
Metabolite Profiling of Chestnut (Castanea crenata) According to Origin and Harvest Time Using 1H NMR Spectroscopy

Abstract
1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
2.2. Sample Collection
2.3. Sample Preparation
2.4. 1H NMR Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Metabolite Profiling of C. crenata by 1H NMR Spectroscopy
3.2. Metabolic Characterization Depending on C. crenata Chestnut Cultivar Origin
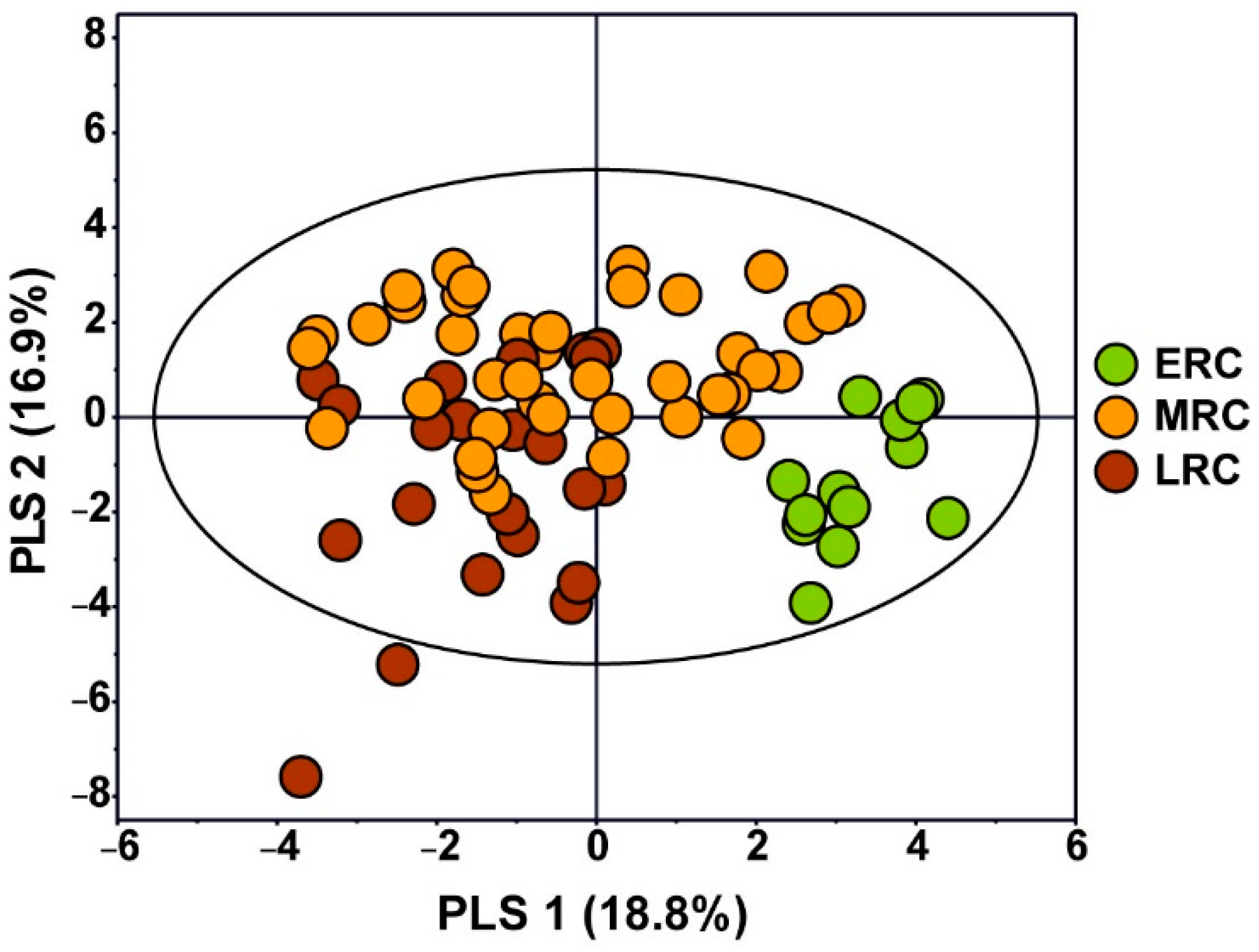
3.3. Metabolic Characterization Depending on Harvest Time of C. crenata Chestnut Cultivars
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chiocchio, I.; Prata, C.; Mandrone, M.; Ricciardiello, F.; Marrazzo, P.; Tomasi, P.; Angeloni, C.; Fiorentini, D.; Malaguti, M.; Poli, F. Leaves and spiny burs of Castanea Sativa from an experimental chestnut grove: Metabolomic analysis and anti-neuroinflammatory activity. Metabolites 2020, 10, 408. [Google Scholar] [CrossRef] [PubMed]
- Tsurunaga, Y.; Takahashi, T. Evaluation of the Antioxidant Activity, Deodorizing Effect, and Antibacterial Activity of ‘Porotan’Chestnut By-Products and Establishment of a Compound Paper. Foods 2021, 10, 1141. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.; Zambon, C.; Techio, V.; Pio, R. Floral characterization and pollen germination protocol for Castanea crenata Siebold & Zucc. S. Afr. J. Bot. 2020, 130, 389–395. [Google Scholar]
- Kang, M.-J.; Shin, A.-Y.; Shin, Y.; Lee, S.-A.; Lee, H.-R.; Kim, T.-D.; Choi, M.; Koo, N.; Kim, Y.-M.; Kyeong, D. Identification of transcriptome-wide, nut weight-associated SNPs in Castanea crenata. Sci. Rep. 2019, 9, 13161. [Google Scholar] [CrossRef]
- Korea Statistical Information Service. Available online: http://kosis.kr (accessed on 14 March 2022).
- Youn, U.-Y.; Shon, M.-S.; Kim, G.-N.; Katagiri, R.; Harata, K.; Ishida, Y.; Lee, S.-C. Antioxidant and anti-adipogenic activities of chestnut (Castanea crenata) byproducts. Food Sci. Biotechnol. 2016, 25, 1169–1174. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Díaz-Hernández, M.; Ciordia-Ara, M.; Ríos-Mesa, D. Chemical composition of chestnut cultivars from Spain. Sci. Hortic. 2006, 107, 306–314. [Google Scholar] [CrossRef]
- Tuyen, P.T.; Xuan, T.D.; Khang, D.T.; Ahmad, A.; Quan, N.V.; Tu Anh, T.T.; Anh, L.H.; Minh, T.N. Phenolic compositions and antioxidant properties in bark, flower, inner skin, kernel and leaf extracts of Castanea crenata Sieb. et Zucc. Antioxidants 2017, 6, 31. [Google Scholar] [CrossRef]
- Noh, J.-R.; Gang, G.-T.; Kim, Y.-H.; Yang, K.-J.; Hwang, J.-H.; Lee, H.-S.; Oh, W.-K.; Song, K.-S.; Lee, C.-H. Antioxidant effects of the chestnut (Castanea crenata) inner shell extract in t-BHP-treated HepG2 cells, and CCl4-and high-fat diet-treated mice. Food Chem. Toxicol. 2010, 48, 3177–3183. [Google Scholar] [CrossRef]
