Abstract
Members of the Lamiaceae family are considered chief sources of bioactive therapeutic agents. They are important ornamental, medicinal, and aromatic plants, many of which are used in traditional and modern medicine and in the food, cosmetic, and pharmaceutical industries. In North Africa, on the Mediterranean side, there is the following particularly interesting Lamiaceous species: Thymus hirtus Willd. sp. Algeriensis Boiss. Et Reut. The populations of this endemic plant are distributed from the subhumid to the lower arid zone and are mainly employed as ethnomedicinal remedies in the following Maghreb countries: Algeria, Libya, Morocco, and Tunisia. In fact, they have been applied as antimicrobial agents, antispasmodics, astringents, expectorants, and preservatives for several food products. The species is commonly consumed as a tea or infusion and is used against hypercholesterolemia, diabetes, respiratory ailments, heart disease, and food poisoning. These medicinal uses are related to constituents with many biological characteristics, including antimicrobial, antioxidant, anticancer, anti-ulcer, anti-diabetic, insecticidal, and anti-inflammatory activities. This review aims to present an overview of the botanical characteristics and geographical distribution of Thymus algeriensis Boiss. Et Reut and its traditional uses. This manuscript also examines the phytochemical profile and its correlation with biological activities revealed by in vitro and in vivo studies.
1. Introduction
Lamiaceae (= Labiatae Adans., the mint family) is one of the groups of plants most widely distributed worldwide, with about 6900–7200 species [1,2]. It is a large family composed of annual, biennial, and perennial herbs, subshrubs, shrubs or trees, large forest forms, and even woody climbers [3]. It includes 242 genera with 200 or even more species, such as Salvia (986), Scutellaria (468), Stachys (375), and Plectranthus L’Hér. (325), Thymus (315), Hyptis Jacq. (295), Teucrium (287), Nepeta (251), and Vitex (223) [4], which are distributed in warm and temperate regions worldwide [5]. In addition, this family has great diversity with a cosmopolitan distribution [6]. They can adapt to different ecosystems and be cultivated easily, but without inhabiting the coldest high-altitude or high-latitude regions [7]. Worldwide, there are the following seven regions of high Lamiaceae diversity: (1) the Mediterranean and southwestern Central Asia; (2) Africa south of the Sahel and Madagascar; (3) China; (4) Australia; (5) South America; (6) North America and Mexico; (7) Indo-Malesia region (Southeast Asia) [3,8].
Lamiaceae is considered one of the most advanced plant families in floral structures [9]. They are plants with a quadrangular, usually fragrant stem, and their leaves are usually opposite without stipules [10]. Lamiaceae are plants that are usually fragrant and aromatic, especially Nepetoideae [3]. They are used in traditional and modern medicine, food, flavoring, perfumery, pesticide, and pharmaceutical industries [11,12,13,14,15]. Indeed, more than 150 plants containing essential oils, oleoresins (solvent-free), and natural extracts (including distillates) have been recognized by the US Food and Drug Administration as safe for human consumption, with no consumption limitations [16]. In this context, the Lamiaceae family deserves special mention as it is endowed with antioxidant [17], anti-inflammatory [18], hepatoprotective [19], antitumor [20], anti-mutagenic [21], radioprotective [22], antibacterial [23], antifungal [24], antiviral [25], and insecticidal properties [14], mainly due to its high content of phenolic compounds. Extracts of these plants, and their essential oils, are rich in a wide variety of secondary metabolites that have also shown potential properties for the treatment of depression [26,27] and pain [28], as well as diabetes [29,30] and cardiovascular disorders [11].
In Northwest Africa, more precisely in the greater Maghreb, represented by Algeria, Libya, Morocco, Mauritania, and Tunisia, the Lamiaceae are composed of a multitude of species distributed almost throughout the region [31,32]. The Maghreb is situated at the floristic crossroads between the Atlantic Ocean and the Mediterranean Sea and is separated from the Middle East by a broad band of Libyan and Egyptian deserts. Botanical influences from different directions could explain the Maghreban floristic biodiversity. Several regions of the Mediterranean climate are found, from desert to mountain, with transitions from plains to steppes [33,34]. The Maghreb has about 8000 km of coastline, bathed by the Atlantic Ocean to the west and the Mediterranean Sea to the north. The temperature and humidity differences between these two currents are the reason for vegetation and climate contrasts between the west and the north of the Maghreb. Boreal Europe lies to the north, Africa to the south, and Asia to the southeast. This particular geographical situation makes northern Africa one of the main routes for the dispersion of flora and fauna.
The Maghreb has a large and diversified heritage of aromatic and medicinal plants widely used in traditional medicine by local populations. This richness is also reflected in a wide culture of phytopharmacology, particularly among herbalists and folk healers. The Maghreb countries have a shared cultural heritage. Traditional, religious, and socio-economic associations mean that the way of life is comparable in all four territories. This similarity is reflected in the traditional uses of plants belonging to the Lamiaceae family. Indeed, its species are the most frequently cited in ethnopharmacological surveys, particularly in Algeria, Morocco, and Tunisia [33,34,35,36,37]. The Lamiaceae are represented by 29 genera in Algeria, 11 in Mauritania, 32 in Morocco, 22 in Libya, and in Tunisia by 26 genera [31,32]. In this review, we consider specifically the genus Thymus L., widespread in many territories in the Mediterranean regions, with much endemism [38,39,40,41,42] as those that grow in Northwest Africa and distributed on the whole coastline until the arid zones [31,43]. It is represented by 17 species and subspecies in Algeria, of which three are endemic [32,44]. These species are present from the north of Algiers to the Saharan Atlas and from the Constantine region to Oran [45]. Twenty-nine Moroccan species and subspecies, of which sixteen are endemic, are listed and found mainly in the plains or mountains, in rocky areas, scrublands, lawns, or scrub [32,46]. Only three Tunisian Thymus species grow at altitudes ranging from 120 to 1100 m in a subhumid or lower arid bioclimate [32,46,47]. Finally, in Mauritania, the Thymus genera do not exist [32,46].
According to taxonomic studies [31,43,46], twelve species and subspecies are distributed between Algeria and Morocco. In contrast, there are only three common species between Tunisia and Algeria and only one between the four countries, namely, Thymus algeriensis Boiss. and Reut. Many studies have reported that the by-products and compounds isolated from this species exhibit various biological activities, such as antimicrobial, anti-inflammatory, antioxidant, and cytotoxic [48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63]. In short, this review will help provide information on the traditional uses, phytochemistry, and pharmacology of T. algeriensis. These data will be valuable to discover gaps and explore potentials that require further research on the chemistry and health benefits of this species, and to generate more interest in the species T. algeriensis in the future.
2. Methodology
The updated literature was obtained from the following databases: PubMed, Web of Science, Elsevier, ACS Publications, Springer, NCBI, Wiley Online Library, google scholar, and other published documents (books and Ph.D. and MSc theses). The keywords used to search the literature sources include “Thymus”, “Thymus algeriensis”, and “Thymus hirtus algeriensis” in combination with Maghreb, Algeria, Libya, Morocco, Tunisia, “botanical description”, “phytochemical compounds”, “traditional uses”, and “pharmacological activities”. We collected all published works from the library from January 1900 to 30 June 2022. Only published data were included in this study, and all reports in all languages were included without exception.
Occurrence data were downloaded from the Global Biodiversity Information Facility (www.GBIF.org (accessed on 8 June 2022) GBIF Occurrence Download (https://doi.org/10.15468/dl.75vs2f (accessed on 8 June 2022), and distribution maps were taken from the site (https://africanplantdatabase.ch/ (accessed on 21 June 2022), whose data were confirmed with that of the Global Biodiversity Information Facility and that of the Maghreb eflora project website (https://efloramaghreb.org/ (accessed on 21 June 2022).
