
In Vitro Antibacterial Mechanism of High-Voltage Electrostatic Field against Acinetobacter johnsonii

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. HVEF Treatment
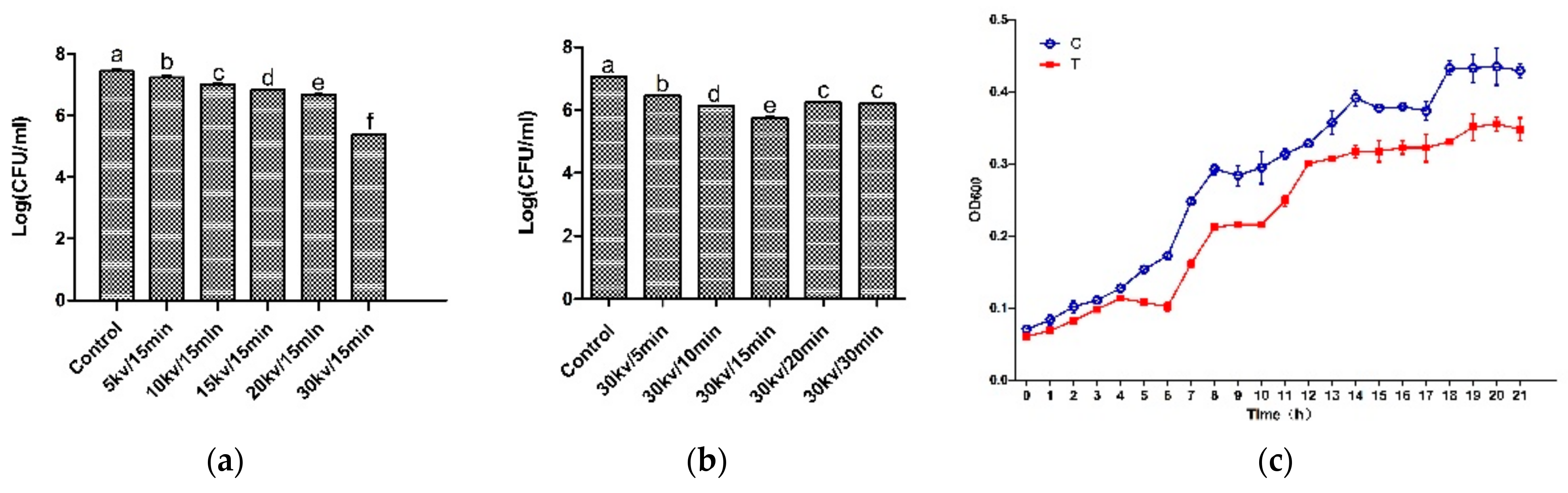
2.3. Bacterial Growth Curve
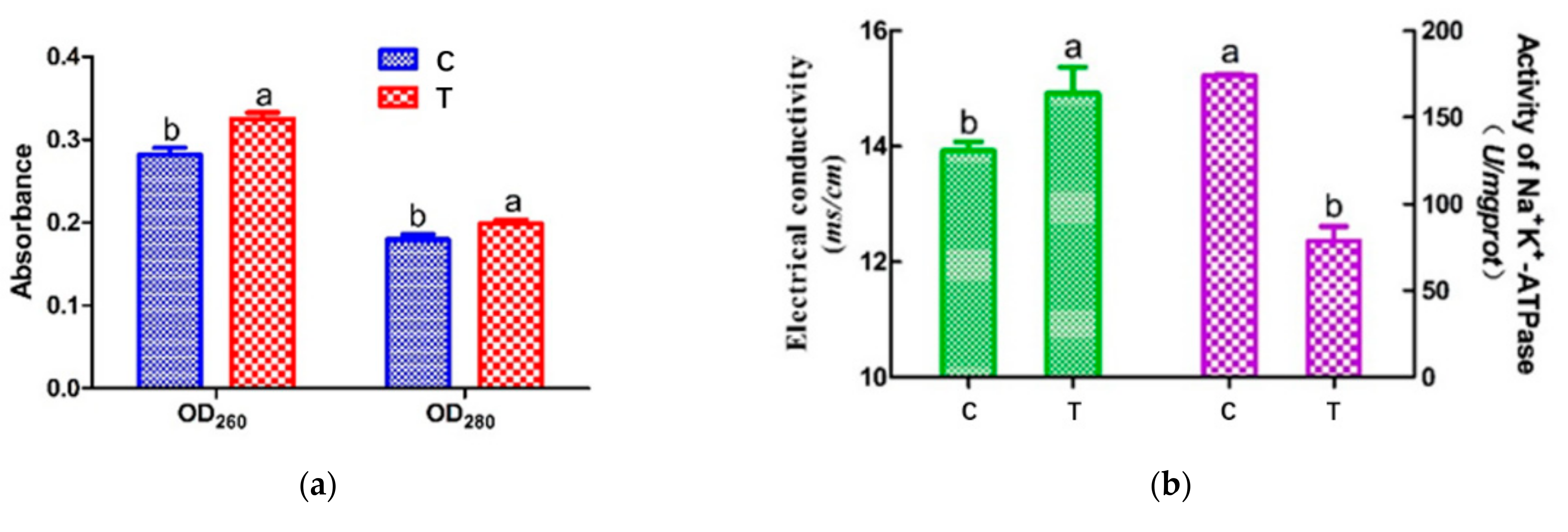
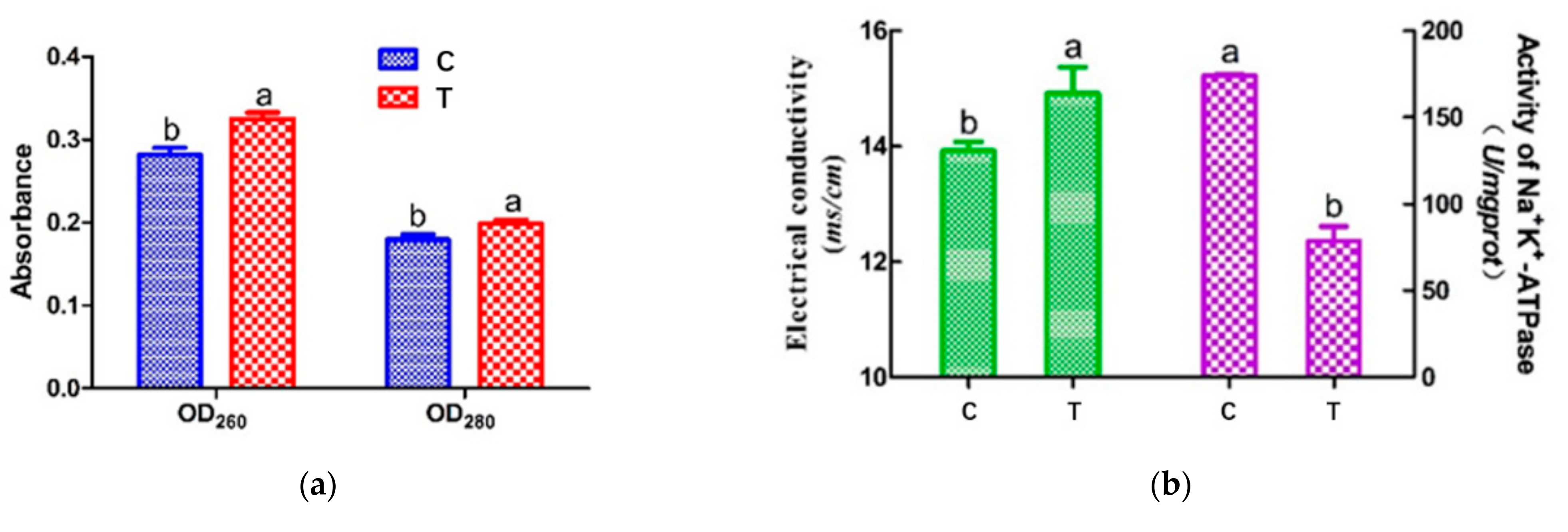
2.4. Cell Leakage Determination through UV Absorption
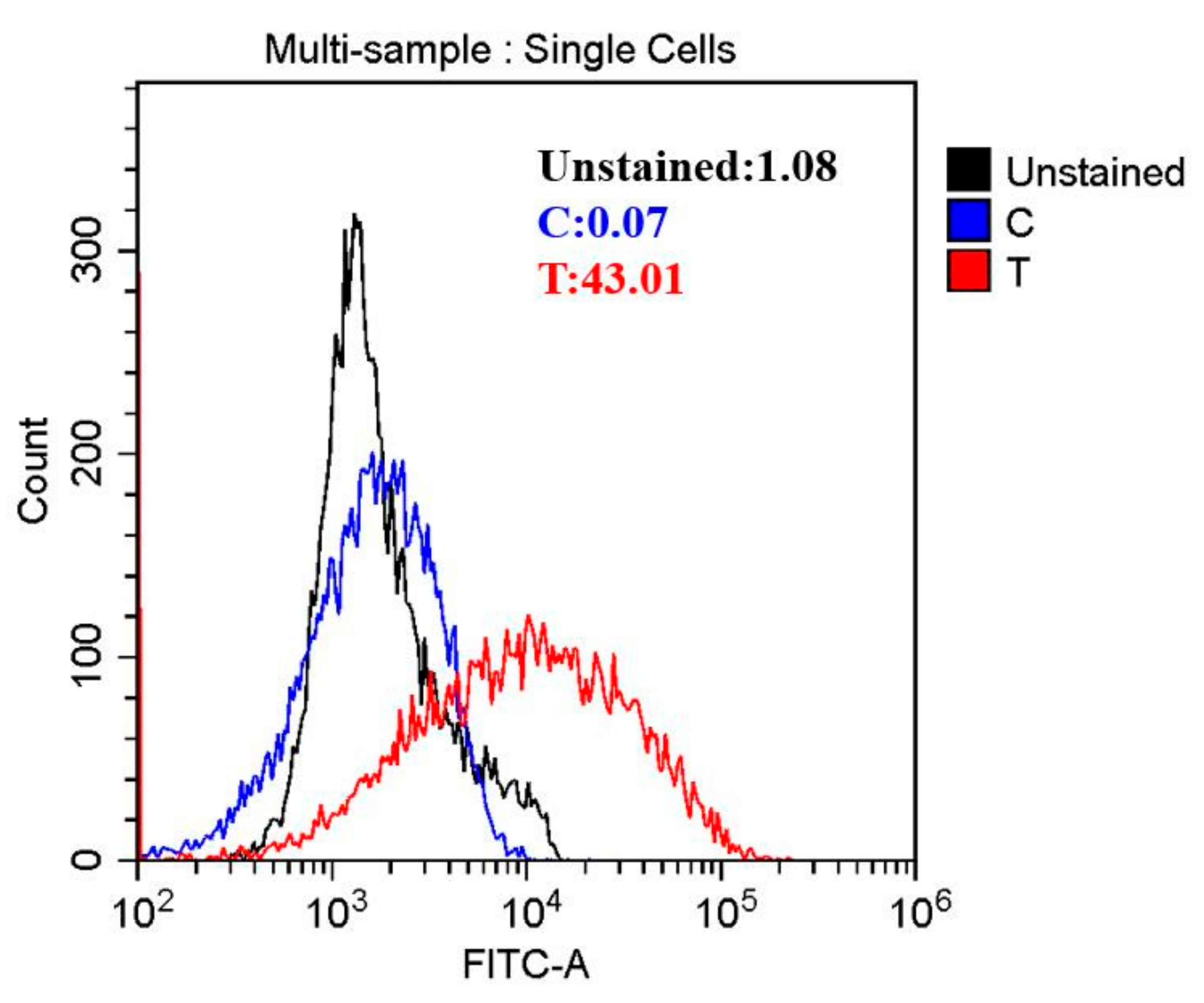
2.5. Cell Membrane Permeability Determination
2.6. Activity of Na+ K+-ATPase
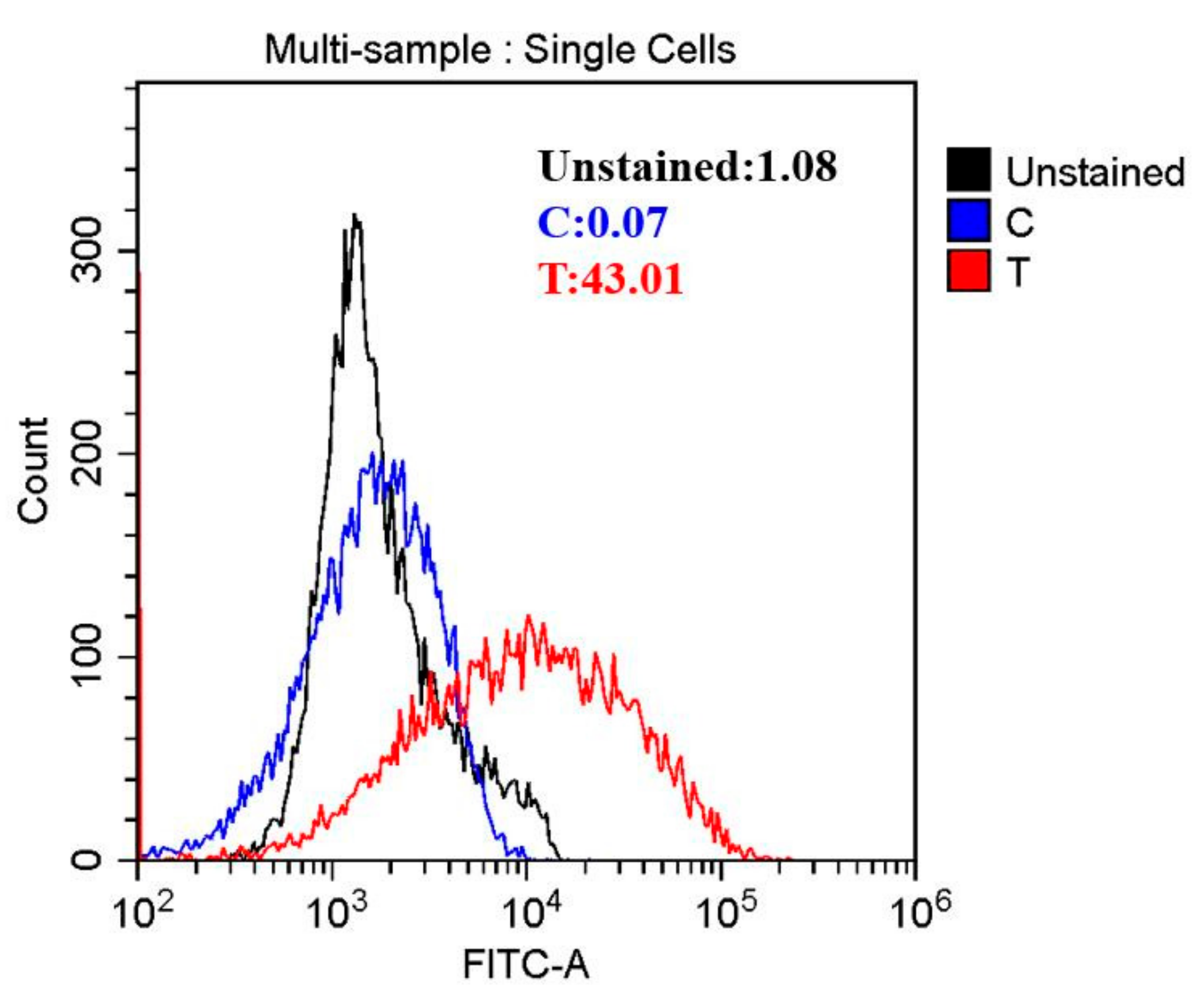
2.7. Intracellular ROS Concentration
2.8. SEM, TEM Observation and Negative Staining of Bacterium
2.9. Gene Expression Analysis
2.10. Statistical Analysis
3. Results and Discussion
3.1. Effect of an HVEF on A. Johnsonii Growth
3.2. Cell Leakage Determination by UV Absorption
3.3. Electrical Conductivity (ETC) and Na+ K+-ATPase Activity of Cell Membrane
3.4. Oxidative Stress Induced by HVEF
3.5. Morphological Changes of A. johnsonii
3.6. Gene Expression Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calvo, T.; Alvarez-Ordonez, A.; Prieto, M.; Gonzalez-Raurich, M.; Lopez, M. Influence of processing parameters and stress adaptation on the inactivation of Listeria monocytogenes by Non-Thermal Atmospheric Plasma (NTAP). Food Res. Int. 2016, 89, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Feng, X.; Sui, X.; Lin, H.; Han, Y. Inactivation mechanism of Vibrio parahaemolyticus via supercritical carbon dioxide treatment. Food Res. Int. 2017, 100, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.H.; Baek, K.H.; Heo, Y.S.; Yong, H.I.; Jo, C. Synergistic bactericidal effect of clove oil and encapsulated atmospheric pressure plasma against Escherichia coli O157:H7 and Staphylococcus aureus and its mechanism of action. Food Microbiol. 