

Evaluation of the Potential Anti-Inflammatory Activity of Black Rice in the Framework of Celiac Disease

 , , , , , , , ,
, , , , , , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Rice Samples and Preparation of the Hydro-Alcoholic Extracts
2.3. Peptic/Tryptic In Vitro Digestion
2.4. Characterization of Phenolic Profile and Antioxidant Capacity of Rice Extracts
2.4.1. Determination of Total Phenolic Content
2.4.2. Determination of Total Anthocyanin Content
2.4.3. Evaluation of the Antioxidant Capacity
DPPH Assay
TEAC Assay
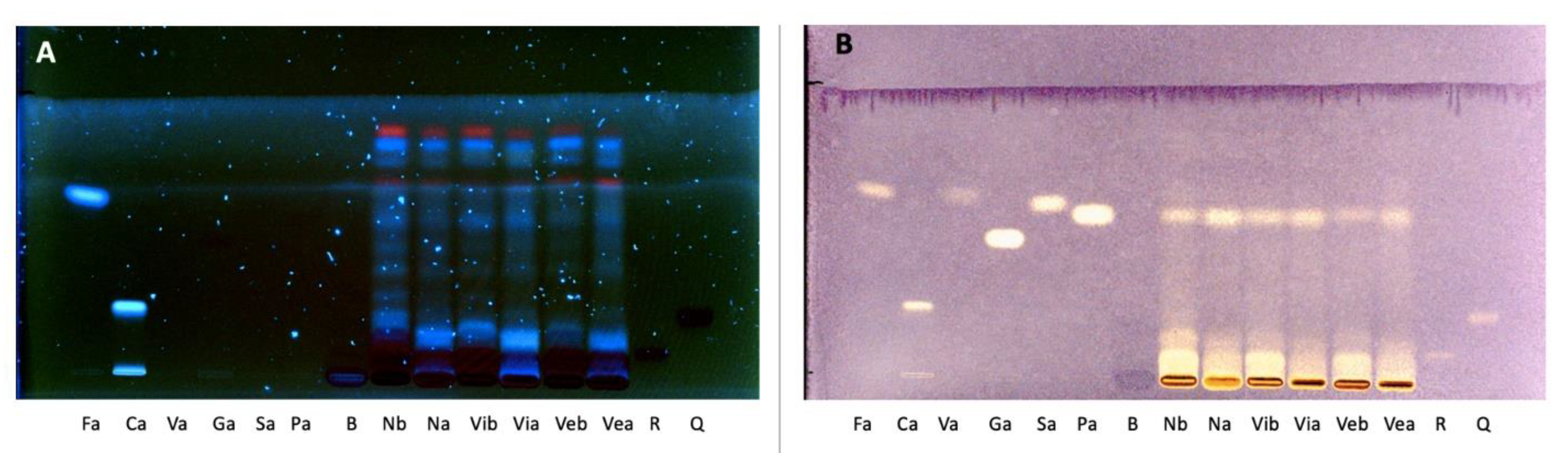
2.4.4. High-Performance Thin Layer Chromatography (HPTLC)
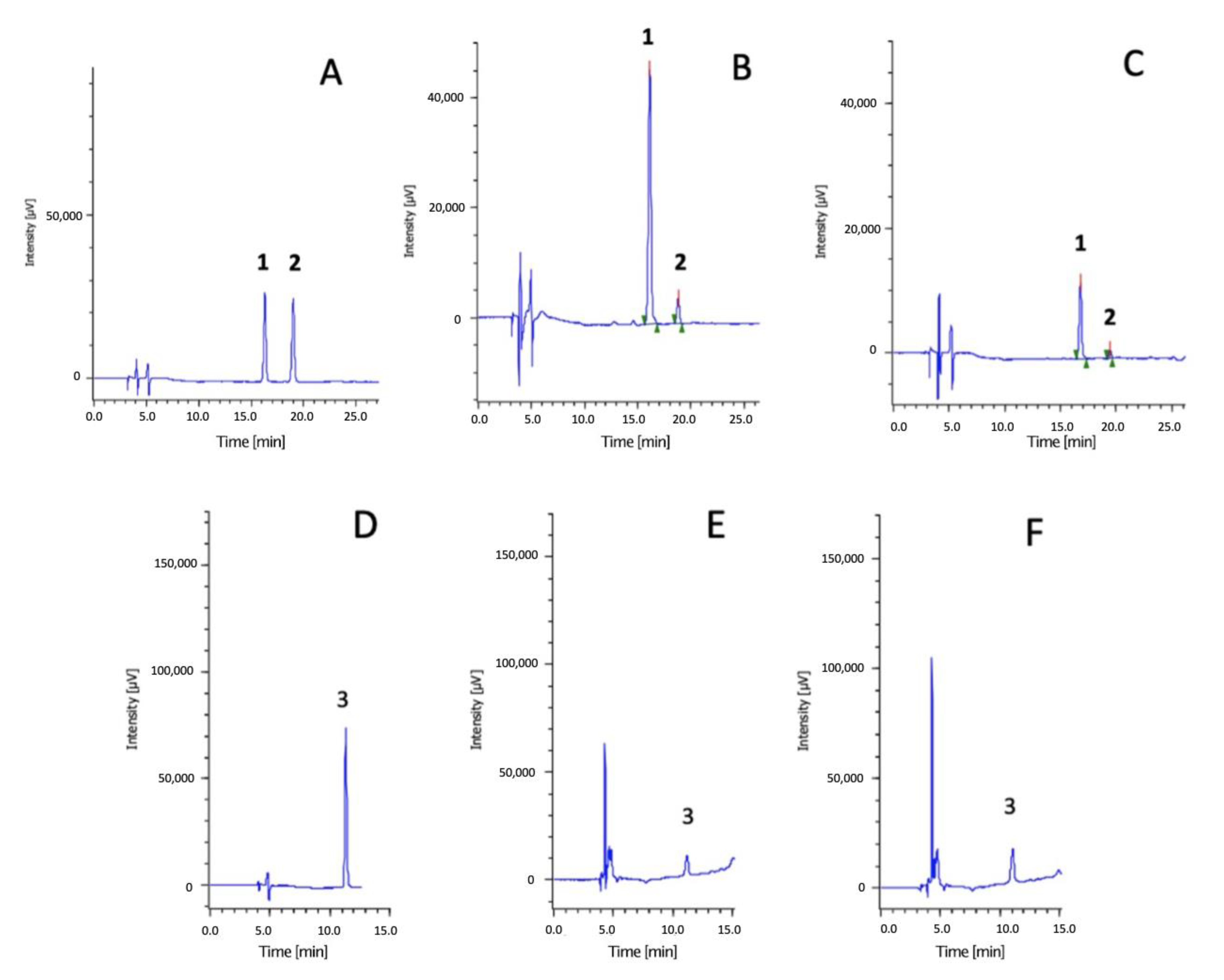
2.4.5. High-Performance Liquid Chromatography-Diode Array Detector (HPLC-DAD) Analysis
2.5. In Vitro Evaluation of the Anti-Inflammatory and Antioxidant Activity
2.5.1. Cell Culture and Treatment
2.5.2. Measurement of Cell Viability
2.5.3. Measurement of Soluble Inflammatory Markers by ELISA Assay
2.5.4. Measurement of the NF-κB-Driven Transcription
2.5.5. Reactive Oxygen Species Analysis in Caco-2 Cells
2.6. K562(S) Cell Agglutination Test
2.7. Statistical Analysis
3. Results and Discussion
3.1. Characterization of Rice Extracts
3.1.1. Evaluation of Phenolic Profile and Antioxidant Capacity
3.1.2. Identification and Quantification of Phenolic Compounds Applying Chromatographic Techniques
3.2. In Vitro Evaluation of the Biological Activity of Extracts
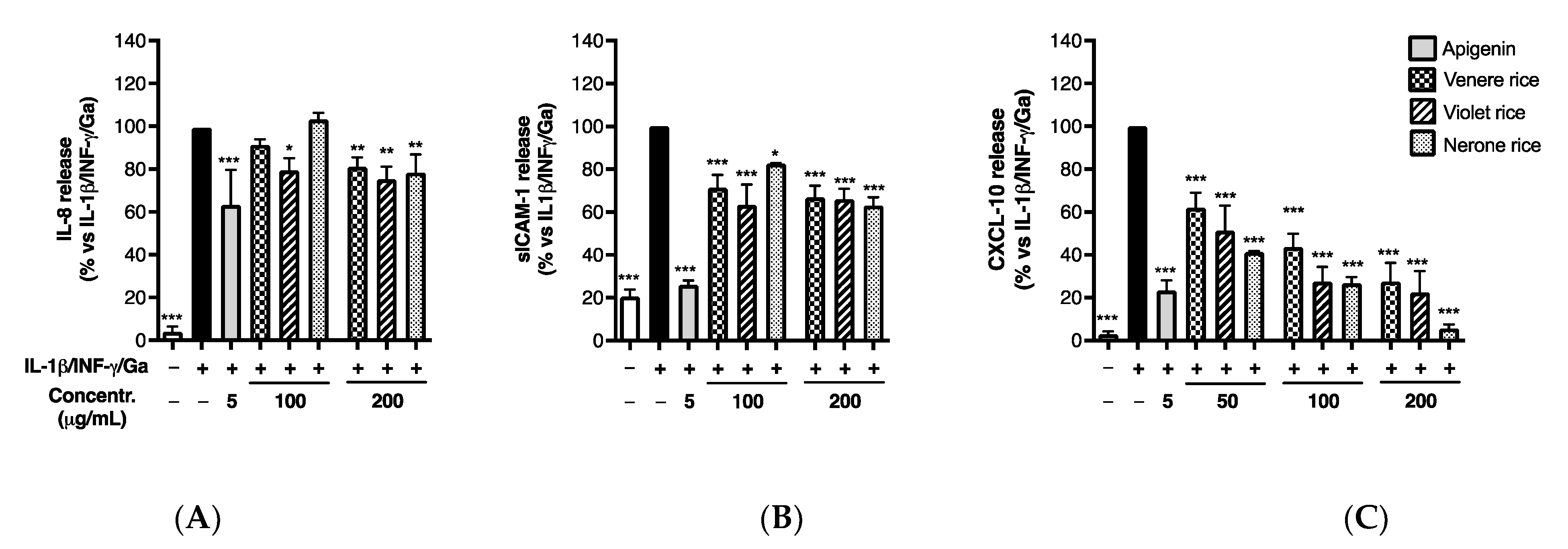
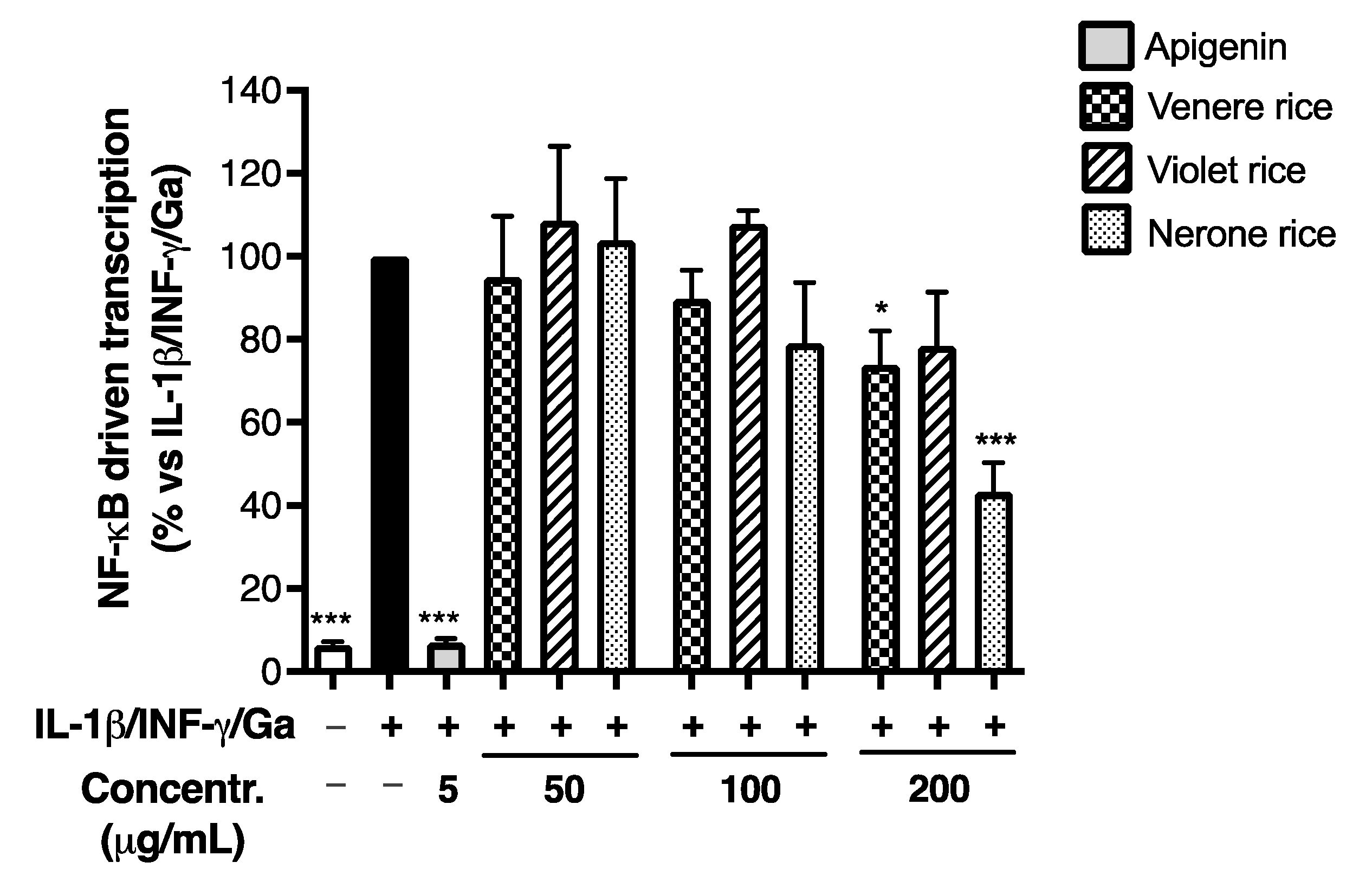
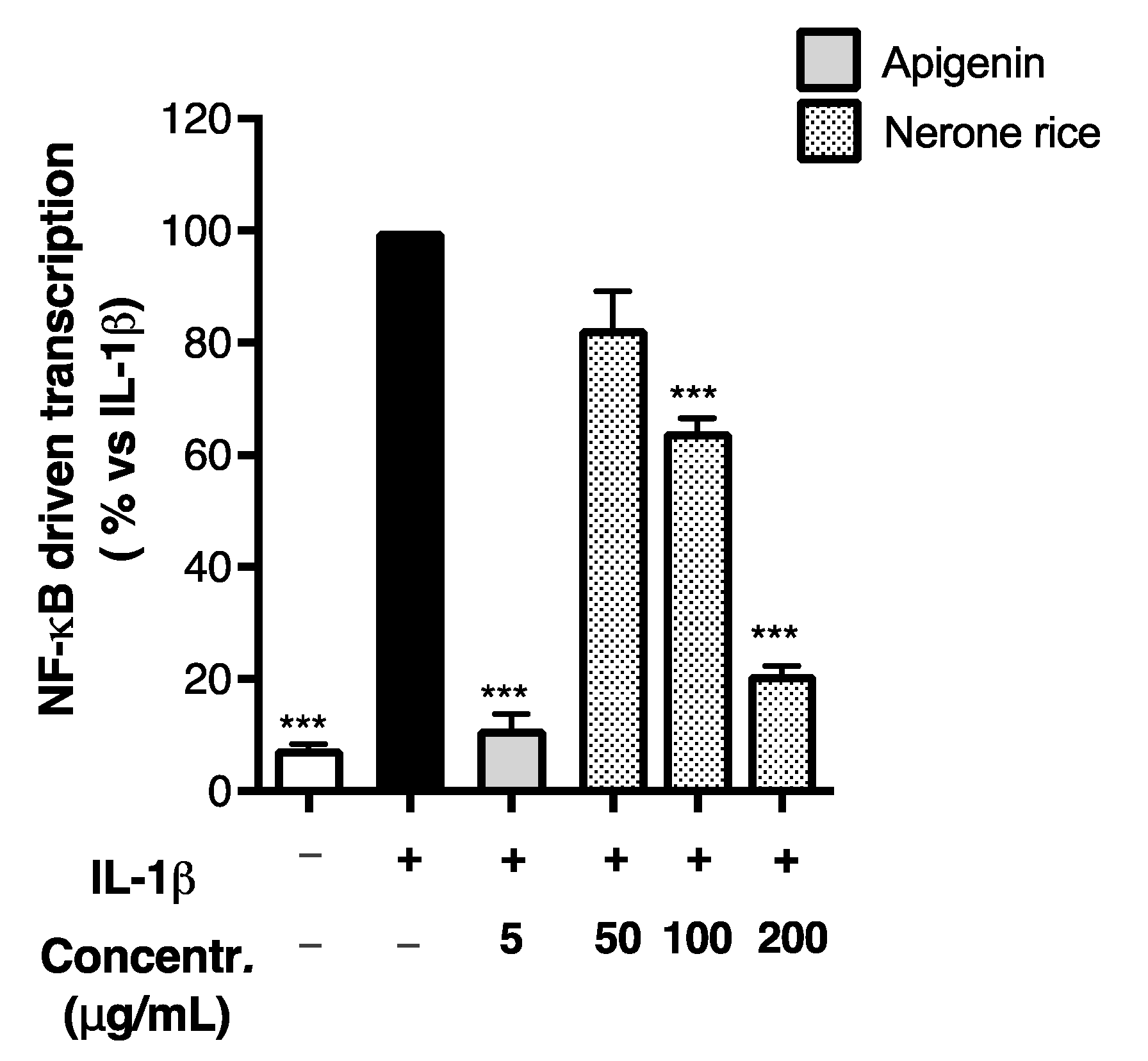
3.2.1. Effect of Rice Extracts on Inflammatory Markers in Caco-2 Cells
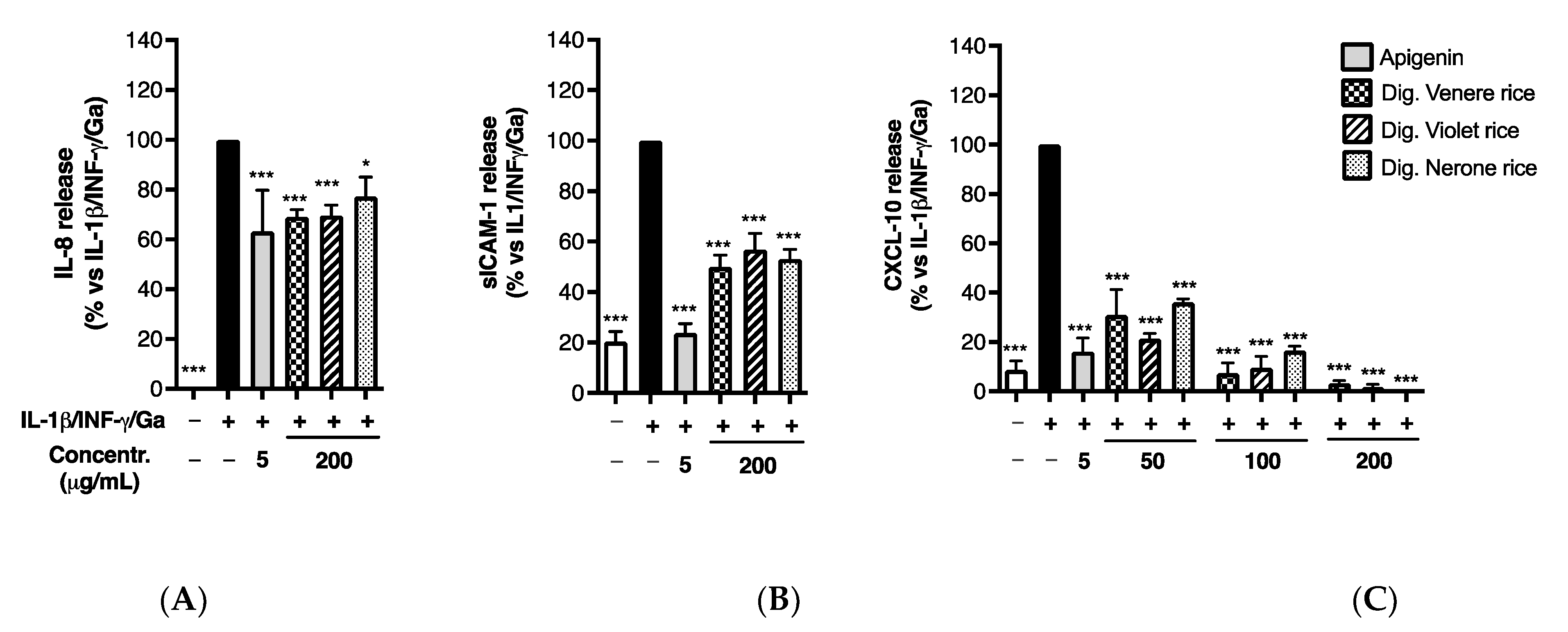
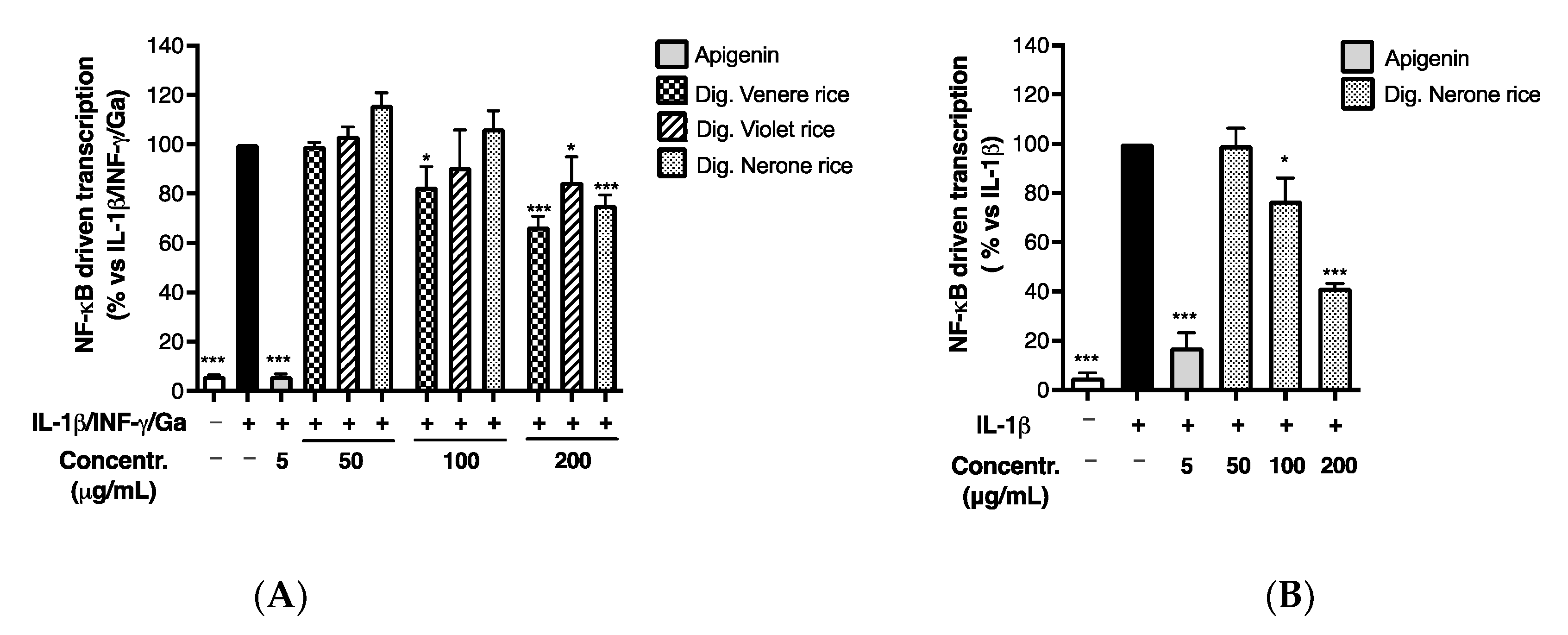
3.2.2. Impact of In Vitro Simulated Digestion on Anti-Inflammatory Activity
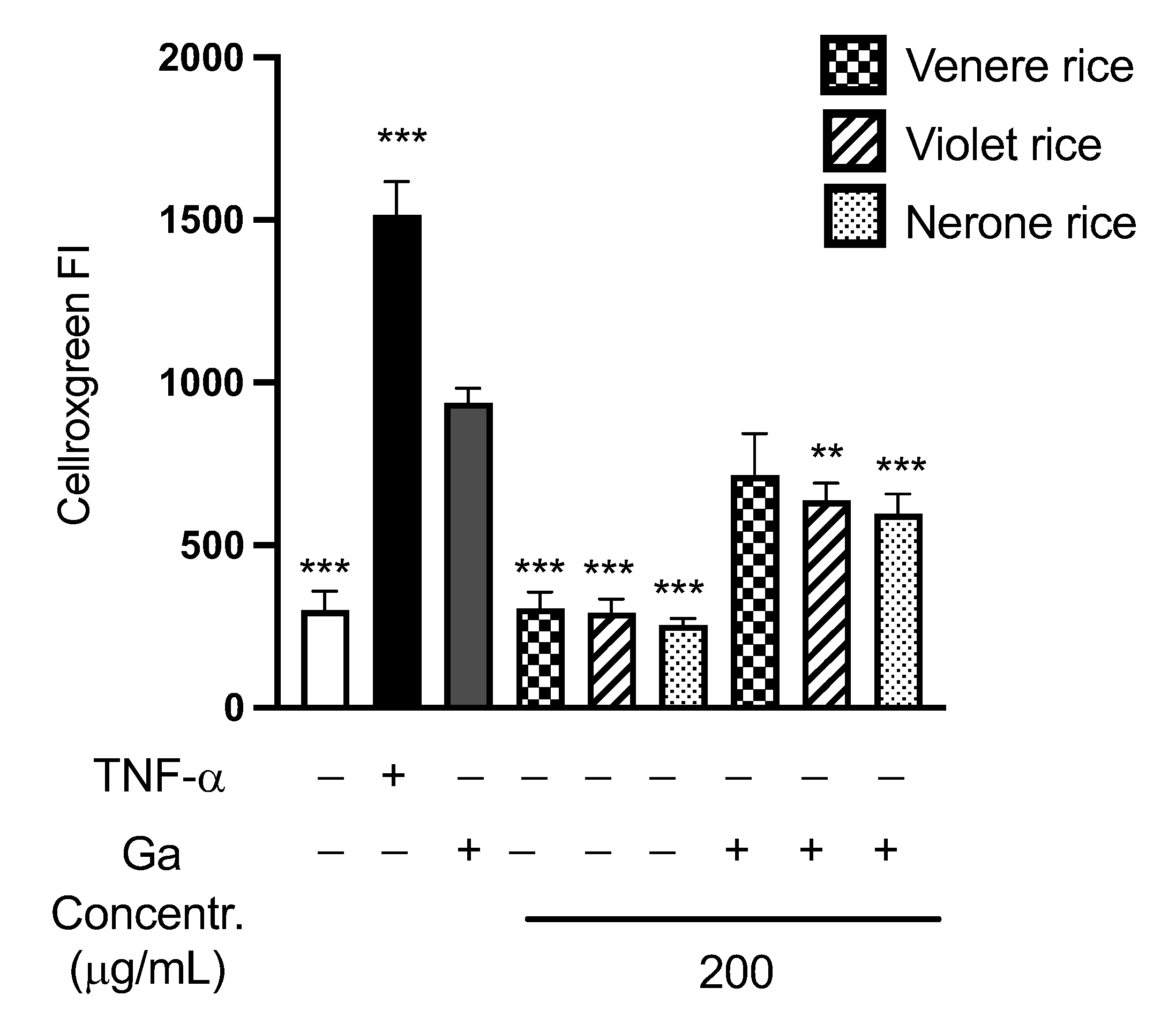
3.2.3. In Vitro Antioxidant Activity of Rice Extracts
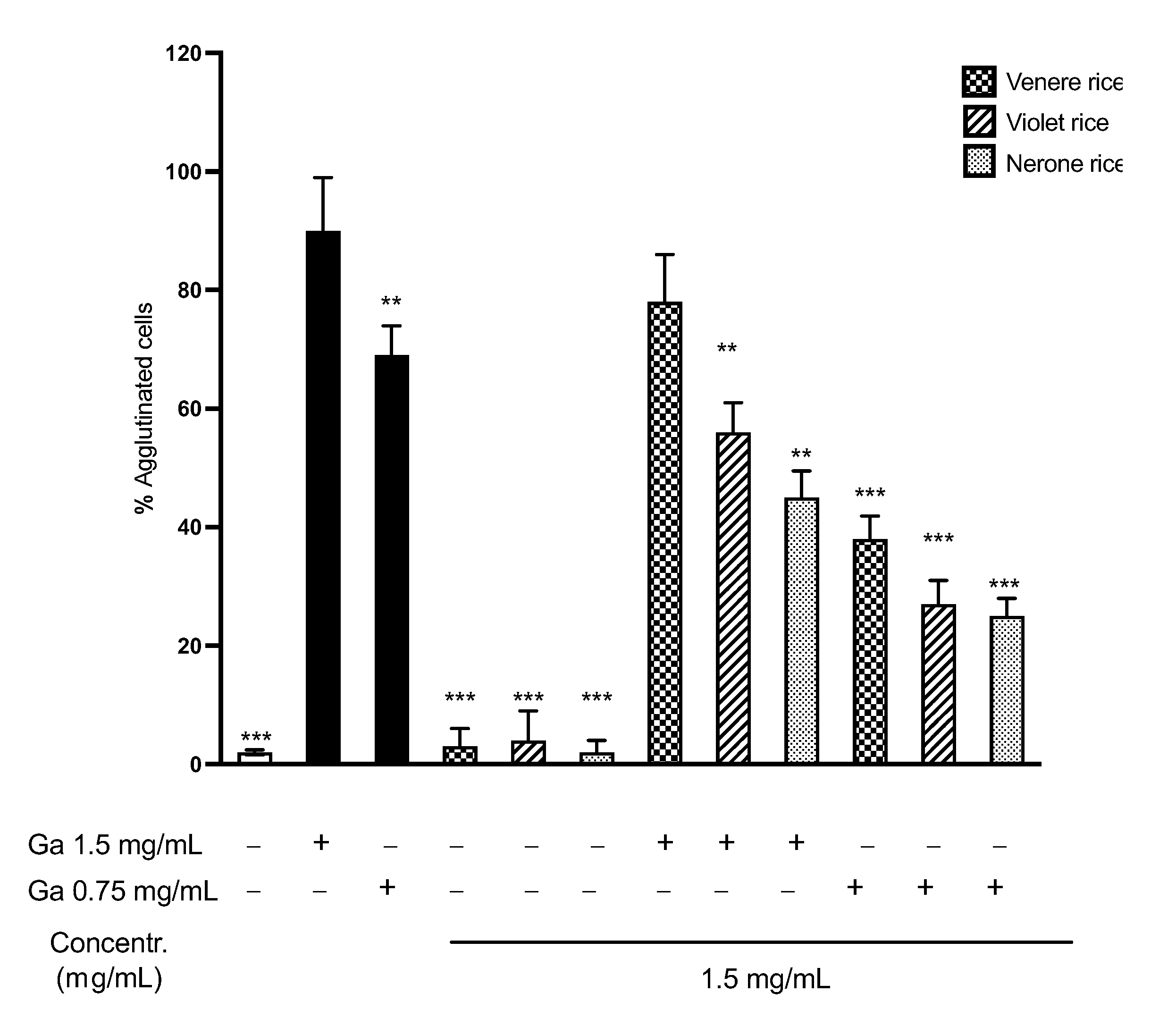
3.2.4. K562(S) Cell Agglutination Test
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, P.; Arora, A.; Strand, T.A.; Leffler, D.A.; Catassi, C.; Green, P.H.; Kelly, C.P.; Ahuja, V.; Makharia, G.K. Global Prevalence of Celiac Disease: Systematic Review and Meta-analysis. Clin. Gastroenterol. Hepatol. 2018, 16, 823–836.e2. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, G.; Bacchetti, T.; Masciangelo, S.; Saturni, L. Celiac disease, inflammation and oxidative damage: A nutrigenetic approach. Nutrients 2012, 4, 243–257. [Google Scholar] [PubMed]
