
Characterisation of Listeria monocytogenes Isolates from Hunted Game and Game Meat from Finland

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Listeria Monocytogenes Isolates
2.2. Whole Genome Sequencing (WGS)
2.3. Characterisation of Listeria Monocytogenes Isolates
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lecuit, M. Listeria Monocytogenes, a Model in Infection Biology. Cell. Microbiol. 2020, 22, e13186. [Google Scholar] [CrossRef] [PubMed]
- Quereda, J.J.; Morón-García, A.; Palacios-Gorba, C.; Dessaux, C.; García-del Portillo, F.; Pucciarelli, M.G.; Ortega, A.D. Pathogenicity and Virulence of Listeria Monocytogenes: A Trip from Environmental to Medical Microbiology. Virulence 2021, 12, 2509–2545. [Google Scholar] [CrossRef]
- EFSA; ECDC. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar]
- Matereke, L.T.; Okoh, A.I. Listeria Monocytogenes Virulence, Antimicrobial Resistance and Environmental Persistence: A Review. Pathogens 2020, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Félix, B.; Sevellec, Y.; Palma, F.; Douarre, P.E.; Felten, A.; Radomski, N.; Mallet, L.; Blanchard, Y.; Leroux, A.; Soumet, C. A European-Wide Dataset to Uncover Adaptive Traits of Listeria Monocytogenes to Diverse Ecological Niches. Sci. Data 2022, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Acciari, V.A.; Ruolo, A.; Torresi, M.; Ricci, L.; Pompei, A.; Marfoglia, C.; Valente, F.M.; Centorotola, G.; Conte, A.; Salini, R. Genetic Diversity of Listeria Monocytogenes Strains Contaminating Food and Food Producing Environment as Single Based Sample in Italy (Retrospective Study). Int. J. Food Microbiol. 2022, 366, 109562. [Google Scholar] [CrossRef]
- Osek, J.; Lachtara, B.; Wieczorek, K. Listeria Monocytogenes in Foods—From Culture Identification to Whole-genome Characteristics. Food Sci. Nutr. 2022, 10, 2825–2854. [Google Scholar] [CrossRef]
- Gomes-Neves, E.; Abrantes, A.C.; Vieira-Pinto, M.; Müller, A. Wild Game Meat—A Microbiological Safety and Hygiene Challenge? Curr. Clin. Microbiol. Rep. 2021, 8, 31–39. [Google Scholar] [CrossRef]
- Obwegeser, T.; Stephan, R.; Hofer, E.; Zweifel, C. Shedding of Foodborne Pathogens and Microbial Carcass Contamination of Hunted Wild Ruminants. Vet. Microbiol. 2012, 159, 149–154. [Google Scholar] [CrossRef]