- Kim, M.; Lee, U.; Kim, S.; Hwang, M.; Lee, M. Comparison of nut characteristics between Korean native chestnut accessions and prevailing cultivars cultivated in Korea. In Proceedings of the III International Chestnut Congress 693, Chaves, Portugal, 20–23 October 2004; pp. 299–304. [Google Scholar]
- Yamamoto, T.; Shimada, T.; Kotobuki, K.; Morimoto, Y.; Yoshida, M. Genetic characterization of Asian chestnut varieties assessed by AFLP. Jpn. J. Breed. 1998, 48, 359–363. [Google Scholar] [CrossRef]
- Yamamoto, T.; Tanaka, T.; Kotobuki, K.; Matsuta, N.; Suzuki, M.; Hayashi, T. Characterization of simple sequence repeats in Japanese chestnut. J. Hortic. Sci. Biotechnol. 2003, 78, 197–203. [Google Scholar] [CrossRef]
- Seo, D.-J.; Chung, M.-J.; Kim, D.-J.; You, J.-K.; Choe, M. Nutritional constituent analysis of Korean chestnuts. J. Korean Soc. Food Sci. Nutr. 2009, 38, 166–176. [Google Scholar] [CrossRef]
- Jeong, H.-R.; Jo, Y.-N.; Jeong, J.-H.; Jin, D.-E.; Song, B.-G.; Jin, Y.-R.; Kim, M.-J.; Lee, U.; Heo, H.-J. Change in the chemical composition of chestnuts (Castanea crenata) from different periods. Korean J. Food Sci. Technol. 2012, 44, 393–400. [Google Scholar] [CrossRef]
- Hall, R.D. Plant metabolomics: From holistic hope, to hype, to hot topic. New Phytol. 2006, 169, 453–468. [Google Scholar] [CrossRef]
- Hall, R.D. Food metabolomics: META-PHOR. A new European research initiative. Agro FOOD Industry Hi Tech 2007, 18, 14. [Google Scholar]
- Son, H.-S.; Hwang, G.-S.; Kim, K.M.; Ahn, H.-J.; Park, W.-M.; Van Den Berg, F.; Hong, Y.-S.; Lee, C.-H. Metabolomic studies on geographical grapes and their wines using 1H NMR analysis coupled with multivariate statistics. J. Agric. Food Chem. 2009, 57, 1481–1490. [Google Scholar] [CrossRef]
- Porzel, A.; Farag, M.A.; Mülbradt, J.; Wessjohann, L.A. Metabolite profiling and fingerprinting of Hypericum species: A comparison of MS and NMR metabolomics. Metabolomics 2014, 10, 574–588. [Google Scholar] [CrossRef]
- Dixon, R.A.; Gang, D.R.; Charlton, A.J.; Fiehn, O.; Kuiper, H.A.; Reynolds, T.L.; Tjeerdema, R.S.; Jeffery, E.H.; German, J.B.; Ridley, W.P. Applications of metabolomics in agriculture. J. Agric. Food Chem. 2006, 54, 8984–8994. [Google Scholar] [CrossRef] [PubMed]
- Calò, F.; Girelli, C.R.; Wang, S.C.; Fanizzi, F.P. Geographical Origin Assessment of Extra Virgin Olive Oil via NMR and MS Combined with Chemometrics as Analytical Approaches. Foods 2022, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Hama, J.R.; Fitzsimmons-Thoss, V. Determination of Unsaturated Fatty Acids Composition in Walnut (Juglans regia L.) Oil Using NMR Spectroscopy. Food Anal. Methods 2022, 1–11. [Google Scholar] [CrossRef]
