [PubMed]
- Henderson, T.T.; Taylor, J.L.; Thorstensen, J.R.; Tucker, M.G.; Kavanagh, J.J. Enhanced availability of serotonin limits muscle activation during high-intensity, but not low-intensity, fatiguing contractions. J. Neurophysiol. 2022, 128, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, S. Dopamine surprises. Nat. Neurosci. 2022, 25, 531. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, D.D.L.; Freitas, F.C.; Aguiar, A.S.; Ward, C., Jr.; Guglielmo, L.G.A.; Prediger, R.D.; Cronin, S.J.F.; Walz, R.; Andrews, N.A.; Latini, A. Impaired dopamine metabolism is linked to fatigability in mice and fatigue in Parkinson’s disease patients. Brain Commun. 2021, 3, fcab116. [Google Scholar] [CrossRef]
- Caravaggio, F.; Barnett, A.J.; Nakajima, S.; Iwata, Y.; Kim, J.; Borlido, C.; Mar, W.; Gerretsen, P.; Remington, G.; Graff-Guerrero, A. The effects of acute dopamine depletion on resting-state functional connectivity in healthy humans. Eur. Neuropsychopharmacol. 2022, 57, 39–49. [Google Scholar] [CrossRef]
- Kok, A. Cognitive control, motivation and fatigue: A cognitive neuroscience perspective. Brain Cogn. 2022, 160, 105880. [Google Scholar] [CrossRef]
- Rabelo, P.C.R.; Cordeiro, L.M.S.; Aquino, N.S.S.; Fonseca, B.B.B.; Coimbra, C.C.; Wanner, S.P.; Szawka, R.E.; Soares, D.D. Rats with higher intrinsic exercise capacities exhibit greater preoptic dopamine levels and greater mechanical and thermoregulatory efficiencies while running. J. Appl. Physiol. 2019, 126, 393–402. [Google Scholar] [CrossRef]
- Welsch, P.; Üçeyler, N.; Klose, P.; Walitt, B.; Häuser, W. Serotonin and noradrenaline reuptake inhibitors (SNRIs) for fibromyalgia. Cochrane Database Syst. Rev. 2018, 2, CD010292. [Google Scholar]
- Xu, Y.; Lian, Y.; Li, J.; Zhang, Y.; Liu, Y.; Wang, X.; Ma, J.; Li, F. KangPiLao decoction modulates cognitive and emotional disorders in rats with central fatigue through the GABA/Glu pathway. Front. Pharmacol. 2022, 13, 939169. [Google Scholar] [CrossRef]
- Rönnbäck, L.; Johansson, B. Long-lasting pathological mental fatigue after brain injury-a dysfunction in glutamate neurotransmission? Front. Behav. Neurosci. 2022, 15, 791984. [Google Scholar] [CrossRef]
- Fortes, L.S.; Faro, H.; de Lima-Junior, D.; Albuquerque, M.R.; Ferreira, M.E.C. Non-invasive brain stimulation over the orbital prefrontal cortex maintains endurance performance in mentally fatigued swimmers. Physiol. Behav. 2022, 250, 113783. [Google Scholar] [CrossRef]
- Ayache, S.S.; Serratrice, N.; Abi Lahoud, G.N.; Chalah, M.A. Fatigue in multiple sclerosis: A review of the exploratory and therapeutic potential of non-invasive brain stimulation. Front. Neurol. 2022, 13, 813965. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.; Mach, N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: A systematic review for athletes. J. Int. Soc. Sports Nutr. 2016, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Peng, X.; Zhou, Z.; Tang, C.; Liu, W. Effects of Bushen Yiyuan recipe on testosterone synthesis in Leydig cells of rats with exercise-induced low serum testosterone levels. Pharm. Biol. 2022, 60, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- Arazi, H.; Aboutalebi, S.; Taati, B.; Cholewa, J.M.; Candow, D.G. Effects of short-term betaine supplementation on muscle endurance and indices of endocrine function following acute high-intensity resistance exercise in young athletes. J. Int. Soc. Sports Nutr. 2022, 19, 1–16. [Google Scholar] [CrossRef]
- Bansal, R.; Gubbi, S.; Koch, C.A. COVID-19 and chronic fatigue syndrome: An endocrine perspective. J. Clin. Transl. Endocrinol. 2022, 27, 100284. [Google Scholar] [CrossRef]
- Mukherjee, S.; Jana, S.; Khawas, S.; Kicuntod, J.; Marschall, M.; Ray, B.; Ray, S. Synthesis, molecular features and biological activities of modified plant polysaccharides. Carbohydr. Polym. 2022, 289, 119299. [Google Scholar] [CrossRef]
- Peng, Y.; Zhao, L.; Hu, K.; Yang, Y.; Ma, J.; Zhai, Y.; Jiang, Y.; Zhang, D. Anti-fatigue effects of Lycium barbarum polysaccharide and effervescent tablets by regulating oxidative stress and energy metabolism in rats. Int. J. Mol. Sci. 2022, 23, 10920. [Google Scholar] [CrossRef]
- Bai, L.; Tan, C.; Ren, J.; Liu, J.; Zou, W.; Liu, G.; Sheng, Y. Cordyceps militaris acidic polysaccharides improve learning and memory impairment in mice with exercise fatigue through the PI3K/NRF2/HO-1 signalling pathway. Int. J. Biol. Macromol. 2023, 227, 158–172. [Google Scholar] [CrossRef]
