


Simulated In Vitro Digestive Characteristics of Raw Yam Tubers in Japanese Diet: Changes in Protein Profile, Starch Digestibility, Antioxidant Capacity and Microstructure

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Samples Preparation
2.3. Moisture Content
2.4. Simulated In Vitro Gastrointestinal Digestion
2.5. Starch Hydrolysis
2.6. Total Protein Content and Protein Digestibility
2.7. Soluble Protein Fractions and Distribution
2.8. Antioxidant Capacity
2.9. Total Phenolic Content (TPC)
2.10. Free Amino Acids
2.11. Morphological Observation
2.12. Statistical Analysis
3. Results and Discussion
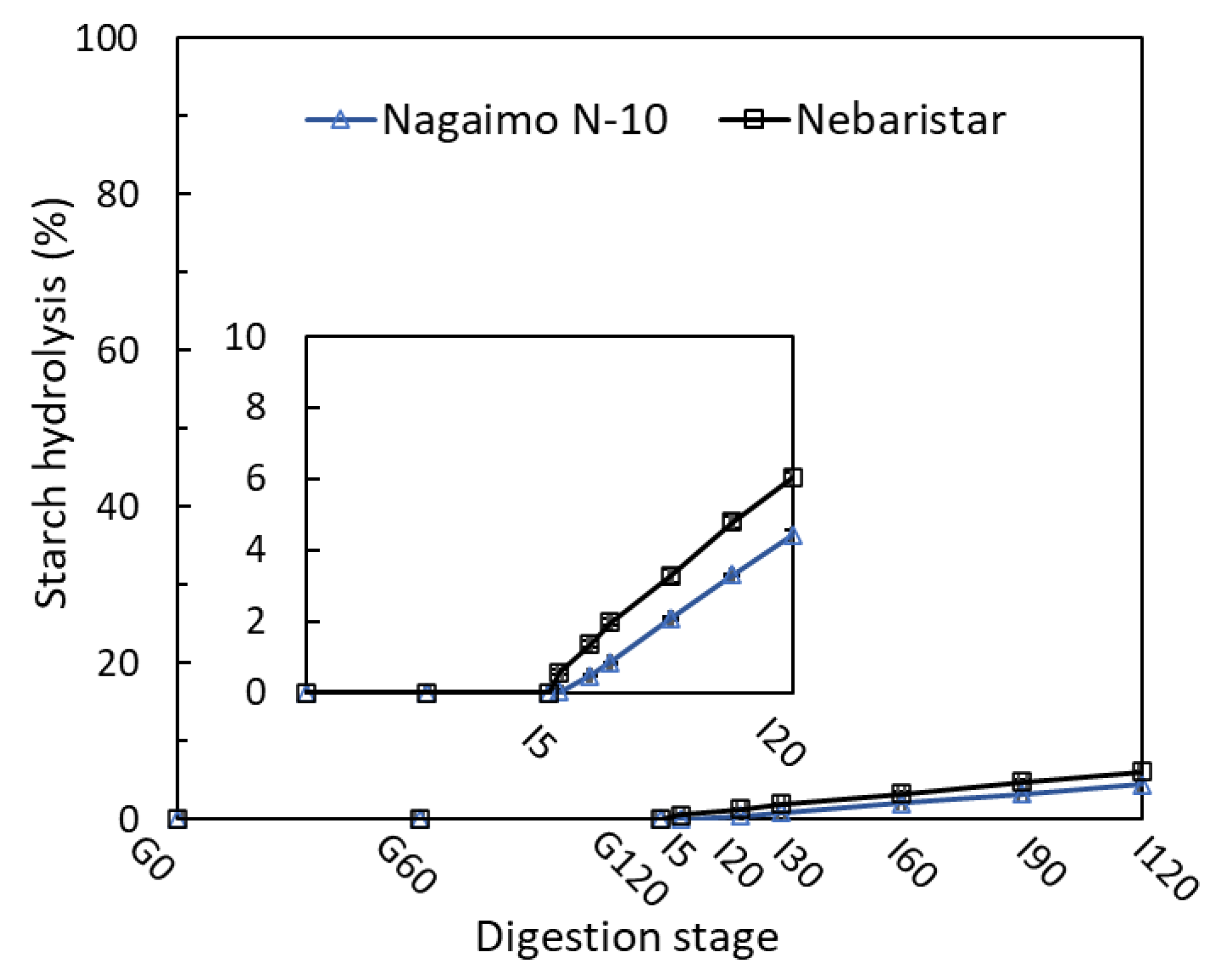
3.1. Starch Digestibility
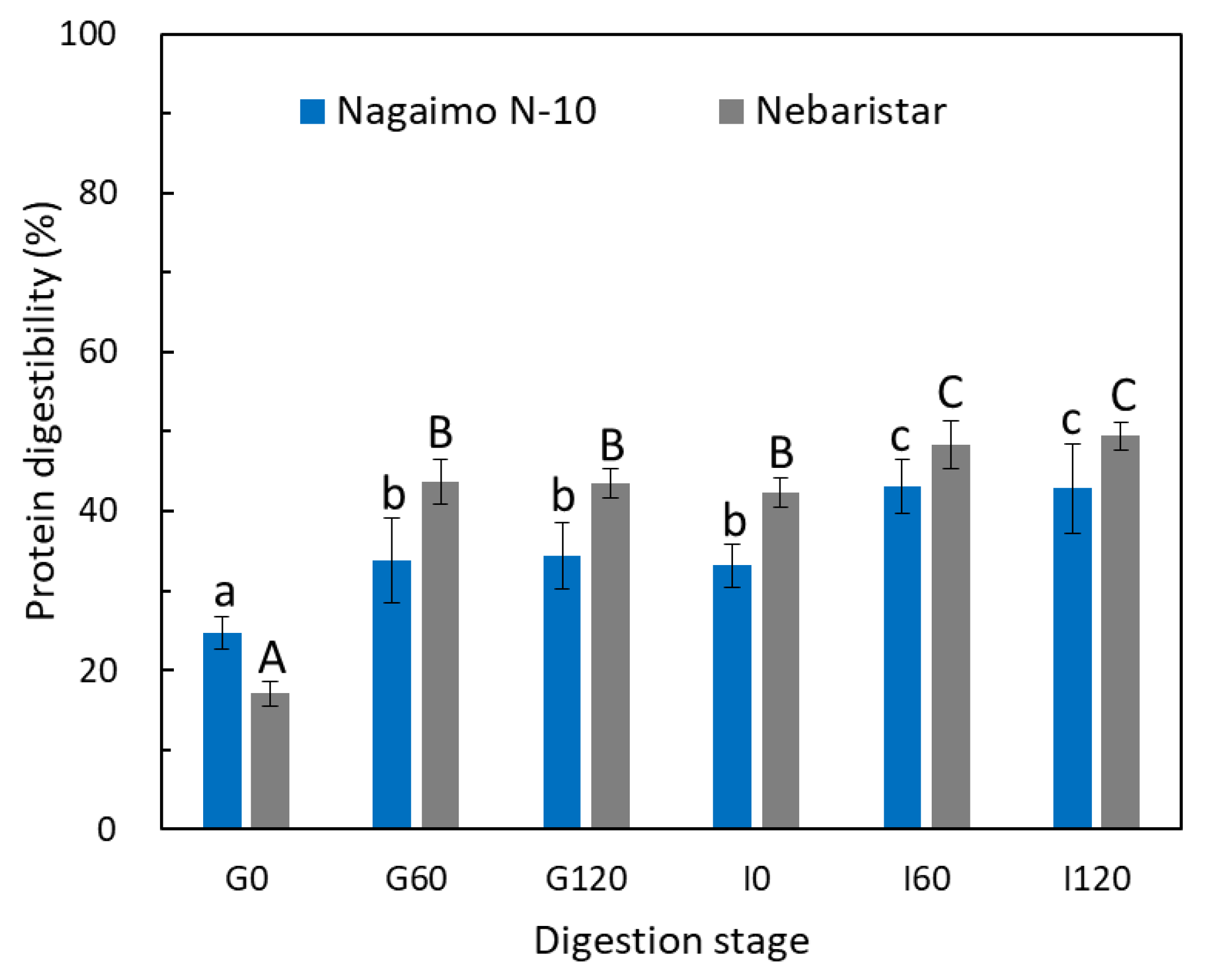
3.2. Protein Digestion
3.2.1. Protein Digestibility
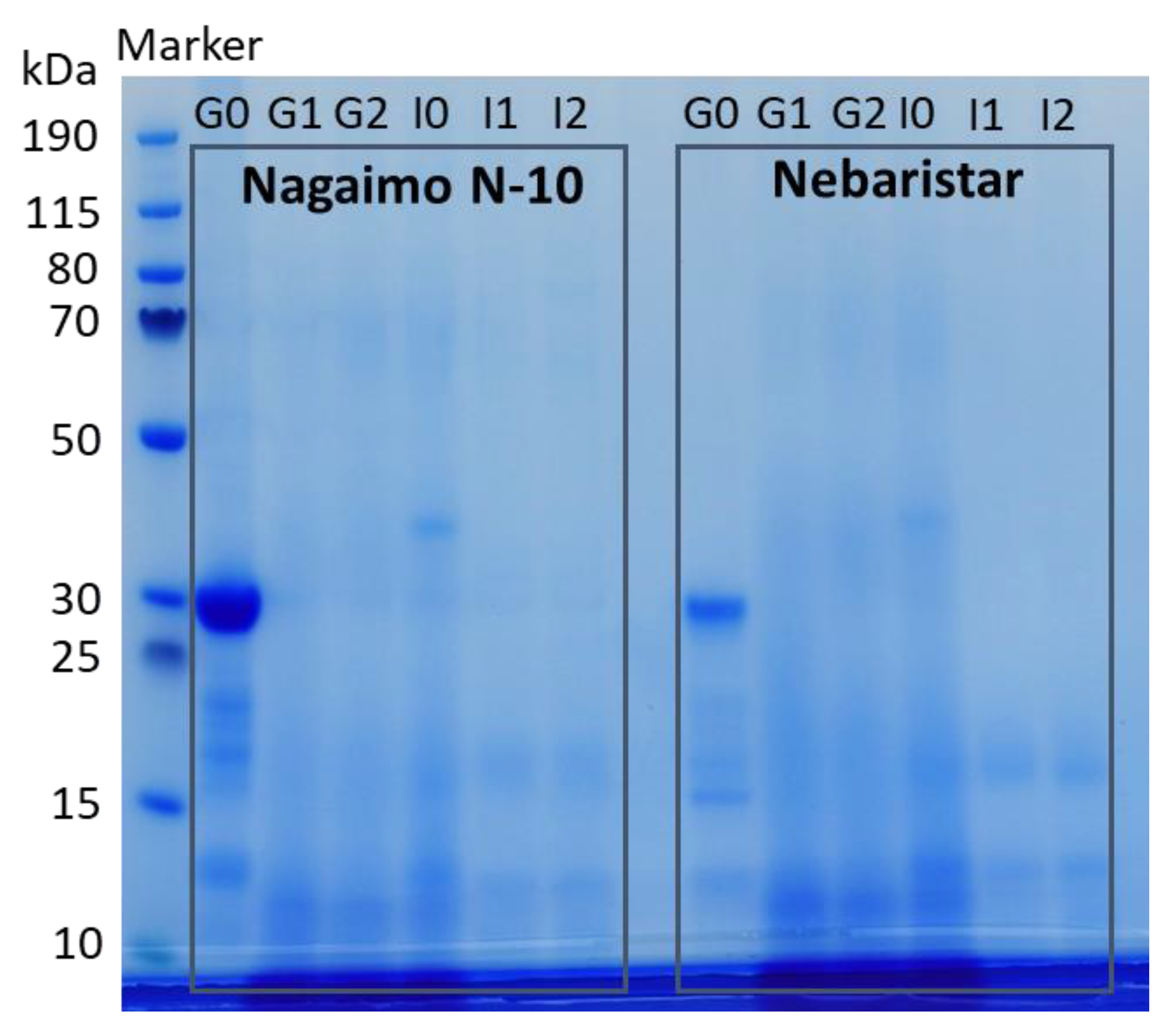
3.2.2. Soluble Protein Fractions and Distribution
3.2.3. Changes in Amino Acid Composition during Simulated In Vitro Digestion
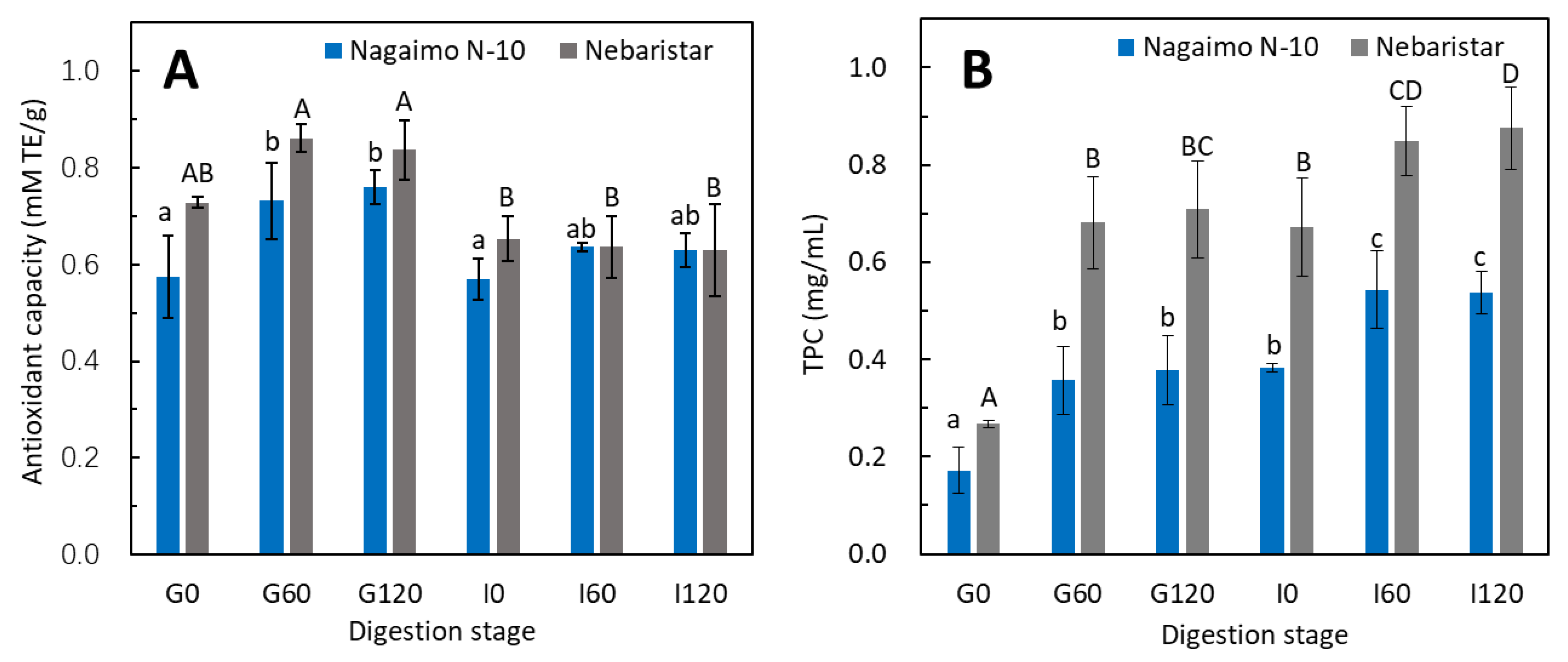
3.3. Changes in Total Antioxidant Capacity and Total Phenolic Content during Simulated In Vitro Digestion
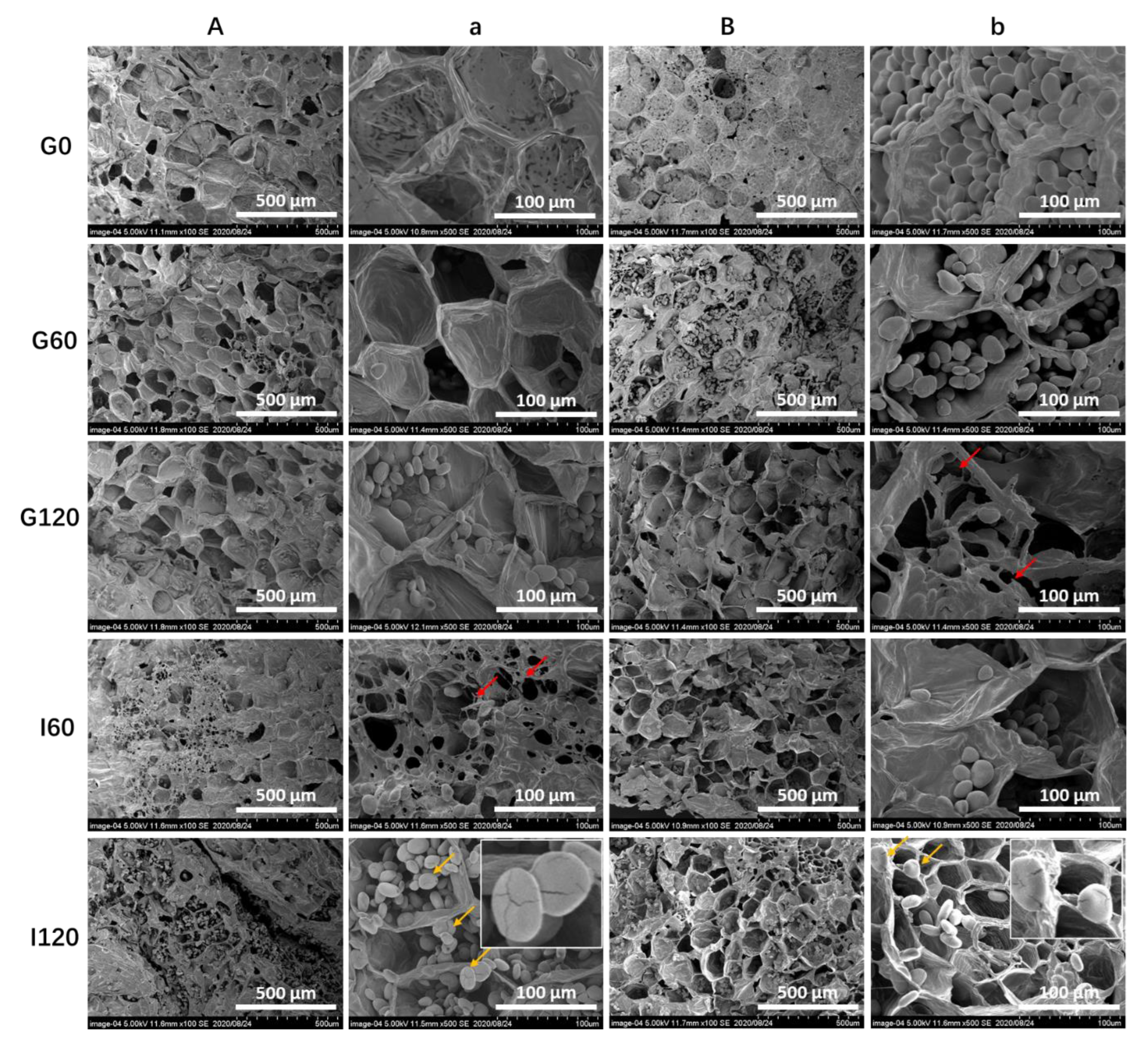
3.4. Changes in Microstructures of Yam Tubers during Simulated In Vitro Digestion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nagaimo N-10 | Nebaristar | |
|---|---|---|
| Moisture content (%) | 82.2 ± 1.2 | 78.0 ± 2.4 |
| Crude protein (dry basis %) | 11.1 ± 0.2 | 11.3 ± 0.3 |
| Total starch (dry basis %) | 67.3 ± 1.6 | 72.4 ± 0.7 |

References
- FAO. FAOSTAT Crops and Livestock Products Data. 2020. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 25 November 2022).