Species data were collected from Euro+Med PlantBase and the website (https://europlusmed.org/ (accessed on 17 June 2022). Scientific names and synonyms were validated by the Kew Plants of the World database (www.plantsoftheworldonline.org (accessed on 16 June 2022).
We used chemical structures of PubChem (https://pubchem.ncbi.nlm.nih.gov/ (accessed 10 July 2022) and SciFinder (www.scifinder.cas.org (accessed 15 July 2022).
3. Thymus Genera: An Overview
The word Thymus comes from the Greek “thyo”, which means “offering” (to be burnt) and “perfume” because of the pleasant smell that the plant gives off naturally when burnt [64,65]. In ancient times, the Sumerians and Egyptians used it for embalming their dead (the mummification process). The Romans burned thyme to purify the air and keep pests away [66,67]. The name “Thyme” comes from the Greek word “Thymos” [68], meaning smell. In the Azores, Madeira and the western part of the Iberian Peninsula, Thyme has the Portuguese names “tomentelo” or “tormentelo”, “tomelo do pais”, “tomentelo do pais” or “tomilho” [65]. In the mountains of Ethiopia, it is known as “rausi”. Moreover, the African northwest has the following several Arabic vernacular names: “Djertil”, “Hamzoucha”, “Mezouqach”, and “Khieta”, and Berber names such as “Azoukni”, “Tazuknite”, “Rebba”, “Djouchchen”, and “Touchna” [69]. In Morocco, it is also called “azukenni” [65].
The genus Thymus, described by Carl Linnaeus in Species Plantarum, belongs to the monophyletic group of the subfamily Nepetoideae Kostel, the tribe Mentheae Dumort and the subtribe Menthinae Endl [3,70]. The species can be sub-shrubs or shrubs, often herbaceous above, usually gynodioic and aromatic. They grow spontaneously on dry, rocky slopes and in scrubland. The stems of species are generally ± quadrangular, hairy all round, on two opposite sides, or only on the corners. The leaves are small, entire, and frequently revolute and form compact, highly branched clumps that rise to about 20 cm above the ground [45]. They have inflorescences of whorls forming a terminal, condensed, often spiciform or interrupted thyrse [46]. These characteristics can have a high degree of polymorphism, demonstrating the complexity of the genus Thymus from a taxonomic and systematic point of view. Indeed, this hybridization has been observed between species belonging to different sections and between species with varying ploidy levels, resulting in different chemical compositions [65,71]. Eight sections can be distinguished in the genus from a taxonomic and geographical point of view, as follows: Micantes, Mastichina, Piperella, Teucrioides, Pseudothymbra, Thymus, Hyphodromi, and Serpyllum. Five of them (sections Micantes, Mastichina, Piperella, Pseudothymbra, and Teucrioides) are endemic to the West Mediterranean area (Iberian Peninsula, Northwest Africa) [72].
4. Thymus algeriensis Boiss. and Reut.
4.1. Distribution
The medicinal properties of Thymus species have been investigated in numerous scientific studies, including in vitro et in vivo experiments. The results reveal that they have a unique combination of beneficial functions due to the Mediterranean climate [73,74,75]. In this geographical area, the western Mediterranean, under the influence of the Atlantic Ocean, the climatic conditions are favorable for thyme vegetation. Therefore, they are mainly found on the Mediterranean coast [76]. Thymus species are sun-loving, heliophilous plants, a fact that reflects the ecology of the genus. Thymus plants frequently live on rocks or stones, and the soil must be well-drained [70]. They need very different substrates. T. algeriensis usually lives on calcareous soils, characteristic of the Maghreb [77]. The Algerian species are found in the eastern Tell, in the bedrock areas, and on the high mountain plateaus up to the border of the pre-Sahara Tassili [43,78]. They are mainly present in subhumid and arid zones from the North-East of Algeria to the Tunisian border [45]; a few isolated individuals occur in Mascara province and from the Oran region to the Moroccan borders (Figure 1).
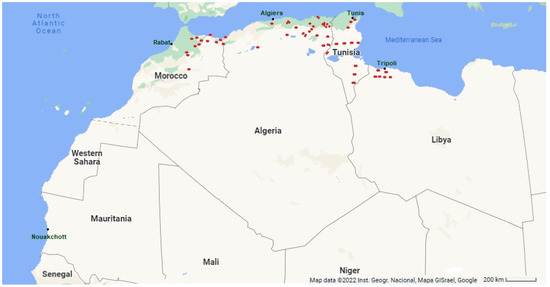
Figure 1.
In Red Thymus algeriensis Boiss. and Reut. Maghreb distribution (Coordinates N26° 14.526120 E5° 9.313440) [43].
The Moroccan T. algeriensis is found in the Mediterranean part of the country, in the Tangier-Tetouan-Al Hoceïma and Fez-Meknes regions, and more particularly in the so-called Oriental area (Béni Snassen forest), in the northeast near the Algerian border [43,78,79]. A few specimens have been found in the Middle Atlas, in the so-called Béni Mellal-Khénifra region (Figure 1). In Tunisia, the species is found in almost all bioclimates. It occurs in the north-eastern Mediterranean in the sub-humid zones of the Boukournine Mountains at 200 m altitude, up to the upper semi-arid zones at the foot of the Reças Mountains (Lead Mountain) at 150 m altitude and the Zaghouane Mountains at about 300 m altitude (Figure 1). Many populations of T. Algerians have been observed in the lower arid bioclimate. They formed a kind of vegetative belt from the city of Sfax to the Magel Bel Abbes near the Algerian border. In the southeastern part of the country, however, it occurs in the upper and lower Saharan bioclimate regions of Ksar Jedid and Remada [43,78]. In the Libyan zone, T. algeriensis only appears in the north-western part of the country (Figure 1), specifically in the Mediterranean coastal bioclimate and the coastal steppes climate in Qsar bin ghashir and Abu ar Rish. In the highlands, it has been detected in the Beni Walid, El Urban, Al Urqub, and Rahiba regions [43,78].
4.2. Systematic Classification and Botanical Aspects
According to Morales [65], T. algeriensis Boiss. and Reut. are classified in section Hyphodromi (A. Kerner) Haläcsy and subsection Subbracteati (Klokov) Jalas [80,81]. It belongs to the order Lamiale, subfamily Nepetoideae, and tribe Menthae (Figure 2).

Figure 2.
Systematic classification and botanical aspects of Thymus algeriensis Boiss. and Reut flowers and leaves. (a,b) from Tunisia, (c) from Algeria, (d) Systematic classification of T. algeriensis [82].
T. algeriensis is a subshrub that can reach a height of 50 cm. It is a short-lived diploid species (2n = 2x = 30), which is also aromatic, perennial, and gynodioic [80,83,84] It is characterized by small, dark green, opposite, lanceolate, short petiole leaves. It reproduces by seed through vegetative means. T. algeriensis is hermaphroditic (male fertile) or female (male sterile) [83]. Pollination is usually via bees (allogamous species). Self-pollination can also occur in hermaphrodite plants [85]. The vegetative stage occurs in January and February, and its flowering occurs between April and June [86]. Its flowers are small (5–6 mm) with a glandular calyx, oval-shaped bracteoles, and a pinkish-purple corolla.