- Ramírez-sánchez, A.D.; Tan, I.L.; Gonera-De Jong, B.C.; Visschedijk, M.C.; Jonkers, I.; Withoff, S. Molecular biomarkers for celiac disease: Past, present and future. Int. J. Mol. Sci. 2020, 21, 8528. [Google Scholar] [CrossRef]
- Van De Walle, J.; Hendrickx, A.; Romier, B.; Larondelle, Y.; Schneider, Y.J. Inflammatory parameters in Caco-2 cells: Effect of stimuli nature, concentration, combination and cell differentiation. Toxicol. In Vitro 2010, 24, 1441–1449. [Google Scholar] [CrossRef]
- Moretti, S.; Mrakic-Sposta, S.; Roncoroni, L.; Vezzoli, A.; Dellanoce, C.; Monguzzi, E.; Branchi, F.; Ferretti, F.; Lombardo, V.; Doneda, L.; et al. Oxidative stress as a biomarker for monitoring treated celiac disease article. Clin. Transl. Gastroenterol. 2018, 9, 157. [Google Scholar] [CrossRef] [PubMed]
- Rowicka, G.; Czaja-Bulsa, G.; Chełchowska, M.; Riahi, A.; Strucińska, M.; Weker, H.; Ambroszkiewicz, J. Oxidative and antioxidative status of children with celiac disease treated with a gluten free-diet. Oxid. Med. Cell. Longev. 2018, 2018, 1324820. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, G.; Bacchetti, T.; Saturni, L.; Manzella, N.; Candelaresi, C.; Benedetti, A.; Di Sario, A. Lipid Peroxidation and Paraoxonase-1 Activity in Celiac Disease. J. Lipids 2012, 2012, 587479. [Google Scholar] [CrossRef] [PubMed]
- Szaflarska-Popławska, A.; Siomek, A.; Czerwionka-Szaflarska, M.; Gackowski, D.; Rózalski, R.; Guz, J.; Szpila, A.; Zarakowska, E.; Oliński, R. Oxidatively damaged DNA/oxidative stress in children with celiac disease. Cancer Epidemiol. Biomarkers Prev. 2010, 19, 1960–1965. [Google Scholar] [CrossRef]
- Odetti, P.; Valentini, S.; Aragno, I.; Garibaldi, S.; Pronzato, M.A.; Rolandi, E.; Barreca, T. Oxidative stress in subjects affected by celiac disease. Free Radic. Res. 1998, 29, 17–24. [Google Scholar] [CrossRef]
- Capozzi, A.; Vincentini, O.; Gizzi, P.; Porzia, A.; Longo, A.; Felli, C.; Mattei, V.; Mainiero, F.; Silano, M.; Sorice, M.; et al. Modulatory Effect of Gliadin Peptide 10-mer on Epithelial Intestinal CACO-2 Cell Inflammatory Response. PLoS ONE 2013, 8, e66561. [Google Scholar] [CrossRef]
