
Characterization and Evaluation of Antioxidant and Anti-Inflammatory Activities of Flavonoids from the Fruits of Lycium barbarum


 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Extraction and Purification of LBFs
2.3. LC–MS Analysis of LBFs
2.4. Assay of Antioxidant Capacity
2.4.1. Assay for Scavenging Activity against DPPH Free Radicals
2.4.2. Assay of Scavenging Activity on Hydroxyl Radicals
2.4.3. Assay of Scavenging Activity on Superoxide Anion Radicals
2.4.4. Assay for Scavenging Activity on ABTS Radicals
2.4.5. Assay of Total Antioxidant Capacity
2.4.6. Assay for Ferric Reducing Antioxidant Potential (FRAP)
2.5. Assay of Anti-Inflammatory Activity
2.5.1. Cell Culture and Assay of Cell Viability
2.5.2. Assay of NO Free Radicals
2.5.3. Determination of Inflammatory Cytokines
2.5.4. RNA Extraction and qRT-PCR Analysis
2.6. Statistical Analysis
3. Results and Discussion
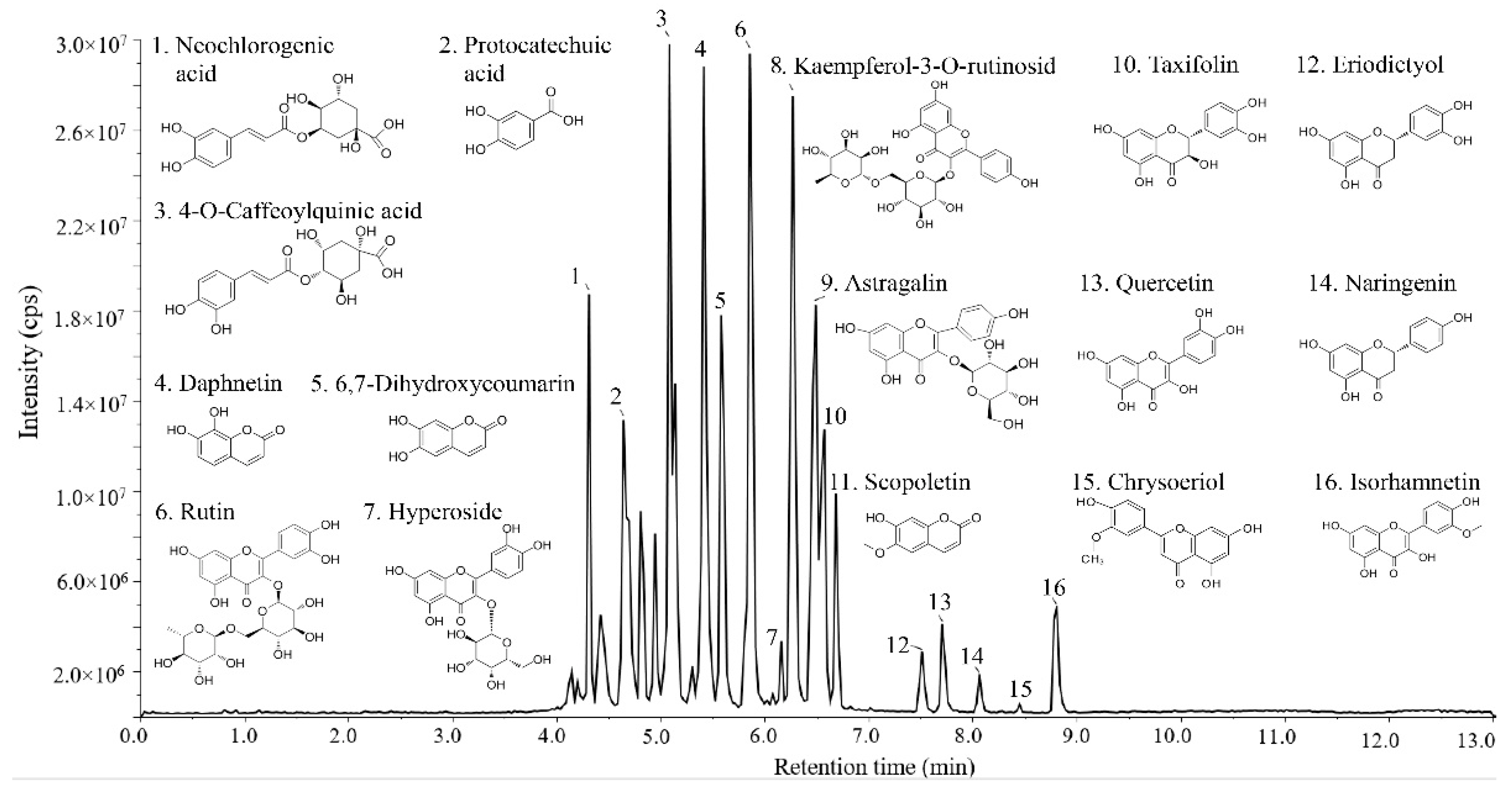
3.1. Identification of Flavonoids by LC–MS
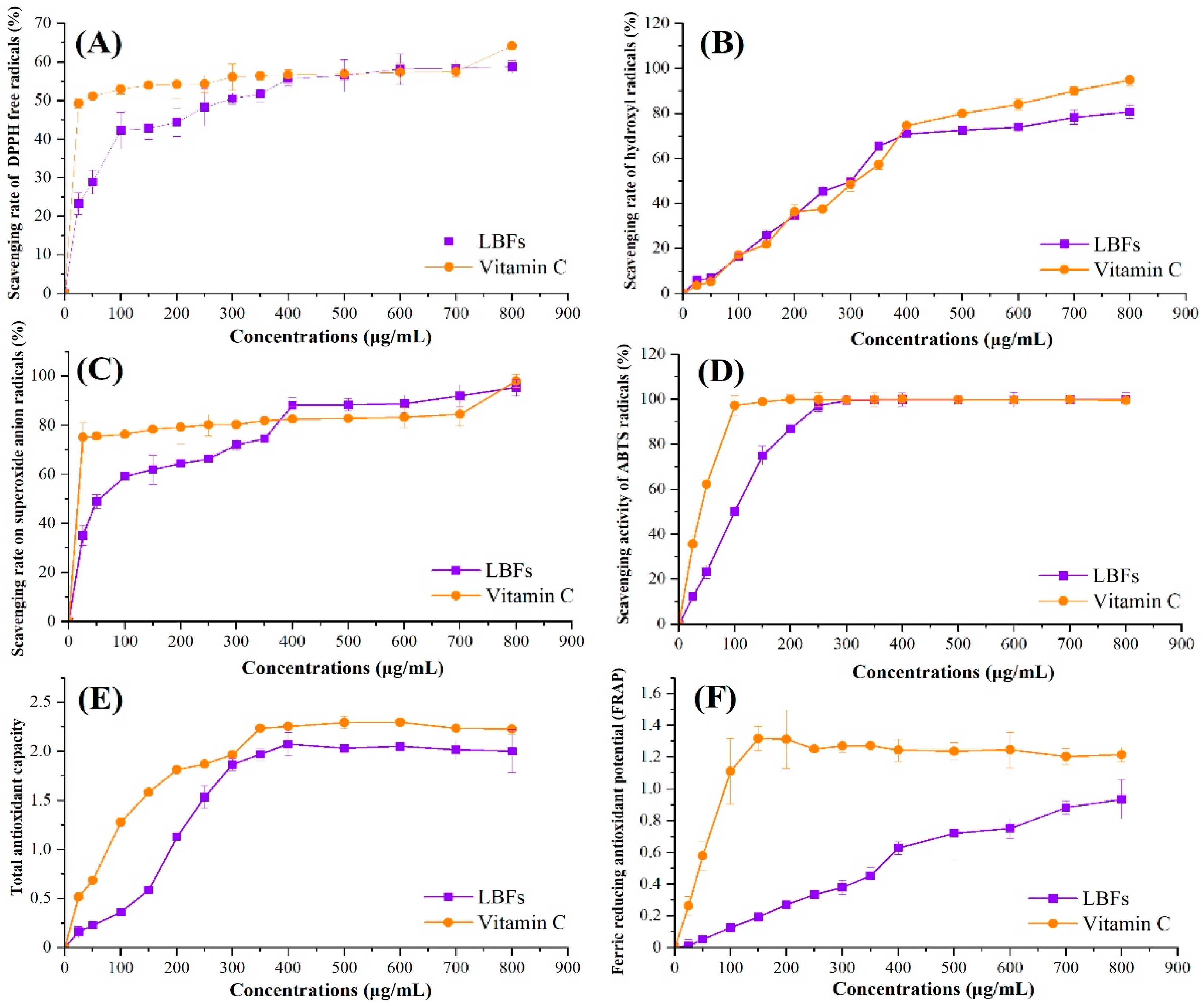
3.2. Antioxidant Activities of LBFs
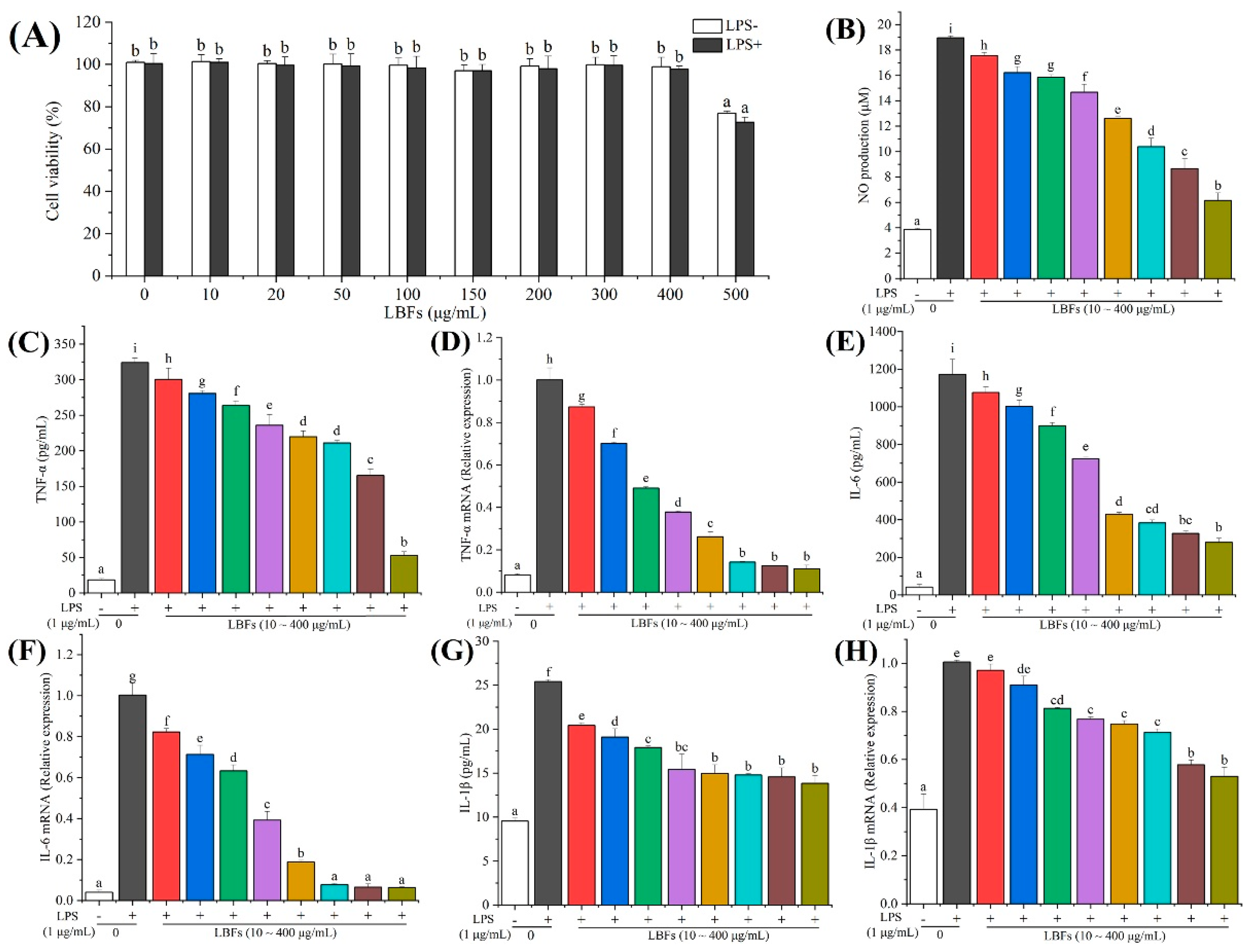
3.3. Anti-Inflammatory Activity of LBFs
3.3.1. Effects of LBFs on Cell Viability
3.3.2. Effects of LBFs on the Production of NO
3.3.3. Effects of LBFs on the Production of LPS-Induced Pro-Inflammatory Cytokines and mRNA Expression
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Magiera, S.; Zaręba, M. Chromatographic determination of phenolic acids and flavonoids in Lycium barbarum L. and evaluation of antioxidant activity. Food Anal. Methods 2015, 8, 2665–2674. [Google Scholar] [CrossRef] [Green Version]
- Cushnie, T.P.T.; Lamb, A.J. Recent advances in understanding the antibacterial properties of flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107. [Google Scholar] [CrossRef]