- Cacciola, N.A.; Cerrato, A.; Capriotti, A.L.; Cavaliere, C.; D’Apolito, M.; Montone, C.M.; Piovesana, S.; Squillaci, G.; Peluso, G.; Laganà, A. Untargeted characterization of chestnut (Castanea sativa Mill.) shell polyphenol extract: A valued bioresource for prostate cancer cell growth inhibition. Molecules 2020, 25, 2730. [Google Scholar] [CrossRef]
- Cerulli, A.; Napolitano, A.; Masullo, M.; Hošek, J.; Pizza, C.; Piacente, S. Chestnut shells (Italian cultivar “Marrone di Roccadaspide” PGI): Antioxidant activity and chemical investigation with in depth LC-HRMS/MSn rationalization of tannins. Food Res. Int. 2020, 129, 108787. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Lee, J.; Kim, H.K.; Moon, B.C.; Ji, Y.; Hwang, G.-S. Metabolite profiling of Curcuma species grown in different regions using 1 H NMR spectroscopy and multivariate analysis. Analyst 2012, 137, 5597–5606. [Google Scholar] [CrossRef]
- Lee, M.Y.; Moon, B.C.; Kwon, Y.K.; Jung, Y.; Oh, T.K.; Hwang, G.S. Discrimination of Polygonatum species and identification of novel markers using 1H NMR-and UPLC/Q-TOF MS-based metabolite profiling. J. Sci. Food Agric. 2016, 96, 3846–3852. [Google Scholar] [CrossRef]
- Jung, Y.; Ha, M.; Lee, J.; Ahn, Y.G.; Kwak, J.H.; Ryu, D.H.; Hwang, G.-S. Metabolite profiling of the response of burdock roots to copper stress. J. Agric. Food Chem. 2015, 63, 1309–1317. [Google Scholar] [CrossRef]
- Worley, B.; Powers, R. Multivariate analysis in metabolomics. Curr. Metab. 2013, 1, 92–107. [Google Scholar]
- Mattila, P.; Salo-Väänänen, P.; Könkö, K.; Aro, H.; Jalava, T. Basic composition and amino acid contents of mushrooms cultivated in Finland. J. Agric. Food Chem. 2002, 50, 6419–6422. [Google Scholar] [CrossRef] [PubMed]
- Phat, C.; Moon, B.; Lee, C. Evaluation of umami taste in mushroom extracts by chemical analysis, sensory evaluation, and an electronic tongue system. Food Chem. 2016, 192, 1068–1077. [Google Scholar] [CrossRef]
- De Vasconcelos, M.d.C.B.; Nunes, F.; Viguera, C.G.; Bennett, R.N.; Rosa, E.A.; Ferreira-Cardoso, J.V. Industrial processing effects on chestnut fruits (Castanea sativa Mill.) 3. Minerals, free sugars, carotenoids and antioxidant vitamins. Int. J. Food Sci. Technol. 2010, 45, 496–505. [Google Scholar] [CrossRef]
- Fernandes, Â.; Antonio, A.L.; Barros, L.; Barreira, J.C.; Bento, A.; Botelho, M.L.; Ferreira, I.C. Low dose γ-irradiation as a suitable solution for chestnut (Castanea sativa Miller) conservation: Effects on sugars, fatty acids, and tocopherols. J. Agric. Food Chem. 2011, 59, 10028–10033. [Google Scholar] [CrossRef]