2021, 93, 103611. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.W.; Loke, X.J.; Chang, C.K.; Ko, W.C.; Hou, C.Y.; Hsieh, C.W. Use of the plasma-treated and chitosan/gallic acid-coated polyethylene film for the preservation of tilapia (Orechromis niloticus) fillets. Food Chem. 2020, 329, 126989. [Google Scholar] [CrossRef] [PubMed]
- Ahmad Shiekh, K.; Benjakul, S. Melanosis and quality changes during refrigerated storage of Pacific white shrimp treated with Chamuang (Garcinia cowa Roxb.) leaf extract with the aid of pulsed electric field. Food Chem. 2020, 309, 125516. [Google Scholar] [CrossRef] [PubMed]
- Bhattacherjee, A.; Hrynets, Y.; Betti, M. Improved bactericidal capacity of UV-B radiation against E. coli strains by photosensitizing bacteria with fructosazine—An advanced Maillard reaction product. Food Chem. 2019, 271, 354–361. [Google Scholar] [CrossRef]
- Qi, M.; Zhao, R.; Liu, Q.; Yan, H.; Zhang, Y.; Wang, S.; Yuan, Y. Antibacterial activity and mechanism of high voltage electrostatic field (HVEF) against Staphylococcus aureus in medium plates and food systems. Food Control 2021, 120, 107566. [Google Scholar] [CrossRef]
- Pan, Y.; Cheng, J.-H.; Lv, X.; Sun, D.-W. Assessing the inactivation efficiency of Ar/O2 plasma treatment against Listeria monocytogenes cells: Sublethal injury and inactivation kinetics. LWT 2019, 111, 318–327. [Google Scholar] [CrossRef]
