

Asymptomatic Carriage of Listeria monocytogenes by Animals and Humans and Its Impact on the Food Chain

Abstract
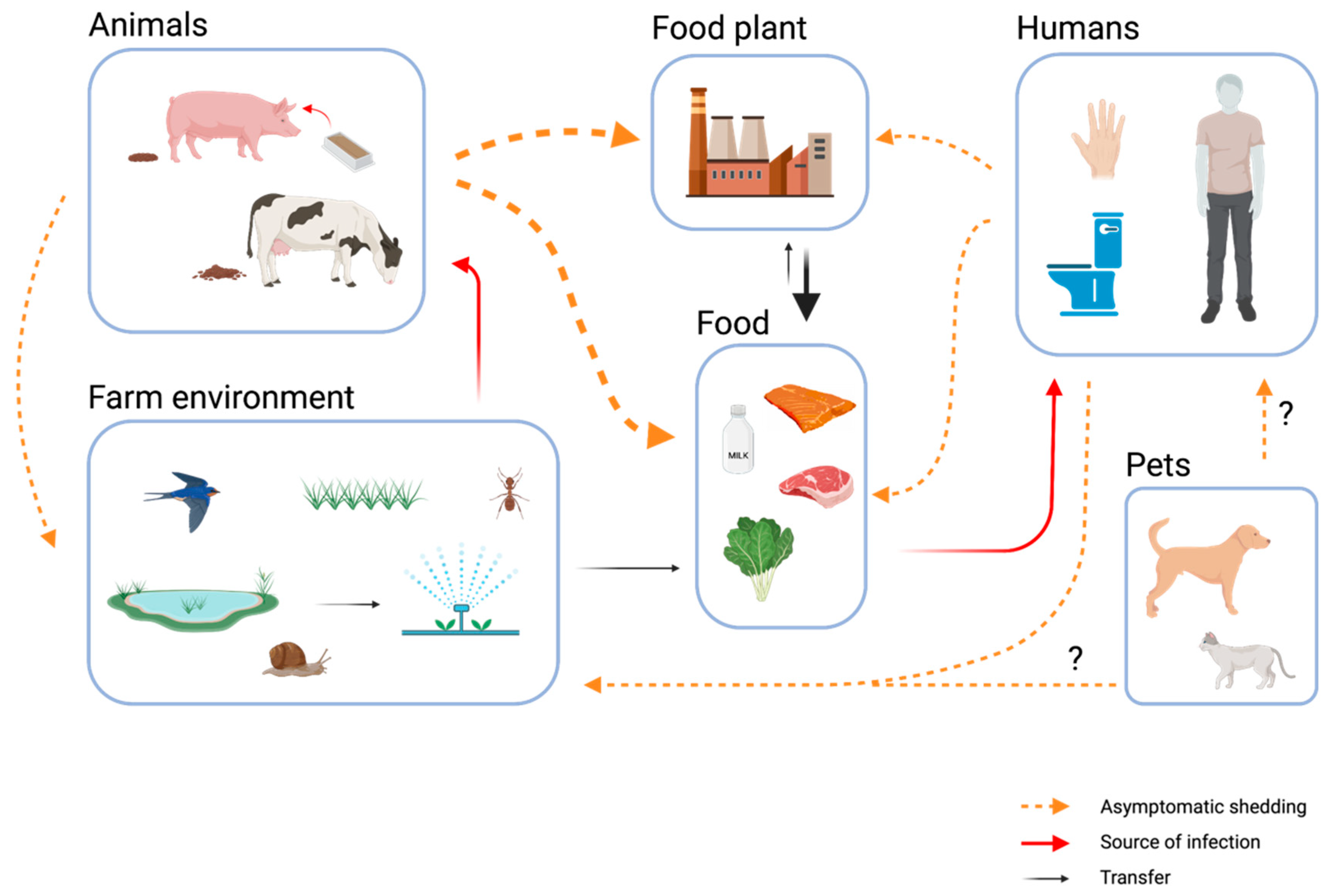
1. Introduction
2. Domestic Animals as Asymptomatic Carriers
3. Carriage of L. monocytogenes by Wild Animals
4. Asymptomatic Carriage of L. monocytogenes in Humans
5. The Impact of Asymptomatic Carriers on the Presence of L. monocytogenes on Farms and in the Food Processing Environment
6. Future Implications
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Anonymous. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Haase, J.K.; Didelot, X.; Lecuit, M.; Korkeala, H.; Group, L.m.M.S.; Achtman, M. The ubiquitous nature of Listeria monocytogenes clones: A large-scale Multilocus Sequence Typing study. Environ. Microbiol. 2014, 16, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Belias, A.; Sullivan, G.; Wiedmann, M.; Ivanek, R. Factors that contribute to persistent Listeria in food processing facilities and relevant interventions: A rapid review. Food Control 2022, 133, 108579. [Google Scholar] [CrossRef]
- Maury, M.M.; Bracq-Dieye, H.; Huang, L.; Vales, G.; Lavina, M.; Thouvenot, P.; Disson, O.; Leclercq, A.; Brisse, S.; Lecuit, M. Hypervirulent Listeria monocytogenes clones’ adaption to mammalian gut accounts for their association with dairy products. Nat. Commun. 2019, 10, 2488. [Google Scholar] [CrossRef]
- Harrand, A.S.; Jagadeesan, B.; Baert, L.; Wiedmann, M.; Orsi, R.H. Evolution of Listeria monocytogenes in a Food Processing Plant Involves Limited Single-Nucleotide Substitutions but Considerable Diversification by Gain and Loss of Prophages. Appl. Environ. Microbiol. 2020, 86, e02493-19. [Google Scholar] [CrossRef]
- Schlech, W.F.; Fischetti, V.A.; Novick, R.P.; Ferretti, J.J.; Portnoy, D.A.; Miriam Braunstein, M.; Rood, J.I. Epidemiology and Clinical Manifestations of Listeria monocytogenes Infection. Microbiol. Spectr. 2019, 7, 3. [Google Scholar] [CrossRef]
- Olier, M.; Garmyn, D.; Rousseaux, S.; Lemaître, J.P.; Piveteau, P.; Guzzo, J. Truncated Internalin A and Asymptomatic Listeria monocytogenes Carriage: In Vivo Investigation by Allelic Exchange. Infect. Immun. 2005, 73, 644–648. [Google Scholar] [CrossRef]
- Nightingale, K.K.; Windham, K.; Martin, K.E.; Yeung, Y.; Wiedmann, M. Select Listeria monocytogenes Subtypes Commonly Found in Foods Carry Distinct Nonsense Mutations in inlA, Leading to Expression of Truncated and Secreted Internalin A, and Are Associated with a Reduced Invasion Phenotype for Human Intestinal Epithelial Cells. Appl. Environ. Microbiol. 2005, 71, 8764–8772. [Google Scholar] [CrossRef]
- Maury, M.M.; Tsai, Y.H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef]
- Besnard, V.; Federighi, M.; Declerq, E.; Jugiau, F.; Cappelier, J.M. Environmental and physico-chemical factors induce VBNC state in Listeria monocytogenes. Vet. Res. 2002, 33, 359–370. [Google Scholar] [CrossRef]
- Lotoux, A.; Milohanic, E.; Bierne, H. The Viable But Non-Culturable State of Listeria monocytogenes in the One-Health Continuum. Front. Cell. Infect. Microbiol. 2022, 12, 849915. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, S.; Robben, C.; Alter, T.; Rossmanith, P.; Mester, P. How to Evaluate Non-Growing Cells—Current Strategies for Determining Antimicrobial Resistance of VBNC Bacteria. Antibiotics 2021, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Hafner, L.; Pichon, M.; Burucoa, C.; Nusser, S.H.A.; Moura, A.; Garcia-Garcera, M.; Lecuit, M. Listeria monocytogenes faecal carriage is common and depends on the gut microbiota. Nat. Commun. 2021, 12, 6826. [Google Scholar] [CrossRef]
- Ho, A.J.; Ivanek, R.; Grohn, Y.T.; Nightingale, K.K.; Wiedmann, M. Listeria monocytogenes fecal shedding in dairy cattle shows high levels of day-to-day variation and includes outbreaks and sporadic cases of shedding of specific L. monocytogenes subtypes. Prev. Vet. Med. 2007, 80, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Wilkes, G.; Edge, T.A.; Gannon, V.P.J.; Jokinen, C.; Lyautey, E.; Neumann, N.F.; Ruecker, N.; Scott, A.; Sunohara, M.; Topp, E.; et al. Associations among pathogenic bacteria, parasites, and environmental and land use factors in multiple mixed-use watersheds. Water Res. 2011, 45, 5807–5825. [Google Scholar] [CrossRef] [PubMed]
- Lyautey, E.; Lapen, D.R.; Wilkes, G.; McCleary, K.; Pagotto, F.; Tyler, K.; Hartmann, A.; Piveteau, P.; Rieu, A.; Robertson, W.J.; et al. Distribution and characteristics of Listeria monocytogenes isolates from surface waters of the South Nation River watershed, Ontario, Canada. Appl. Environ. Microbiol. 2007, 73, 5401–5410. [Google Scholar] [CrossRef] [PubMed]