- Paulsen, P.; Winkelmayer, R. Seasonal Variation in the Microbial Contamination of Game Carcasses in an Austrian Hunting Area. Eur. J. Wildl. Res. 2004, 50, 157–159. [Google Scholar] [CrossRef]
- Sasaki, Y.; Goshima, T.; Mori, T.; Murakami, M.; Haruna, M.; Ito, K.; Yamada, Y. Prevalence and Antimicrobial Susceptibility of Foodborne Bacteria in Wild Boars (Sus Scrofa) and Wild Deer (Cervus Nippon) in Japan. Foodborne Pathog. Dis. 2013, 10, 985–991. [Google Scholar] [CrossRef]
- Palacios-Gorba, C.; Moura, A.; Leclercq, A.; Gómez-Martín, Á.; Gomis, J.; Jiménez-Trigos, E.; Mocé, M.L.; Lecuit, M.; Quereda, J.J. Listeria spp. Isolated from Tonsils of Wild Deer and Boars: Genomic Characterization. Appl. Environ. Microbiol. 2021, 87, e02651-20. [Google Scholar] [CrossRef] [PubMed]
- Stella, S.; Tirloni, E.; Castelli, E.; Colombo, F.; Bernardi, C. Microbiological Evaluation of Carcasses of Wild Boar Hunted in a Hill Area of Northern Italy. J. Food Prot. 2018, 81, 1519–1525. [Google Scholar] [CrossRef]
- Wacheck, S.; Fredriksson-Ahomaa, M.; König, M.; Stolle, A.; Stephan, R. Wild Boars as an Important Reservoir for Foodborne Pathogens. Foodborne Pathog. Dis. 2010, 7, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Weindl, L.; Frank, E.; Ullrich, U.; Heurich, M.; Kleta, S.; Ellerbroek, L.; Gareis, M. Listeria Monocytogenes in Different Specimens from Healthy Red Deer and Wild Boars. Foodborne Pathog. Dis. 2016, 13, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, P.; Smulders, F.; Kukleci, E.; Tichy, A. Bacterial Surface Counts and Visually Assessed Cleanliness of Carcasses from Hunted Roe Deer (Capreolus Capreolus). Wien. Tierarztl. Mon. 2022, 109, doc3. [Google Scholar]
- Sauvala, M.; Laaksonen, S.; Laukkanen-Ninios, R.; Jalava, K.; Stephan, R.; Fredriksson-Ahomaa, M. Microbial Contamination of Moose (Alces Alces) and White-Tailed Deer (Odocoileus Virginianus) Carcasses Harvested by Hunters. Food Microbiol. 2019, 78, 82–88. [Google Scholar] [CrossRef]
- Alía, A.; Andrade, M.J.; Córdoba, J.J.; Martín, I.; Rodríguez, A. Development of a Multiplex Real-Time PCR to Differentiate the Four Major Listeria Monocytogenes Serotypes in Isolates from Meat Processing Plants. Food Microbiol. 2020, 87, 103367. [Google Scholar] [CrossRef]