- Chen, G.; Fan, M.; Wu, J.; Li, N.; Guo, M. Antioxidant and anti-inflammatory properties of flavonoids from lotus plumule. Food Chem. 2019, 277, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wang, C.; Mai, C.; Chen, J.; Lai, X.; He, L.; Huang, S.; Zhang, X. Flavonoids from Livistona chinensis fruit ameliorates LPS/D-GalN-induced acute liver injury by inhibiting oxidative stress and inflammation. J. Funct. Foods 2019, 61, 103460. [Google Scholar] [CrossRef]
- Baba, S.A.; Malik, S.A. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Sci. 2015, 9, 449–454. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Dong, C.; Nie, S.; Li, F.; Wang, Z.; Shen, M.; Xie, M. Extraction, chemical composition and antioxidant activity of flavonoids from Cyclocarya paliurus (Batal.) Iljinskaja leaves. Food Chem. 2015, 186, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.W.; Stratton, M.S.; Ferguson, B.S. Dietary natural products as epigenetic modifiers in aging-associated inflammation and disease. Nat. Prod. Rep. 2020, 37, 653–676. [Google Scholar] [CrossRef]
- Bisson, J.; Simmler, C.; Graham, J.; McAlpine, J.B.; Friesen, B.J.; Lankin, D.C.; Chen, S.N.; Orjala, J.; Pauli, G.F. A new dedicated center to connect scientists and advance natural product research. Planta Med. 2016, 82, P1029. [Google Scholar] [CrossRef]
- Gao, Y.; Wei, Y.; Wang, Y.; Gao, F.; Chen, Z. Lycium Barbarum: A traditional Chinese herb and a promising anti-aging agent. Aging Dis. 2017, 8, 778. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Guo, S.; Zhang, F.; Yan, H.; Qian, D.; Wang, H.; Jin, L.; Duan, J. Comparison of functional components and antioxidant activity of Lycium barbarum L. fruits from different regions in China. Molecules 2019, 24, 2228. [Google Scholar] [CrossRef] [Green Version]
- Mocan, A.; Cairone, F.; Locatelli, M.; Cacciagrano, F.; Carradori, S.; Vodnar, D.C.; Crișan, G.; Simonetti, G.; Cesa, S. Polyphenols from Lycium barbarum (Goji) fruit European cultivars at different maturation steps: Extraction, HPLC-DAD analyses, and biological evaluation. Antioxidants 2019, 8, 562. [Google Scholar] [CrossRef] [Green Version]