4.3. Uses in Folk Medecine
In traditional Algerian medicine, T. algeriensis has been used as an astringent, expectorant, and healing agent and a blood circulation stimulant and aphrodisiac [87,88]. Infusion, decoction, and powder of the aerial parts are used in Naâma, southwest Algeria for treating colds as an anti-inflammatory, to manage hypercholesterolemia and menstrual cycle problems, and recently against COVID-19 [34]. El Kantara’s area (Algerian Sahara gate) is traditionally employed to flavor coffee, buttermilk, and tea. Infusing leaves and flowers are used against abdominal stomach pain, wound infections, and food poisoning. It is also antihypertensive and manages heart diseases [89]. In Morocco, T. algeriensis is a medicinal species indicated in the traditional treatment of diabetes [90]. It is a tonic stimulant against cough, fever, and wound infections [91,92]. It also treats asthma, bad breath, chest pain, lung disorders, and rhinosinusitis. In addition, some of his by-products were used as antitussives [33] and anti-inflammatory agents by topical or oral administration [93,94,95]. Traditionally in Tunisia, T. algeriensis is used as a culinary herb, fresh or dried [96], or as condiments or flavoring mainly added to black tea [35]. It is widely used in popular Tunisian medicine as a protective treatment against digestive tract diseases and abortion [97].
4.4. Phytochemistry
4.4.1. Essential oil Chemical Composition
The species belonging to the Lamiaceae family are reservoirs of molecules with high chemical diversity [12,98,99,100,101]. Many compounds have been identified to understand the relationship between their structures with biological activities and therapeutic potential. This significant chemical variability is mainly present in the species of the genus Thymus and also within the same species depending on the environmental, plant, and soil characteristics in which they grow [81,102,103]. These are several horticultural species, most of which are used as culinary herbs to flavor food. The flavoring comes from the fact that they produce essential oils with particular spicy aromas and flavors that are in demand by the cosmetic, nutrition, and health industries [104,105,106,107,108].
Essential oils are colorless and lipophilic liquids. They are odoriferous as a combination of volatile compounds derived from the secondary metabolism of plants [109]. They are mainly monoterpenes, sesquiterpenes, and diterpenes. Phenylpropanoids, fatty acids and their esters, alcohols, acids, epoxides, aldehydes, ketones, amines, and sulphides were also identified [110,111]. According to Venditti et al. [112], these compounds can be accompanied by polysaccharides and polyphenols.
Plants synthesize and emit a wide variety of volatile organic compounds that act as chemical signals, controlling their external environment. Stresses such as pathogen infections [113] or subterranean microbial communities [114] can induce the particular emission of volatile compounds from plants. They attract insect pollinators and repel predators [109]. Some of these signals are generated and emitted directly after herbivores damage plants. The volatile substances released often serve as indirect defenses, attracting insects and mites. The last attack or parasitize the herbivores and thus reduce further damage to the plant [115]. Additionally, essential oils inhibit seed germination and help plants communicate with each other. Indeed, several scientific reports have shown that plants can perceive other volatile substances emitted by nearby plants under attack by herbivores [116,117,118]. They respond to the information emitted by activating their defenses. In some cases, high expression of several genes involved in defense metabolism has been observed [119,120,121].
All species of Thymus produce essential oils [81]. They synthesize and store them in specialized anatomical structures called glandular trichomes [122]. These epidermal outgrowths contain special secretory cells heterogeneously distributed on all parts of the plant as follows: flowers, buds, seeds, leaves, twigs, bark, herbs, wood, fruits, and roots [123]. In the study of Guesmi et al. [86], scanning electron microscopy analysis demonstrated that T. algeriensis leaves contain glandular and non-glandular trichomes. Classifications can be based on the following two points: the cell number and the configuration of their walls in the head of the gland [124]. The glandular trichomes of T. algeriensis are of the peltate and capitate types. In peltate trichomes, the secretory material accumulates in the subcutaneous space and is finally released by rupturing the cuticle [125]. These structures are considered the sites of essential oil production [126]. In contrast, the secretory material is extruded through the cuticle in capitate trichomes [125]. They can produce essential oils and polysaccharides [126].
Numerous studies have been conducted on Thymus species to identify their chemical composition. The outcome is that they are rich in essential oils, characterized by a remarkable variability in chemical composition [127,128]. Oil chemotypes comprise monoterpenes, sesquiterpenes, and their oxygenated and hydrocarbon derivatives. An essential oil may contain one, two, or three of these main types of compounds and thus constitute a particular chemotype [129].
The structures of some (58) of the volatile and phenolic compounds identified in the essential oils of T. algeriensis are shown in Figure 3. GC-MS allowed the identification of an average of 43 volatile compounds in the Algerian T. algeriensis essential oils. They are mainly monoterpenes and sesquiterpenes responsible for the perfume [51,54,55,61,130,131,132,133,134,135,136,137]. Camphor, carvacrol acetate, elemol, linalool terpinene-4-ol, α-terpinyl acetate, thymol, α-pinene, β-myrcene, and γ-terpinene are the main constituents of Algerian essential oils, extracted by hydrodistillation, steam distillation, or by microwave (Table 1 and Figure 3). They are primarily found in the aerial parts of the plants. According to their location, there are three categories of essential oils. Based on major compounds, the populations of the northern region (Blida, Ain Defla, Medea) are characterized by the chemotypes linalool, terpinene-4-ol/camphor, terpinyl acetate/(trans)-nerolidol, thymol, and γ-terpinene/cymene [134,135,136,137]. The T. algeriensis of central regions (M’sila, Laghouat) contains camphor, camphor/borneol, carvacrol acetate/limonene, and α-terpinyl acetate as the main chemotypes [61,130,132,138]. Camphor, elemol/camphor, β-myrcene/camphor, germacrene D, and α-pinene are the chemotypes of the eastern region plants (Batna, Biskra, Guelma, Oum El Bouaghi, Soukahras) [51,53,54,55,131,139,140]. Monoterpenes such as 1,8-cineole, bornyl acetate, cis-sabinene hydrate, geranyl acetate, linalyl acetate, neryl acetate, perilla aldehyde, sabinene, trans-ocimene, trans-sabinene hydrate, trans-verbenol, α-terpinene, and α-terpineol have also been identified (Figure 3). Sesquiterpenes like the-nerolidol, 7-epi-α-eudesmol, acorenone, allo-aromadendrene, caryophyllene oxide, 5-neo-Cedranol, trans-caryophyllene, viridiflorol, α-cadinol, α-caryophyllene, β-caryophyllene, β-eudesmol, β-farnesene, and δ-cadinene are among the volatile oil components detected in Algerian oils (Figure 3).
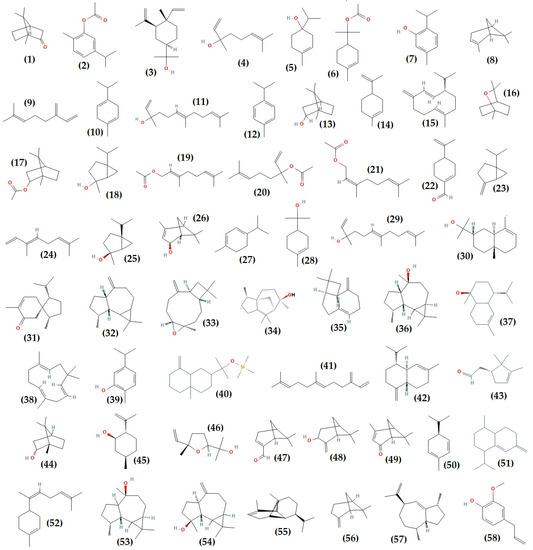
Figure 3.