- Li, F.; Ye, Q.; Chen, M.; Zhou, B.; Xiang, X.; Wang, C.; Shang, Y.; Zhang, J.; Pang, R.; Wang, J. Mining of Novel Target Genes through Pan-Genome Analysis for Multiplex PCR Differentiation of the Major Listeria Monocytogenes Serotypes. Int. J. Food Microbiol. 2021, 339, 109026. [Google Scholar] [CrossRef] [PubMed]
- Hyden, P.; Pietzka, A.; Lennkh, A.; Murer, A.; Springer, B.; Blaschitz, M.; Indra, A.; Huhulescu, S.; Allerberger, F.; Ruppitsch, W. Whole Genome Sequence-Based Serogrouping of Listeria Monocytogenes Isolates. J. Biotechnol. 2016, 235, 181–186. [Google Scholar] [CrossRef]
- Lachtara, B.; Wieczorek, K.; Osek, J. Genetic Diversity and Relationships of Listeria Monocytogenes Serogroup IIa Isolated in Poland. Microorganisms 2022, 10, 532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, W.; Wang, J.; Xu, B.; Liu, H.; Dong, Q.; Zhang, X. 10-Year Molecular Surveillance of Listeria Monocytogenes Using Whole-Genome Sequencing in Shanghai, China, 2009–2019. Front. Microbiol. 2020, 11, 551020. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Idland, L.; Heir, E.; Møretrø, T.; Aspholm, M.; Lindbäck, T.; Langsrud, S. Whole-Genome Sequencing Analysis of Listeria Monocytogenes from Rural, Urban, and Farm Environments in Norway: Genetic Diversity, Persistence, and Relation to Clinical and Food Isolates. Appl. Environ. Microbiol. 2022, 88, e02136-e21. [Google Scholar] [CrossRef] [PubMed]
- Sauvala, M.; Woivalin, E.; Kivistö, R.; Laukkanen-Ninios, R.; Laaksonen, S.; Stephan, R.; Fredriksson-Ahomaa, M. Hunted Game Birds–Carriers of Foodborne Pathogens. Food Microbiol. 2021, 98, 103768. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson-Ahomaa, M.; London, L.; Skrzypczak, T.; Kantala, T.; Laamanen, I.; Biström, M.; Maunula, L.; Gadd, T. Foodborne Zoonoses Common in Hunted Wild Boars. EcoHealth 2020, 17, 512–522. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of Methods for Genomic Taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef]
- Ragon, M.; Wirth, T.; Hollandt, F.; Lavenir, R.; Lecuit, M.; Le Monnier, A.; Brisse, S. A New Perspective on Listeria Monocytogenes Evolution. PLoS Pathog. 2008, 4, e1000146. [Google Scholar] [CrossRef]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V. Whole Genome-Based Population Biology and Epidemiological Surveillance of Listeria Monocytogenes. Nat. Microbiol. 2016, 2, 16185. [Google Scholar] [CrossRef]