- Kan, X.; Yan, Y.; Ran, L.; Lu, L.; Mi, J.; Zhang, Z.; Li, X.; Zeng, X.; Cao, Y. Ultrasonic-assisted extraction and high-speed counter-current chromatography purification of zeaxanthin dipalmitate from the fruits of Lycium barbarum L. Food Chem. 2020, 310, 125854. [Google Scholar] [CrossRef]
- Amagase, H.; Farnsworth, N.R. A review of botanical characteristics, phytochemistry, clinical relevance in efficacy and safety of Lycium barbarum fruit (Goji). Food Res. Int. 2011, 44, 1702–1717. [Google Scholar] [CrossRef]
- Liu, J.; Meng, J.; Du, J.; Liu, X.; Pu, Q.; Di, D.; Chen, C. Preparative separation of flavonoids from Goji berries by mixed-mode macroporous adsorption resins and effect on Aβ-expressing and anti-aging genes. Molecules 2020, 25, 3511. [Google Scholar] [CrossRef]
- Zhou, Z.; Xiao, J.; Fan, H.; Yu, Y.; He, R.; Feng, X.; Kurihara, H.; So, K.; Yao, X.; Gao, H. Polyphenols from wolfberry and their bioactivities. Food Chem. 2017, 214, 644–654. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Gong, G.; Li, F.; Ren, H.; Liu, Y. Adsorption and desorption properties of macroporous resins for flavonoids from the extract of Chinese wolfberry (Lycium barbarum L.). Food Bioprod. Processing 2015, 93, 148–155. [Google Scholar] [CrossRef]
- Chen, S.; Li, X.; Liu, X.; Wang, N.; An, Q.; Ye, X.M.; Zhao, Z.T.; Zhao, M.; Han, Y.; Ouyang, K.H.; et al. Investigation of chemical composition, antioxidant activity, and the effects of Alfalfa flavonoids on growth performance. Oxid. Med. Cell. Longev. 2020, 2020, 8569237. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Shareef, M.M.; Shetty, K.T. Effect of vanadate on different forms of Coomassie brilliant blue and protein assay. Anal. Biochem. 1998, 258, 143–146. [Google Scholar] [CrossRef]
- Škerget, M.; Kotnik, P.; Hadolin, M.; Hraš, A.R.; Simonič, M.; Knez, Ž. Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities. Food Chem. 2005, 89, 191–198. [Google Scholar] [CrossRef]