Chemical structures of the phenolic and volatile compounds (1–58) identified from Thymus algeriensis Boiss. and Reut: camphor (1), carvacrol acetate (2), elemol (3), linalool (4), terpinene-4-ol (5), α-terpinyl acetate (6), thymol (7), α-pinene (8), β-myrcene (9), γ-terpinene (10), (trans)-nerolidol (11), cymene (12), borneol (13), limonene (14), germacrene d (15), 1,8-cineol (16), bornyl acetate (17), cis-sabinene hydrate (18), geranyl acetate (19), linalyl acetate (20), neryl acetate (21), perilla aldehyde (22), sabinene (23), trans-ocimene (24), trans-sabinene hydrate (25), trans-verbenol (26), α-terpinene (27), α-terpineol (28), (e)-nerolidol (29), 7-epi-α-eudesmol (30), acorenone (31), allo-aromadendrene (32), caryophyllene oxide (33), 5-neo-cedranol (34), trans-caryophyllene(35), viridiflorol (36), α-cadinol (37), α-caryophyllene (38), carvacrol (39), β-eudesmol (40), β-farnesene (41), δ-cadinene (42), campholenal (43), endo-borneol (44), iso-pulegol (45), linalool oxide (46), myrtenal (47), pinocarveol (48), verbenone (49), α-phellandrene (50), (+)-epi-bicyclosesquiphellandrene (51), cis-α-bisabolene (52), epiglobulol (53), spathulenol (54), α-copaene (55), β- pinene (56), γ-gurjunene (57), p-eugenol (58).
According to this research (Table 1), there is a clear chemical variability between specimens of T. algeriensis collected in the different Algerian regional groups. These include the North (bioclimate Mediterranean sub-humid) [134,135,136,137], the East (Mediterranean sub-humid, arid to super arid bioclimate) [51,53,54,55,131,139,140] and the Centre (super arid, sub-Saharan to Saharan bioclimate) [60,130,132]. It is even present between provinces belonging to the same bioclimatic group. The chemical composition of the essential oils of T. algeriensis collected in the Aurès region of Algeria is one example among many others. It is unique compared to other oils from the same region. Indeed, it is characterized by its high sesquiterpene content (67.0%). The main compounds of the oil were germacrene D (29.6%), β-caryophyllene (11.0%), and E-β-farnesene (7.8 According to Kebbi et al. [140], this is the first time that these compounds have been described with high content in T. algeriensis oil.
Furthermore, the essential oils of T. algeriensis showed different chemical profiles even for samples taken from the exact location at different levels. In the study by Hazzit et al. [135] the sample collected at 800 m altitude in Chrea National Park is characterized by thymol as the predominant component. In contrast, the sample taken at the same location at 1500 m altitude showed a predominance of terpinyl acetate/nerolidol/α-pinene/borneol/bornyl acetate [135]. Touhami et al. [141] also reported a difference in the chemical composition according to the developmental stage. The plants collected at the flowering stage had high proportions of oxygenated monoterpenes (77.56%) and oxygenated sesquiterpenes (5.98%). On the other hand, at the pre-flowering stage, their levels were lower (61.86%) and (2.10%), respectively. There was also a decrease in the monoterpenes (25.36%) and sesquiterpenes (10.68%) from the pre-blooming stage (16.46%) to the blooming stage (0.00%), respectively. According to Touhami et al. [141], these changes may be due to an increase in photosynthetic activity that induces a high rate of biosynthesis of volatile compounds (mainly phenolic compounds) at the complete flowering stage. Indeed, various studies have shown that the secretion of essential oils and their chemical composition in volatile compounds depends on several factors. They can be the development stage of the plant, its age, plant organ, species, seasonality, circadian rhythms, geographical location, and genetics [142,143,144,145,146]. Other factors related to the harvesting period [147], storage conditions [148], and extraction technique [149] also influence the nature of the essential oil.
Monoterpenes (1,8-cineole, camphor, cis-sabinene hydrate, linalool, terpinen-4-ol, terpinyl acetate, α-pinene), phenol-terpene thymol, and sesquiterpenes (caryophyllene oxide, viridiflorol) are the main compounds identified in the different Tunisian populations of T. algeriensis (Table 1). Some chemotypes are similar to those of the Algerian oils, such as camphor [49,143], linalool [48,85,150,151], thymol [85], and α-pinene [85,152]. Others are different such as 1,8-cineole, 1,8-cineole/camphor, 1,8-cineole/α-pinene, 4-terpineol/camphor, camphor/4-terpineol, caryophyllene oxide, cis-sabinene hydrate, cis-sabinene hydrate/1,8-cineole, eucalyptol, eucalyptol/2-carene, eucalyptol/viridiflorol, terpinen-4-ol, terpinyl acetate/1,8-cineole, viridiflorol, viridiflorol/cyclo-hexene,1-(1-butenyl), and viridiflorol/α-pinene [48,50,86,143,150,151,152,153].
Monoterpenes (2-carene, borneol, bornyl acetate, camphene, campholenal, cis-sabinene hydrate, endo-borneol, geraniol, iso-pulegol, linalool oxide, linalyl acetate, myrtenal, o-cymene, p-cymene, pinocarveol, sabinene, terpinyl acetate, verbenone, α-phellandrene, α-terpineol, α-terpinylacetate, β-ocimene, β-phellandrene, β-pinene, and γ-terpinene) have been identified in Tunisian populations of T. algeriensis (Figure 3). The (+)-epi-bicyclosesquiphellandrene, allo-aromadendrene, cis-α-bisabolene, elemol, epiglobulol, germacrene B, ledol, spathulenol, α-cadinol, α-copaene, α-humulene, β-caryophyllene, β-eudesmol, γ-cadinene, and γ-gurjunene are the sesquiterpenoid compounds that have also been detected in plants [48,50,86,143,150,151,152,153]. The phenolic compound carvacrol was only seen at 2.55% in the flowers and leaves of a species at the vegetative and flowering stage in the Oued Oum Ali region [143]. Furthermore, the phenylpropanoid compound, p-Eugenol (Figure 3), was only present in the leaves (tr-14.40%) and roots (tr-15.80%) of a population collected in Korbous, Jdidi Jebel Mountain, and Hammem Sousse [152].

Table 1.
Main components of the essential oils isolated from different Maghreb populations of Thymus algeriensis Boiss. and Reut (Algeria, Libya, Morocco, and Tunisia).
According to Table 1, a high chemical differentiation among populations was observed. In the study by Guesmi et al. [86], the different developmental phases greatly influenced the chemical composition of the essential oils of T. algeriensis. It was found that oils from leaves collected during the vegetative phase contained the highest level of volatile compounds and were at their lowest in older leaves collected in the post-flowering period. Monoterpene hydrocarbons and oxygenated monoterpenes decreased from the vegetative to the fruiting period. In contrast, the content of oxygenated sesquiterpenes was significantly lower during the vegetative stage.
In the studies by Zouari et al. [143], the upper parts of T. algeriensis were collected from different locations during the vegetative cycle. The results showed that 18 of the 71 compounds had a statistically significant variation between population locations and phenological stages. The chemical differentiation between the populations observed in the study was high. According to the authors, the distribution of observed chemotypes was related to population location and not to bioclimate, indicating that local selective environmental factors were responsible for the diversity of chemotypes [143].
In the study by Ben-El Hadj Ali et al. [152], qualitative and quantitative differences between different organs and samples were revealed. A large chemical variability was observed in collected plants and oil compound classes. The detection of distinct oil compounds was only in stems (cis-sabinene hydrate, β-gurjenene, and α-humulene), leaves (i.e., verbenone, γ-terpinene), and roots (geraniol and trans-α-bisabolene). According to the authors, this divergence could be due either to differential oil accumulation or physiological and biochemical interactions within and between organs during morphogenesis. It could also result from differential gene expression of organs and metabolic processes [85].