- Gök, V.; Aktop, S.; Özkan, M.; Tomar, O. The effects of atmospheric cold plasma on inactivation of Listeria monocytogenes and Staphylococcus aureus and some quality characteristics of pastırma—A dry-cured beef product. Innov. Food Sci. Emerg. Technol. 2019, 56, 102188. [Google Scholar] [CrossRef]
- Xu, Z.; Cheng, C.; Shen, J.; Lan, Y.; Hu, S.; Han, W.; Chu, P.K. In vitro antimicrobial effects and mechanisms of direct current air-liquid discharge plasma on planktonic Staphylococcus aureus and Escherichia coli in liquids. Bioelectrochemistry 2018, 121, 125–134. [Google Scholar] [CrossRef]
- Hertwig, C.; Leslie, A.; Meneses, N.; Reineke, K.; Rauh, C.; Schlüter, O. Inactivation of Salmonella Enteritidis PT30 on the surface of unpeeled almonds by cold plasma. Innov. Food Sci. Emerg. Technol. 2017, 44, 242–248. [Google Scholar] [CrossRef]
- Lin, L.; Wang, X.; Cui, H. Synergistic efficacy of pulsed magnetic fields and Litseacubeba essential oil treatment against Escherichia coli O157:H7 in vegetable juices. Food Control 2019, 106, 106686. [Google Scholar] [CrossRef]
- Dalvi-Isfahan, M.; Hamdami, N.; Le-Bail, A.; Xanthakis, E. The principles of high voltage electric field and its application in food processing: A review. Food Res. Int. 2016, 89, 48–62. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Jia, G.; Tatsumi, E.; Liu, H. Effect of corona wind, current, electric field and energy consumption on the reduction of the thawing time during the high-voltage electrostatic-field (HVEF) treatment process. Innov. Food Sci. Emerg. Technol. 2016, 34, 135–140. [Google Scholar] [CrossRef]