- Song, Y.; Fu, Y.; Huang, S.; Liao, L.; Wu, Q.; Wang, Y.; Ge, F.; Fang, B. Identification and antioxidant activity of bovine bone collagen-derived novel peptides prepared by recombinant collagenase from Bacillus cereus. Food Chem. 2021, 349, 129143. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Zhou, Z.; Shui, G.; Lam, S.M. Extensive profiling of polyphenols from two trollius species using a combination of untargeted and targeted approaches. Metabolites 2020, 10, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moo-Huchin, V.M.; Moo-Huchin, M.I.; Estrada-León, R.J.; Cuevas-Glory, L.; Estrada-Mota, I.A.; Ortiz-Vázquez, E.; Betancur-Ancona, D.; Sauri-Duch, E. Antioxidant compounds, antioxidant activity and phenolic content in peel from three tropical fruits from Yucatan, Mexico. Food Chem. 2015, 166, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Li, S. In vitro antioxidant analysis and characterization of antler velvet extract. Food Chem. 2009, 114, 1321–1327. [Google Scholar] [CrossRef]
- Xie, M.; Hu, B.; Wang, Y.; Zeng, X. Grafting of gallic acid onto Chitosan enhances antioxidant activities and alters rheological properties of the copolymer. J. Agric. Food Chem. 2014, 62, 9128–9136. [Google Scholar] [CrossRef]
- Liu, L.; Sun, Y.; Laura, T.; Liang, X.; Ye, H.; Zeng, X. Determination of polyphenolic content and antioxidant activity of kudingcha made from Ilex kudingcha C.J. Tseng. Food Chem. 2009, 112, 35–41. [Google Scholar] [CrossRef]
- Tang, J.; Yan, Y.; Ran, L.; Mi, J.; Sun, Y.; Lu, L.; Gao, Y.; Zeng, X.; Cao, Y. Isolation, antioxidant property and protective effect on PC12 cell of the main anthocyanin in fruit of Lycium ruthenicum Murray. J. Funct. Foods 2017, 30, 97–107. [Google Scholar] [CrossRef]
- Fang, Z.; Zhang, Y.; Lü, Y.; Ma, G.; Chen, J.; Liu, D.; Ye, X. Phenolic compounds and antioxidant capacities of bayberry juices. Food Chem. 2009, 113, 884–888. [Google Scholar] [CrossRef]