- Gupta, K.B.; Upadhyay, S.; Saini, R.G.; Mantha, A.K.; Dhiman, M. Inflammatory response of gliadin protein isolated from various wheat varieties on human intestinal cell line. J. Cereal Sci. 2018, 81, 91–98. [Google Scholar] [CrossRef]
- Gupta, K.B.; Mantha, A.K.; Dhiman, M. Mitigation of Gliadin-Induced Inflammation and Cellular Damage by Curcumin in Human Intestinal Cell Lines. Inflammation 2021, 44, 873–889. [Google Scholar] [CrossRef] [PubMed]
- Van Buiten, C.B.; Lambert, J.D.; Elias, R.J. Green Tea Polyphenols Mitigate Gliadin-Mediated Inflammation and Permeability in Vitro. Mol. Nutr. Food Res. 2018, 62, e1700879. [Google Scholar] [CrossRef] [PubMed]
- Valitutti, F.; Iorfida, D.; Anania, C.; Trovato, C.M.; Montuori, M.; Cucchiara, S.; Catassi, C. Cereal consumption among subjects with celiac disease: A snapshot for nutritional considerations. Nutrients 2017, 9, 396. [Google Scholar] [CrossRef] [PubMed]
- Jnawali, P.; Kumar, V.; Tanwar, B. Celiac disease: Overview and considerations for development of gluten-free foods. Food Sci. Hum. Wellness 2016, 5, 169–176. [Google Scholar] [CrossRef]
- Shen, Y.; Jin, L.; Xiao, P.; Lu, Y.; Bao, J. Total phenolics, flavonoids, antioxidant capacity in rice grain and their relations to grain color, size and weight. J. Cereal Sci. 2009, 49, 106–111. [Google Scholar] [CrossRef]
- Umadevi, M.; Pushpa, R.; Sampathkumar, K.P.; Bhowmik, D. Rice-Traditional Medicinal Plant in India. J. Pharmacogn. Phytochem. 2012, 1, 6–12. [Google Scholar]
- Mbanjo, E.G.N.; Kretzschmar, T.; Jones, H.; Ereful, N.; Blanchard, C.; Boyd, L.A.; Sreenivasulu, N. The Genetic Basis and Nutritional Benefits of Pigmented Rice Grain. Front. Genet. 2020, 11, 229. [Google Scholar] [CrossRef]
- Tanaka, J.; Nakamura, S.; Tsuruma, K.; Shimazawa, M.; Shimoda, H.; Hara, H. Purple Rice (Oryza sativa L.) extract and its constituents inhibit VEGF-induced angiogenesis. Phyther. Res. 2012, 26, 214–222. [Google Scholar] [CrossRef]
- Kasote, D.; Tiozon, R.N.; Sartagoda, K.J.D.; Itagi, H.; Roy, P.; Kohli, A.; Regina, A.; Sreenivasulu, N. Food Processing Technologies to Develop Functional Foods With Enriched Bioactive Phenolic Compounds in Cereals. Front. Plant Sci. 2021, 12, 771276. [Google Scholar] [CrossRef]