- Epping, J.; Laibach, N. An underutilized orphan tuber crop-Chinese yam: A review. Planta 2020, 252, 58. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Lyons, J.B.; Chilaka, C.A. The Dioscorea Genus (Yam)-An Appraisal of Nutritional and Therapeutic Potentials. Foods 2020, 9, 1304. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Nagashima, T. Functional properties of dioscorin, a soluble viscous protein from Japanese yam (Dioscorea opposita thunb.) tuber mucilage Tororo. Z. Nat. C 2006, 61, 792–798. [Google Scholar] [CrossRef]
- Nagai, T.; Nagashima, T.; Suzuki, N. Purification and Partial Characterization of Major Viscous Protein from Yam (Dioscorea opposite Thunb.) Tuber Mucilagetororo. Int. J. Food Prop. 2007, 10, 515–526. [Google Scholar] [CrossRef]
- Conlan, R.S.; Griffiths, L.-A.; Napier, J.A.; Shewry, P.R.; Mantell, S.; Ainsworth, C. Isolation and characterisation of cDNA clones representing the genes encoding the major tuber storage protein (dioscorin) of yam (Dioscorea cayenensis Lam.). Plant Mol. Biol. 1995, 28, 369–380. [Google Scholar] [CrossRef]
- Silva do Nascimento, E.; Caju de Oliveira, J.M.; Jocelino Gomes de Lacerda, J.T.; Montenegro, S.B.; Caetano-Silva, M.E.; Dias, M.; Alberto de Almeida Gadelha, C. Yam (Dioscorea cayennensis) protein concentrate: Production, characterization and in vitro evaluation of digestibility. LWT 2021, 140, 110771. [Google Scholar] [CrossRef]
- Wu, Z.G.; Jiang, W.; Nitin, M.; Bao, X.Q.; Chen, S.L.; Tao, Z.M. Characterizing diversity based on nutritional and bioactive compositions of yam germplasm (Dioscorea spp.) commonly cultivated in China. J. Food Drug Anal. 2016, 24, 367–375. [Google Scholar] [CrossRef]
