

Effects of a Novel Starter Culture on Quality Improvement and Putrescine, Cadaverine, and Histamine Inhibition of Fermented Shrimp Paste


Abstract
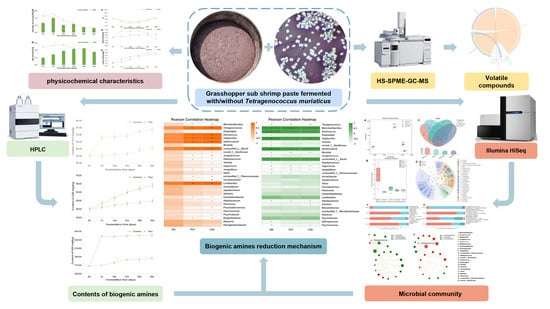
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Starter Culture
2.3. Preparation of Grasshopper Sub Shrimp Paste Samples
2.4. Physicochemical and Microbial Analysis
2.5. Biogenic Amine Measurement by HPLC
2.6. Microbial Community Analysis by Illumina High-Throughput Sequencing
2.7. Identification of Volatile Compounds by HS-SPME-GC-MS
2.8. Statistical Analysis
2.9. Nucleotide Sequence Accession Numbers
3. Results
3.1. Analysis of Bacterial Community in Shrimp Paste Samples
3.2. Effect of Starter Culture on the Interspecies Correlation
3.3. Contribution of the Starter Culture to BA Reduction
3.4. Effects of Starter Culture on Physicochemical Properties and Microbial Counts of Shrimp Paste
3.5. Impacts of Starter Culture on Volatile Compounds of Shrimp Paste
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, W.; Mi, S.; Liu, X.; Sang, Y.; Sun, J. Variations in the physicochemical properties and bacterial community composition during fermentation of low-salt shrimp paste. Food Res. Int. 2022, 154, 111034. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lu, K.; Dong, X.; Xie, W. Virgibacillus sp. SK37 and Staphylococcus nepalensis JS11 as potential starters to improve taste of shrimp paste. LWT Food Sci. Technol. 2022, 154, 112657. [Google Scholar] [CrossRef]
- Sang, X.; Li, K.; Zhu, Y.; Ma, X.; Hao, H.; Bi, J.; Zhang, G.; Hou, H. The Impact of Microbial Diversity on Biogenic Amines Formation in Grasshopper Sub Shrimp Paste during the Fermentation. Front. Microbiol. 2020, 11, 782. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lu, K.; Zi, J.; Yang, X.; Zheng, Z.; Xie, W. Halophilic bacteria as starter cultures: A new strategy to accelerate fermentation and enhance flavor of shrimp paste. Food Chem. 2022, 393, 133393. [Google Scholar] [CrossRef]
- Park, Y.K.; Lee, J.H.; Mah, J.H. Occurrence and reduction of biogenic amines in traditional Asian fermented soybean foods: A review. Food Chem. 2019, 278, 1–9. [Google Scholar] [CrossRef]
- Anal, A.K.; Perpetuini, G.; Petchkongkaew, A.; Tan, R.; Avallone, S.; Tofalo, R.; Nguyen, H.V.; Chu-Ky, S.; Ho, P.H.; Phan, T.T.; et al. Food safety risks in traditional fermented food from South-East Asia. Food Control 2020, 109, 106922. [Google Scholar] [CrossRef]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic Amine Production by Lactic Acid Bacteria: A Review. Foods 2019, 8, 17. [Google Scholar] [CrossRef]
- Jaguey-Hernandez, Y.; Aguilar-Arteaga, K.; Ojeda-Ramirez, D.; Anorve-Morga, J.; Gonzalez-Olivares, L.G.; Castaneda-Ovando, A. Biogenic amines levels in food processing: Efforts for their control in foodstuffs. Food Res. Int. 2021, 144, 110341. [Google Scholar] [CrossRef]
- Doeun, D.; Davaatseren, M.; Chung, M.S. Biogenic amines in foods. Food Sci. Biotechnol. 2017, 26, 1463–1474. [Google Scholar] [CrossRef]
- Jin, Y.H.; Lee, J.H.; Park, Y.K.; Lee, J.H.; Mah, J.H. The Occurrence of Biogenic Amines and Determination of Biogenic Amine-Producing Lactic Acid Bacteria in Kkakdugi and Chonggak Kimchi. Foods 2019, 8, 73. [Google Scholar] [CrossRef]