Nevertheless, this variability has been explained by geographical regions, local abiotic (topography, moisture, temperature, and edaphic factors), and selective biotic factors (associated fauna and flora) as well as genetic factors [152,159]. The latter was evaluated in eight Tunisian populations of T. algeriensis using 47 terpenoids and 154 RAPD (random amplified polymorphic DNA) markers amplified by seven selected primers. The populations were collected in different geographical regions of the sub-humid, upper semi-arid, middle semi-arid, lower semi-arid, and upper arid bioclimates. High genetic diversity within populations and high genetic differentiation between them, based on RAPDs, were revealed due to habitat fragmentation, the small size of most populations, and the low level of gene flow between them. Genetic and chemical structures are consistent with geographical distances indicating isolation by distance [85].
Moroccan essential oils are characterized by the chemotype camphor, carvacrol, borneol, thymol, and geranyl acetate (Table 1) [52,56,62,155,156,157,158]. The latter is present but in low levels (from 2.65 to 4.26%) only in the aerial parts (stems, flowers, and leaves) of the Algerian species from the El-Guetfa region (M’sila) and was extracted by hydrodistillation. It is also present in plant leaves but extracted by microwave distillation [61].
4.4.2. Phenolic Compounds
Plants produce an incredible variety of natural products with very different structures. These products are commonly referred to as “secondary metabolites” as opposed to “primary metabolites”, which are essential for plant growth and development [160]. They play a dynamic role in adapting plants to their environment [161]. In addition to their physiological function in plants, natural products also strongly impact human culture and have been used throughout history as condiments, pigments, and pharmaceuticals [7]. In recent years, rapid progress has been made in understanding the genomics and synthesis of natural products, their regulation and function, the evolution of metabolic diversity, and the investigation of the biological activities of secondary metabolites [162,163,164,165]. In addition, interest and sales of alternative therapies and medicines rich in phytochemicals have grown significantly and have been a fast-growing market in all parts of the industrialized world [166].
On the other hand, consumer demand for fresh herbs and natural products continuously increases, and plants from the Thymus genus have been considered relevant in this context [167,168]. Many species have been the subject of experimental studies confirming the effectiveness of some of their traditional applications [75]. In fact, they have well-established protective health benefits, mostly related to their highly complex phytochemistry [167,169,170,171]. Several groups and subgroups coexist within this genus, each with its own phytochemical characteristics and peculiarities. Some produce mainly lipids, volatile terpenoids (present in the essential oil), and those that are known to have principally non-volatile metabolites “phenolic compounds” in the polar fraction and are low in an essential oil [172]. Species of the genus Thymus are considered a high source of the latter compounds. They are characterized by many structures and functions but generally possess an aromatic ring bearing one or more hydroxyl substituents [173]. They are produced via the shikimic acid pathway and are usually involved in plant adaptation to environmental stress conditions [174]. There are different ways to classify them as they exist in many heterogeneous structures, ranging from single molecules to highly polymerized compounds. The secondary metabolite pattern of T. algeriensis comprises phenolic acids and flavonoids. The Figure 3, Figure 4, Figure 5 and Figure 6 show the chemical structures of these compounds identified in the different Algerian, Tunisian, and Moroccan species.
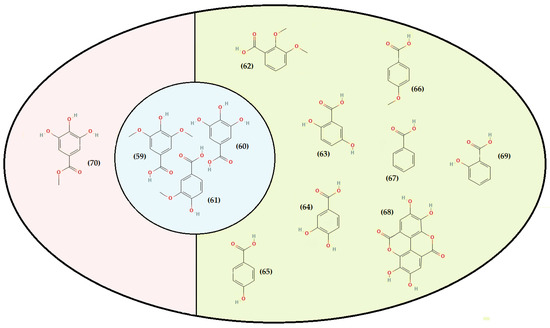
Figure 4.
Representation of the hydroxybenzoic acids’ structure identified in T. algeriensis. Green from Algeria, red from Tunisia, and blue common between the three countries (Algeria, Tunisia, and Morocco). Syringic acid (59), gallic acid (60), vanillic acid (61), 2,3-dimethoxybenzoic acid (62), 2,5 dihydroxybenzoic acid (63), 3,4-dihydroxybenzoic acid (64), 4-hydroxybenzoic acid (65), anisic acid (66), benzoic acid (67), ellagic acid (68), salicylic acid (69), and methyl galate (70).
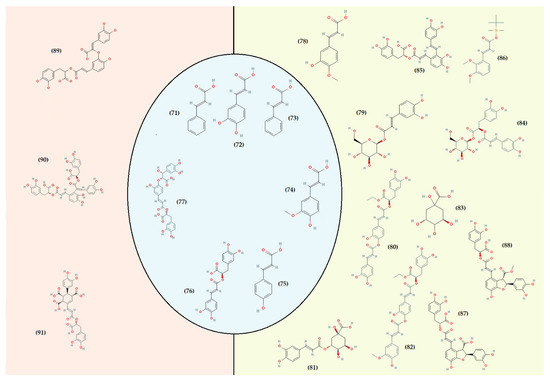
Figure 5.
Representation of the hydrocinnamic acid’s structure identified in T. algeriensis. Green from Algeria, red from Tunisia, and blue common between the three countries (Algeria, Tunisia, and Morocco). Caffeic acid (71), cinnamic acid (72), ferulic acid (73), p-coumaric acid (74), rosmarinic acid (75), salvianolic acid K (76), trans-cinnamic acid (77), 3-hydroxy-4-methoxycinnamic acid (78), caffeic acid glucoside (79), caffeoyl ethylrosmarinate (80), chlorogenic acid (81), feruloyl ethylrosmarinate (82), quinic acid (83), rosmarinic acid glucoside (84), salvianolic acid A (85), trans-2.3-dimethoxycinnamic acid (86), lithospermic acid A (87), monomethyl lithospermate (88), caffeoyl rosmarinic acid (89), salvianolic acid E (90), yunnaneic acid E (91).
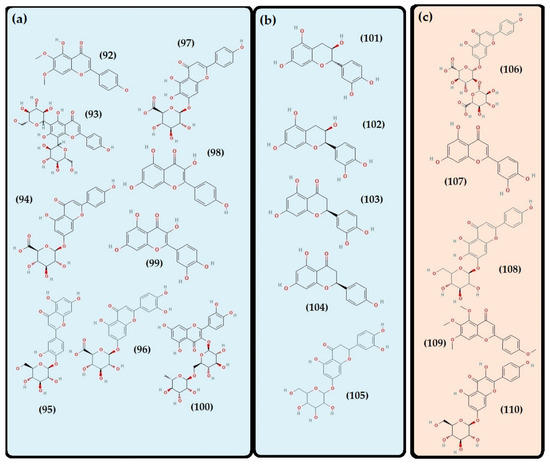
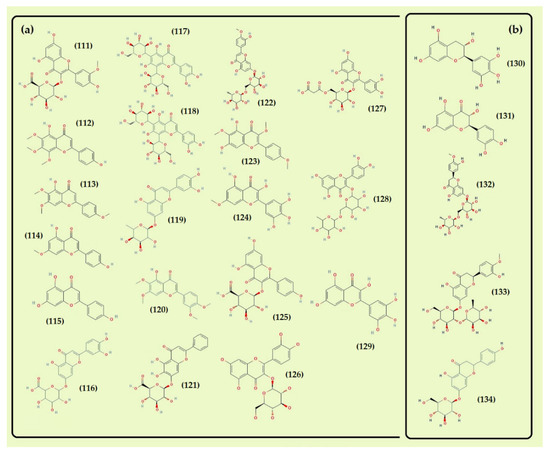
Figure 6.