- AOAC. Association of Official Analytical Chemists. Official Methods of Analysis; AOAC: Arlington, VA, USA, 1990. [Google Scholar]
- Tamura, M.; Singh, J.; Kaur, L.; Ogawa, Y. Impact of structural characteristics on starch digestibility of cooked rice. Food Chem. 2016, 191, 91–97. [Google Scholar] [CrossRef]
- Zhang, C.; Quek, S.Y.; Fu, N.; Su, Y.; Kilmartin, P.A.; Chen, X.D. Storage stability and in vitro digestion of microencapsulated powder containing fermented noni juice and probiotics. Food Biosci. 2020, 37, 100740. [Google Scholar] [CrossRef]
- Ketnawa, S.; Ogawa, Y. Evaluation of protein digestibility of fermented soybeans and changes in biochemical characteristics of digested fractions. J. Funct. Foods 2019, 52, 640–647. [Google Scholar] [CrossRef]
- Zhang, C.; Khoo, S.L.A.; Swedlund, P.; Ogawa, Y.; Shan, Y.; Quek, S.Y. Fabrication of spray-dried microcapsules containing noni juice using blends of maltodextrin and gum acacia: Physicochemical properties of powders and bioaccessibility of bioactives during In vitro digestion. Foods 2020, 9, 1316. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, X.; Zhang, B.; Chen, P.X.; Liu, R.; Tsao, R. Characterisation of phenolics, betanins and antioxidant activities in seeds of three Chenopodium quinoa Willd. genotypes. Food Chem. 2015, 166, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ada Khoo, S.L.; Chen, X.D.; Quek, S.Y. Microencapsulation of fermented noni juice via micro-fluidic-jet spray drying: Evaluation of powder properties and functionalities. Powder Technol. 2020, 361, 995–1005. [Google Scholar] [CrossRef]