- Schoder, D.; Wagner, M. Growing awareness of asymptomatic carriage of Listeria monocytogenes. Wien. Tierarztl. Mon. 2012, 99, 322–329. [Google Scholar]
- van Bree, F.P.J.; Bokken, G.C.A.M.; Mineur, R.; Franssen, F.; Opsteegh, M.; van der Giessen, J.W.B.; Lipman, L.J.A.; Overgaauw, P.A.M. Zoonotic bacteria and parasites found in raw meat-based diets for cats and dogs. Vet. Rec. 2018, 182, 50. [Google Scholar] [CrossRef]
- Autio, T.; Markkula, A.; Hellstrom, S.; Niskanen, T.; Lunden, J.; Korkeala, H. Prevalence and genetic diversity of Listeria monocytogenes in the tonsils of pigs. J. Food Prot. 2004, 67, 805–808. [Google Scholar] [CrossRef]
- Oswaldi, V.; Luth, S.; Dzierzon, J.; Meemken, D.; Schwarz, S.; Fessler, A.T.; Felix, B.; Langforth, S. Distribution and Characteristics of Listeria spp. in Pigs and Pork Production Chains in Germany. Microorganisms 2022, 10, 512. [Google Scholar] [CrossRef]
- Hellstrom, S.; Laukkanen, R.; Siekkinen, K.M.; Ranta, J.; Maijala, R.; Korkeala, H. Listeria monocytogenes contamination in pork can originate from farms. J. Food Prot. 2010, 73, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Miró, S.; Tecles, F.; Ramón, M.; Escribano, D.; Hernández, F.; Madrid, J.; Orengo, J.; Martínez-Subiela, S. Causes, consequences and biomarkers of stress in swine: An update. BMC Vet. Res. 2016, 12, 171. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, A.; Ocejo, M.; Oporto, B. Salmonella spp. and Listeria monocytogenes shedding in domestic ruminants and characterization of potentially pathogenic strains. Vet. Microbiol. 2017, 210, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Boerlin, P.; Boerlin-Petzold, F.; Jemmi, T. Use of listeriolysin O and internalin A in a seroepidemiological study of listeriosis in Swiss dairy cows. J. Clin. Microbiol. 2003, 41, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Gorba, C.; Moura, A.; Gomis, J.; Leclercq, A.; Gomez-Martin, A.; Bracq-Dieye, H.; Moce, M.L.; Tessaud-Rita, N.; Jimenez-Trigos, E.; Vales, G.; et al. Ruminant-associated Listeria monocytogenes isolates belong preferentially to dairy-associated hypervirulent clones: A longitudinal study in 19 farms. Environ. Microbiol. 2021, 23, 7617–7631. [Google Scholar] [CrossRef]
- Husu, J.R. Epidemiological studies on the occurrence of Listeria monocytogenes in the feces of dairy cattle. Zent. Vet. B 1990, 37, 276–282. [Google Scholar] [CrossRef]
- Husu, J.R.; Seppanen, J.T.; Sivela, S.K.; Rauramaa, A.L. Contamination of raw milk by Listeria monocytogenes on dairy farms. Zent. Vet. B 1990, 37, 268–275. [Google Scholar] [CrossRef]
- Ivaneka, R.; Grohn, Y.T.; Ho, A.J.J.; Wiedmann, M. Markov chain approach to analyze the dynamics of pathogen fecal shedding—Example of Listeria monocytogenes shedding in a herd of dairy cattle. J. Theor. Biol. 2007, 245, 44–58. [Google Scholar] [CrossRef]
- Vilar, M.J.; Yus, E.; Sanjuán, M.L.; Diéguez, F.J.; Rodríguez-Otero, J.L. Prevalence of and Risk Factors for Listeria Species on Dairy Farms. J. Dairy Sci. 2007, 90, 5083–5088. [Google Scholar] [CrossRef]
- Nightingale, K.K.; Fortes, E.D.; Ho, A.J.; Schukken, Y.H.; Grohn, Y.T.; Wiedmann, M. Evaluation of farm management practices as risk factors for clinical listeriosis and fecal shedding of Listeria monocytogenes in ruminants. J. Am. Vet. Med. Assoc. 2005, 227, 1808–1814. [Google Scholar] [CrossRef]
- Nightingale, K.K.; Schukken, Y.H.; Nightingale, C.R.; Fortes, E.D.; Ho, A.J.; Her, Z.; Grohn, Y.T.; McDonough, P.L.; Wiedmann, M. Ecology and transmission of Listeria monocytogenes infecting ruminants and in the farm environment. Appl. Environ. Microbiol. 2004, 70, 4458–4467. [Google Scholar] [CrossRef] [PubMed]
- Schoder, D.; Melzner, D.; Schmalwieser, A.; Zangana, A.; Winter, P.; Wagner, M. Important vectors for Listeria monocytogenes transmission at farm dairies manufacturing fresh sheep and goat cheese from raw milk. J. Food Prot. 2011, 74, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Moura, A.; Lefrancq, N.; Wirth, T.; Leclercq, A.; Borges, V.; Gilpin, B.; Dallman, T.J.; Frey, J.; Franz, E.; Nielsen, E.M.; et al. Emergence and global spread of Listeria monocytogenes main clinical clonal complex. Sci. Adv. 2021, 7, eabj9805. [Google Scholar] [CrossRef] [PubMed]
- Zundel, E.; Bernard, S. Listeria monocytogenes translocates throughout the digestive tract in asymptomatic sheep. J. Med. Microbiol. 2006, 55, 1717–1723. [Google Scholar] [CrossRef][Green Version]
- Lin, C.H.; Adams, P.J.; Huang, J.F.; Sun, Y.F.; Lin, J.H.; Robertson, I.D. Contamination of chicken carcasses and the abattoir environment with Listeria monocytogenes in Taiwan. Br. Poult. Sci. 2021, 62, 701–709. [Google Scholar] [CrossRef]