- Sangiovanni, E.; Di Lorenzo, C.; Colombo, E.; Colombo, F.; Fumagalli, M.; Frigerio, G.; Restani, P.; Dell’Agli, M. The effect of in vitro gastrointestinal digestion on the anti-inflammatory activity of Vitis vinifera L. leaves. Food Funct. 2015, 6, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, A.; Ponnuchamy, M.; Kumar, P.S.; Kapoor, A.; Vo, D.V.N.; Prabhakar, S. Techniques and modeling of polyphenol extraction from food: A review. Environ. Chem. Lett. 2021, 19, 3409–3443. [Google Scholar] [PubMed]
- Ballabio, C.; Uberti, F.; Di Lorenzo, C.; Brandolini, A.; Penas, E.; Restani, P. Biochemical and immunochemical characterization of different varieties of amaranth (Amaranthus L. ssp.) as a safe ingredient for gluten-free products. J. Agric. Food Chem. 2011, 59, 12969–12974. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic-phosphothungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- AOAC International. AOAC Official Method 2005.02 Total Monomeric Anthocyanin Pigment Content of Fruit Juices, Beverages, Natural Colorants, and Wines pH Differential Method. In Official Methods of Analysis of AOAC International; Horwitz, W., Latimer, G.W.J., Eds.; AOAC International: Rockville, MD, USA, 2006; pp. 37–39. ISBN 0-935584-77-3. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Colombo, F.; Di Lorenzo, C.; Petroni, K.; Silano, M.; Pilu, R.; Falletta, E.; Biella, S.; Restani, P. Pigmented corn varieties as functional ingredients for gluten-free products. Foods 2021, 10, 1770. [Google Scholar] [CrossRef]
- Martinelli, G.; Angarano, M.; Piazza, S.; Fumagalli, M.; Magnavacca, A.; Pozzoli, C.; Khalilpour, S.; Dell’agli, M.; Sangiovanni, E. The Nutraceutical Properties of Sumac (Rhus coriaria L.) against Gastritis: Antibacterial and Anti-Inflammatory Activities in Gastric Epithelial Cells Infected with H. pylori. Nutrients 2022, 14, 1757. [Google Scholar] [CrossRef]
- Peter Hoensch, H.; Weigmann, B. Regulation of the intestinal immune system by flavonoids and its utility in chronic inflammatory bowel disease. World J. Gastroenterol. 2018, 24, 877–881. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Vincentini, O.; Pinto, D.; Polo, A.; Maialetti, F.; Porrelli, A.; Gobbetti, M. Nutrients Bioaccessibility and Anti-inflammatory Features of Fermented Bee Pollen: A Comprehensive Investigation. Front. Microbiol. 2021, 12, 622091. [Google Scholar] [CrossRef]