- Shen, Y.; Wu, Y.; Wang, Y.; Li, L.; Li, C.; Zhao, Y.; Yang, S. Contribution of autochthonous microbiota succession to flavor formation during Chinese fermented mandarin fish (Siniperca chuatsi). Food Chem. 2021, 348, 129107. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Li, L.; Yang, X.; Chen, S.; Wu, Y.; Zhao, Y.; Wang, J.; Wei, Y.; Yang, D. Application of UHPLC-Q/TOF-MS-based metabolomics in the evaluation of metabolites and taste quality of Chinese fish sauce (Yu-lu) during fermentation. Food Chem. 2019, 296, 132–141. [Google Scholar] [CrossRef]
- Sang, X.; Ma, X.; Hao, H.; Bi, J.; Zhang, G.; Hou, H. Evaluation of biogenic amines and microbial composition in the Chinese traditional fermented food grasshopper sub shrimp paste. LWT Food Sci. Technol. 2020, 134, 109979. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Y.; Li, X.; Bi, J.; Zhang, G.; Hao, H.; Hou, H. Impacts of salt-tolerant Staphylococcus nepalensis 5-5 on bacterial composition and biogenic amines accumulation in fish sauce fermentation. Int. J. Food Microbiol. 2022, 361, 109464. [Google Scholar] [CrossRef]
- Saelao, S.; Maneerat, S.; Thongruck, K.; Watthanasakphuban, N.; Wiriyagulopas, S.; Chobert, J.M.; Haertle, T. Reduction of tyramine accumulation in Thai fermented shrimp (kung-som) by nisin Z-producing Lactococcus lactis KTH0-1S as starter culture. Food Control 2018, 90, 249–258. [Google Scholar] [CrossRef]
- Hua, Q.; Gao, P.; Xu, Y.; Xia, W.; Sun, Y.; Jiang, Q. Effect of commercial starter cultures on the quality characteristics of fermented fish-chili paste. LWT Food Sci. Technol. 2020, 122, 109016. [Google Scholar] [CrossRef]
- Zhang, Y.; Qin, Y.; Wang, Y.; Huang, Y.; Li, P.; Li, P. Lactobacillus plantarum LPL-1, a bacteriocin producing strain, changed the bacterial community composition and improved the safety of low-salt fermented sausages. LWT Food Sci. Technol. 2020, 128, 109385. [Google Scholar] [CrossRef]
- Hua, Q.; Sun, Y.; Xu, Y.; Gao, P.; Xia, W. Bacterial community succession and biogenic amine changes during fermentation of fish-chili paste inoculated with different commercial starter cultures. Int. J. Food Sci. Technol. 2021, 56, 6752–6764. [Google Scholar] [CrossRef]
- Li, C.; Li, W.; Li, L.; Chen, S.; Wu, Y.; Qi, B. Microbial community changes induced by a newly isolated salt-tolerant Tetragenococcus muriaticus improve the volatile flavor formation in low-salt fish sauce. Food Res. Int. 2022, 156, 111153. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, S.H.; Chun, B.H.; Jeong, S.E.; Jeon, C.O. Tetragenococcus halophilus MJ4 as a starter culture for repressing biogenic amine (cadaverine) formation during saeu-jeot (salted shrimp) fermentation. Food Microbiol. 2019, 82, 465–473. [Google Scholar] [CrossRef]
- Lee, J.H.; Heo, S.; Jeong, K.; Lee, B.; Jeong, D.W. Genomic insights into the non-histamine production and proteolytic and lipolytic activities of Tetragenococcus halophilus KUD23. FEMS Microbiol. Lett. 2017, 365, fnx252. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.W.; Heo, S.; Lee, J.H. Safety assessment of Tetragenococcus halophilus isolates from doenjang, a Korean high-salt-fermented soybean paste. Food Microbiol. 2017, 62, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Sang, X.; Ma, X.; Zhang, Y.; Hao, H.; Bi, J.; Zhang, G.; Hou, H. Assessment of the Distribution and Safety of Tetragenococcus muriaticus for Potential Application in the Preparation of Chinese Grasshopper Sub Shrimp Paste. Front. Microbiol. 2021, 12, 628838. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Lu, H.; He, Z.; Sang, Y.; Sun, J. Quality characteristics and bacterial community of a Chinese salt-fermented shrimp paste. LWT Food Sci. Technol. 2021, 136, 110358. [Google Scholar] [CrossRef]