- Pohjola, L.; Nykäsenoja, S.; Kivistö, R.; Soveri, T.; Huovilainen, A.; Hänninen, M.L.; Fredriksson-Ahomaa, M. Zoonotic Public Health Hazards in Backyard Chickens. Zoonoses Public Health 2016, 63, 420–430. [Google Scholar] [CrossRef]
- Elsayed, M.M.; Elkenany, R.M.; Zakaria, A.I.; Badawy, B.M. Epidemiological study on Listeria monocytogenes in Egyptian dairy cattle farms’ insights into genetic diversity of multi-antibiotic-resistant strains by ERIC-PCR. Environ. Sci. Pollut. Res. 2022, 29, 54359–54377. [Google Scholar] [CrossRef]
- Weber, A.; Potel, J.; SchaferSchmidt, R.; Prell, A.; Datzmann, C. Investigations on the occurrence of Listeria monocytogenes in fecal samples of domestic and companion animals. Zent. Hyg. Umw. 1995, 198, 117–123. [Google Scholar]
- Oswaldi, V.; Dzierzon, J.; Thieme, S.; Merle, R.; Meemken, D. Slaughter pigs as carrier of Listeria monocytogenes in Germany. J. Consum. Prot. Food Saf. 2021, 16, 109–115. [Google Scholar] [CrossRef]
- Adorisio, E.; Tassone, D.D.; Conte, A.; Annicchiarico, L.S. Lysteria monocytogenes in samples of swines sent to the human alimentary chain. Ann. Ig. Med. Prev. Comunita 2003, 15, 575–581. [Google Scholar]
- Iida, T.; Kanzaki, M.; Maruyama, T.; Inoue, S.; Kaneuchi, C. Prevalence of Listeria monocytogenes in intestinal contents of healthy animals in japan. J. Vet. Med. Sci. 1991, 53, 873–875. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Iwabuchi, E.; Yamamoto, S.; Muramatsu, M.; Takashima, I.; Hirai, K. Prevalence and characteristics of Listeria monocytogenes in feces of black beef cattle reared in three geographically distant areas in Japan. Foodborne Pathog. Dis. 2014, 11, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Obaidat, M.M.; Stringer, A.P. Prevalence, molecular characterization, and antimicrobial resistance profiles of Listeria monocytogenes, Salmonella enterica, and Escherichia coli O157:H7 on dairy cattle farms in Jordan. J. Dairy Sci. 2019, 102, 8710–8720. [Google Scholar] [CrossRef]
- Stipetic, K.; Chang, Y.C.; Peters, K.; Salem, A.; Doiphode, S.H.; McDonough, P.L.; Chang, Y.F.; Sultan, A.; Mohammed, H.O. The risk of carriage of Salmonella spp. and Listeria monocytogenes in food animals in dynamic populations. Vet. Med. Sci. 2016, 2, 246–254. [Google Scholar] [CrossRef]
- Bandelj, P.; Jamnikar-Ciglenecki, U.; Ocepek, M.; Blagus, R.; Vengust, M. Risk factors associated with fecal shedding of Listeria monocytogenes by dairy cows and calves. J. Vet. Intern. Med. 2018, 32, 1773–1779. [Google Scholar] [CrossRef]
- Locatelli, A.; Lewis, M.A.; Rothrock, M.J., Jr. The Distribution of Listeria in Pasture-Raised Broiler Farm Soils Is Potentially Related to University of Vermont Medium Enrichment Bias toward Listeria innocua over Listeria monocytogenes. Front. Vet. Sci. 2017, 4, 227. [Google Scholar] [CrossRef]
- Mohammed, H.O.; Stipetic, K.; McDonough, P.L.; Gonzalez, R.N.; Nydam, D.V.; Atwill, E.R. Identification of potential on-farm sources of Listeria monocytogenes in herds of dairy cattle. Am. J. Vet. Res. 2009, 70, 383–388. [Google Scholar] [CrossRef]
- Yoshida, T.; Sugimoto, T.; Sato, M.; Hirai, K. Incidence of Listeria monocytogenes in wild animals in Japan. J. Vet. Med. Sci. 2000, 62, 673–675. [Google Scholar] [CrossRef][Green Version]
- Bouttefroy, A.; Lemaitre, J.P.; Rousset, A. Prevalence of Listeria sp. in droppings from urban rooks (Corvus frugilegus). J. Appl. Microbiol. 1997, 82, 641–647. [Google Scholar] [CrossRef]
- Hellstrom, S.; Kiviniemi, K.; Autio, T.; Korkeala, H. Listeria monocytogenes is common in wild birds in Helsinki region and genotypes are frequently similar with those found along the food chain. J. Appl. Microbiol. 2008, 104, 883–888. [Google Scholar] [CrossRef]
- Lyautey, E.; Hartmann, A.; Pagotto, F.; Tyler, K.; Lapen, D.R.; Wilkes, G.; Piveteau, P.; Rieu, A.; Robertson, W.J.; Medeiros, D.T.; et al. Characteristics and frequency of detection of fecal Listeria monocytogenes shed by livestock, wildlife, and humans. Can. J. Microbiol. 2007, 53, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Weindl, L.; Frank, E.; Ullrich, U.; Heurich, M.; Kleta, S.; Ellerbroek, L.; Gareis, M. Listeria monocytogenes in Different Specimens from Healthy Red Deer and Wild Boars. Foodborne Pathog. Dis. 2016, 13, 391–397. [Google Scholar] [CrossRef]
- Nowakiewicz, A.; Zieba, P.; Ziolkowska, G.; Gnat, S.; Muszynska, M.; Tomczuk, K.; Dziedzic, B.M.; Ulbrych, L.; Troscianczyk, A. Free-Living Species of Carnivorous Mammals in Poland: Red Fox, Beech Marten, and Raccoon as a Potential Reservoir of Salmonella, Yersinia, Listeria spp. and Coagulase-Positive Staphylococcus. PLoS ONE 2016, 11, e0155533. [Google Scholar] [CrossRef] [PubMed]