- Li, C.; Zhu, X.; Zhang, J.; Xu, T.; Zhang, H.; Zheng, Z.; Kumar, R.R. Polysaccharides from apple pomace exhibit anti-fatigue activity through increasing glycogen content. J. Food Sci. Technol. 2023, 60, 283–291. [Google Scholar] [CrossRef]
- Zhao, H.P.; Zhang, Y.; Liu, Z.; Chen, J.Y.; Zhang, S.Y.; Yang, X.D.; Zhou, H.L. Acute toxicity and anti-fatigue activity of polysaccharide-rich extract from corn silk. Biomed. Pharmacother. 2017, 90, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Chi, A.; Shen, Z.; Zhu, W.; Sun, Y.; Kang, Y.; Guo, F. Characterization of a protein-bound polysaccharide from Herba Epimedii and its metabolic mechanism in chronic fatigue syndrome. J. Ethnopharmacol. 2017, 203, 241–251. [Google Scholar] [CrossRef]
- Tang, W.; Jin, L.; Xie, L.; Huang, J.; Wang, N.; Chu, B.; Dai, X.; Liu, Y.; Wang, R.; Zhang, Y. Structural characterization and antifatigue effect in vivo of Maca (Lepidium meyenii Walp.) polysaccharide. J. Food Sci. 2017, 82, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, Q.; Meng, Q.; Wang, L.; Xiong, W.; Zhang, L. Anti-fatigue activity of polysaccharide fractions from Lepidium meyenii Walp. (maca). Int. J. Biol. Macromol. 2017, 95, 1305–1311. [Google Scholar] [CrossRef]
- Chung, Y.H.; Park, T.K.; Yim, S.H.; Lee, J.H.; Bang, J.S.; Shin, Y.K.; Bae, J.; Sim, S.S.; Hwang, K.W.; Shin, C.Y.; et al. Polysaccharide-rich extract of Phragmites rhizome attenuates water immersion stress and forced swimming fatigue in rodent animal model. J. Med. Food 2019, 22, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Sun, Y.; Cao, L.; Chen, L.; Chen, J.; Cheng, X.; Wang, C. Antifatigue and antihypoxia activities of oligosaccharides and polysaccharides from Codonopsis pilosula in mice. Food Funct. 2020, 11, 6352–6362. [Google Scholar] [CrossRef]
- Xie, F. Effects of anti-fatigue in vivo and anti-oxidant in vitro of polysaccharides from Polygala tenuifolia Willd. on exhaustive exercise mice. Sci. Technol. Food Ind. 2021, 42, 332–336. [Google Scholar]
- Yu, Y.; Nie, J.; Zhao, B.; Tan, J.; Lv, C.; Lu, J. Structure characterization and anti-fatigue activity of an acidic polysaccharide from Panax ginseng C. A. Meyer. J. Ethnopharmacol. 2023, 301, 115831. [Google Scholar] [CrossRef]
- Liu, G.; Yang, X.; Zhang, J.; Liang, L.; Miao, F.; Ji, T.; Ye, Z.; Chu, M.; Ren, J.; Xu, X. Synthesis, stability and anti-fatigue activity of selenium nanoparticles stabilized by Lycium barbarum polysaccharides. Int. J. Biol. Macromol. 2021, 179, 418–428. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, B.; Yan, G.; Wu, H.; Han, Y.; Cui, H. Chemical properties and anti-fatigue effect of polysaccharide from Pholiota nameko. J. Food Biochem. 2022, 46, e14015. [Google Scholar] [CrossRef]
- Zhu, M.; Zhu, H.; Ding, X.; Liu, S.; Zou, Y. Analysis of the anti-fatigue activity of polysaccharides from Spirulina platensis: Role of central 5-hydroxytryptamine mechanisms. Food Funct. 2020, 11, 1826–1834. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Xiong, M.; Li, Y.; Qian, J.; Li, Y.; Han, X.; Tan, J.; Luo, Y.; Wang, Q.; Qin, C. Chitosan oligosaccharide combined with running benefited the immune status of rats. Int. Immunopharmacol. 2020, 88, 106915. [Google Scholar] [CrossRef]
- Spaulding, H.R.; Yan, Z. AMPK and the adaptation to exercise. Annu. Rev. Physiol. 2022, 84, 209–227. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Ji, H.; Song, W.; Peng, S.; Zhan, S.; Wei, L.; Chen, M.; Zhang, D.; Liu, S. The anti-fatigue effect of the Auxis thazard oligopeptide via modulation of the AMPK/PGC-1α pathway in mice. Food Funct. 2022, 13, 1641–1650. [Google Scholar] [CrossRef]
- Rodriguez-Lopez, C.; Santalla, A.; Valenzuela, P.L.; Real-Martínez, A.; Villarreal-Salazar, M.; Rodriguez-Gomez, I.; Pinós, T.; Ara, I.; Lucia, A. Muscle glycogen unavailability and fat oxidation rate during exercise: Insights from McArdle disease. J. Physiol. 2023, 601, 551–566. [Google Scholar] [CrossRef] [PubMed]
- Heden, T.D.; Chow, L.S.; Hughey, C.C.; Mashek, D.G. Regulation and role of glycophagy in skeletal muscle energy metabolism. Autophagy 2022, 18, 1078–1089. [Google Scholar] [CrossRef]