- He, X.; Liu, R.; Tatsumi, E.; Nirasawa, S.; Liu, H. Factors affecting the thawing characteristics and energy consumption of frozen pork tenderloin meat using high-voltage electrostatic field. Innov. Food Sci. Emerg. Technol. 2014, 22, 110–115. [Google Scholar] [CrossRef]
- Nishimura, Y.; Ino, T.; Iizuka, H. Acinetobacter radioresistens sp. nov. isolated from cotton and soil. Int. J. Syst. Bacteriol. 1988, 38, 209–211. [Google Scholar] [CrossRef] [Green Version]
- Maboni, G.; Seguel, M.; Lorton, A.; Sanchez, S. Antimicrobial resistance patterns of Acinetobacter spp. of animal origin reveal high rate of multidrug resistance. Vet. Microbiol. 2020, 245, 108702. [Google Scholar] [CrossRef]
- Wang, T.; Costa, V.; Jenkins, S.G.; Hartman, B.J.; Westblade, L.F. Acinetobacter radioresistens infection with bacteremia and pneumonia. IDCases 2019, 15, e00495. [Google Scholar] [CrossRef]
- Gupta, K.K.; Mandal, R.; Jagtap, S.; Ramadas, K. Glucovanillin: A potent inhibitor of lipase from Acinetobacter radioresistens. Inform. Med. Unlocked 2018, 10, 126–133. [Google Scholar] [CrossRef]
- Cao, J.; Wang, Q.; Ma, T.; Bao, K.; Yu, X.; Duan, Z.; Shen, X.; Li, C. Effect of EGCG-gelatin biofilm on the quality and microbial composition of tilapia fillets during chilled storage. Food Chem. 2020, 305, 125454. [Google Scholar] [CrossRef]