- Ruppitsch, W.; Pietzka, A.; Prior, K.; Bletz, S.; Fernandez, H.L.; Allerberger, F.; Harmsen, D.; Mellmann, A. Defining and Evaluating a Core Genome Multilocus Sequence Typing Scheme for Whole-Genome Sequence-Based Typing of Listeria Monocytogenes. J. Clin. Microbiol. 2015, 53, 2869–2876. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A General Classification Scheme for Bacterial Virulence Factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Terentjeva, M.; Šteingolde, Ž.; Meistere, I.; Elferts, D.; Avsejenko, J.; Streikiša, M.; Gradovska, S.; Alksne, L.; Ķibilds, J.; Bērziņš, A. Prevalence, Genetic Diversity and Factors Associated with Distribution of Listeria Monocytogenes and Other Listeria spp. in Cattle Farms in Latvia. Pathogens 2021, 10, 851. [Google Scholar] [CrossRef] [PubMed]
- Filipello, V.; Mughini-Gras, L.; Gallina, S.; Vitale, N.; Mannelli, A.; Pontello, M.; Decastelli, L.; Allard, M.W.; Brown, E.W.; Lomonaco, S. Attribution of Listeria Monocytogenes Human Infections to Food and Animal Sources in Northern Italy. Food Microbiol. 2020, 89, 103433. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Langsrud, S.; Møretrø, T. Microbial Diversity and Ecology of Biofilms in Food Industry Environments Associated with Listeria Monocytogenes Persistence. Curr. Opin. Food Sci. 2021, 37, 171–178. [Google Scholar] [CrossRef]
- Parra-Flores, J.; Holý, O.; Bustamante, F.; Lepuschitz, S.; Pietzka, A.; Contreras-Fernández, A.; Castillo, C.; Ovalle, C.; Alarcón-Lavín, M.P.; Cruz-Córdova, A. Virulence and Antibiotic Resistance Genes in Listeria Monocytogenes Strains Isolated from Ready-to-Eat Foods in Chile. Front. Microbiol. 2021, 12, 796040. [Google Scholar] [CrossRef]
- Disson, O.; Moura, A.; Lecuit, M. Making Sense of the Biodiversity and Virulence of Listeria Monocytogenes. Trends Microbiol. 2021, 29, 811–822. [Google Scholar] [CrossRef]
- Gorski, L.; Cooley, M.B.; Oryang, D.; Carychao, D.; Nguyen, K.; Luo, Y.; Weinstein, L.; Brown, E.; Allard, M.; Mandrell, R.E. Prevalence and Clonal Diversity of over 1,200 Listeria Monocytogenes Isolates Collected from Public Access Waters near Produce Production Areas on the Central California Coast during 2011 to 2016. Appl. Environ. Microbiol. 2022, 88, e00357-22. [Google Scholar] [CrossRef]
- Castro, H.; Douillard, F.P.; Korkeala, H.; Lindström, M. Mobile Elements Harboring Heavy Metal and Bacitracin Resistance Genes Are Common among Listeria Monocytogenes Strains Persisting on Dairy Farms. Msphere 2021, 6, e00383-21. [Google Scholar] [CrossRef]