Part I. Structure of the flavonoids detected in Thymus algeriensis from Maghreb countries. In blue, the flavonoids identified in common between the countries. (a) Flavone and flavonol compounds: cirsimaritin (92), apigenin 6,8-di-C-hexosides, (93) apigenin-7-O-glucuronide (94), apigenin-8-C-glucoside (95), luteolin glucuronide (96), scutellarin (97), kaempferol (98), quercetin (99), rutin (100). (b) Flavanol, flavanone, and flavanone glucoside: catechin (101), epicatechin (102), eriodictyol (103), naringenin (104), eriodictyol 7-O-glucoside (105). In red, flavonoids identified in Tunisian T. algeriensis. (c) Flavone and flavonol compounds: apigenin diglucuronide (106), luteolin (107), scutellarein-O-hexoside-hexuronide (108), tetramethyl-scutellarein (109), kaempferol-O-hexoside (110). Part II. In green, flavonoids identified in Algerian Thymus algeriensis. (a) Flavone and flavonol compounds: isorhamnetin pentosyl glucuronide (111), xanthomicrol (112), salvigenin (113), genkwanin (114), apigenin (115), luteolin feruloyl glucuronide (116), luteolin pentoside (117), luteolin pentosyl-glucoside (118), luteolin-7-O-rhamnoside (119), 5-desmethylsinensetin (120), baicalin (121), diosmin (122), santin (123), europetin (124), kaempferol-O-glucuronide (125), quercetin-3-β-D-glucoside (126), quercetin-O-malonyhexoside (127), quercetin-3-O-rutinoside (128), myricetin (129). (b) Flavanol, flavanonol, flavanone, and flavanone glucoside: gallocatechin (130), taxifolin (131), hesperidin (132), neohesperidin (133), naringenin-O-hexoside (134).
There is a wealth of published data for identifying and quantitatively determining phenolic constituents in Thymus plant extracts [175,176,177,178,179,180,181,182]. They have been widely investigated for their biological activities in physiological systems, e.g., antioxidant, anti-aging, anticarcinogenic, pro-apoptosis, anti-inflammation, and anti-atherosclerosis. In addition, they are effective in cardiovascular protection, improving endothelial function, and inhibiting angiogenesis and cell proliferation [75,127,128,171]. Furthermore, the content of these compounds in T. algeriensis plant extracts (Table 2) depends mainly on the chemotype and origin of the plant raw material, as well as the choice of solvent and extraction procedures [48,61,183,184]. Irrigation conditions, harvest time, storage conditions, and drying treatments are also factors that can affect the final phenolic composition of extracts and their respective biological activities [185]. Thus, many analytical procedures have been developed to quantify phenolic compounds in herbs [186,187].
Table 2.
Total phenolic, flavonoid, phenolic acids, flavonol, anthocyanin, and tannin content of Maghreb Thymus algeriensis Boiss. and Reut (Algeria, Morocco, and Tunisia) plant parts extract.
- Phenolic acids
A wide range of phenolic acids (benzoic and cinnamic acid series) was identified in T. algeriensis. They are distinguished from other phenols by their acidic character [192,193]. These compounds are involved in various functions related to plant physiology, including nutrient uptake, protein synthesis, enzymatic activity, photosynthesis, structural components, and allelopathy [194,195]. Phenolic acids are widely present in plant foods (e.g., fruits, vegetables, and cereals), which comprise a significant proportion of the human diet. They exist in association with other structural (cellulose, protein, lignin), polyphenolic or terpene plant components, and smaller organic molecules (e.g., glucose, quinic, maleic, or tartaric acids). They thus constitute the aglycone part of the compound [194,196].
Phenolic acids are, after flavonoids, the most investigated secondary plant metabolites due to their extensive presence in the diet and rapid metabolism in the human body [197]. Beyond their known protective antioxidant behavior attributed to several phenolic hydroxyl groups in their chemical structure, other biological activities of phenolic acids have been reported [197,198,199]. Therefore, they have received much attention in pharmaceutical and medicinal research as they seem to play a role in preventing several human diseases [200].
Depending on the constituent carbon frameworks, phenolic acids can be divided into the following two categories: benzoic acid derivatives (i.e., hydroxybenzoic acids) and cinnamic acid derivatives (i.e., hydroxycinnamic acids) [201]. They are characterized by a 6 and 9 carbon skeleton containing a carboxyl group attached to the benzene ring with one or more hydroxyl or methoxyl groups attached. The cinnamic acids, in addition, have an unsaturated propionic acid side chain attached to the benzene ring [202]. A summary of phenolic acids and derivatives identified in the different Algerian, Tunisian, and Moroccan species of T. algeriensis is presented in Table 3. In addition, their structure (from 59 to 90) is identified in each country and that in common is described in Figure 4 and Figure 5.
Most hydroxybenzoic acids include a C6-C1 backbone directly obtained from benzoic acid [199]. In the Algerian T. algeriensis (Figure 4), we found the presence of p-hydroxybenzoic acid, 2,5-dihydroxybenzoic acid, 3,4-dihydroxybenzoic acid, and 2,3-dimethoxybenzoic acid, which were not detected in the Tunisian species [58,132,184,203]. There are also anisic, ellagic, and salicylic acids [55,132,183,203]. Methyl gallate is only found in Tunisian populations (Figure 4) [48,61].
Furthermore, the content of hydroxybenzoic acids (gallic acid, syringic acid, and vanillic acid) in T. algeriensis populations generally varies between Algeria [132,184] and Tunisia [48,57]. However, this difference can also be related to the extraction method. The efficiency of an extraction technique depends on several critical parameters. In fact, the solvent, the nature of the material, the light, the duration of the extraction period, the pH, the temperature, the size of the material, the solvent/substrate ratio, and the liquid-liquid or solid-liquid ratio can influence the chemical composition [204]. In addition to selecting the optimal extraction method, the choice of a suitable technique for detecting and quantifying phenolic compounds is also of high relevance. There is a large amount of published research on this topic, but some analyses still have difficulties [205,206]. Despite this, there is great potential for developing specific methods [207,208].
Table 3.
Major phytochemicals isolated and characterized from Maghreb Thymus algeriensis Boiss. and Reut (Algeria, Morocco, and Tunisia).
Some of the hydroxybenzoic acids (Figure 4) detected can also be found in derived forms, such as amides and esters or glycosides [210]. They exhibit multiple physiological functions and high pharmacological potential, mainly attributed to the presence of multiple hydroxyl groups in their chemical structure, making them suitable free radical scavengers [197,211].
In contrast to the hydroxybenzoic acids, the group of hydroxycinnamic acids is the most important class of phenolic acids (Figure 5). They have a structure with three carbon side chains (C6–C3) originating from the phenylalanine and tyrosine pathways [199]. Their derivatives serve as precursor molecules for stilbenes, chalcones, flavonoids, lignans, and anthocyanins [212]. In Tunisian and Algerian populations of T. algeriensis, the common hydroxycinnamic acids detected and/or quantified are caffeic acid, cinnamic acid, ferulic acid, p-coumaric acid, rosmarinic acid, and salvianolic acid k [48,50,55,56,58,59,61,88,132,183,184,188,203,209,213].