- Najdenski, H.; Dimova, T.; Zaharieva, M.M.; Nikolov, B.; Petrova-Dinkova, G.; Dalakchieva, S.; Popov, K.; Hristova-Nikolova, I.; Zehtindjiev, P.; Peev, S.; et al. Migratory birds along the Mediterranean–Black Sea Flyway as carriers of zoonotic pathogens. Can. J. Microbiol. 2018, 64, 915–924. [Google Scholar] [CrossRef]
- Gorham, T.J.; Lee, J. Pathogen Loading From Canada Geese Faeces in Freshwater: Potential Risks to Human Health Through Recreational Water Exposure. Zoonoses Public Health 2016, 63, 177–190. [Google Scholar] [CrossRef]
- Quessy, S.; Messier, S. Prevalence of Salmonella spp., Campylobacter spp. and Listeria spp. in ring-billed gulls (Larus delawarensis). J. Wildl. Dis. 1992, 28, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wang, Y.; Wang, Y.; Li, H.; Luo, L.; Wang, P.; Zhang, L.; Li, H.; Liu, J.; Lu, L.; et al. Prevalence and Characteristics of Listeria ivanovii Strains in Wild Rodents in China. Vector-Borne Zoonotic Dis. 2019, 19, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, L.; Lan, R.; Salazar, J.K.; Liu, J.; Xu, J.; Ye, C. Isolation and characterization of Listeria species from rodents in natural environments in China. Emerg. Microbes Infect. 2017, 6, e44. [Google Scholar] [CrossRef]
- Laaksonen, S.; Oksanen, A.; Julmi, J.; Zweifel, C.; Fredriksson-Ahomaa, M.; Stephan, R. Presence of foodborne pathogens, extended-spectrum beta-lactamase -producing Enterobacteriaceae, and methicillin-resistant Staphylococcus aureus in slaughtered reindeer in northern Finland and Norway. Acta Vet. Scand. 2017, 59, 2. [Google Scholar] [CrossRef][Green Version]
- Weber, A.; Potel, J.; Schafer-Schmidt, R. The occurrence of Listeria monocytogenes in fecal samples of pigeons. Berl. Und Muenchener Tieraerztliche Wochenschr. 1995, 108, 26–27. [Google Scholar]
- Onjong, H.A.; Ngayo, M.O.; Mwaniki, M.; Wambui, J.; Njage, P.M.K. Microbiological Safety of Fresh Tilapia (Oreochromis niloticus) from Kenyan Fresh Water Fish Value Chains. J. Food Prot. 2018, 81, 1973–1981. [Google Scholar] [CrossRef] [PubMed]
- Gnat, S.; Troscianczyk, A.; Nowakiewicz, A.; Majer-Dziedzic, B.; Ziolkowska, G.; Dziedzic, R.; Zieba, P.; Teodorowski, O. Experimental studies of microbial populations and incidence of zoonotic pathogens in the faeces of red deer (Cervus elaphus). Lett. Appl. Microbiol. 2015, 61, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Wacheck, S.; Fredriksson-Ahomaa, M.; Konig, M.; Stolle, A.; Stephan, R. Wild boars as an important reservoir for foodborne pathogens. Foodborne Pathog. Dis. 2010, 7, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Orsi, R.H.; Chen, R.; Gunderson, M.; Roof, S.; Wiedmann, M.; Childs-Sanford, S.E.; Cummings, K.J. Characterization of Listeria monocytogenes isolated from wildlife in central New York. Vet. Med. Sci. 2022, 8, 1319–1329. [Google Scholar] [CrossRef]
- Ferreira, V.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M.J. Listeria monocytogenes persistence in food-associated environments: Epidemiology, strain characteristics, and implications for public health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.-L.; Opitz, H.M.; Bayer, R.C.; Kling, L.J.; Halteman, W.A.; Martin, R.E.; Slabyj, B.M. Listeria monocytogenes in an Atlantic Salmon (Salmo salar) Processing Environment. J. Food Prot. 2005, 68, 1635–1640. [Google Scholar] [CrossRef]
- Fagerlund, A.; Wagner, E.; Møretrø, T.; Heir, E.; Moen, B.; Rychli, K.; Langsrud, S. Pervasive Listeria monocytogenes Is Common in the Norwegian Food System and Is Associated with Increased Prevalence of Stress Survival and Resistance Determinants. Appl. Environ. Microbiol. 2022, 88, e0086122. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, H.; Wirtanen, G. Ecology of Listeria spp. in a fish farm and molecular typing of Listeria monocytogenes from fish farming and processing companies. Int. J. Food Microbiol. 2006, 112, 138–146. [Google Scholar] [CrossRef]
- Simothy, L.; Mahomoodally, F.; Neetoo, H. A study on the potential of ants to act as vectors of foodborne pathogens. AIMS Microbiol. 2018, 4, 319–333. [Google Scholar] [CrossRef]
- Pushkareva, V.I.; Ermolaeva, S.A. Listeria monocytogenes virulence factor Listeriolysin O favors bacterial growth in co-culture with the ciliate Tetrahymena pyriformis, causes protozoan encystment and promotes bacterial survival inside cysts. BMC Microbiol. 2010, 10, 26. [Google Scholar] [CrossRef]
- Statista GmbH. Anzahl der Reptilien in Europa Nach Ländern im Jahr 2021. Available online: https://de.statista.com/statistik/daten/studie/454104/umfrage/reptilien-in-europa-nach-laendern/ (accessed on 7 October 2022).