- DeLoughery, Z.; Luczak, M.W.; Zhitkovich, A. Monitoring Cr intermediates and reactive oxygen species with fluorescent probes during chromate reduction. Chem. Res. Toxicol. 2014, 27, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Silano, M.; Vincentini, O.; Luciani, A.; Felli, C.; Caserta, S.; Esposito, S.; Villella, V.R.; Pettoello-Mantovani, M.; Guido, S.; Maiuri, L. Early tissue transglutaminase-mediated response underlies K562(S)-cell gliadin-dependent agglutination. Pediatr. Res. 2012, 71, 532–538. [Google Scholar] [CrossRef]
- Min, B.; McClung, A.; Chen, M.H. Effects of hydrothermal processes on antioxidants in brown, purple and red bran whole grain rice (Oryza sativa L.). Food Chem. 2014, 159, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Vicente, A.; Gil-Izquierdo, A.; García-Viguera, C. In vitro gastrointestinal digestion study of pomegranate juice phenolic compounds, anthocyanins, and vitamin C. J. Agric. Food Chem. 2002, 50, 2308–2312. [Google Scholar] [CrossRef] [PubMed]
- McDougall, G.J.; Fyffe, S.; Dobson, P.; Stewart, D. Anthocyanins from red wine—Their stability under simulated gastrointestinal digestion. Phytochemistry 2005, 66, 2540–2548. [Google Scholar] [CrossRef]
- McGhie, T.K.; Walton, M.C. The bioavailability and absorption of anthocyanins: Towards a better understanding. Mol. Nutr. Food Res. 2007, 51, 702–713. [Google Scholar] [CrossRef]
- Sun, D.; Huang, S.; Cai, S.; Cao, J.; Han, P. Digestion property and synergistic effect on biological activity of purple rice (Oryza sativa L.) anthocyanins subjected to a simulated gastrointestinal digestion in vitro. Food Res. Int. 2015, 78, 114–123. [Google Scholar] [CrossRef]
- Melini, V.; Panfili, G.; Fratianni, A.; Acquistucci, R. Bioactive compounds in rice on Italian market: Pigmented varieties as a source of carotenoids, total phenolic compounds and anthocyanins, before and after cooking. Food Chem. 2019, 277, 119–127. [Google Scholar] [CrossRef]