- Li, D.D.; Ma, J.M.; Li, M.J.; Gao, L.L.; Fan, Y.N.; Zhang, Y.N.; Tao, X.J.; Yang, J.J. Supplementation of Lycium barbarum polysaccharide combined with aerobic exercise ameliorates high-fat-induced nonalcoholic steatohepatitis via AMPK/PPARα/PGC-1α pathway. Nutrients 2022, 14, 3247. [Google Scholar] [CrossRef]
- Shen, K.; Pender, C.L.; Bar-Ziv, R.; Zhang, H.; Wickham, K.; Willey, E.; Durieux, J.; Ahmad, Q.; Dillin, A. Mitochondria as cellular and organismal signaling hubs. Annu. Rev. Cell Dev. Biol. 2022, 38, 179–218. [Google Scholar] [CrossRef]
- He, P.; Chen, L.; Qin, X.; Du, G.; Li, Z. Astragali Radix-Codonopsis Radix-Jujubae Fructus water extracts ameliorate exercise-induced fatigue in mice via modulating gut microbiota and its metabolites. J. Sci. Food Agric. 2022, 102, 5141–5152. [Google Scholar] [CrossRef]
- Flockhart, M.; Nilsson, L.C.; Tais, S.; Ekblom, B.; Apró, W.; Larsen, F.J. Excessive exercise training causes mitochondrial functional impairment and decreases glucose tolerance in healthy volunteers. Cell Metab. 2021, 33, 957–970.e6. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, C.; Hou, J.; Su, P.; Yang, Y.; Xia, B.; Zhao, X.; He, R.; Wang, L.; Cao, C.; et al. Red ginseng extract improves skeletal muscle energy metabolism and mitochondrial function in chronic fatigue mice. Front. Pharmacol. 2022, 13, 1077249. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cao, F.; Wu, Q.; Luo, Y.; Guo, T.; Han, S.; Huang, M.; Hu, Z.; Bai, J.; Luo, F.; et al. Dietary supplementation of octacosanol improves exercise-induced fatigue and its molecular mechanism. J. Agric. Food Chem. 2021, 69, 7603–7618. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Choi, Y.; Jeong, E.; Park, J.; Kim, J.; Tanaka, M.; Choi, J. Physiological significance of elevated levels of lactate by exercise training in the brain and body. J. Biosci. Bioeng. 2023, 135, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wu, Q.; Yu, W.; Ye, F.; Cao, Y.; Akan, O.D.; Wu, X.; Xie, T.; Lu, H.; Cao, F.; et al. Gastrodin ameliorates exercise-induced fatigue via modulating Nrf2 pathway and inhibiting inflammation in mice. Food Biosci. 2023, 51, 102262. [Google Scholar] [CrossRef]
- Powers, S.K.; Goldstein, E.; Schrager, M.; Ji, L.L. Exercise training and skeletal muscle antioxidant enzymes: An update. Antioxidants 2022, 12, 39. [Google Scholar] [CrossRef]
- Eggen, M.D.; Merboth, P.; Neukirchner, H.; Glomb, M.A. Lipid peroxidation has major impact on malondialdehyde-derived but only minor influence on glyoxal and methylglyoxal-derived protein modifications in carbohydrate-rich foods. J. Agric. Food Chem. 2022, 70, 10271–10283. [Google Scholar] [CrossRef]
- Qin, D.D.; Yang, F.Y.; Hu, Z.M.; Liu, J.L.; Wu, Q.; Luo, Y.; Yang, L.F.; Han, S.; Luo, F.J. Peptide T8 isolated from yak milk residue ameliorates H2O2-induced oxidative stress through Nrf2 signaling pathway in HUVEC cells. Food Biosci. 2021, 44, 101408. [Google Scholar] [CrossRef]
- Tuo, X.; Deng, Z.; Huang, G.; Gong, H.; Xie, H. Astragalus polysaccharide attenuates overexercise-induce myocardial injury via activating AMPK signaling pathway to suppress inflammation and oxidative stress. An. Acad. Bras. Cienc. 2021, 94, e20210314. [Google Scholar] [CrossRef]
- Zabriskie, H.A.; Blumkaitis, J.C.; Moon, J.M.; Currier, B.S.; Stefan, R.; Ratliff, K.; Harty, P.S.; Stecker, R.A.; Rudnicka, K.; Jäger, R.; et al. Yeast beta-glucan supplementation downregulates markers of systemic inflammation after heated treadmill exercise. Nutrients 2020, 12, 1144. [Google Scholar] [CrossRef]
- Han, C.; Shen, H.; Yang, Y.; Sheng, Y.; Wang, J.; Li, W.; Zhou, X.; Guo, L.; Zhai, L.; Guan, Q. Antrodia camphorata polysaccharide resists 6-OHDA-induced dopaminergic neuronal damage by inhibiting ROS-NLRP3 activation. Brain Behav. 2020, 10, e01824. [Google Scholar] [CrossRef]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Li, P.; Wang, D.; Wang, T.; Hao, D.; Qu, X. Alterations in intestinal microbiota diversity, composition, and function in patients with sarcopenia. Sci. Rep. 2021, 11, 4628. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.S.B.; Raes, J.; Bork, P. The human gut microbiome: From association to modulation. Cell 2018, 172, 1198–1215. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Luo, F.; Lin, Q. Dietary nutrition and gut microflora: A promising target for treating diseases. Trends Food Sci. Technol. 2018, 75, 72–80. [Google Scholar] [CrossRef]