- Barreira, J.; Pereira, J.A.; Oliveira, M.; Ferreira, I.C. Sugars profiles of different chestnut (Castanea sativa Mill.) and almond (Prunus dulcis) cultivars by HPLC-RI. Plant Foods Hum. Nutr. 2010, 65, 38–43. [Google Scholar] [CrossRef]
- Suárez, M.H.; Galdón, B.R.; Mesa, D.R.; Romero, C.D.; Rodríguez, E.R. Sugars, organic acids and total phenols in varieties of chestnut fruits from Tenerife (Spain). Food Nutr. Sci. 2012, 3, 705–715. [Google Scholar] [CrossRef]
- Ribeiro, B.; Rangel, J.; Valentão, P.c.; Andrade, P.B.; Pereira, J.A.; Bölke, H.; Seabra, R.M. Organic acids in two Portuguese chestnut (Castanea sativa Miller) varieties. Food Chem. 2007, 100, 504–508. [Google Scholar] [CrossRef]
- Servillo, L.; Giovane, A.; Casale, R.; Balestrieri, M.L.; Cautela, D.; Paolucci, M.; Siano, F.; Volpe, M.G.; Castaldo, D. Betaines and related ammonium compounds in chestnut (Castanea sativa Mill.). Food Chem. 2016, 196, 1301–1309. [Google Scholar] [CrossRef]
- Du, C.; Chen, W.; Wu, Y.; Wang, G.; Zhao, J.; Sun, J.; Ji, J.; Yan, D.; Jiang, Z.; Shi, S. Effects of GABA and vigabatrin on the germination of Chinese chestnut recalcitrant seeds and its implications for seed dormancy and storage. Plants 2020, 9, 449. [Google Scholar] [CrossRef] [PubMed]
- Bouche, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef]
- Kinnersley, A.M.; Turano, F.J. Gamma aminobutyric acid (GABA) and plant responses to stress. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
- Bouché, N.; Lacombe, B.t.; Fromm, H. GABA signaling: A conserved and ubiquitous mechanism. Trends Cell Biol. 2003, 13, 607–610. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Gilliham, M.; Xu, B. γ-Aminobutyric acid (GABA) signalling in plants. Cell. Mol. Life Sci. 2017, 74, 1577–1603. [Google Scholar] [CrossRef]
- Oupadissakoon, C.; Young, C.T.; Giesbrecht, F.G.; Perry, A. Effect of location and time of harvest on free amino acid and free sugar contents of Florigiant peanuts. Peanut Sci. 1980, 7, 61–67. [Google Scholar] [CrossRef]
- Jwa, C.-S.; Yang, Y.-T.; Koh, J.-S. Changes in free sugars, organic acids, free amino acids and minerals by harvest time and parts of Acanthopanax koreanum. Appl. Biol. Chem. 2000, 43, 106–109. [Google Scholar]
- Loewus, F.A.; Loewus, M.W. Myo-inositol: Its biosynthesis and metabolism. Annu. Rev. Plant Physiol. 1983, 34, 137–161. [Google Scholar] [CrossRef]