- Wan, P.; Xie, M.; Chen, G.; Dai, Z.; Hu, B.; Zeng, X.; Sun, Y. Anti-inflammatory effects of dicaffeoylquinic acids from Ilex kudingcha on lipopolysaccharide-treated RAW264.7 macrophages and potential mechanisms. Food Chem. Toxicol. 2019, 126, 332–342. [Google Scholar] [CrossRef]
- Kim, M.; Choi, S.; Lee, P.; Hur, J. Neochlorogenic acid inhibits lipopolysaccharide-induced activation and pro-inflammatory responses in BV2 microglial cells. Neurochem. Res. 2015, 40, 1792–1798. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, Y.; Shi, Y. Visualizing the spatial distribution of endogenous molecules in wolfberry fruit at different development stages by matrix-assisted laser desorption/ionization mass spectrometry imaging. Talanta 2021, 234, 122687. [Google Scholar] [CrossRef]
- Ali, M.C.; Chen, J.; Zhang, H.; Li, Z.; Zhao, L.; Qiu, H. Effective extraction of flavonoids from Lycium barbarum L. fruits by deep eutectic solvents-based ultrasound-assisted extraction. Talanta 2019, 203, 16–22. [Google Scholar] [CrossRef]
- Xie, C.; Li, Y.; Gao, J.; Wang, Y. Esculetin regulates the phenotype switching of airway smooth muscle cells. Phytother. Res. 2019, 33, 3008–3015. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, B.; Jia, Z.; Scarlett, C.J.; Sheng, Z. Adsorption/desorption characteristics and enrichment of quercetin, luteolin and apigenin from Flos populi using macroporous resin. Rev. Bras. Farmacog. 2019, 29, 69–76. [Google Scholar] [CrossRef]
- Cid-Ortega, S.; Monroy-Rivera, J.A. Extraction of kaempferol and its glycosides using supercritical fluids from plant sources: A review. Food Technol. Biotechnol. 2018, 56, 480–493. [Google Scholar] [CrossRef]
- Cho, I.; Gong, J.; Kang, M.; Lee, E.; Park, J.H.Y.; Park, S.; Kang, Y. Astragalin inhibits airway eotaxin-1 induction and epithelial apoptosis through modulating oxidative stress-responsive MAPK signaling. BMC Pulm. Med. 2014, 14, 122. [Google Scholar] [CrossRef] [Green Version]