- Przewłócka, K.; Folwarski, M.; Kaźmierczak-Siedlecka, K.; Skonieczna-Żydecka, K.; Kaczor, J.J. Gut-muscle axis exists and may affect skeletal muscle adaptation to training. Nutrients 2020, 12, 1451. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.; et al. MetaHIT Consortium. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Aran, L.; Bulli, G.; Curcio, F.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Sarcopenia: Assessment of disease burden and strategies to improve outcomes. Clin. Interv. Aging 2018, 13, 913–927. [Google Scholar] [CrossRef]
- Jäger, R.; Zaragoza, J.; Purpura, M.; Iametti, S.; Marengo, M.; Tinsley, G.M.; Anzalone, A.J.; Oliver, J.M.; Fiore, W.; Biffi, A.; et al. Probiotic administration increases amino acid absorption from plant protein: A placebo-controlled, randomized, double-blind, multicenter, crossover study. Probiotics Antimicrob. Proteins 2020, 12, 1330–1339. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Xu, F.; Liu, G.; Pang, B.; Liao, N.; Li, H.; Shi, J. Gut microbiota as a potential target for developing anti-fatigue foods. Crit. Rev. Food Sci. Nutr. 2023, 63, 3065–3080. [Google Scholar] [CrossRef]
- Sorboni, S.G.; Moghaddam, H.S.; Jafarzadeh-Esfehani, R.; Soleimanpour, S. A comprehensive review on the role of the gut microbiome in human neurological disorders. Clin. Microbiol. Rev. 2022, 35, e0033820. [Google Scholar] [CrossRef]
- Porter, N.T.; Martens, E.C. The critical roles of polysaccharides in gut microbial ecology and physiology. Annu. Rev. Microbiol. 2017, 71, 349–369. [Google Scholar] [CrossRef]
- Liu, H.; Liao, C.; Wu, L.; Tang, J.; Chen, J.; Lei, C.; Zheng, L.; Zhang, C.; Liu, Y.Y.; Xavier, J.; et al. Ecological dynamics of the gut microbiome in response to dietary fiber. ISME J. 2022, 16, 2040–2055. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, H.; Zhang, K.; Lu, Y.; Wu, Q.; Chen, J.; Li, Y.; Wu, Q.; Chen, Y. Extraction, purification, structural characterization, and gut microbiota relationship of polysaccharides: A review. Int. J. Biol. Macromol. 2022, 213, 967–986. [Google Scholar] [CrossRef]
- Tang, M.; Cheng, L.; Liu, Y.; Wu, Z.; Zhang, X.; Luo, S. Plant Polysaccharides modulate immune function via the gut microbiome and may have potential in COVID-19 therapy. Molecules 2022, 27, 2773. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Jiang, W.; Yang, F.; Cheng, Y.; Guo, Y.; Yao, W.; Zhao, Y.; Qian, H. The combination of microbiome and metabolome to analyze the cross-cooperation mechanism of Echinacea purpurea polysaccharide with the gut microbiota in vitro and in vivo. Food Funct. 2022, 13, 10069–10082. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, M.; Li, Z.; Wei, J.; Liu, F.; Zheng, W.; Zhu, X.; Chai, X.; Zhao, S. Eucommiae cortex polysaccharides mitigate obesogenic diet-induced cognitive and social dysfunction via modulation of gut microbiota and tryptophan metabolism. Theranostics 2022, 12, 3637–3655. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Li, Q.M.; Zha, X.Q.; Luo, J.P. Dendrobium fimbriatum Hook polysaccharide ameliorates dextran-sodium-sulfate-induced colitis in mice via improving intestinal barrier function, modulating intestinal microbiota, and reducing oxidative stress and inflammatory responses. Food Funct. 2022, 13, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, L.; Zhu, M.; Yang, B.; Yang, Y.; Jia, X.; Feng, L. Moutan Cortex polysaccharide ameliorates diabetic kidney disease via modulating gut microbiota dynamically in rats. Int. J. Biol. Macromol. 2022, 206, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Wang, T.; Li, Y.; Xue, H.; Zou, J.; Cai, J.; Shi, R.; Wu, J.; Ma, Y. Gardenia jasminoides Ellis polysaccharide ameliorates cholestatic liver injury by alleviating gut microbiota dysbiosis and inhibiting the TLR4/NF-κB signaling pathway. Int. J. Biol. Macromol. 2022, 205, 23–36. [Google Scholar] [CrossRef]
- Ticinesi, A.; Lauretani, F.; Tana, C.; Nouvenne, A.; Ridolo, E.; Meschi, T. Exercise and immune system as modulators of intestinal microbiome: Implications for the gut-muscle axis hypothesis. Exerc. Immunol. Rev. 2019, 25, 84–95. [Google Scholar]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Guan, T.; Li, S.; Guan, Q.; Shi, J.S.; Lu, Z.M.; Xu, Z.H.; Geng, Y. Spore powder of Paecilomyces hepialid shapes gut microbiota to relieve exercise-induced fatigue in mice. Nutrients 2022, 14, 2973. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, H.; Hossainpour, H. Application and development of fecal microbiota transplantation in the treatment of gastrointestinal and metabolic diseases: A review. Saudi J. Gastroenterol. 2023, 29, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Moretti, C.; Benjamin, N.; Weitzberg, E. Symbiotic bacteria enhance exercise performance. Br. J. Sports Med. 2021, 55, 243. [Google Scholar] [CrossRef]