- Zhuang, S.; Li, Y.; Hong, H.; Liu, Y.; Shu, R.; Luo, Y. Effects of ethyl lauroyl arginate hydrochloride on microbiota, quality and biochemical changes of container-cultured largemouth bass (Micropterus salmonides) fillets during storage at 4 degrees C. Food Chem. 2020, 324, 126886. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Sun, W.; Xiong, G.; Shi, L.; Jiao, C.; Wu, W.; Li, X.; Qiao, Y.; Liao, L.; Ding, A.; et al. Effects of HVEF treatment on microbial communities and physicochemical properties of catfish fillets during chilled storage. LWT 2020, 131, 109667. [Google Scholar] [CrossRef]
- Huang, H.; Xiong, G.; Shi, L.; Wu, W.; Li, X.; Qiao, Y.; Liao, L.; Ding, A.; Wang, L. Application of HVEF treatment in bacteriostasis against Acinetobacter radioresistens. Food Control 2021, 124, 107914. [Google Scholar] [CrossRef]
- Bouyahya, A.; Abrini, J.; Dakka, N.; Bakri, Y. Essential oils of Origanum compactum increase membrane permeability, disturb cell membrane integrity, and suppress quorum-sensing phenotype in bacteria. J. Pharm. Anal. 2019, 9, 301–311. [Google Scholar] [CrossRef]
- Zhao, J.; Peng, T.; Liang, S.; Ma, M.; Zeng, Z.; Yu, P.; Gong, D.; Deng, S. Antibacterial activity and action mechanism of microencapsulated dodecyl gallate with methyl-β-cyclodextrin. Food Control 2020, 109, 106953. [Google Scholar] [CrossRef]
- Tang, S.; Liu, H.; Yin, H.; Liu, X.; Peng, H.; Lu, G.; Dang, Z.; He, C. Effect of 2, 2′, 4, 4′-tetrabromodiphenyl ether (BDE-47) and its metabolites on cell viability, oxidative stress, and apoptosis of HepG2. Chemosphere 2018, 193, 978–988. [Google Scholar] [CrossRef]