- Angelis, A.; Hubert, J.; Aligiannis, N.; Michalea, R.; Abedini, A.; Nuzillard, J.; Gangloff, S.; Skaltsounis, A.; Renault, J. Bio-guided isolation of methanol-soluble metabolites of common spruce (Picea abies) bark by products and investigation of their dermo-cosmetic properties. Molecules 2016, 21, 1586. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Xiong, Q.; Gan, D.; Jiao, Y.; Liu, J.; Ma, L.; Zeng, X. Antioxidant activity and potential hepatoprotective effect of polysaccharides from Cyclina sinensis. Carbohyd. Polym. 2013, 91, 262–268. [Google Scholar] [CrossRef]
- Hwang, S.J.; Yoon, W.B.; Lee, O.; Cha, S.J.; Kim, J.D. Radical-scavenging-linked antioxidant activities of extracts from black chokeberry and blueberry cultivated in Korea. Food Chem. 2014, 146, 71–77. [Google Scholar] [CrossRef]
- Żebrowska, J.; Dyduch-Siemińska, M.; Gawroński, J.; Jackowska, I.; Pabich, M. Genetic estimates of antioxidant properties in the conventionally and in vitro propagated strawberry (Fragaria × ananassa Duch). Food Chem. 2019, 299, 125110. [Google Scholar] [CrossRef]
- Protti, M.; Gualandi, I.; Mandrioli, R.; Zappoli, S.; Tonelli, D.; Mercolini, L. Analytical profiling of selected antioxidants and total antioxidant capacity of goji (Lycium spp.) berries. J. Pharmaceut. Biomed. 2017, 143, 252–260. [Google Scholar] [CrossRef]
- Fehér, J.; Kovács, I.; Balacco Gabrieli, C. Role of gastrointestinal inflammations in the development and treatment of depression. Orv. Hetil. 2011, 152, 1477–1485. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Shin, J.; Jang, D.S.; Lee, K. Cnidilide, an alkyl phthalide isolated from the roots of Cnidium officinale, suppresses LPS-induced NO, PGE2, IL-1β, IL-6 and TNF-α production by AP-1 and NF-κB inactivation in RAW264.7 macrophages. Int. Immunopharmacol. 2016, 40, 146–155. [Google Scholar] [CrossRef]