- Gismervik, K.; Aspholm, M.; Rorvik, L.M.; Bruheim, T.; Andersen, A.; Skaar, I. Invading slugs (Arion vulgaris) can be vectors for Listeria monocytogenes. J. Appl. Microbiol. 2015, 118, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Schuchat, A.; Swaminathan, B.; Broome, C.V. Epidemiology of human listeriosis. Clin. Microbiol. Rev. 1991, 4, 169–183. [Google Scholar] [CrossRef]
- Grif, K.; Hein, I.; Wagner, M.; Brandl, E.; Mpamugo, O.; McLauchlin, J.; Dierich, M.P.; Allerberger, F. Prevalence and characterization of Listeria monocytogenes in the feces of healthy Austrians. Wien. Klin. Wochenschr. 2001, 113, 737–742. [Google Scholar] [PubMed]
- Sauders, B.D.; Pettit, D.; Currie, B.; Suits, P.; Evans, A.; Stellrecht, K.; Dryja, D.M.; Slate, D.; Wiedmann, M. Low prevalence of Listeria monocytogenes in human stool. J. Food Prot. 2005, 68, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Grif, K.; Patscheider, G.; Dierich, M.P.; Allerberger, F. Incidence of fecal carriage of Listeria monocytogenes in three healthy volunteers: A one-year prospective stool survey. Eur. J. Clin. Microbiol. Infect. Dis. 2003, 22, 16–20. [Google Scholar] [CrossRef]
- EFSA Panelon Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K.; et al. Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar] [CrossRef]
- MacGowan, A.P.; Marshall, R.J.; MacKay, I.M.; Reeves, D.S. Listeria faecal carriage by renal transplant recipients, haemodialysis patients and patients in general practice: Its relation to season, drug therapy, foreign travel, animal exposure and diet. Epidemiol. Infect. 1991, 106, 157–166. [Google Scholar] [CrossRef]
- Freitag, I.G.R.; de Castro Lisboa Pereira, R.; Machado, E.S.; Hofer, E.; Vallim, D.C.; Hofer, C.B. Prevalence of Listeria monocytogenes fecal carriers in HIV-infected and -uninfected pregnant women from Brazil. Braz. J. Microbiol. 2021, 52, 2081–2084. [Google Scholar] [CrossRef]
- Lamont, R.J.; Postlethwaite, R. Carriage of Listeria monocytogenes and related species in pregnant and non-pregnant women in Aberdeen, Scotland. J. Infect. 1986, 13, 187–193. [Google Scholar] [CrossRef]
- Mascola, L.; Ewert, D.P.; Eller, A. Listeriosis: A previously unreported medical complication in women with multiple gestations. Am. J. Obstet. Gynecol. 1994, 170, 1328–1332. [Google Scholar] [CrossRef]
- Sarr, M.; Alou, M.T.; Delerce, J.; Khelaifia, S.; Diagne, N.; Diallo, A.; Bassene, H.; Bréchard, L.; Bossi, V.; Mbaye, B.; et al. A Listeria monocytogenes clone in human breast milk associated with severe acute malnutrition in West Africa: A multicentric case-controlled study. PLoS Negl. Trop. Dis. 2021, 15, e0009555. [Google Scholar] [CrossRef] [PubMed]
- Tahoun, A.; Abou Elez, R.M.M.; Abdelfatah, E.N.; Elsohaby, I.; El-Gedawy, A.A.; Elmoslemany, A.M. Listeria monocytogenes in raw milk, milking equipment and dairy workers: Molecular characterization and antimicrobial resistance patterns. J. Glob. Antimicrob. Resist. 2017, 10, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.E. Listeria isolations from feces of patients with diarrhea and from healthy food handlers. Infection 1990, 18, 97–99. [Google Scholar] [CrossRef] [PubMed]
- Meghdadi, H.; Khosravi, A.D.; Sheikh, A.F.; Alami, A.; Nassirabady, N. Isolation and characterization of Listeria monocytogenes from environmental and clinical sources by culture and PCR-RFLP methods. Iran. J. Microbiol. 2019, 11, 7–12. [Google Scholar] [CrossRef]
- Akkaya, L.; Alisarli, M.; Cetinkaya, Z.; Kara, R.; Telli, R. Occurrence of Escherichia coli O157: H7/O157, Listeria monocytogenes and Salmonella spp. in beef slaughterhouse environments, equipment and workers. J. Muscle. Foods 2008, 19, 261–274. [Google Scholar] [CrossRef]
- Pala, K.; Oezakin, C.; Akis, N.; Sinirtas, M.; Gedikoglu, S.; Aytekin, H. Asymptomatic carriage of bacteria in food workers in Nilufer district, Bursa, Turkey. Turk. J. Med. Sci. 2010, 40, 133–139. [Google Scholar] [CrossRef]
- Alam, M.S.; Gangiredla, J.; Hasan, N.A.; Barnaba, T.; Tartera, C. Aging-Induced Dysbiosis of Gut Microbiota as a Risk Factor for Increased Listeria monocytogenes Infection. Front. Immunol. 2021, 12, 672353. [Google Scholar] [CrossRef]
- European Food Safety Authority. Analysis of the baseline survey on the prevalence of Listeria monocytogenes in certain ready-to-eat (RTE) foods in the EU, 2010-2011 Part A: Listeria monocytogenes prevalence estimates. EFSA J. 2013, 11, 3241. [Google Scholar] [CrossRef]
- Wagner, M.; Auer, B.; Trittremmel, C.; Hein, I.; Schoder, D. Survey on the Listeria contamination of ready-to-eat food products and household environments in Vienna, Austria. Zoonoses Public Health 2007, 54, 16–22. [Google Scholar] [CrossRef]
- Esteban, J.I.; Oporto, B.; Aduriz, G.; Juste, R.A.; Hurtado, A. Faecal shedding and strain diversity of Listeria monocytogenes in healthy ruminants and swine in Northern Spain. BMC Vet. Res. 2009, 5, 2. [Google Scholar] [CrossRef]