- Ryu, J.Y.; Choi, H.M.; Yang, H.I.; Kim, K.S. Dysregulated autophagy mediates sarcopenic obesity and its complications via AMPK and PGC1α signaling pathways: Potential involvement of gut dysbiosis as a pathological link. Int. J. Mol. Sci. 2020, 21, 6887. [Google Scholar] [CrossRef]
- Zhong, L.; Ma, N.; Zheng, H.; Ma, G.; Zhao, L.; Hu, Q. Tuber indicum polysaccharide relieves fatigue by regulating gut microbiota in mice. J. Funct. Foods 2019, 63, 103580. [Google Scholar] [CrossRef]
- Cai, J.; Sun, L.; Gonzalez, F.J. Gut microbiota-derived bile acids in intestinal immunity, inflammation, and tumorigenesis. Cell Host Microbe 2022, 30, 289–300. [Google Scholar] [CrossRef]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The microbiota-gut-brain axis: From motility to mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Zheng, Z.; Xie, G.; Liu, H.; Tan, G.; Li, L.; Liu, W. Fermented ginseng leaf enriched with rare ginsenosides relieves exercise-induced fatigue via regulating metabolites of muscular interstitial fluid, satellite cells-mediated muscle repair and gut microbiota. J. Funct. Foods 2021, 83, 104509. [Google Scholar] [CrossRef]
- Li, G.; Jin, B.; Fan, Z. Mechanisms involved in gut microbiota regulation of skeletal muscle. Oxid. Med. Cell. Longev. 2022, 2022, 2151191. [Google Scholar] [CrossRef]
- Chew, W.; Lim, Y.P.; Lim, W.S.; Chambers, E.S.; Frost, G.; Wong, S.H.; Ali, Y. Gut-muscle crosstalk. A perspective on influence of microbes on muscle function. Front. Med. 2023, 9, 1065365. [Google Scholar] [CrossRef]
- Nay, K.; Jollet, M.; Goustard, B.; Baati, N.; Vernus, B.; Pontones, M.; Lefeuvre-Orfila, L.; Bendavid, C.; Rué, O.; Mariadassou, M.; et al. Gut bacteria are critical for optimal muscle function: A potential link with glucose homeostasis. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E158–E171. [Google Scholar] [CrossRef]
- Lahiri, S.; Kim, H.; Garcia-Perez, I.; Reza, M.M.; Martin, K.A.; Kundu, P.; Cox, L.M.; Selkrig, J.; Posma, J.M.; Zhang, H.; et al. The gut microbiota influences skeletal muscle mass and function in mice. Sci. Transl. Med. 2019, 11, eaan5662. [Google Scholar] [CrossRef] [PubMed]
- Thye, A.Y.; Law, J.W.; Tan, L.T.; Thurairajasingam, S.; Chan, K.G.; Letchumanan, V.; Lee, L.H. Exploring the gut microbiome in myasthenia gravis. Nutrients 2022, 14, 1647. [Google Scholar] [CrossRef] [PubMed]
- Ticinesi, A.; Lauretani, F.; Milani, C.; Nouvenne, A.; Tana, C.; Del Rio, D.; Maggio, M.; Ventura, M.; Meschi, T. Aging gut microbiota at the cross-road between nutrition, physical frailty, and sarcopenia: Is there a gut-muscle axis? Nutrients 2017, 9, 1303. [Google Scholar] [CrossRef]
- Chen, L.H.; Chang, S.S.; Chang, H.Y.; Wu, C.H.; Pan, C.H.; Chang, C.C.; Chan, C.H.; Huang, H.Y. Probiotic supplementation attenuates age-related sarcopenia via the gut-muscle axis in SAMP8 mice. J. Cachexia Sarcopenia Muscle 2022, 13, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Liu, Y.; Yang, G.; Meng, B.; Yi, Z.; Yang, G.; Chen, M.; Hou, P.; Wang, H.; Xu, X. Moderate-intensity physical exercise affects the exercise performance and gut microbiota of mice. Front. Cell. Infect. Microbiol. 2021, 11, 712381. [Google Scholar] [CrossRef]
- Prokopidis, K.; Chambers, E.; Ni Lochlainn, M.; Witard, O.C. Mechanisms linking the gut-muscle axis with muscle protein metabolism and anabolic resistance: Implications for older adults at risk of sarcopenia. Front. Physiol. 2021, 12, 770455. [Google Scholar] [CrossRef]
- Yin, Y.; Guo, Q.; Zhou, X.; Duan, Y.; Yang, Y.; Gong, S.; Han, M.; Liu, Y.; Yang, Z.; Chen, Q.; et al. Role of brain-gut-muscle axis in human health and energy homeostasis. Front. Nutr. 2022, 9, 947033. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.C.; Chen, M.J.; Huang, H.W.; Wu, W.K.; Lee, Y.W.; Kuo, H.C.; Huang, C.C. Probiotic Lactiplantibacillus plantarum tana isolated from an international weightlifter enhances exercise performance and promotes antifatigue effects in mice. Nutrients 2022, 14, 3308. [Google Scholar] [CrossRef] [PubMed]