- Peanparkdee, M.; Patrawart, J.; Iwamoto, S. Physicochemical stability and in vitro bioaccessibility of phenolic compounds and anthocyanins from Thai rice bran extracts. Food Chem. 2020, 329, 127157. [Google Scholar] [CrossRef]
- Correa-Betanzo, J.; Allen-Vercoe, E.; McDonald, J.; Schroeter, K.; Corredig, M.; Paliyath, G. Stability and biological activity of wild blueberry (Vaccinium angustifolium) polyphenols during simulated in vitro gastrointestinal digestion. Food Chem. 2014, 165, 522–531. [Google Scholar] [CrossRef]
- Ti, H.; Zhang, R.; Li, Q.; Wei, Z.; Zhang, M. Effects of cooking and in vitro digestion of rice on phenolic profiles and antioxidant activity. Food Res. Int. 2015, 76, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin-Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A.; Soszka, A. Changes in phenolic compounds and antioxidant activity of fruit musts and fruit wines during simulated digestion. Molecules 2020, 25, 5574. [Google Scholar] [CrossRef] [PubMed]
- Bhuvaneswari, S.; Gopala Krishnan, S.; Bollinedi, H.; Saha, S.; Ellur, R.K.; Vinod, K.K.; Singh, I.M.; Prakash, N.; Bhowmick, P.K.; Nagarajan, M.; et al. Genetic Architecture and Anthocyanin Profiling of Aromatic Rice From Manipur Reveals Divergence of Chakhao Landraces. Front. Genet. 2020, 11, 570731. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Donnarumma, G.; Napolitano, A.; Galvano, F.; Gallo, A.; Scalfi, L.; Fogliano, V. Protocatechuic acid is the major human metabolite of cyanidin-glucosides. J. Nutr. 2007, 137, 2043–2048. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, H.; Zhang, W.; Ding, Y.; Zhao, T.; Zhang, M.; Mao, G.; Feng, W.; Wu, X.; Yang, L. Bioaccessibility and biotransformation of anthocyanin monomers following: In vitro simulated gastric-intestinal digestion and in vivo metabolism in rats. Food Funct. 2019, 10, 6052–6061. [Google Scholar] [CrossRef]
- Peanparkdee, M.; Yamauchi, R.; Iwamoto, S. Characterization of Antioxidants Extracted from Thai Riceberry Bran Using Ultrasonic-Assisted and Conventional Solvent Extraction Methods. Food Bioprocess Technol. 2018, 11, 713–722. [Google Scholar] [CrossRef]
- Drago, S.; El Asmar, R.; Di Pierro, M.; Clemente, M.G.; Tripathi, A.; Sapone, A.; Thakar, M.; Iacono, G.; Carroccio, A.; D’Agate, C.; et al. Gliadin, zonulin and gut permeability: Effects on celiac and non-celiac intestinal mucosa and intestinal cell lines. Scand. J. Gastroenterol. 2006, 41, 408–419. [Google Scholar] [CrossRef]