- Sun, X.-h.; Hao, L.-r.; Xie, Q.-c.; Lan, W.-q.; Zhao, Y.; Pan, Y.-J.; Wu, V.C.H. Antimicrobial effects and membrane damage mechanism of blueberry (Vaccinium corymbosum L.) extract against Vibrio parahaemolyticus. Food Control 2020, 111, 107020. [Google Scholar] [CrossRef]
- Roh, S.H.; Oh, Y.J.; Lee, S.Y.; Kang, J.H.; Min, S.C. Inactivation of Escherichia coli O157:H7, Salmonella, Listeria monocytogenes, and Tulane virus in processed chicken breast via atmospheric in-package cold plasma treatment. LWT 2020, 127, 109429. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, Y.; Cheng, J.-H.; Sun, D.-W. Inactivation of Listeria Monocytogenes at various growth temperatures by ultrasound pretreatment and cold plasma. LWT 2020, 118, 108635. [Google Scholar] [CrossRef]
- Govaert, M.; Smet, C.; Vergauwen, L.; Ećimović, B.; Walsh, J.L.; Baka, M.; Van Impe, J. Influence of plasma characteristics on the efficacy of Cold Atmospheric Plasma (CAP) for inactivation of Listeria monocytogenes and Salmonella Typhimurium biofilms. Innov. Food Sci. Emerg. Technol. 2019, 52, 376–386. [Google Scholar] [CrossRef]
- Debao, N.; Wang, Q.-Y.; Ren, E.-F.; Zeng, X.-A.; Wang, L.-H.; He, T.-F.; Wen, Q.-H.; Brennan, C.S. Multi-target antibacterial mechanism of eugenol and its combined inactivation with pulsed electric fields in a hurdle strategy on Escherichia coli. Food Control 2019, 106, 106742. [Google Scholar] [CrossRef]
- Korachi, M.; Gurol, C.; Aslan, N. Atmospheric plasma discharge sterilization effects on whole cell fatty acid profiles of Escherichia coli and Staphylococcus aureus. J. Electrost. 2010, 68, 508–512. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, C.; Cui, H.; Lin, L. Antimicrobial mechanism of pulsed light for the control of Escherichia coli O157:H7 and its application in carrot juice. Food Control 2019, 106, 106751. [Google Scholar] [CrossRef]