Rosmarinic acid is the dominant caffeic acid derivative in the different extracts of T. algeriensis, a common compound in the Thymus plants (Table 3, Figure 5). It is more abundant in the infusion (58.20 ± 0.30 mg/g), decoction (54.40 ± 0.90 mg/g) and hydroethanolic extract (29.70 ± 0.70 mg/g) prepared from the Algerian species [183] than in the methanolic extracts of the Tunisian species (1157.80−383.80 µg/mL) [62]. According to Jaouadi et al. [62], high levels of rosmarinic acid are detected only in some Tunisian populations, notably those characterized by an upper semi-arid (1157 µg/mL extract), lower semi-arid (1083 µg/mL extract) and upper arid bioclimate (957 and 807.2 µg/mL). Rosmarinic hexoside acid (2.80−7.06 mg/g) is also found in some Algerian populations and rosmarinic caffeoyl acid (39.20–232.20 µg/mL) in Tunisian ones [61,183,188]. Caffeic acid was quantified at 52.79 ng/mL in the n-butanol extract by HPLC-TOF/MS [132] and at 33.3 µg/g by HPLC in the ethanolic extract of the Algerian species [203]. However, it was detected at concentrations of 26.00 ± 14.00 μg/g by HPLC in the methanolic extract of a Tunisian population [57]. Regarding ferulic acid, it is more dominant in the extracts prepared by microwaves (140.64 ± 0.73 µg/g) [184] than in the other preparations (hydro methanolic, ethanolic, n-butanol, and chloroform) [48,55,57,132,184]. The same observation was made for the compounds p-coumaric acid (106.99 ± 0.77 µg/g) and o-coumaric acid (341.55 ± 1.17 µg/g) [184]. Furthermore, HPLC did not detect cinnamic acid in the methanolic extract of the Tunisian species [48]. On the other hand, it was quantified at 20.51 µg/g in the ethanolic extracts of the Tunisian species by HPLC and 5.4% in the hydromethanolic extracts of the Algerian species by HPLC/UV [132]. Carvacrol, a diterpene phenol (Figure 3), was quantified between 48.76% and 76.03% by GC-MS in ethyl acetate and petroleum ether extracts of some Moroccan populations of T. algeriensis [56]. As for the Tunisian species, from 1374.70 ± 5.00 to 2221.60 ± 2.50 µg/mL were detected in the methanolic extracts by UHPLC-DAD-ESI/MSn [62]. On the other hand, HPLC-PDAs chloroform extract of the Algerian species revealed only 0.43 ± 0.01 µg/g of carvacrol [184].
Other phenolic acids in Algerian plants are 3-hydroxy-4-methoxycinnamic acid, trans-2,3-dimethoxycinnamic acid, trans-cinnamic acid, chlorogenic acid, feruloyl ethyl rosmarinate, quinic acid, yunnaneic acid E, lithospermic acid A (isomer I and II), monomethyl lithospermate, and salvianolic acid A [55,132,184,188,203,209]. Many of these substances are well investigated and have demonstrated multiple physiological functions, such as antioxidant, anti-inflammatory, and antimicrobial functions [197,198,214,215,216]. They play an important role in preventing and treating obesity, diabetes, and related disorders [217]. In addition, several studies have shown a relationship between their consumption and the risk of developing certain cancers [218,219].
- Flavonoids
Flavonoids are a large group of hydroxylated polyphenolics with a benzo-γ-pyrone structure found throughout plants. Their synthesis pathway is part of the broader phenylpropanoid pathway, which produces a range of other secondary metabolites, such as lignins, lignans, and stilbenes [220]. All flavonoids share a chemical structure based on the flavan system (C6-C3-C6). Plants synthesize them in response to various abiotic and biotic stresses [221,222]. They function as UV filters, signaling molecules, allelopathic agents, frost and drought protection agents, phytoalexins, detoxifiers, antimicrobials, and anti-herbivore factors [223,224]. Flavonoids are food components with health-promoting properties due to their antioxidant potential through their reducing capacities and/or possible influences on intracellular redox status [225]. These activities depend on their structural diversity. Flavonoid compounds differ structurally by their degree of hydroxylation, the presence of other substitutions and conjugations, and the degree of polymerization [226,227]. About 10,000 flavonoids have been identified, the third-largest group of natural products after alkaloids [228].
Considering the data present in Table 2 and Table 3, T. algeriensis appears to be a rich source of these compounds, containing large amounts of flavonoids. These include flavones, flavone glycosides, flavonols, flavanones, flavanone glycosides, flavanols, and flavanonols. The structure of each group of flavonoids (from 92 to 134) identified is represented in Figure 6. Qualitative and quantitative analysis of different extracts of T. algeriensis revealed several common chemical structures between the Algerian, Tunisian, and Moroccan populations. It also appears from the data in Table 2 and Table 3 that the Algerian species is more prosperous than the Tunisians [48,50,55,56,57,58,59,61,88,132,183,184,188,203,209]. However, the content of total flavonoids present in T. algeriensis can be affected by several factors, such as physiological, genetic, environmental, growth and storage conditions, etc., as well as the choice of solvent and extraction procedures [228,229,230,231,232]. Therefore, properly comparing the different phenolic compositions of Algerian, Moroccan, and Tunisian medicinal plants is difficult, namely, because the plant samples are of different territorial or regional origins. Moreover, the different standard methods for total flavonoid determination (by UV, IR, or NMR spectrophotometry) and the development of chromatographic methods (HPLC-DAD, HPLC-UV, HPLC-MS, LC-MS, LC-DAD, HPLC-PAD, LC-MS-MS, etc.) make it very difficult to carry out the procedure of comparing different results [99,233]. Various identification approaches allow the accumulation of a large amount of data on the composition of medicinal plant by-products. DNA markers are among the methods that can help solve this problem. In addition to morphological, anatomical, and chemical traits, they can differentiate authentic material and are thus an ideal approach for identifying medicinal plant species and populations/varieties of the same species [234].
Flavones are one of the main subgroups of flavonoids and are mainly dominant in the preparations of T. algeriensis (Table 3, Figure 6). Their structure has a ketone in the 4-position of the C-ring and a double bond between the 2- and 3-positions. Most of these compounds have a hydroxyl group in position 5 (Figure 6). The hydroxylation of position 7, A-ring or 3′ and 4′, B-ring can vary according to the taxonomic classification of the plants [224]. Cirsimaritin, apigenin, luteolin, and scutellarin were distinguished (Table 3). Apigenin (4′,5,7-trihydroxyflavone) is more concentrated in aqueous extracts (infusions and decoctions) than hydroethanolic or methanolic extracts. It is also more abundant in the Algerian populations than in the Tunisians [61,183,188].
Flavones are widely present in leaves, flowers, and fruits as glycosides [224]. Apigenin-C-dihexoside is detected in 20.70 ± 0.10 mg/g, 18.80 ± 0.10 mg/g, and 10.0 ± 0.50 mg/g, respectively, by LC-DAD-ESI/MS in the infusion, decoction, and hydroethanolic preparations of the Algerian species [183]. On the other hand, the Tunisian species showed 62.60–10.40 µg/mL of Apigenin-C-di-hexoside and 1.40–112.80 µg/mL of Apigenin-O-hexuronide in the methanolic extracts characterized by UHPLC-DAD-ESI/MSn [62]. The following other Apigenin derivatives were detected in T. algeriensis: Apigenin diglucuronide and Apigenin glucoside glucuronide in the Tunisian species [50] and Apigenin-8-C-glucoside in the Algerian species [183]. In contrast, luteolin and its derivatives (luteolin glucoside, luteolin feruloyl glucuronide, luteolin glucuronid, luteolin pentoside, luteolin pentosyl-glucoside, and luteolin-7-O-rhamnoside) have been reported in T. algeriensis plants (Algeria, Morocco, and Tunisia) [50,59,61,88,183,209]. The other flavone derivatives, isorhamnetin pentosyl glucuronide, xanthomicrol, salvigenin, genkwanin, and baicalin, are, however, only detected in the extracts of Algerian plants [55,58,203,209].
The aerial parts of T. algeriensis are also rich in flavonols and flavonol-glycosides based on kaempferol and quercetin skeletons (Table 3, Figure 6). They are the building blocks of proanthocyanins. In effect, anthocyanins and flavonols are derived from phenylalanine and share common precursors, dihydroflavonols, which are substrates for flavonol synthase and dihydroflavonol 4-reductase.