- Fenlon, D.R.; Wilson, J.; Donachie, W. The incidence and level of Listeria monocytogenes contamination of food sources at primary production and initial processing. J. Appl. Bacteriol. 1996, 81, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Fenlon, D.R. Wild birds and silage as reservoirs of Listeria in the agricultural environment. J. Appl. Bacteriol. 1985, 59, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Honjoh, K.; Iwazako, Y.; Lin, Y.; Kijima, N.; Miyamoto, T. Possibilities for Contamination of Tomato Fruit by Listeria monocytogenes during Cultivation. Food Sci. Technol. Res. 2016, 22, 349–357. [Google Scholar] [CrossRef][Green Version]
- Weller, D.; Belias, A.; Green, H.; Roof, S.; Wiedmann, M. Landscape, Water Quality, and Weather Factors Associated With an Increased Likelihood of Foodborne Pathogen Contamination of New York Streams Used to Source Water for Produce Production. Front. Sustain. Food Syst. 2020, 3, 124. [Google Scholar] [CrossRef]
- Weller, D.; Wiedmann, M.; Strawn, L.K. Spatial and Temporal Factors Associated with an Increased Prevalence of Listeria monocytogenes in Spinach Fields in New York State. Appl. Environ. Microbiol. 2015, 81, 6059–6069. [Google Scholar] [CrossRef]
- Weller, D.; Wiedmann, M.; Strawn, L.K. Irrigation Is Significantly Associated with an Increased Prevalence of Listeria monocytogenes in Produce Production Environments in New York State. J. Food Prot. 2015, 78, 1132–1141. [Google Scholar] [CrossRef]
- Gu, G.; Strawn, L.K.; Ottesen, A.R.; Ramachandran, P.; Reed, E.A.; Zheng, J.; Boyer, R.R.; Rideout, S.L. Correlation of Salmonella enterica and Listeria monocytogenes in Irrigation Water to Environmental Factors, Fecal Indicators, and Bacterial Communities. Front. Microbiol. 2020, 11, 557289. [Google Scholar] [CrossRef]
- Huang, Y.; Hansen, L.T.; Ragush, C.M.; Jamieson, R.C. Disinfection and removal of human pathogenic bacteria in arctic waste stabilization ponds. Environ. Sci. Pollut. Res. Int. 2018, 25, 32881–32893. [Google Scholar] [CrossRef]
- Walland, J.; Lauper, J.; Frey, J.; Imhof, R.; Stephan, R.; Seuberlich, T.; Oevermann, A. Listeria monocytogenes infection in ruminants: Is there a link to the environment, food and human health? A review. Schweiz Arch. Tierheilkd 2015, 157, 319–328. [Google Scholar] [CrossRef]
- Oliver, S.P.; Jayarao, B.M.; Almeida, R.A. Foodborne pathogens in milk and the dairy farm environment: Food safety and public health implications. Foodborne Pathog. Dis. 2005, 2, 115–129. [Google Scholar] [CrossRef]
- de Valk, H.; Vaillant, V.; Jacquet, C.; Rocourt, J.; Le Querrec, F.; Stainer, F.; Quelquejeu, N.; Pierre, O.; Pierre, V.; Desenclos, J.C.; et al. Two consecutive nationwide outbreaks of Listeriosis in France, October 1999-February 2000. Am. J. Epidemiol. 2001, 154, 944–950. [Google Scholar] [CrossRef] [PubMed][Green Version]
- McLauchlin, J.; Hall, S.M.; Velani, S.K.; Gilbert, R.J. Human listeriosis and pate: A possible association. BMJ 1991, 303, 773–775. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Giovannacci, I.; Ragimbeau, C.; Queguiner, S.; Salvat, G.; Vendeuvre, J.L.; Carlier, V.; Ermel, G. Listeria monocytogenes in pork slaughtering and cutting plants: Use of RAPD, PFGE and PCR-REA for tracing and molecular epidemiology. Int. J. Food Microbiol. 1999, 53, 127–140. [Google Scholar] [CrossRef]
- Beloeil, P.A.; Chauvin, C.; Toquin, M.T.; Fablet, C.; Le Notre, Y.; Salvat, G.; Madec, F.; Fravalo, P. Listeria monocytogenes contamination of finishing pigs: An exploratory epidemiological survey in France. Vet. Res. 2003, 34, 737–748. [Google Scholar] [CrossRef]
- Falardeau, J.; Johnson, R.P.; Pagotto, F.; Wang, S. Occurrence, characterization, and potential predictors of verotoxigenic Escherichia coli, Listeria monocytogenes, and Salmonella in surface water used for produce irrigation in the Lower Mainland of British Columbia, Canada. PLoS ONE 2017, 12, e0185437. [Google Scholar] [CrossRef]
- Ojeniyi, B.; Wegener, H.C.; Jensen, N.E.; Bisgaard, M. Listeria monocytogenes in poultry and poultry products: Epidemiological investigations in seven Danish abattoirs. J. Appl. Bacteriol. 1996, 80, 395–401. [Google Scholar] [CrossRef]
- Moosavy, M.H.; Shavisi, N.; Warriner, K.; Mosta-Favi, E. Bacterial Contamination of Iranian Paper Currency. Iran. J. Public Health 2013, 42, 1067–1070. [Google Scholar]
- Marshall, J.C.; Soboleva, T.K.; Jamieson, P.; French, N.P. Estimating Bacterial Pathogen Levels in New Zealand Bulk Tank Milk. J. Food Prot. 2016, 79, 771–780. [Google Scholar] [CrossRef]
- Jongman, M.; Korsten, L. Microbial quality and suitability of roof-harvested rainwater in rural villages for crop irrigation and domestic use. J. Water Health 2016, 14, 961–971. [Google Scholar] [CrossRef]
- Ahlstrom, C.A.; Manuel, C.S.; Den Bakker, H.C.; Wiedmann, M.; Nightingale, K.K. Molecular ecology of Listeria spp., Salmonella, Escherichia coli O157:H7 and non-O157 Shiga toxin-producing E. coli in pristine natural environments in Northern Colorado. J. Appl. Microbiol. 2018, 124, 511–521. [Google Scholar] [CrossRef]
- Dungan, R.S.; Klein, M.; Leytem, A.B. Quantification of bacterial indicators and zoonotic pathogens in dairy wastewater ponds. Appl. Environ. Microbiol. 2012, 78, 8089–8095. [Google Scholar] [CrossRef] [PubMed]
- Belias, A.; Strawn, L.K.; Wiedmann, M.; Weller, D. Small Produce Farm Environments Can Harbor Diverse Listeria monocytogenes and Listeria spp. Populations. J. Food Prot. 2021, 84, 113–121. [Google Scholar] [CrossRef]