- Yao, Y.; Zhou, X.; Hadiatullah, H.; Zhang, J.; Zhao, G. Determination of microbial diversities and aroma characteristics of Beitang shrimp paste. Food Chem. 2021, 344, 128695. [Google Scholar] [CrossRef]
- Ma, X.; Bi, J.; Li, X.; Zhang, G.; Hao, H.; Hou, H. Contribution of Microorganisms to Biogenic Amine Accumulation during Fish Sauce Fermentation and Screening of Novel Starters. Foods 2021, 10, 2572. [Google Scholar] [CrossRef]
- Ryu, J.-A.; Kim, E.; Yang, S.-M.; Lee, S.; Yoon, S.-R.; Jang, K.-S.; Kim, H.-Y. High-throughput sequencing of the microbial community associated with the physicochemical properties of meju (dried fermented soybean) and doenjang (traditional Korean fermented soybean paste). LWT Food Sci. Technol. 2021, 146, 111473. [Google Scholar] [CrossRef]
- Belleggia, L.; Aquilanti, L.; Ferrocino, I.; Milanovic, V.; Garofalo, C.; Clementi, F.; Cocolin, L.; Mozzon, M.; Foligni, R.; Haouet, M.N.; et al. Discovering microbiota and volatile compounds of surströmming, the traditional Swedish sour herring. Food Microbiol. 2020, 91, 103503. [Google Scholar] [CrossRef]
- Zhao, C.C.; Eun, J.B. Shotgun metagenomics approach reveals the bacterial community and metabolic pathways in commercial hongeo product, a traditional Korean fermented skate product. Food Res. Int. 2020, 131, 109030. [Google Scholar] [CrossRef]
- Suzuki, T.; Matsutani, M.; Matsuyama, M.; Unno, R.; Matsushita, H.; Sugiyama, M.; Yamasato, K.; Koizumi, Y.; Ishikawa, M. Growth and metabolic properties of halophilic and alkaliphilic lactic acid bacterial strains of Marinilactibacillus psychrotolerans isolated from surface-ripened soft cheese. Int. Dairy J. 2021, 112, 104840. [Google Scholar] [CrossRef]
- Ruiz-Capillas, C.; Herrero, A.M. Impact of Biogenic Amines on Food Quality and Safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef]
- Ma, Q.; Lu, X.; Wang, W.; Hubbe, M.A.; Liu, Y.; Mu, J.; Wang, J.; Sun, J.; Rojas, O.J. Recent developments in colorimetric and optical indicators stimulated by volatile base nitrogen to monitor seafood freshness. Food Packag. Shelf Life 2021, 28, 100634. [Google Scholar] [CrossRef]
- Yu, J.; Lu, K.; Sun, J.; Xie, W.; Song, L. The Flavor and Antioxidant Activity Change Pattern of Shrimp Head Paste during Fermentation. J. Ocean Univ. China 2022, 21, 195–203. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, L.; Liu, H.; Liu, H.; Zhou, Y.; Li, M.; Gao, R. Analysis of the changes of volatile flavor compounds in a traditional Chinese shrimp paste during fermentation based on electronic nose, SPME-GC-MS and HS-GC-IMS. Food Sci. Hum. Wellness 2023, 12, 173–182. [Google Scholar] [CrossRef]
- Yu, J.; Lu, K.; Zi, J.; Yang, X.; Xie, W. Characterization of aroma profiles and aroma-active compounds in high-salt and low-salt shrimp paste by molecular sensory science. Food Biosci. 2022, 45, 101470. [Google Scholar] [CrossRef]