- Hans, S.; Purkait, D.; Nandan, S.; Bansal, M.; Hameed, S.; Fatima, Z. Rec A disruption unveils cross talk between DNA repair and membrane damage, efflux pump activity, biofilm formation in Mycobacterium smegmatis. Microb. Pathog. 2020, 149, 104262. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, Q.; Yu, G.; Li, L.; Zhao, X.; Huang, X.; Mei, W. Polypyridyl Ruthenium(II) complex-induced mitochondrial membrane potential dissipation activates DNA damage-mediated apoptosis to inhibit liver cancer. Eur. J. Med. Chem. 2019, 164, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Yan, A.; Yang, K.; Wang, Z.; Li, X.; Jia, Y. Antibacterial activity of phenyllactic acid against Listeria monocytogenes and Escherichia coli by dual mechanisms. Food Chem. 2017, 228, 533–540. [Google Scholar] [CrossRef]
- Nobels, I.; Dardenne, F.; De Coen, W.; Blust, R. Application of a multiple endpoint bacterial reporter assay to evaluate toxicological relevant endpoints of perfluorinated compounds with different functional groups and varying chain length. Toxicol. Vitr. 2010, 24, 1768–1774. [Google Scholar] [CrossRef]
- Chen, L.; Xie, Q.; Nathan, C. Alkyl Hydroperoxide Reductase Subunit C (AhpC) Protects Bacterial and Human Cells against Reactive Nitrogen Intermediates. Mol. Cell 1998, 1, 795–805. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Class | Description | C | T |
|---|---|---|---|