- Jensen, A.; Thomsen, L.E.; Jorgensen, R.L.; Larsen, M.H.; Roldgaard, B.B.; Christensen, B.B.; Vogel, B.F.; Gram, L.; Ingmer, H. Processing plant persistent strains of Listeria monocytogenes appear to have a lower virulence potential than clinical strains in selected virulence models. Int. J. Food Microbiol. 2008, 123, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Lunden, J.M.; Autio, T.J.; Sjoberg, A.M.; Korkeala, H.J. Persistent and nonpersistent Listeria monocytogenes contamination in meat and poultry processing plants. J. Food Prot. 2003, 66, 2062–2069. [Google Scholar] [CrossRef] [PubMed]
- Kastbjerg, V.G.; Larsen, M.H.; Gram, L.; Ingmer, H. Influence of sublethal concentrations of common disinfectants on expression of virulence genes in Listeria monocytogenes. Appl. Environ. Microbiol. 2010, 76, 303–309. [Google Scholar] [CrossRef]
- Rodrigues, D.; Cerca, N.; Teixeira, P.; Oliveira, R.; Ceri, H.; Azeredo, J. Listeria monocytogenes and Salmonella enterica enteritidis biofilms susceptibility to different disinfectants and stress-response and virulence gene expression of surviving cells. Microb. Drug Resist. 2011, 17, 181–189. [Google Scholar] [CrossRef]
- Prates, D.D.; Haubert, L.; Wurfel, S.D.; Cavicchioli, V.Q.; Nero, L.A.; da Silva, W.P. Listeria monocytogenes in dairy plants in Southern Brazil: Occurrence, virulence potential, and genetic diversity. J. Food Saf. 2019, 39, e12695. [Google Scholar] [CrossRef]
- Alessandria, V.; Rantsiou, K.; Dolci, P.; Cocolin, L. Molecular methods to assess Listeria monocytogenes route of contamination in a dairy processing plant. Int. J. Food Microbiol. 2010, 141, S156–S162. [Google Scholar] [CrossRef]
- Ibba, M.; Cossu, F.; Spanu, V.; Virdis, S.; Spanu, C.; Scarano, C.; De Santis, E.P.L. Listeria monocytogenes contamination in dairy plants: Evaluation of Listeria monocytogenes environmental contamination in two cheese-making plants using sheeps milk. Ital. J. Food Saf. 2013, 2, 109–112. [Google Scholar] [CrossRef]
- Mafu, A.A.; Roy, D.; Goulet, J.; Magny, P. Attachment of Listeria monocytogenes to Stainless Steel, Glass, Polypropylene, and Rubber Surfaces After Short Contact Times. J. Food Prot. 1990, 53, 742–746. [Google Scholar] [CrossRef]
- Lawrence, L.M.; Gilmour, A. Characterization of Listeria monocytogenes isolated from poultry products and from the poultry-processing environment by random amplification of polymorphic DNA and multilocus enzyme electrophoresis. Appl. Environ. Microbiol. 1995, 61, 2139–2144. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Country | Animal | Target | Sample (n) | Positive (%) | Ref. |
|---|---|---|---|---|---|
| Austria | Sheep/Goat | L. spp. | Feces (53) | 42.6 | [32] |
| L. monocytogenes | 13 | ||||
| Egypt | Cattle | L. monocytogenes | Feces (660) | 6.8 | [37] |
| Milk (660) | 5.9 | ||||
| Finland | Chicken | L monocytogenes | Cloacal swabs (457) | 1.3 | [36] |
| Germany | Cattle | L. monocytogenes | Feces (138) | 33 | [38] |
| Sheep | (100) | 8 | |||
| Hens | (100) | 8 | |||
| Pigs | (34) | 5.9 | |||
| Horses | (400) | 4.8 | |||
| Dogs | (300) | 1.3 | |||
| Cats | (275) | 0.4 | |||
| Germany | Pigs | L. monocytogenes | Tonsils (430) | 1.6 | [39] |
| L. innocua | 1.2 | ||||
| Italy | Pigs | L. monocytogenes | Salivary gland, lymph nodes, tonsils (189) | 13.2 | [40] |
| Japan | Cattle | L. monocytogenes | Feces (1705) | 1.9 | [41] |
| Pigs | 0.6 | ||||
| Dogs | 0.9 | ||||
| Rats | 6.5 | ||||
| Japan | Cattle | L. monocytogenes | Feces (1738) | 6 | [42] |
| Jordan | Cattle | L. monocytogenes | Feces (610) | 1.5 | [43] |
| Qatar | Camel | L. monocytogenes | Feces (50) | 4 | [44] |
| Slovenia | Cows | L. monocytogenes | Feces (540) | 18.2 | [45] * |
| Calves | (511) | 8.4 | |||
| Spain | Cattle (beef) | L. monocytogenes | Feces (301) | 42.3 | [23] |
| Cattle (dairy) | 46.3 | ||||
| Sheep | 23.5 | ||||
| Spain | Cattle | L. monocytogenes | Feces (953) | 4.3 | [25] |
| Sheep | Feces (483) | 5.8 | |||
| Goat | Feces (333) | 0.3 | |||
| Switzerland | Cattle | Ab to LO | Serum (1652) | 11 | [24] |
| and IA ** | |||||
| Taiwan | Chicken | L. monocytogenes | Carcass rinse (246) | 11.4 | [35] |
| USA | Cattle | L. monocytogenes | Feces (825) | 31 | [14] *** |
| USA | Cattle | L. monocytogenes | Feces (528) | 20.2 | [31] **** |
| USA | Broiler | L. monocytogenes | Feces (555) | 14.9 | [46] |
| USA (Central NY State) | Cattle | L. monocytogenes | Milk (1412) | 13 | [47] |
| Udder swab (1408) | 19 | ||||
| Feces (1414) | 43 |
| Country | Animal | Target | Sample (n) | Positive (%) | Ref. |
|---|---|---|---|---|---|
| Austria/ Germany | Red deer | L. monocytogenes | samples* (45) | 42 | [52] |
| Wild boar | (49) | 25 | |||
| Bulgaria | Birds (Riparia riparia, Motacilla flava) | L. monocytogenes | Feces (673) | 0.6 | [54] |
| Canada | Geese | L. spp. | Feces (495) | 9.5 | [55] |
| L. monocytogenes | 4.0 | ||||
| Canada | Gulls (Laurus delawarensis) | L. monocytogenes | Cloacal swabs (264) | 9.5 | [56] |
| China | Rodents | L. m | Feces (702) | 0.3 | [57] |
| L. ivanovii | 3.7 | ||||
| L. innocua | 6.7 | ||||
| China | Rodents | L. spp. | Feces (341) | 9 | [58] |