- Farber, J.M.; Zwietering, M.; Wiedmann, M.; Schaffner, D.; Hedberg, C.W.; Harrison, M.A.; Hartnett, E.; Chapman, B.; Donnelly, C.W.; Goodburn, K.E. Alternative Approaches to the Risk Management of Listeria Monocytogenes in Low Risk Foods. Food Control 2021, 123, 107601. [Google Scholar] [CrossRef]
- Chen, Y.; Gonzalez-Escalona, N.; Hammack, T.S.; Allard, M.W.; Strain, E.A.; Brown, E.W. Core Genome Multilocus Sequence Typing for Identification of Globally Distributed Clonal Groups and Differentiation of Outbreak Strains of Listeria Monocytogenes. Appl. Environ. Microbiol. 2016, 82, 6258–6272. [Google Scholar] [CrossRef]
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria Pathogenesis and Molecular Virulence Determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef]
- Schiavano, G.F.; Ateba, C.N.; Petruzzelli, A.; Mele, V.; Amagliani, G.; Guidi, F.; De Santi, M.; Pomilio, F.; Blasi, G.; Gattuso, A. Whole-Genome Sequencing Characterization of Virulence Profiles of Listeria Monocytogenes Food and Human Isolates and in Vitro Adhesion/Invasion Assessment. Microorganisms 2021, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Hanes, R.M.; Huang, Z. Investigation of Antimicrobial Resistance Genes in Listeria Monocytogenes from 2010 through to 2021. Int. J. Environ. Res. Public Health 2022, 19, 5506. [Google Scholar] [CrossRef] [PubMed]
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Abu Ghoush, M.H.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of Antibiotic Resistance in Listeria Monocytogenes Isolated from Food Products: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277–1292. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
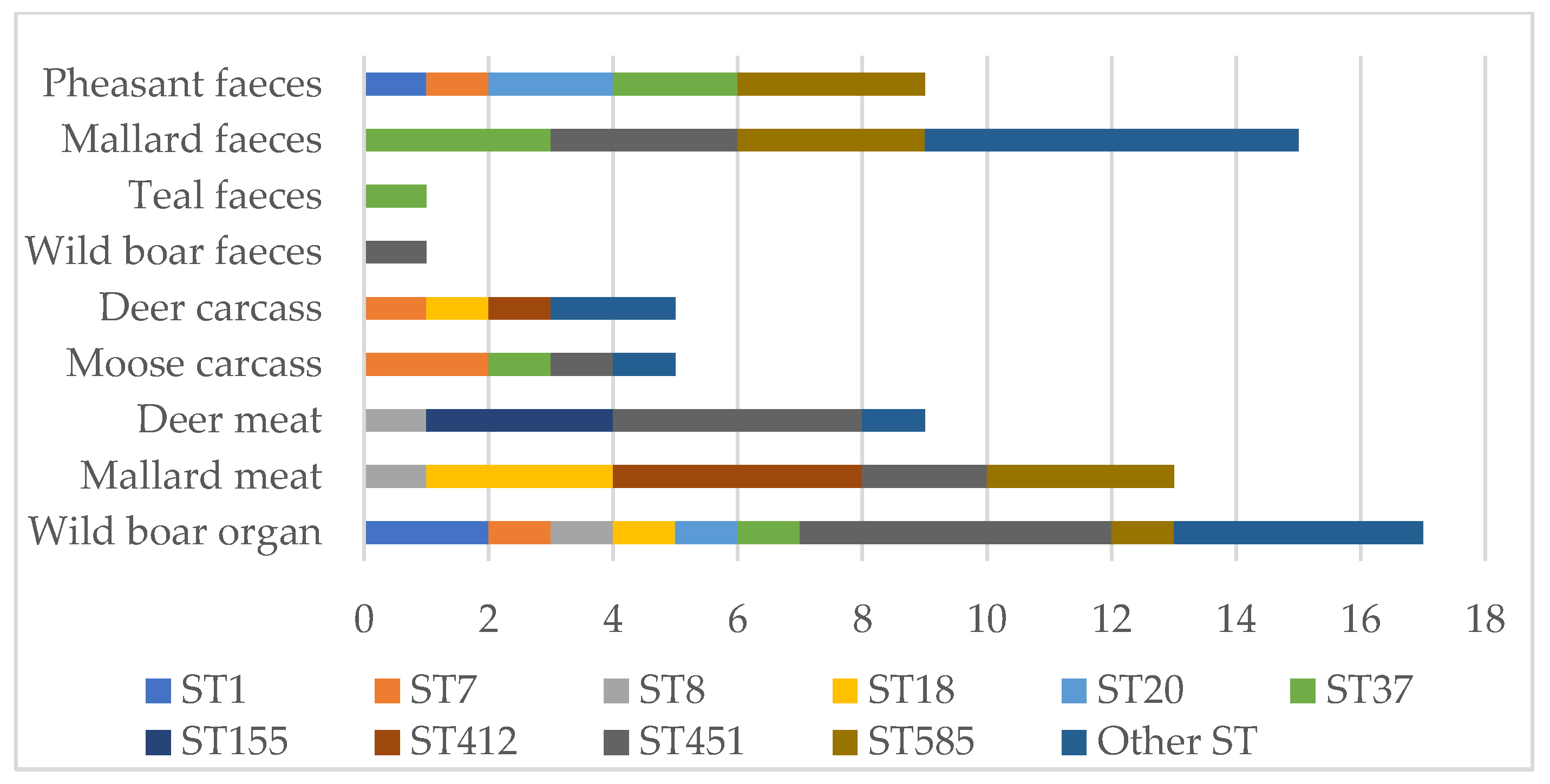
| Scheme 2012 | Sampling Year | Number of Samples | Positive Samples | Reference | |
|---|---|---|---|---|---|
| Moose carcass | 2012–2014 | 100 | 5 | (5%) | [17] |
| Deer carcass | 2013–2015 | 100 | 5 | (5%) | [17] |
| Deer meat | 2019–2020 | 50 | 9 | (18%) | Not published |
| Wild boar organ | 2016 | 130 | 40 | (31%) | [25] |
| Pheasant faeces | 2013–2014 | 101 | 9 | (9%) | [24] |
| Teal faeces | 2013 | 30 | 1 | (3%) | [24] |
| Mallard faeces | 2013–2014 | 110 | 15 | (14%) | [24] |
| Mallard meat | 2016 | 100 | 13 | (13%) | [24] |