- Shi, Q.; Cao, J.; Fang, L.; Zhao, H.; Liu, Z.; Ran, J.; Zheng, X.; Li, X.; Zhou, Y.; Ge, D.; et al. Geniposide suppresses LPS-induced nitric oxide, PGE2 and inflammatory cytokine by downregulating NF-κB, MAPK and AP-1 signaling pathways in macrophages. Int. Immunopharmacol. 2014, 20, 298–306. [Google Scholar] [CrossRef]
- Damlar, I.; Esen, E.; Tatli, U. Effects of glucosamine-chondroitin combination on synovial fluid IL-1β, IL-6, TNF-α and PGE2 levels in internal derangements of temporomandibular joint. Med. Oral Patol. Oral Cir. Bucal 2015, 20, e278–e283. [Google Scholar] [CrossRef]
- Patwardhan, R.S.; Sharma, D.; Thoh, M.; Checker, R.; Sandur, S.K. Baicalein exhibits anti-inflammatory effects via inhibition of NF-κB transactivation. Biochem. Pharmacol. 2016, 108, 75–89. [Google Scholar] [CrossRef]
- Chanput, W.; Krueyos, N.; Ritthiruangdej, P. Anti-oxidative assays as markers for anti-inflammatory activity of flavonoids. Int. Immunopharmacol. 2016, 40, 170–175. [Google Scholar] [CrossRef]
- Lee, K.J.; Oh, Y.C.; Cho, W.K.; Ma, J.Y. Antioxidant and anti-inflammatory activity determination of one hundred kinds of pure chemical compounds using offline and online screening HPLC assay. Evid.-Based Complement. Altern. 2015, 2015, 165457. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence | Annealing T (°C) |
|---|---|---|
| TNF-α | FW: CTCATGCACCACCATCAAGG | 60 |
| RV: ACCTGACCACTCTCCCTTTG | ||
| IL-1β | FW: AGCTTCAAATCTCGCAGCAG | 59 |
| RV: TCTCCACAGCCACAATGAGT | ||
| IL-6 | FW: CTCTGGCGGAGCTATTGAGA | 60 |
| RV: AAGTCTCCTGCGTGGAGAAA | ||
| GAPDH | FW: GGACTTACAGAGGTCCGCTT | 59 |
| RV: CTATAGGGCCTGGGTCAGTG |
| No. | Molecular Formula | Theoretical m/z [M − H] | Retention Time | Component Name |
|---|---|---|---|---|
| 1 | C16H18O9 | 353.309 | 4.31 | Neochlorogenic acid |
| 2 | C7H6O4 | 153.120 | 4.64 | Protocatechuic acid |
| 3 | C16H18O9 | 353.309 | 5.08 | 4-O-Caffeoylquinic acid |
| 4 | C9H6O4 | 177.140 | 5.40 | Daphnetin |
| 5 | C9H6O4 | 177.141 | 5.58 | 6,7-Dihydroxycoumarin |