- Muto, E.; Dell’Agli, M.; Sangiovanni, E.; Mitro, N.; Fumagalli, M.; Crestani, M.; De Fabiani, E.; Caruso, D. Olive oil phenolic extract regulates interleukin-8 expression by transcriptional and posttranscriptional mechanisms in Caco-2 cells. Mol. Nutr. Food Res. 2015, 59, 1217–1221. [Google Scholar] [CrossRef]
- Schuerer-Maly, C.-C.; Eckmann, L.; Kagnoff, M.F.; Falco, M.T.; Malyt, F.-E. Colonic epithelial cell lines as a source of interleukin-8: Stimulation by inflammatory cytokines and bacterial lipopolysaccharide. Immunology 1994, 81, 85–91. [Google Scholar]
- Wu, T.; Grootaert, C.; Pitart, J.; Vidovic, N.K.; Kamiloglu, S.; Possemiers, S.; Glibetic, M.; Smagghe, G.; Raes, K.; Van de Wiele, T.; et al. Aronia (Aronia melanocarpa) Polyphenols Modulate the Microbial Community in a Simulator of the Human Intestinal Microbial Ecosystem (SHIME) and Decrease Secretion of Proinflammatory Markers in a Caco-2/endothelial Cell Coculture Model. Mol. Nutr. Food Res. 2018, 62, e1800607. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, D.; Cimino, F.; Fratantonio, D.; Molonia, M.S.; Bashllari, R.; Busà, R.; Saija, A.; Speciale, A. Cyanidin-3-O-Glucoside Modulates the In Vitro Inflammatory Crosstalk between Intestinal Epithelial and Endothelial Cells. Mediat. Inflamm. 2017, 2017, 3454023. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Digestion | TAC mg CY/g | TPC mg GAE/g | AOA | |
|---|---|---|---|---|---|
| DPPH mg GAE/g | TEAC mg TE/g | ||||
| Nerone | Before | 66.26 ± 1.54 | 159.92 ± 16.68 | 40.97 ± 0.85 | 301.71 ± 6.50 |
| After | 29.23 ± 2.80 ** | 175.87 ± 4.94 | 46.60 ± 0.32 * | 345.37 ± 23.04 * | |
| Violet | Before | 32.61 ± 0.41 | 124.09 ± 4.22 | 45.52 ± 0.49 | 240.54 ± 18.72 |
| After | 20.31 ± 1.37 ** | 89.53 ± 6.51 ** | 29.39 ± 2.26 ** | 170.35 ± 8.94 * | |
| Venere | Before | 34.28 ± 2.67 | 135.58 ± 2.11 | 43.19 ± 3.68 | 267.23 ± 11.48 |
| After | 21.57 ± 1.49 ** | 120.61 ± 10.99 | 38.27 ± 3.50 | 233.40 ± 4.84 * | |
| Samples | Digestion | Anthocyanins (mg/g) | Phenolic Acids (mg/g) | ||||
|---|---|---|---|---|---|---|---|
| CY | PE | Total (CY + PE) | % Variation | PA | % Variation | ||