- Vigh-Larsen, J.F.; Ørtenblad, N.; Spriet, L.L.; Overgaard, K.; Mohr, M. Muscle glycogen metabolism and high-intensity exercise performance: A narrative review. Sports Med. 2021, 51, 1855–1874. [Google Scholar] [CrossRef]
- Lee, M.C.; Hsu, Y.J.; Ho, H.H.; Kuo, Y.W.; Lin, W.Y.; Tsai, S.Y.; Chen, W.L.; Lin, C.L.; Huang, C.C. Effectiveness of human-origin Lactobacillus plantarum PL-02 in improving muscle mass, exercise performance and anti-fatigue. Sci. Rep. 2021, 11, 19469. [Google Scholar] [CrossRef] [PubMed]
- Yeh, W.L.; Hsu, Y.J.; Ho, C.S.; Ho, H.H.; Kuo, Y.W.; Tsai, S.Y.; Huang, C.C.; Lee, M.C. Lactobacillus plantarum PL-02 supplementation combined with resistance training improved muscle mass, force, and exercise performance in mice. Front. Nutr. 2022, 9, 896503. [Google Scholar] [CrossRef]
- de Sire, R.; Rizzatti, G.; Ingravalle, F.; Pizzoferrato, M.; Petito, V.; Lopetuso, L.; Graziani, C.; de Sire, A.; Mentella, M.C.; Mele, M.C.; et al. Skeletal muscle-gut axis: Emerging mechanisms of sarcopenia for intestinal and extra intestinal diseases. Minerva Gastroenterol. Dietol. 2018, 64, 351–362. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources | Polysaccharides | Total Sugar Content | Monosaccharide Composition | Preparation | Model/ Number | Method/Dose/ Time | Characteristics | Ref. |
|---|---|---|---|---|---|---|---|---|
| corn silk | PCS | 58.6% | Rha, Ara, Xyl, Man, Glu, and Gal; ratio is 0.17:0.30:0.26:0.35: 1.00:0.57 | Water-extracting, alcohol precipitating, and ultrasonic-assisted | Kunming mice, (n = 20) | oral gavage; 50, 100, 200, 400 mg/kg; 4 weeks | body weight, food consumption, weights of main organ →; swimming time to exhaustion ↑; LDH, HG, GLU, TC, Cr ↑; BUN, LA, ALT, ALP ↓; AST, TG → | [71] |
| Herba epimedii | HEP2-a | 60.1% | Man, Rha, Glu, Gal, Ara, and galacturonic acid in percentages of 4.41%, 5.43%, 31.26, 27.07%, 23.43%, 8.40% | soaked in 95% alcohol, extracted with distilled water at 80 °C | CFS rat, (n = 10) | oral gavage; 50, 100, 200 mg/kg; 4 weeks | body weight, number of standing ↑; time needed to locate the platform; motionless time in the tail-suspension test ↓; noradrenaline, creatine ↑ | [72] |
| Dendrobium officinale | DOP | 30% | / | / | inbred strain male BALB/c mice, (n = 8) | oral; 50 mg/kg; 30 days | endurance, body weight, food intake ↑; LDH, BUN, MDA, CK, TG, and LD in serum ↓; serum SOD/GSH-Px and gastrocnemius glycogen ↑; cell variability of T and B lymphocytes ↑ | [41] |
| Lepidium meyenii Walp. (Maca) | MP | 99.2% | D-GalA: D-Glc: L-Ara: D-Man: D-Gal: L-Rha = 35.07:29.98:16.98:13.01:4.21:0.75 (mol, %) | extracted with hot water at 90 °C | ICR mice, (n = 20) | Intragastrically; 25, 50, 100 mg/kg bw/d; 30 days | viscera indices, body weight gain →; swimming durations, average swimming speeds ↑; GSH-Px, CK ↑; LDH, BUN, MDA, LA ↓ | [73] |
| Lepidium meyenii Walp. (maca) | MPS-1, MPS-2 | 93.2%; 91.5% | MPS-1: xylose, arabinose, galactose, glucose, with the mole ratio 1:1.7:3.3:30.5; MPS-2: arabinose, galactose, glucose, with the mole ratio 1:1.3:36.8 | extracted with distilled water at 80 °C | male Kunming mice, (n = 10) | oral gavage; 20, 100 mg/kg·d; 30 days | exhausting swimming time ↑; LG ↑; BLA, BUN, LDH ↓ | [74] |
| Epidium meyenii Walp. (Maca) | ME | 19.72% | / | extracted in boiling water | ICR mice, (n = 10); H2O2-induced C2C12 cell 6 h | oral; 10 mL per kg bw; 4 weeks; 0.10 mg mL−1 | grip-strength, exercise time in the rotarod test ↑; BLA, BUN, LDH, ROS ↓; skeletal muscle damage ↓; NAD+, NADH ↑; no cytotoxicity, cytoprotective and 68.8% anti-oxidation effects, ROS ↓; mitochondrial quality ↑ | [16] |
| Phragmites rhizoma | PEP | 85.6% | / | hot water decoction | male ICR mice, (n = 6) | oral; 1 g/kg; 10 days | body weight →; time to reach fatigue, swimming endurance capacity ↑; glucose, TG, T. chol., GSH, SOD, CAT, GPx ↑; LDH, CK, cortisol, MPO, TBARS ↓ | [75] |
| Codonopsis pilosula | POL | 15.52% | / | extracted with 95% ethanol and decocted with boiling water | male ICR mice, (n = 8) | oral gavage; 0.25, 0.5, 1.0 g/kg; 21 days | swimming time, hypoxia tolerance ↑; MG, LG, GSH ↑; BUN, LDH, MDA ↓ | [76] |
| Polygala tenuifolia Willd. | / | / | / | extracted with 95% ethanol reflux and 67 °C water | male mice, (n = 10) | gavage, 0.1, 0.2, 0.4 mg/g, 30 days | weight-loaded swimming time ↑; LG, MG, LDH ↑; BLA, BUN ↓; scavenging rate of hydroxyl and DPPH free radical ↑ | [77] |