- Loewus, F. Biogenesis of Plant Cell Wall Polysaccharides; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Horbowicz, M.; Obendorf, R.L. Seed desiccation tolerance and storability: Dependence on flatulence-producing oligosaccharides and cyclitols—Review and survey. Seed Sci. Res. 1994, 4, 385–405. [Google Scholar] [CrossRef]
- Bohnert, H.J.; Jensen, R.G. Strategies for engineering water-stress tolerance in plants. Trends Biotechnol. 1996, 14, 89–97. [Google Scholar] [CrossRef]
- Obendorf, R.L. Oligosaccharides and galactosyl cyclitols in seed desiccation tolerance. Seed Sci. Res. 1997, 7, 63–74. [Google Scholar] [CrossRef]
- Peterbauer, T.; Puschenreiter, M.; Richter, A. Metabolism of galactosylononitol in seeds of Vigna umbellata. Plant Cell Physiol. 1998, 39, 334–341. [Google Scholar] [CrossRef][Green Version]
- Peterbauer, T.; Richter, A. Galactosylononitol and stachyose synthesis in seeds of adzuki bean: Purification and characterization of stachyose synthase. Plant Physiol. 1998, 117, 165–172. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Number of Samples | Cultivar Name | Origin | Crossbreeding | Origin Group | Harvest Time |
|---|---|---|---|---|---|---|
| Chungchengnam-do (latitude 36° N, Longitude 127° E) | 5 | Okkwang | Korea | crenata a | KN | Mid ripening |
| 5 | Jahong | Korea | crenata a | KN | Mid ripening | |
| 5 | Mipung | Korea | crenata a | KN | Late ripening | |
| 5 | Juok | Korea | crenata hybrid (Gwangjujoyul a × Okkwang a) | KN | Mid ripening | |
| 3 | Pyeonggi | Korea | crenata hybrid (Riheiguri × Ginyosi b) | KH | Mid ripening | |
| 4 | Daebo | Korea | crenata hybrid (Sangmyeon 1 a × Riheiguri) | KH | Mid ripening | |
| 4 | Idea | Korea | crenata hybrid (Ibuki × Sandae a) | KH | Late ripening | |
| 5 | Otanba | Japan | crenata b | JN | Mid ripening | |
| 5 | Ginyosi | Japan | crenata b | JN | Late ripening | |
| 5 | Tanzawa | Japan | crenata hybrid (Otomune b × Taishouwase b) | JN | Early ripening | |
| 5 | Tsukuba | Japan | crenata hybrid (Ganne b × Hayadama b) | JN | Mid ripening | |
| 4 | Ishizuuchi | Japan | crenata hybrid (Ganne b × Kasaharawase b) | JN | Late ripening | |
| 5 | Porotan | Japan | (crenata × bungeana) × crenata hybrid (550-40 × Tanzawa b) | JH | Early ripening | |
| 3 | Moriwase | Japan | crenata × bungeana hybrid (Pyungyangyul × Toyotamawase b) | JH | Early ripening | |
| 4 | Riheiguri | Japan | crenata × mollissima hybrid | JH | Mid ripening | |
| 4 | Hyogo57 | Japan | crenata × mollissima hybrid | JH | Mid ripening | |
| 5 | Banseki c | Japan | - | - | Late ripening |
| No. | Metabolites | 1H Chemical Shifts b | Formula |
|---|---|---|---|
| Amino acids | |||
| 1 | Alanine | 1.5 (d), 3.8 (q) | C3H7NO2 |
| 2 | Arginine | 1.7 (m), 1.9 (m), 3.2 (t), 3.8 (t) | C6H14N4O2 |
| 3 | Asparagine | 2.8 (q), 2.9 (dd), 3.9 (q), 6.9 (s) | C4H8N2O3 |