- Guo, P.; Yu, J.; Copeland, L.; Wang, S.; Wang, S. Mechanisms of starch gelatinization during heating of wheat flour and its effect on in vitro starch digestibility. Food Hydrocoll. 2018, 82, 370–378. [Google Scholar] [CrossRef]
- Tian, J.; Chen, S.; Shi, J.; Chen, J.; Liu, D.; Cai, Y.; Ye, X. Microstructure and digestibility of potato strips produced by conventional frying and air-frying: An in vitro study. Food Struct. 2017, 14, 30–35. [Google Scholar] [CrossRef]
- Myoda, T.; Matsuda, Y.; Suzuki, T.; Nakagawa, T.; Nagai, T.; Nagashima, T. Identification of soluble proteins and interaction with mannan in mucilage of Dioscorea opposita Thunb. (Chinese yam tuber). Food Sci. Technol. Res. 2006, 12, 299–302. [Google Scholar] [CrossRef]
- Zou, J.; Xu, M.; Zou, Y.; Yang, B. Physicochemical properties and microstructure of Chinese yam (Dioscorea opposita Thunb.) flour. Food Hydrocoll. 2021, 113, 106448. [Google Scholar] [CrossRef]
- Yu, B.; Li, J.; Tao, H.; Zhao, H.; Liu, P.; Cui, B. Physicochemical properties and in vitro digestibility of hydrothermal treated Chinese yam (Dioscorea opposita Thunb.) starch and flour. Int. J. Biol. Macromol. 2021, 176, 177–185. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Brodkorb, A. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Ajibola, C.F.; Malomo, S.A.; Fagbemi, T.N.; Aluko, R.E. Polypeptide composition and functional properties of African yam bean seed (Sphenostylis stenocarpa) albumin, globulin and protein concentrate. Food Hydrocoll. 2016, 56, 189–200. [Google Scholar] [CrossRef]