| L. monocytogenes | 3.2 | ||||
| L. innocua | 2.9 | ||||
| Finland | Birds | L. monocytogenes | Feces (212) | 36 | [50] |
| Finland/Norway | Reindeer | L. monocytogenes | Feces (470) | 3.2 | [59] |
| France | Rooks | L. monocytogenes | Feces (112) | 33 | [49] |
| L. innocua | 24 | ||||
| L. seeligeri | 8 | ||||
| Germany | Pigeons | L. monocytogenes | Feces (350) | 0.8 | [60] |
| L. innocua | 2.3 | ||||
| L. seeligeri | 0.6 | ||||
| Japan | Crows | L. monocytogenes | Feces (301) | 1.7 | [48] |
| L. innocua | 43 | ||||
| Kenya | Nile tilapia | L. monocytogenes | Muscle (167) | 1.2 | [61] |
| Poland | Red deer | L. monocytogenes | Feces (120) | 1.75 | [62] |
| Poland | Red fox, beech marten, racoon | L. monocytogenes | Rectal swab (286) | 3.5 | [53] |
| (65) | 6.1 | ||||
| (70) | 4.3 | ||||
| Switzerland | Wild boars | L. monocytogenes | Tonsils (153) | 17 | [63] |
| Feces (153) | 1 | ||||
| USA (central New York) | Reptiles | L. monocytogenes | Feces (17) | 12 | [64] |
| Mammals | (64) | 8 | |||
| Birds | (242) | 4.5 |
| Country | Target | Sample (n) | Positive (%) | Reference |
|---|---|---|---|---|
| Austria | L. monocytogenes | Feces, healthy people (505) | 0.2 | [74] |
| Brazil | L. monocytogenes | Feces, pregnant women (213) | 2.4 | [79] |
| L. spp. | 7.5 | |||
| Egypt | L. monocytogenes | Hand swabs, farm workers (100) | 16 | [83] |
| L. innocua | 2 | |||
| France | hly gene | Feces (900) | 10 | [13] |
| Germany | L. monocytogenes | Feces, patients with diarrhea (1000) | 0.6 | [84] |
| L. innocua | 1.7 | |||
| Germany | L. monocytogenes | Feces, healthy people (2000) | 0.8 | [84] |
| L. innocua | 2 | |||
| Iran | L. monocytogenes | Feces (80) | 7.5 | [85] |
| Vaginal swabs (80 samples from women with at least two abortions) | 11.3 | |||
| Senegal | L. monocytogenes | Breast milk, mothers of SAM children (120) | 100 | [82] |
| Breast milk, mothers of healthy children (32) | 37.5 | |||
| Turkey | L. monocytogenes | Hand and clothes swabs, abattoir workers (70) | 5.7 | [86] |
| Turkey | L. monocytogenes | Feces (1061) | 0.9 | [87] |
| UK | L. monocytogenes | Feces, patients with gastroenteritis (171) | 1.8 | [78] |
| USA | L. monocytogenes | Feces (827) | 0.12 | [75] |
| Country | Target | Sample (n) | Positive (%) | Ref. |
|---|---|---|---|---|
| Austria | L. spp. | Working Boots (53) | 51 | [32] |
| Floor (53) | 39.3 | |||
| L. monocytogenes | Working Boots (53) | 15.7 | ||
| Floor (53) | 7.9 | |||
| Canada | L. monocytogenes | Irrigation water (223) | 10.3 | [106] |
| Canada (Arctic region) | L. monocytogenes | Wastewater stabilization ponds (109) | 99 | [99] |
| Denmark | L. monocytogenes | Abattoir poultry (3080) | 8.0 | [107] |
| Egypt | L. monocytogenes | Water (36) | 8.3 | [37] |
| Silage (36) | 27.8 | |||
| Manure (36) | 19.4 | |||
| Soil (36) | 8.3 | |||
| Milking equipment (432) | 6.9 | |||
| Germany | L. monocytogenes | Slaughterhouse (environment and equipment, 77) | 0.9 | [20] |
| Iran | L. monocytogenes | Water (180) | 16.7 | [85] |
| Iran | L. monocytogenes | Iranian currency (108) | 0.93 | [108] |
| Jordan | L. monocytogenes | Bulk tank milk (305) | 7.5 | [43] |
| New Zealand | L. monocytogenes | Bulk tank milk (400) | 4.0 | [109] * |
| South Africa | L. monocytogenes | Roof-harvested rainwater (264) | 22 | [110] |
| Taiwan | L. monocytogenes | Abattoir environment (246) | 0 | [35] |
| USA | L. monocytogenes | Soil (555) | 15.3 | [46] |
| USA | L. spp.** | Stream water (196) | 28 | [95] |
| L. monocytogenes | 10 | |||
| USA (Colorado, wilderness area) | L. monocytogenes | Soil, water, sediment, surface soil and wildlife fecal samples (572) | 0.23 | [111] |
| L. spp. | 1.5 | |||
| USA (Idaho) | L. monocytogenes | Dairy wastewater ponds (30) | 6.7 | [112] |
| USA (New York State) | L. monocytogenes | Water (132) | 48 | [113] |
| L. innocua | 10.6 | |||
| L. monocytogenes | Feces (77) | 29 | ||
| L. innocua | 8 | |||
| USA (NYS) | L. spp. | Spinach field soil (1092) | 12 | [96] |
| L. monocytogenes | 7.8 | |||
| USA (NYS) | L. spp. | Pond/river water used for irrigation (9) | 44 | [97] |
| L. monocytogenes | 22 | |||
| USA (Virginia) | L. monocytogenes | irrigation water | [98] | |
| pond (48) | 27.1 | |||
| well (48) | 4.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schoder, D.; Guldimann, C.; Märtlbauer, E. Asymptomatic Carriage of Listeria monocytogenes by Animals and Humans and Its Impact on the Food Chain. Foods 2022, 11, 3472. https://doi.org/10.3390/foods11213472
Schoder D, Guldimann C, Märtlbauer E. Asymptomatic Carriage of Listeria monocytogenes by Animals and Humans and Its Impact on the Food Chain. Foods. 2022; 11(21):3472. https://doi.org/10.3390/foods11213472
Chicago/Turabian StyleSchoder, Dagmar, Claudia Guldimann, and Erwin Märtlbauer. 2022. "Asymptomatic Carriage of Listeria monocytogenes by Animals and Humans and Its Impact on the Food Chain" Foods 11, no. 21: 3472. https://doi.org/10.3390/foods11213472
APA StyleSchoder, D., Guldimann, C., & Märtlbauer, E. (2022). Asymptomatic Carriage of Listeria monocytogenes by Animals and Humans and Its Impact on the Food Chain. Foods, 11(21), 3472. https://doi.org/10.3390/foods11213472