| 4 | Aspartate | 2.6 (q), 2.8 (dd), 3.8 (dd) | C4H7NO4 |
| 5 | Glutamate | 2.0 (m), 2.1 (m), 2.4 (m), 3.8 (q) | C5H9NO4 |
| 6 | Glutamine | 2.44 (m), 2.12 (m) | C5H10N2O3 |
| 7 | Histidine | 3.2 (m), 4.0 (t), 7.1 (s), 7.9 (s) | C6H9N3O2 |
| 8 | Isoleucine | 0.9 (t), 1.0 (d), 1.2 (m), 1.5 (m), 2.0 (m), 3.7 (d) | C6H13NO2 |
| 9 | Leucine | 0.9(d), 1.0 (d), 1.7 (m), 1.7 (m), 1.7 (m), 3.7 (q) | C6H13NO2 |
| 10 | Phenylalanine | 3.1 (q), 3.3 (q), 4.0 (q), 7.3 (d), 7.4 (m) | C9H11NO2 |
| 11 | Threonine | 1.3 (d), 3.6 (d), 4.3 (q) | C4H9NO3 |
| 12 | Trigonelline | 8.0 (t), 8.8 (dd), 9.1 (s) | C7H7NO2 |
| 13 | Tryptophan | 7.1 (t), 7.2 (t), 7.3 (s), 7.5 (d), 7.7 (d) | C11H12N2O2 |
| 14 | Tyrosine | 3.1 (q), 3.2 (q), 3.9 (q), 6.8 (d), 7.2 (d) | C9H11NO3 |
| 15 | Valine | 1.0 (d), 2.3 (m), 3.6 (d) | C5H11NO2 |
| Organic acids | |||
| 16 | Citric acid | 2.5 (d), 2.7 (d) | C6H8O7; |
| 17 | Formic acid | 8.4 (s) | CH2O2 |
| 18 | Fumaric acid | 6.5 (s) | C4H4O4 |
| 19 | Malic acid | 2.4 (q), 2.7 (dd), 4.3 (d) | C4H6O5 |
| 20 | Malonic acid | 3.1 (s) | C3H4O4 |
| 21 | Succinic acid | 2.3 (s) | C4H6O4 |
| Sugars | |||
| 22 | Fructose | 3.5–4.1 (m) | C6H12O6 |
| 23 | Glucose | 3.2 (q), 3.4 (m), 3.5 (q), 3.7 (m), 4.6 (d), 5.2 (d) | C6H12O6 |
| 24 | Sucrose | 3.4 (t), 3.5 (dd), 3.6 (dd), 3.7 (t), 3.8 (m), 4.0 (t), 4.2 (d), 5.4 (d) | C12H22O11 |
| Others | |||
| 25 | 4-Aminobutyrate | 1.9 (m), 2.3 (t), 3.0 (t) | C4H9NO2 |
| 26 | Betaine | 3.3 (s), 3.9 (s) | (CH3)3N+ CH2COO− |
| 27 | Choline | 3.2 (s), 3.5 (m), 4.0 (m) | C5H14NO |
| 28 | Ethanol | 1.18 (t), 3.6 (q) | C2H5OH |
| 29 | Ethanolamine | 3.1 (t), 3.8 (t) | C2H7NO |
| 30 | Myo-inositol | 3.2 (t), 3.5 (dd), 3.6 (t), 4.0 (t) | C6H12O6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.M.; Nam, M.; Kim, M.-S. Metabolite Profiling of Chestnut (Castanea crenata) According to Origin and Harvest Time Using 1H NMR Spectroscopy. Foods 2022, 11, 1325. https://doi.org/10.3390/foods11091325
Yu JM, Nam M, Kim M-S. Metabolite Profiling of Chestnut (Castanea crenata) According to Origin and Harvest Time Using 1H NMR Spectroscopy. Foods. 2022; 11(9):1325. https://doi.org/10.3390/foods11091325
Chicago/Turabian StyleYu, Ja Myung, Miso Nam, and Min-Sun Kim. 2022. "Metabolite Profiling of Chestnut (Castanea crenata) According to Origin and Harvest Time Using 1H NMR Spectroscopy" Foods 11, no. 9: 1325. https://doi.org/10.3390/foods11091325
APA StyleYu, J. M., Nam, M., & Kim, M.-S. (2022). Metabolite Profiling of Chestnut (Castanea crenata) According to Origin and Harvest Time Using 1H NMR Spectroscopy. Foods, 11(9), 1325. https://doi.org/10.3390/foods11091325