| Source | Isolation Year | Number of Isolates | Serotype | MLST |
|---|---|---|---|---|
| Moose carcass | 2012–2013 | 5 | 1/2a | ST7, 29, 37, 451 |
| Deer carcass | 2013–2014 | 5 | 1/2a (2), 4b (3) | ST4, 18, 315, 412 |
| Deer meat | 2019 | 9 | 1/2a | ST8, 11, 155 |
| Mallard faeces | 2013–2014 | 15 | 1/2a (13), 1/2b (2) | ST11, 37, 224, 391, 585 |
| Mallard meat | 2016 | 13 | 1/2a | ST8, 18, 412, 451, 585 |
| Teal faeces | 2013 | 1 | 1/2a | ST37 |
| Pheasant faeces | 2013–2014 | 9 | 1/2a (8), 4b (1) | ST1, 7, 20, 37, 585 |
| Wild boar faeces | 2020 | 1 | 1/2a | ST451 |
| Wild boar organ | 2016 | 17 | 1/2a (15), 4b (2) | ST1, 7, 8, 18, 20, 21, 37, 91, 399, 451, 573 |
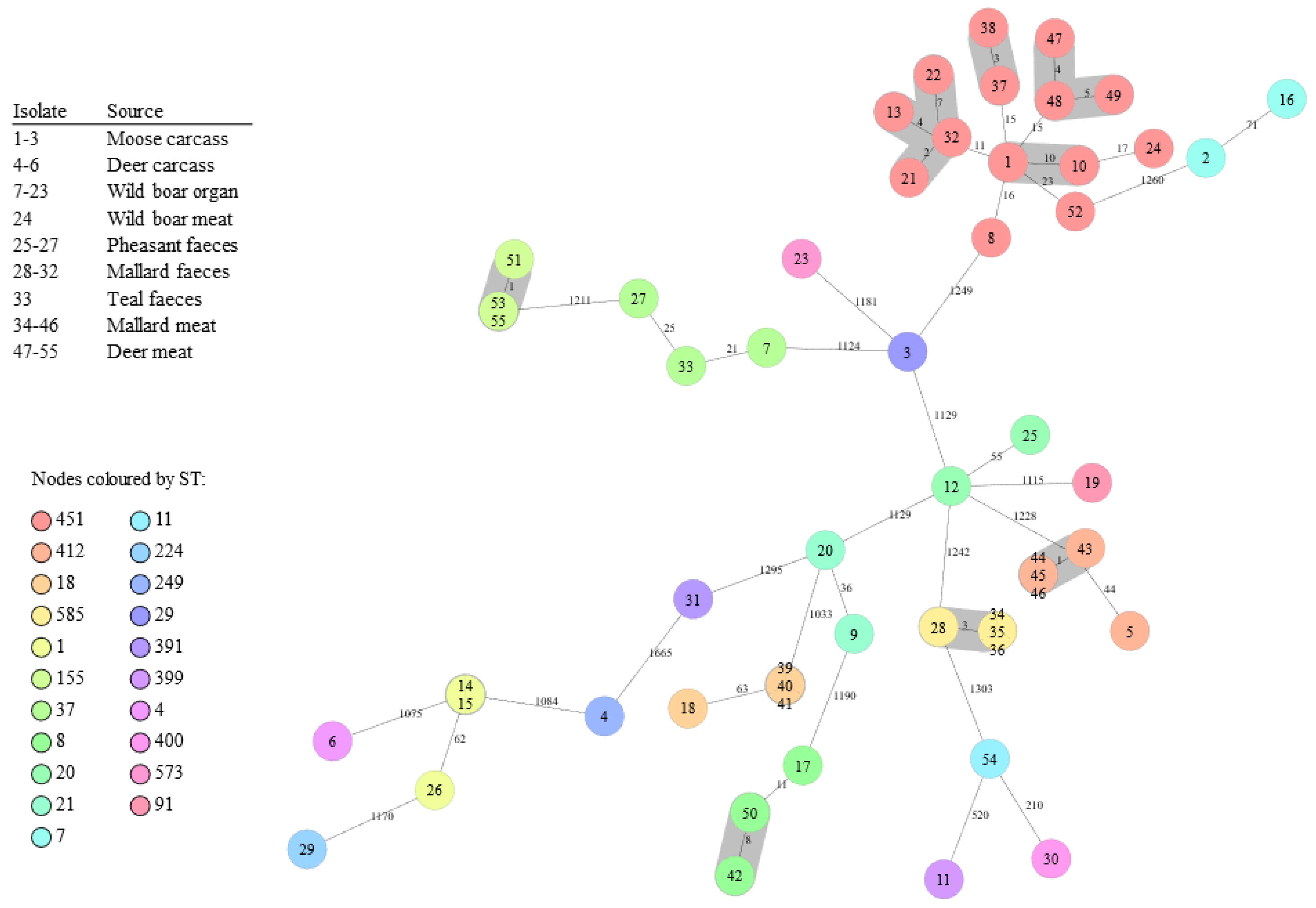
| MLST | Finland b | Clonal Complex | Lineage | Serotype | Number of Isolates | Source (Isolation Year) |
|---|---|---|---|---|---|---|
| ST1a | 2016, 2017 | CC1 | I | 4b | 3 | Pheasant faeces (2013), Wild boar organ (2016) |
| ST4 | CC4 | I | 4b | 1 | Deer carcass (2014) | |
| ST7 | 2018–2021 | CC7 | II | 1/2a | 4 | Moose carcass (2012), Pheasant faeces (2013), Wild boar organ (2016) |
| ST8 | 2017–2018, 2020–2021 | CC8 | II | 1/2a | 3 | Mallard meat (2016), Wild boar organ (2016), Deer meat (2019) |
| ST11 | CC11 | II | 1/2a | 1 | Deer meat (2019) | |
| ST18 | 2016 | CC18 | II | 1/2a | 5 | Deer carcass (2013), Wild boar organ (2016), Mallard meat (2019) |
| ST20 | CC20 | II | 1/2a | 3 | Pheasant faeces (2013), Wild boar organ (2016) | |
| ST21 | CC21 | II | 1/2a | 2 | Wild boar organ (2016) | |
| ST29 | CC29 | II | 1/2a | 1 | Moose carcass (2013) | |
| ST37 | 2016, 2018–2020 | CC37 | II | 1/2a | 8 | Moose carcass (2013), Teal faeces (2013), Mallard faeces (2013–2014), Pheasant faeces (2013–2014) Wild boar organ (2016) |
| ST91 | 2021 | CC14 | II | 1/2a | 1 | Wild boar organ (2019) |
| ST155 | 2020 | CC155 | II | 1/2a | 3 | Deer meat (2019) |
| ST224 | CC224 | I | 1/2b | 2 | Mallard faeces (2013) | |
| ST249 | CC315 | I | 4b | 2 | Deer carcass (2013) | |
| ST391 | CC89 | II | 1/2a | 2 | Mallard faeces (2013) | |
| ST399 | CC14 | II | 1/2a | 1 | Wild boar organ (2016) | |
| ST400 | CC11 | II | 1/2a | 2 | Mallard faeces (2013) | |
| ST412 | CC412 | II | 1/2a | 5 | Deer carcass (2013), Mallard meat (2019) | |