- Gong, X.; Hui, X.; Wu, G.; Morton, J.D.; Brennan, M.A.; Brennan, C.S. In Vitro digestion characteristics of cereal protein concentrates as assessed using a pepsin-pancreatin digestion model. Food Res. Int. 2022, 152, 110715. [Google Scholar] [CrossRef]
- Wang, X.; Ye, A.; Dave, A.; Singh, H. In vitro digestion of soymilk using a human gastric simulator: Impact of structural changes on kinetics of release of proteins and lipids. Food Hydrocoll. 2021, 111, 106235. [Google Scholar] [CrossRef]
- Nikoo, M.; Regenstein, J.M.; Noori, F.; Piri Gheshlaghi, S. Autolysis of rainbow trout (Oncorhynchus mykiss) by-products: Enzymatic activities, lipid and protein oxidation, and antioxidant activity of protein hydrolysates. LWT 2021, 140, 110702. [Google Scholar] [CrossRef]
- Möller, N.P.; Scholz-Ahrens, K.E.; Roos, N.; Schrezenmeir, J. Bioactive peptides and proteins from foods: Indication for health effects. Eur. J. Nutr. 2008, 47, 171–182. [Google Scholar] [CrossRef]
- Wu, G. Dietary protein intake and human health. Food Funct. 2016, 7, 1251–1265. [Google Scholar] [CrossRef]
- Wu, G.; Meininger, C.J.; Knabe, D.A.; Baze, F.W.; Rhoads, M.J. Arginine nutrition in development, health and disease. Curr. Opin. Clin. Nutr. Metab. Care 2000, 3, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Diep, T.T.; Yoo, M.J.Y.; Rush, E. Effect of In Vitro Gastrointestinal Digestion on Amino Acids, Polyphenols and Antioxidant Capacity of Tamarillo Yoghurts. Int. J. Mol. Sci. 2022, 23, 2526. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Suwannachot, J.; Ogawa, Y. In Vitro gastrointestinal digestion of crisphead lettuce: Changes in bioactive compounds and antioxidant potential. Food Chem. 2020, 311, 125885. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Xiong, H.; Selomulya, C.; Chen, X.D.; Zhong, H.; Wang, S.; Zhou, Q. Enzymatic hydrolysis of rice dreg protein: Effects of enzyme type on the functional properties and antioxidant activities of recovered proteins. Food Chem. 2012, 134, 1360–1367. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In Vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Jamali, B.; Bjornsdottir, I.; Nordfang, O.; Hansen, S.H. Investigation of racemisation of the enantiomers of glitazone drug compounds at different pH using chiral HPLC and chiral CE. J. Pharm. Biomed. Anal. 2008, 46, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein–phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin–Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Romano, A.; D’Amelia, V.; Gallo, V.; Palomba, S.; Carputo, D.; Masi, P. Relationships between composition, microstructure and cooking performances of six potato varieties. Food Res. Int. 2018, 114, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Mennah-Govela, Y.A.; Bornhorst, G.M. Mass transport processes in orange-fleshed sweet potatoes leading to structural changes during in vitro gastric digestion. J. Food Eng. 2016, 191, 48–57. [Google Scholar] [CrossRef]





| Nagaimo N-10 | Nebaristar | |||||
|---|---|---|---|---|---|---|
| Amino Acid | Before Digestion | After Digestion | Increment (Fold) | Before Digestion | After Digestion | Increment (Fold) |
| nmol/g Fresh Yam | nmol/g Fresh Yam | |||||