| Nerone | Before | 43.440 ± 0.944 | 3.724 ± 0.048 | 47.165 ± 0.969 | 1.546 ± 0.016 | ||
| After | 11.384 ± 0.136 | 1.187 ± 0.010 | 12.571 ± 0.126 *** | −73.3 | 3.140 ± 0.032 *** | +103.0 | |
| Violet | Before | 22.334 ± 0.928 | 0.786 ± 0.024 | 23.120 ± 0.928 | 2.056 ± 0.097 | ||
| After | 6.258 ± 0.058 | 0.212 ± 0.002 | 6.470 ± 0.057 *** | −72.0 | 3.017 ± 0.056 *** | +46.75 | |
| Venere | Before | 19.242 ± 0.090 | 1.012 ± 0.007 | 20.254 ± 0.091 | 1.823 ± 0.010 | ||
| After | 4.723 ± 0.046 | 0.205 ± 0.001 | 4.928 ± 0.047 *** | −75.7 | 2.769 ± 0.037 *** | +51.91 | |
| Black Rice Variety | IC50 (μg/mL) |
|---|---|
| Ve | 76.44 ± 13.59 *** |
| Vi | 48.07 ± 13.02 *** |
| N | 21.53 ± 7.29 *** |
| Nerone Extract | IC50 (μg/mL) vs. IL-1β/IFN-γ/Ga | IC50 (μg/mL) vs. IL-1β |
|---|---|---|
| Before digestion | 165.90 ± 33.75 | 108.4 ± 9.57 |
| After digestion | >200 μg/mL | 164 ± 14.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piazza, S.; Colombo, F.; Bani, C.; Fumagalli, M.; Vincentini, O.; Sangiovanni, E.; Martinelli, G.; Biella, S.; Silano, M.; Restani, P.; et al. Evaluation of the Potential Anti-Inflammatory Activity of Black Rice in the Framework of Celiac Disease. Foods 2023, 12, 63. https://doi.org/10.3390/foods12010063
Piazza S, Colombo F, Bani C, Fumagalli M, Vincentini O, Sangiovanni E, Martinelli G, Biella S, Silano M, Restani P, et al. Evaluation of the Potential Anti-Inflammatory Activity of Black Rice in the Framework of Celiac Disease. Foods. 2023; 12(1):63. https://doi.org/10.3390/foods12010063
Chicago/Turabian StylePiazza, Stefano, Francesca Colombo, Corinne Bani, Marco Fumagalli, Olimpia Vincentini, Enrico Sangiovanni, Giulia Martinelli, Simone Biella, Marco Silano, Patrizia Restani, and et al. 2023. "Evaluation of the Potential Anti-Inflammatory Activity of Black Rice in the Framework of Celiac Disease" Foods 12, no. 1: 63. https://doi.org/10.3390/foods12010063
APA StylePiazza, S., Colombo, F., Bani, C., Fumagalli, M., Vincentini, O., Sangiovanni, E., Martinelli, G., Biella, S., Silano, M., Restani, P., Dell’Agli, M., & Di Lorenzo, C. (2023). Evaluation of the Potential Anti-Inflammatory Activity of Black Rice in the Framework of Celiac Disease. Foods, 12(1), 63. https://doi.org/10.3390/foods12010063