| Group Ⅰ Oxidative Stress Response | AhpC | 33.22 ± 19.22 b | 198.62 ± 141.87 a |
| KatE | 72.40 ± 18.57 a | 82.68 ± 13.56 a | |
| Nfo | / | / | |
| SodA | 20.81 ± 4.90 b | 1363.17 ± 167.52 a | |
| Group Ⅱ DNA Damage Protection/Repair | RecG | 100.28 ± 29.61 b | 702.27 ± 151.29 a |
| RadA | 16.25 ± 1.79 b | 73.10 ± 7.64 a | |
| RecN | 143.55 ± 33.46 b | 269.88 ± 127.55 a | |
| Dps | 12.59 ± 4.18 b | 494.88 ± 159.48 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.; Gao, T.; Qian, X.; Wu, W.; Fan, X.; Shi, L.; Xiong, G.; Ding, A.; Li, X.; Qiao, Y.; et al. In Vitro Antibacterial Mechanism of High-Voltage Electrostatic Field against Acinetobacter johnsonii. Foods 2022, 11, 955. https://doi.org/10.3390/foods11070955
Huang H, Gao T, Qian X, Wu W, Fan X, Shi L, Xiong G, Ding A, Li X, Qiao Y, et al. In Vitro Antibacterial Mechanism of High-Voltage Electrostatic Field against Acinetobacter johnsonii. Foods. 2022; 11(7):955. https://doi.org/10.3390/foods11070955
Chicago/Turabian StyleHuang, Han, Tianqi Gao, Xiaoqing Qian, Wenjing Wu, Xiuzhi Fan, Liu Shi, Guangquan Xiong, Anzi Ding, Xin Li, Yu Qiao, and et al. 2022. "In Vitro Antibacterial Mechanism of High-Voltage Electrostatic Field against Acinetobacter johnsonii" Foods 11, no. 7: 955. https://doi.org/10.3390/foods11070955
APA StyleHuang, H., Gao, T., Qian, X., Wu, W., Fan, X., Shi, L., Xiong, G., Ding, A., Li, X., Qiao, Y., Liao, L., & Wang, L. (2022). In Vitro Antibacterial Mechanism of High-Voltage Electrostatic Field against Acinetobacter johnsonii. Foods, 11(7), 955. https://doi.org/10.3390/foods11070955