| Asp | 465.2 ± 8.1 | 599.9 ± 14.7 | 1.3 | 669.3 ± 4.9 | 802.2 ± 29.3 | 1.2 |
| Thr | 770.7 ± 16.8 | 957.4 ± 20.1 | 1.2 | 1374.7 ± 82.8 | 1443.7 ± 18.9 | 1.1 |
| Ser | 4941.8 ± 92.2 | 5485.8 ± 135.4 | 1.1 | 5881.7 ± 181.8 | 6218.2 ± 78.8 | 1.1 |
| Glu | 420.4 ± 0.6 | 624.7 ± 17.7 | 1.5 | 496.3 ± 15.7 | 608.4 ± 40.6 | 1.2 |
| Gly | 1069.7 ± 14.4 | 1273.4 ± 7.3 | 1.2 | 984.0 ± 10.7 | 1131.5 ± 15.2 | 1.1 |
| Ala | 4797.5 ± 60.2 | 5426.4 ± 68.6 | 1.1 | 3255.0 ± 34.9 | 3647.4 ± 48.0 | 1.1 |
| Val | 753.2 ± 12.6 | 1114.1 ± 11.0 | 1.5 | 824.5 ± 11.6 | 1243.4 ± 17.7 | 1.5 |
| Met | 177.7 ± 0.9 | 341.5 ± 3.1 | 1.9 | 242.5 ± 3.3 | 495.3 ± 20.1 | 2.0 |
| Ile | 473.3 ± 3.7 | 806.5 ± 9.8 | 1.7 | 611.3 ± 7.3 | 1089.2 ± 17.1 | 1.8 |
| Leu | 548.8 ± 4.2 | 1607.8 ± 23.6 | 2.9 | 848.7 ± 10.8 | 2920.7 ± 36.9 | 3.4 |
| Tyr | 115.6 ± 1.2 | 685.6 ± 10.1 | 5.9 | 139.8 ± 1.8 | 1211.5 ± 15.8 | 8.7 |
| Phe | 427.3 ± 3.6 | 1170.4 ± 6.7 | 2.7 | 484.8 ± 5.6 | 1964.9 ± 7.8 | 4.1 |
| His | 253.5 ± 1.1 | 333.9 ± 3.2 | 1.3 | 345.4 ± 6.1 | 438.2 ± 9.9 | 1.3 |
| Lys | 384.1 ± 2.1 | 966.8 ± 13.8 | 2.5 | 755.1 ± 7.5 | 1770.5 ± 16.8 | 2.3 |
| Arg | 3257.0 ± 18.5 | 4253.6 ± 56.6 | 1.3 | 8631.8 ± 118.2 | 10,317.1 ± 131.6 | 1.2 |
| Pro | 102.2 ± 6.8 | 123.7 ± 1.6 | 1.2 | 225.1 ± 2.8 | 241.1 ± 16.3 | 1.1 |
| Cysta * | 109.5 ± 0.0 | 148.8 ± 4.4 | 1.4 | 252.6 ± 3.3 | 315.9 ± 1.3 | 1.3 |
| In total | 19,067.5 ± 247.1 | 25,920.4 ± 407.7 | 1.4 | 26,022.6 ± 509.5 | 35,859.2 ± 522.1 | 1.4 |
| EAA | 3788.7 ± 41.4 | 7298.6 ± 84.7 | 1.9 | 5487.0 ± 129.8 | 11,366.0 ± 137.4 | 2.1 |