- Odeyemi, O.A.; Burke, C.M.; Bolch, C.C.J.; Stanley, R. Seafood spoilage microbiota and associated volatile organic compounds at different storage temperatures and packaging conditions. Int. J. Food Microbiol. 2018, 280, 87–99. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biogenic Amines | LOD/(mg/kg) | RSD/% | Regression Equation | R2 |
|---|---|---|---|---|
| PUT | 0.80 | 2.36 | y = 29.554x − 5.7576 | 0.9991 |
| CAD | 0.80 | 2.74 | y = 30.636x − 32.384 | 0.9993 |
| HIS | 1.20 | 1.24 | y = 26.431x − 26.907 | 0.9990 |
| No. | RT (min) | Compound Name | Relative Percentage (%) | |
|---|---|---|---|---|
| Control | Test | |||
| Pyrazines | ||||
| A1 | 15.09 | 3-Ethyl-2,5-dimethyl-pyrazine | 55.40 | 56.31 |
| A2 | 11.41 | Trimethyl-pyrazine | 17.08 | 18.73 |
| A3 | 7.11 | 2,5-Dimethyl-pyrazine | 16.50 | 17.45 |
| A4 | 10.45 | 2,6-Dimethyl-pyrazine | 0.27 | 0.20 |
| A5 | 19.48 | 3,5-Diethyl-2-methyl-pyrazine | 2.64 | ND |
| A6 | 19.31 | 2,3,5-Trimethyl-6-ethylpyrazine | ND | 3.27 |
| Hydrocarbons | ||||
| B1 | 1.89 | Oxirane, 2-butyl-3-methyl-, cis- | 0.22 | ND |
| B2 | 37.35 | Hexa-t-butylthiatrisiletane | ND | 0.03 |
| B3 | 38.29 | Silane, [[4-[1,2-bis[(trimethylsilyl)oxy] ethyl]-1,2-phenylene] bis(oxy)] bis [trimethyl-] | ND | 0.04 |
| Esters | ||||
| C1 | 5.08 | Arsenous acid, tris(trimethylsilyl) ester | 0.49 | ND |
| C2 | 32.58 | Benzenesulfonic acid, 4-methyl-, bicyclo [3.2.1] oct-6-yl ester | 0.05 | ND |
| Ethers | ||||
| D1 | 38.29 | 18-Methyl-nonadecane-1,2-dio, trimethylsilyl ether | 0.06 | ND |
| Acids | ||||
| E1 | 1.59 | 12-Methylaminolauric acid | ND | 1.36 |
| Ketones | ||||
| F1 | 25.76 | Thebacon | ND | 0.07 |
| Alcohols | ||||
| G1 | 1.40 | Cyclobutanol | ND | 1.62 |
| G2 | 51.51 | trans-Farnesol | ND | 0.18 |
| Other compounds | ||||
| H1 | 1.42 | 2-Propanamine | 3.82 | 0.59 |
| H2 | 1.61 | Octodrine | 3.20 | ND |
| H3 | 3.78 | 1,2,4-trimethyl-Piperazine | 0.17 | ND |
| H4 | 49.20 | 4H-1-Benzopyran-2-carbonyl chloride, 4-oxo- | 0.04 | ND |
| H5 | 25.76 | 2-Chloro-4-(4-methoxyphenyl)-6-(4-nitrophenyl) pyrimidine | 0.08 | ND |
| H6 | 3.75 | 1,3,3-Trimethyl-diaziridine | ND | 0.07 |
| H7 | 29.84 | 1-Ethoxy-2-methyl-benzene | ND | 0.06 |
| H8 | 47.68 | 5-Fluoroorotyl-N, N-dimethylhydrazide | ND | 0.01 |
| H9 | 52.08 | 1H-Indole, 2-(1,1-dimethyl-2-propenyl)-6-(3-methyl-2-butenyl)- | ND | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhang, Y.; Ma, X.; Zhang, G.; Hou, H. Effects of a Novel Starter Culture on Quality Improvement and Putrescine, Cadaverine, and Histamine Inhibition of Fermented Shrimp Paste. Foods 2023, 12, 2833. https://doi.org/10.3390/foods12152833
Li X, Zhang Y, Ma X, Zhang G, Hou H. Effects of a Novel Starter Culture on Quality Improvement and Putrescine, Cadaverine, and Histamine Inhibition of Fermented Shrimp Paste. Foods. 2023; 12(15):2833. https://doi.org/10.3390/foods12152833
Chicago/Turabian StyleLi, Xinyu, Yang Zhang, Xinxiu Ma, Gongliang Zhang, and Hongman Hou. 2023. "Effects of a Novel Starter Culture on Quality Improvement and Putrescine, Cadaverine, and Histamine Inhibition of Fermented Shrimp Paste" Foods 12, no. 15: 2833. https://doi.org/10.3390/foods12152833
APA StyleLi, X., Zhang, Y., Ma, X., Zhang, G., & Hou, H. (2023). Effects of a Novel Starter Culture on Quality Improvement and Putrescine, Cadaverine, and Histamine Inhibition of Fermented Shrimp Paste. Foods, 12(15), 2833. https://doi.org/10.3390/foods12152833