| 6 | C27H30O16 | 609.518 | 5.86 | Rutin |
| 7 | C21H20O12 | 463.376 | 6.16 | Hyperoside |
| 8 | C27H30O15 | 593.518 | 6.26 | Kaempferol-3-o-rutinosid |
| 9 | C21H20O11 | 447.377 | 6.51 | Astragalin |
| 10 | C15H12O7 | 303.252 | 6.56 | Taxifolin |
| 11 | C10H8O4 | 191.168 | 6.72 | Scopoletin |
| 12 | C15H12O6 | 287.252 | 7.65 | Eriodictyol |
| 13 | C15H10O7 | 301.236 | 7.72 | Quercetin |
| 14 | C15H12O5 | 271.253 | 8.07 | Naringenin |
| 15 | C16H12O6 | 299.263 | 8.36 | Chrysoeriol |
| 16 | C16H12O7 | 315.262 | 8.79 | Isorhamnetin |
| LBFs | VC | |
|---|---|---|
| EC50/DPPH (mg/mL) | 294.864 ± 2.470 b | 40.436 ± 1.607 a |
| EC50/HR (mg/mL) | 275.048 ± 2.439 a | 265.520 ± 2.424 a |
| EC50/SR (mg/mL) | 66.463 ± 1.820 b | 1.919 ± 0.283 a |
| EC50/ABTS (mg/mL) | 82.663 ± 1.917 b | 34.348 ± 1.536 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.; Hu, Y.; Yan, Y.; Zhou, W.; Chen, G.; Zeng, X.; Cao, Y. Characterization and Evaluation of Antioxidant and Anti-Inflammatory Activities of Flavonoids from the Fruits of Lycium barbarum. Foods 2022, 11, 306. https://doi.org/10.3390/foods11030306
Yang T, Hu Y, Yan Y, Zhou W, Chen G, Zeng X, Cao Y. Characterization and Evaluation of Antioxidant and Anti-Inflammatory Activities of Flavonoids from the Fruits of Lycium barbarum. Foods. 2022; 11(3):306. https://doi.org/10.3390/foods11030306
Chicago/Turabian StyleYang, Tingting, Yuhang Hu, Yamei Yan, Wangting Zhou, Guijie Chen, Xiaoxiong Zeng, and Youlong Cao. 2022. "Characterization and Evaluation of Antioxidant and Anti-Inflammatory Activities of Flavonoids from the Fruits of Lycium barbarum" Foods 11, no. 3: 306. https://doi.org/10.3390/foods11030306
APA StyleYang, T., Hu, Y., Yan, Y., Zhou, W., Chen, G., Zeng, X., & Cao, Y. (2022). Characterization and Evaluation of Antioxidant and Anti-Inflammatory Activities of Flavonoids from the Fruits of Lycium barbarum. Foods, 11(3), 306. https://doi.org/10.3390/foods11030306