| HBAA | 7505.0 ± 99.9 | 11,424.9 ± 139.0 | 1.5 | 6884.3 ± 81.8 | 13,129.5 ± 180.9 | 1.9 |
| HLAA | 4780.3 ± 30.5 | 6778.9 ± 105.9 | 1.4 | 10,897.9 ± 152.3 | 13,936.4 ± 228.3 | 1.3 |
| AAA | 796.5 ± 5.9 | 2190.0 ± 20.1 | 2.7 | 970.0 ± 13.6 | 3614.7 ± 33.6 | 3.7 |
| AOAA | 1185.8 ± 13.5 | 2803.9 ± 29.3 | 2.4 | 1690.2 ± 23.0 | 4667.0 ± 71.3 | 2.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Ketnawa, S.; Thuengtung, S.; Cai, Y.; Qin, W.; Ogawa, Y. Simulated In Vitro Digestive Characteristics of Raw Yam Tubers in Japanese Diet: Changes in Protein Profile, Starch Digestibility, Antioxidant Capacity and Microstructure. Foods 2022, 11, 3892. https://doi.org/10.3390/foods11233892
Zhang C, Ketnawa S, Thuengtung S, Cai Y, Qin W, Ogawa Y. Simulated In Vitro Digestive Characteristics of Raw Yam Tubers in Japanese Diet: Changes in Protein Profile, Starch Digestibility, Antioxidant Capacity and Microstructure. Foods. 2022; 11(23):3892. https://doi.org/10.3390/foods11233892
Chicago/Turabian StyleZhang, Chuang, Sunantha Ketnawa, Sukanya Thuengtung, Yidi Cai, Wei Qin, and Yukiharu Ogawa. 2022. "Simulated In Vitro Digestive Characteristics of Raw Yam Tubers in Japanese Diet: Changes in Protein Profile, Starch Digestibility, Antioxidant Capacity and Microstructure" Foods 11, no. 23: 3892. https://doi.org/10.3390/foods11233892
APA StyleZhang, C., Ketnawa, S., Thuengtung, S., Cai, Y., Qin, W., & Ogawa, Y. (2022). Simulated In Vitro Digestive Characteristics of Raw Yam Tubers in Japanese Diet: Changes in Protein Profile, Starch Digestibility, Antioxidant Capacity and Microstructure. Foods, 11(23), 3892. https://doi.org/10.3390/foods11233892