| ST451 | 2017–2021 | CC11 | II | 1/2a | 16 | Moose carcass (2012), Mallard faeces (2013–2014), Mallard meat (2016), Wild boar organ (2016), Deer meat (2019), Wild boar faeces (2020) |
| ST573 | CC573 | II | 1/2a | 1 | Wild boar organ (2016) | |
| ST585 | ST585 | II | 1/2a | 9 | Pheasant faeces (2013–2014), Mallard faeces (2013–2014), Mallard meat (2016) |
| ST a | SG | CT b | CL c | VP d | No. | Source | Year | |
|---|---|---|---|---|---|---|---|---|
| 1 | IVb | 1430 | 2c | 1 | Pheasant | Faeces | 2013 | |
| 1 | IVb | 25365 | 1 | 2c | 2 | Wild boar | Organ | 2016 |
| 4 | IVb | 27292 | 1d | 1 | Deer | Carcass | 2014 | |
| 7 | IIa | 21218 | 2a | 1 | Wild boar | Organ | 2016 | |
| 7 | IIa | 22874 | 1e | 1 | Moose | Carcass | 2012 | |
| 8 | IIa | 19030 | 2a | 1 | Wild boar | Organ | 2016 | |
| 8 | IIa | 28125 | 8 | 1e | 2 | Mallard, Deer | Meat | 2016, 2019 |
| 11 | IIa | 30149 | 1b | 1 | Deer | Meat | 2019 | |
| 18 | IIa | 26674 | 18 | 1a | 3 | Mallard | Meat | 2016 |
| 18 | IIa | 28197 | 1a | 1 | Wild boar | Organ | 2016 | |
| 20 | IIa | 21358 | 1a | 1 | Wild boar | Organ | 2016 | |
| 20 | IIa | 26681 | 0 | 1 | Pheasant | Faeces | 2013 | |
| 21 | IIa | 9841 | 2a | 1 | Wild boar | Organ | 2016 | |
| 21 | IIa | 25363 | 2a | 1 | Wild boar | Organ | 2016 | |
| 29 | IIa | 23920 | 1e | 1 | Moose | Carcass | 2013 | |
| 37 | IIa | 22893 | 1e | 1 | Teal | Faeces | 2013 | |
| 37 | IIa | 28230 | 1e | 1 | Pheasant | Faeces | 2014 | |
| 37 | IIa | 30787 | 2a | 1 | Wild boar | Organ | 2016 | |
| 91 | IIa | 20232 | 1a | 1 | Wild boar | Organ | 2016 | |
| 155 | IIa | 26763 | 155 | 0 | 3 | Deer | Meat | 2019 |
| 224 | IIb | 8887 | 1c | 1 | Mallard | Faeces | 2013 | |
| 249 | IVb | 20958 | 1d | 1 | Deer | Carcass | 2013 | |
| 391 | IIa | 29935 | 0 | 1 | Mallard | Meat | 2013 | |
| 399 | IIa | 22031 | 2b | 1 | Wild boar | Organ | 2016 | |
| 400 | IIa | 28173 | 1b | 1 | Mallard | Meat | 2013 | |
| 412 | IIa | 8287 | 3b | 1 | Deer | Carcass | 2013 | |
| 412 | IIa | 20896 | 412 | 2d | 4 | Mallard | Meat | 2016 |
| 451 | IIa | 24184 | 0 | 1 | Deer | Meat | 2019 | |
| 451 | IIa | 11793 | 0 | 1 | Wild boar | Faeces | 2020 | |
| 451 | IIa | 5208 | 451a | 1a | 3 | Mallard, wild boar | Meat, organ | 2016 |
| 451 | IIa | 11797 | 451b | 0,1a | 2 | Moose, wild boar | Carcass, organ | 2012, 2016 |
| 451 | IIa | 20939 | 451c | 0 | 3 | Deer | Meat | 2019 |
| 451 | IIa | 28250 | 451d | 0,1a | 4 | Mallard, wild boar | Faeces, organ | 2014, 2016 |