| Chinese yam (Dioscorea opposita Thunb.) | CYPs | 73.20% | rhamnose, glucuronic acid, glucose, galactose, and arabinose with a molar ratio of 0.01:0.06:1.00:0.17:0.01 | water-extracting, alcohol-precipitating | Swiss mice, (n = 10) | oral; 100 mg/kg; 14 days | exhausting swimming time, movement distance, body weight ↑, static time ↓; ATP, SOD ↑; IL-lβ, MDA, BUN, LDH ↓; tumor volume and tumor weight ↓ | [18] |
| Semen cassiae | SCP | 5.42%. | / | extracted using aqueous solvent (water) | BALB/c male mice | oral; 100 mg/kg; 4 weeks | swimming duration, body weight, food intake ↑; BUN, CK, TG, LA, LDH, MDA ↓; SOD, GPX, liver and muscle glycogen ↑; lymphocyte proliferation ↑ | [14] |
| Ginseng | WSGP-S3 | 8.01% | Rha, GalA, Glc, Gal, Ara GlcA was 35.48, 6.98, 5.98, 32.02, 16.95, 2.67 mol% | extracted under reflux extraction with 95% ethanol; washed with distilled water, and extracted with hot water | male Kunming mice, (n = 10) | oral; 25, 50, 75 mg/kg; 30 days | body weight →; thymus and spleen index ↑; exhaustive swimming time↑; LG, MG, SOD, CAT, GSH-Px ↑; BUN, LA, MDA ↓; T or B lymphocyte proliferation ↑ | [17] |
| Panax ginseng C. A. Meyer | APS-1 | 96% | Man, Rha, GlcA, GalA, Glc, Gal and Ara with an area ratio of 3: 4:2:17:7:40:27 | extracted with boiling water and separated by DEAE-52 cellulose ion exchange column and gel column chromatography | male C57BL/6J mice, (n = 8) | Intragastric; 50, 100, 150 mg/kg; 15 days | fatigue tolerance time ↑; BLA, LDH, BUN, MDA ↓; SOD, CAT, CK ↑; LKB1, p-AMPK, PGC-1α and Glut4 ↑ | [78] |
| Lycium barbarum | LBP1-SeNPs | 80.1% | arabinose (24.0%), ylose (25.0%), glucose (20.2%), galactose (23.1%) | immersed in 95% ethanol and extracted with ultra-pure water at 90 °C | male ICR mice, (n = 20) | Intragastric; 0.5, 2, 4 Se/kg/d; 30 days | body weigh →; exhaustive swimming time ↑; liver and muscle glycogen, SOD ↑; BUN, BLA, MDA ↓ | [79] |
| Lycium barbarum | LBP | 4.73%, 97.22% | Rha, Ara, Gal, Glu, Xyl and Man with a molar ratio of 1.00:11.35:6.10:0.56:1.08:0.71 | extracted with water, and DEAE-52 cellulose was used for purification | 4-week-old male SD rats, (n = 8) | Gavage; 120, 360 mg/kg/d; 28 days | exhaustive swimming time ↑; LA and CK, serum glucose, ATP and glycogen ↑ | [68] |
| Inonotus obliquus | PIO-1 | 73.2% | mannose, glucose, galactose, xylose, and arabinose with the molar ratio of 1.0:1.9:3.5:18.5:5.7 | hot water extraction | male Kunming mice, (n = 10) | oral; 50 mg/kg/day; 30 days | climbing duration, swimming time ↑; immobility time ↓; BLA, BUN, LDH, 5-HT ↓; GRAF1 ↑ | [19] |
| Sarcodon imbricatus | SI | 35.22% | / | / | Kunming male mice, (n = 10) | oral gavage; 0.25, 0.5, 1 g/kg; 20 days | exercise tolerance↑, immobility in the tail suspension test ↓; glycogen, ATP, SOD, GSH-Px ↑, LD, BUN, ROS, MDA ↓; Nrf2, SOD1, SOD2, HO-1, and CAT ↑ | [44] |
| Ganoderma lucidum | GLPs | / | Fuc, Ara, Gal, Glc, Xyl, Man | extracted by hot water | male Balb/C rats, (n = 10) | Gavage; 100, 150, 200 mg/kg; 4 weeks | body weight →; exercise time ↑; BUN, CK, MDA, BLA ↓; Gly, TG, GPx, SOD, LDH ↑; fat transformation ↑ | [15] |
| Pholiota nameko | PNP | 82% | / | hot water extracting-alcohol precipitating | adult Kun Ming mice, (n = 6) | gavage; 150, 300, 450 mg/kg; 15 days | forced swim time ↑; LDH, ATPase, SOD ↑; MDA ↓ | [80] |
| Cordyceps militaris | CMPB | 36.81% | Man (44.06%), Gal (28.41%), Glc (7.42%), Rha (4.98%), Xyl (4.93%), Ara (4.42%) | extracted, isolated and purified via DEAE-cellulose 52 and Sepharose CL-6B columns | male ICR mice, (n = 15) | Intragastric; 400, 800 mg/kg; 5 weeks | DPPH, superoxide anion, hydroxyl radicals ↑; stick rotation time, learning and memory abilities ↑; LD, LDH, BUN, MDA, ROS ↓; SOD, GSH-Px, eNOS, Glu/GABA, BDNF, PI3K, Nrf2, HO-1 ↑ | [69] |
| Spirulina platensis | PSP | 70.13% | / | / | SD male rats, (n = 10) | Intragastric; 50, 100, 200 mg kg−1; 7 days | exhaustion time ↑; Hb, 5-HT1B ↑; LA, BUN, CK, 5-HT, TPH2 ↓ | [81] |
| Chitosan oligosaccharide | COS | / | / | / | SD rats, (n = 6) | Intragastric; 200 mg/kg; 4 weeks | spleen weight ↑; LAC, the ratio of T cell/CD8+ T cell ↑; ALB, BUN, blood creatinine, lymphocytes, LDH, CREA, TNF, IL-2 ↓; TC, TG, HDL, Na+, K+, and Ca2+, platelet, IL-10, IL-4 → | [82] |