| 573 | IIa | 1569 | 2a | 1 | Wild boar | Faeces | 2016 | |
| 585 | IIa | 28251 | 585 | 1e,2a,3a | 4 | Mallard | Faeces, meat | 2013, 2016 |
| Virulence Profile | Number of Isolates | Sequence Types | Missing Virulence Genes (=1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| act | ami | aut | inlF | inlJ | lapB | vip | |||
| VP0 | 12 | ST20,155,391,451 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| VP1a | 13 | ST18,20,91,451 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| VP1b | 2 | ST11,400 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| VP1c | 1 | ST224 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| VP1d | 2 | ST4,249 a | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| VP1e | 7 | ST7,8,29,37, 585 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| VP2a | 7 | ST7,8,21,37,573,585 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| VP2b | 1 | ST399 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| VP2c | 3 | ST1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 |
| VP2d | 4 | ST412 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| VP3a | 2 | ST585 | 0 | 1 | 0 | 0 | 1 | 0 | 1 |
| VP3b | 1 | ST412 | 1 | 0 | 0 | 0 | 0 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fredriksson-Ahomaa, M.; Sauvala, M.; Kurittu, P.; Heljanko, V.; Heikinheimo, A.; Paulsen, P. Characterisation of Listeria monocytogenes Isolates from Hunted Game and Game Meat from Finland. Foods 2022, 11, 3679. https://doi.org/10.3390/foods11223679
Fredriksson-Ahomaa M, Sauvala M, Kurittu P, Heljanko V, Heikinheimo A, Paulsen P. Characterisation of Listeria monocytogenes Isolates from Hunted Game and Game Meat from Finland. Foods. 2022; 11(22):3679. https://doi.org/10.3390/foods11223679
Chicago/Turabian StyleFredriksson-Ahomaa, Maria, Mikaela Sauvala, Paula Kurittu, Viivi Heljanko, Annamari Heikinheimo, and Peter Paulsen. 2022. "Characterisation of Listeria monocytogenes Isolates from Hunted Game and Game Meat from Finland" Foods 11, no. 22: 3679. https://doi.org/10.3390/foods11223679
APA StyleFredriksson-Ahomaa, M., Sauvala, M., Kurittu, P., Heljanko, V., Heikinheimo, A., & Paulsen, P. (2022). Characterisation of Listeria monocytogenes Isolates from Hunted Game and Game Meat from Finland. Foods, 11(22), 3679. https://doi.org/10.3390/foods11223679