| Sources | Polysaccharides | Total Sugar Content | Monosaccharide Composition | Preparation | Model/ Number | Method/Dose/ Time | Changed Abundance of Gut Microbiota and SCFAs | Characteristics | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Ginseng | WEG | 90.15% | Glc, Gal, Ara, GlcA, GalA, Man, Rha, and Fuc, with the molar ratios of 1588:96:80:94:79:21:13:1 | prepared by hot water extraction | SD rats, (n = 8) | intraperitoneal injection; 1.42 g kg−1; 14 days | TUDCA, TCDCA, UDCA 3-sulfate, CDCA 3-sulfate ↓; butyrate ↑; tryptophan, LPC ↑; Bacteroidetes, Lactobacillus, Bacteroides ↑, Firmicutes ↓; Bifidobacterium, Coprococcus ↑; Anaerotruncus, Streptococcus, Blautia, Clostridium ↓ | body weight ↑; LA, CP, LDH, MDA, BUN, TG ↓; GPx, Glc, SOD ↑; IL-1β ↓, TGR5 ↑ | [21] |
| Lepidium meyenii Walp. (Maca) | MCP | 34.78 mg/mL | / | extracted in boiling water (w/v, 1:8) | ICR mice, (n = 10) | oral, 1, 2, 4 g/kg w; 30 days | Lactobacillus, Akkermansia, Lachnospiraceae, Bacteroides, Blautia, Roseburia, Parabacteroides, Clostridia_UCG-014, Marvinbryantia ↑; Candidatus_Planktophila, Candidatus_Arthromitus ↓ | grip-strength, rotarod ↑; blood sugar, NAD(H), MG, ATP ↑; BLA, BUN, LDH, ROS ↓ | [29] |
| Fermented ginseng leaf | FGL | 19.2 mg/g | / | fermented by S. cerevisiae | male SD rats, (n = 12) | intragastric administration; 10, 20, 50 mg/kg/d; 4 weeks | acetic acid, propionic acid, n-buryric acid, i-butyric acid, total SCFAs in feces ↑; Bacteroidetes, Bacteroidetes/Firmicute ratio, Allobaculum, Akkermansia, Bifidobacterium, Eubacterium ↑; Verrucomicrobiales, Coprobacillus, Erysipelotrichaceae ↓ | endurance time ↑; hypoxanthine, isoprostane ↓; TNF-α, IL-6, IL-10 ↓; SOD, GSH-Px ↑; MDA ↓; MyHC-I, MyHC-II ↓; Pax7, MyoD1 ↑ | [129] |
| Tuber indicum | TIP | 90.25% | Rha, Man, Glu, and Gal, with a molar percentage of 6.73%, 13.36%, 76.78% and 3.13% | extracted with ultrapure water, and precipitated with of 100% ethanol | male C57BL/6 mice, (n = 8) | Gavage; 0.1, 0.3, 0.9 g/kg body weight; 40 days | acetic acid, propionic acid, n-butyric acid, i-butyric acid, total SCFAs ↑; Firmicutes, Proteobacteria ↓; Bacteroidetes, Actinobacteria ↑; Prevotellaceae, Ruminococcaceae, Helicobacteraceae ↓; Porphyromonadaceae, Bacteroidaceae, Rikenellaceae ↑; Odoribacter, Bacteroides, Roseburia↑; Deferribacteres, Lachnospiraceae → | exhaustive swimming time, weight, feed efficiency ↑; LG, MG, BG ↑, BLA, BUN, CK ↓; LDH, Na+K+-ATPase, Ca2+Mg2+-ATPase ↑; p-p38/p38, p-AMPK/AMPK ↓; occluding ↑, ZO-1 → | [126] |
| Astragali Radix, Codonopsis Radix, Jujubae Fructus | ACJ | 43.46 mg/g | / | extracted with water, concentrated under vacuum in a rotary evaporator | male ICR mice, (n = 11) | oral gavage; 4.69, 18.75 g kg−1 d−1; 6 weeks | Actinobacteria, Proteobacteria, TM7, Cyanobacteria ↑; Oscillospira, Hymenobacter, Deinococcus, Bilophila, Streptococcus ↓; Adlercreutzia, Fructobacillus, Desulfovibrio ↑ | Body weight →; exhaustive swimming time ↑; BUN, LA ↓; LG ↑ | [89] |
| Dendrobium officinale | EPDO | 92.6% | Man and Glc ratio is 2.0–4.0 | immersed with 80% ethanol, extracted with deionized water | male ICR mice, (n = 6) | oral gavage; 100, 150, 200 mg/kg; 4 weeks | Bacteroidetes, Actinobacteria ↑; Firmicutes, Proteobacteria ↓; Lactobacillus, Bifidobacterium, Roseburia ↑; Acidaminoccus ↓ | body weight →; swimming time ↑; LG, MG →; SOD ↑; BLA, BUN, MDA ↓ | [30] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Chu, Z.; Luo, Y.; Yang, F.; Cao, F.; Luo, F.; Lin, Q. Dietary Polysaccharides Exert Anti-Fatigue Functions via the Gut-Muscle Axis: Advances and Prospectives. Foods 2023, 12, 3083. https://doi.org/10.3390/foods12163083
Zhou Y, Chu Z, Luo Y, Yang F, Cao F, Luo F, Lin Q. Dietary Polysaccharides Exert Anti-Fatigue Functions via the Gut-Muscle Axis: Advances and Prospectives. Foods. 2023; 12(16):3083. https://doi.org/10.3390/foods12163083
Chicago/Turabian StyleZhou, Yaping, Zhongxing Chu, Yi Luo, Feiyan Yang, Fuliang Cao, Feijun Luo, and Qinlu Lin. 2023. "Dietary Polysaccharides Exert Anti-Fatigue Functions via the Gut-Muscle Axis: Advances and Prospectives" Foods 12, no. 16: 3083. https://doi.org/10.3390/foods12163083
APA StyleZhou, Y., Chu, Z., Luo, Y., Yang, F., Cao, F., Luo, F., & Lin, Q. (2023). Dietary Polysaccharides Exert Anti-Fatigue Functions via the Gut-Muscle Axis: Advances and Prospectives. Foods, 12(16), 3083. https://doi.org/10.3390/foods12163083