Flavonols have, compared to flavones, a hydroxyl group in position 3 of the C ring, which can also be glycosylated or methylated [224]. In T. algeriensis, methanolic extraction was more efficient in extracting kaempferol-O-glucuronide (3,4′,5,7-tetrahydroxyflavone) (862.80–202.90 µg/mL) from the Algerian species compared to the Tunisian ones. Indeed, the content of the flavonol-glycoside in infusion, decoction, and hydroethanolic preparations was (65.0 ± 0.40 mg/g), (62.20 ± 0.90 mg/g), and (16.7 ± 0.20 mg/g), respectively [61,183]. In contrast, quercetin (3,3′,4′,5,7-pentahydroxyflavone) is more abundantly obtained by microwave-assisted extraction (180.72 ± 0.77 µg/g) than by other techniques [55,57,132,184].
In Algerian plants, myricetin (3,5,7,3′,4′,5′-hexahydroxyflavone), diosmin (3′,5,7-trihydroxy-4′-methoxyflavone-7-rhamnoglucoside), and the two o-methyl flavonol compounds, europetin (7-o-methylmyricetin), and santin (5,7-dihydroxy-3,6,4′-trimethoxyflavone) are also identified [55,58,203].
Flavanones, also known as dihydroflavones, have a saturated C-ring because, unlike flavones, the double bond between the 2- and 3-positions is saturated [224]. The flavanone eriodictyol and its glycosyl form, eriodictyol-O-hexoside, have been detected in Tunisian T. algeriensis. They are more extensively expressed in the population under the medium semi-arid (4.10–42.00 µg/mL) and high arid (3.50–52.80 µg/mL) bioclimates, respectively [61,209].
The flavanols catechin (359.80 ± 1.98 µg/g) and epicatechin (2462.75 ± 2.00 µg/g) are more concentrated in the microwaved preparations than in the macerated ones [48,55,57,132,184,203]. Naringenin, however, was only detected in trace amounts in the Algerian species’ infusion, decoction, and hydroethanolic preparations. On the contrary, the chloroform extract (F16), contains a large amount (8.97 ± 0.74 µg/g) compared to the other chloroform fraction (F26) (0.90 ± 0.03 µg/g) and the n-butanol extract (0.47 ± 0.01 µg/g) [61,183,184]. The flavanol gallocatechin, flavanone glycosides (hesperidin, neohesperidin), and flavanonol Taxifolin were also identified in Algerian T. algeriensis [203,209].
Several studies using various in vitro and in vivo methodologies have suggested that flavonoids are the main chemical components responsible for the pharmacological activities of many medicinal plant by-products. Furthermore, it has been determined that flavonoids (60%) and phenolic acids (30%) represent the main phenolic compounds in our diet [235]. In the last decade, flavonoids’ biological, pharmacological, and medicinal properties have been widely reviewed [236,237,238,239]. These molecules have been attributed to positive effects on human and animal health, and the current interest is in disease therapy and chemoprevention [224]. Flavonoids possess antioxidant, vasculoprotective, anti-hepatotoxic, anti-inflammatory, anti-thrombotic, and anti-cholinesterase activities, and many other beneficial properties have also been investigated [224,240].
In addition, flavonoids are effective as anticancer and cardioprotective agents. These activities may be related to the abilities of these compounds to modify the activity of enzyme systems in mammals (kinases, phospholipases, ATPase, lipooxygenases, cyclooxygenases, phosphodiesterases, etc.), with a correlation being observed in some cases between the structure of the flavonoid and its enzymatic activity. Much of these effects can be attributed to the flavonoid’s ability to interact with the nucleotide-binding sites of regulatory enzymes. In addition, flavonoids can bind to Toll-like receptors in the plasma membrane and thus initiate the enzyme induction process [241]. Many protective enzymes have antioxidant functions and can scavenge oxygen and other free radicals [242]. The activity and selectivity of these compounds depend on structural factors (such as oxidation stage, substituents, and the presence of glycosylation), and the search for structural elements defining the beneficial interactions of these compounds has been explored. Using these compounds as molecular models to design selective and potent small molecule inhibitors may be a strategy to overcome the common problem encountered with the therapeutic efficacy of existing drugs [243].
- Other compounds
The organic acid profile (Table 3) revealed seven organic acids (12-hydroxyjasmonic acid 12-O-β-D-glucoside, schizotenuin F, malic acid, fumaric acid, phloretic acid, phloretic acid caffeoyl 3-hydroxy-3-methylglutaroyl, and quinic acid) in the Algerian T. algeriensis [203,209]. Six organic compounds were identified in the Tunisian species (4-hydroxyphenyl acetic acid, tri-hydroxyoctadecedienoic acid isomer, quinic acid, citric acid, 12-hydroxyjasmonic acid, 12-hydroxyjasmonic acid sulphate) [48,50]. Sagerinic acid, in the form of sagerinate, belongs to the cyclobutane lignans class and was also found in Algerian T. algeriensis. Furthermore, a phenolic aldehyde (isovanillin), a stilbenoid (polydatin), a pentacyclic triterpenoid (oleanolic acid), and phytosterols (β-sitosterol) have also been identified [57,58,184].
4.5. Pharmacological Reports
4.5.1. Antibacterial Effects
The antimicrobial potential is the most investigated therapeutic application of T. algeriensis by-products. The antibacterial activity of the essential oils and extracts of the different parts of the plant by the inhibition zone diameter (IZD mm) and/or minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) methods are summarized in Table 4.
Crude extracts and oils isolated from Algerian, Libyan, Moroccan, and Tunisian species possess antibacterial properties, especially against gram-positive bacteria. All species showed broad to moderate inhibition zones according to the well diffusion method results. Fatma et al. [158] investigated the antibacterial activity of essential oils from three different Tunisian regions against Staphylococcus aureus with Penicillin as a control for the inhibitory effect. All the essences showed broad zones of inhibition [Eo1 (22.00 mm), Eo2 (63.00 mm), and Eo3 (63.00 mm)] against the bacterium [48]. The same was observed for a Moroccan essential oil, which gave an IZD equal to 51.00 ± 3.40 mm. On the other hand, in the Ben El Hadj Ali et al. [152] study, three different T. algeriensis essential oils collected from Tunisian regions revealed moderate activity against the same strain of Staphylococcus aureus bacteria ATCC 25923. The IZD of the three oils was 17.20 ± 0.20 mm, 19.40 ± 0.50 mm, and 14.8 ± 0.50 mm [152]. Algerian essential oils also revealed suitable antibacterial activities with IZD values of 18.0 ± 0.70 mm and 17.30 ± 0.58 mm [61].
The microdilution method data indicated that S. aureus is more sensitive to oils and extracts of Algerian species. Indeed, in a study by Rezzoug et al. [132], the essential oil extracted from T. algeriensis from the Saharan Atlas (Laghouat region) revealed a MIC equal to 32.00 µg/mL, and the ethanolic extract showed a MIC of 65.00 µg/mL. In the study by Messaoudi et al. [189], the ethanolic and methanolic extracts also exhibited high antibacterial potential with MICs of 165.00 µg/mL and 40.00 µg/mL, respectively. Furthermore, in a recent study by Ghorbel et al. [50], the antibacterial screening of the aqueous extract and the essential oil of the Tunisian species revealed a MIC of 83.00 µg/mL and 70.00 µg/mL, respectively.
Petroleum ether, chloroform, and n-butanol extracts of the aerial parts of Algerian T. algeriensis exhibited high inhibitory activities against Enterococcus faecalis ATCC29212, with MICs equal to 12.50 µg/mL, 12.50 µg/mL, and 6.25 µg/mL, respectively. The antibacterial activity of organic extracts may be due to flavonoids, phenolic compounds, and triterpenoids, which are well-known for their antimicrobial properties [58]. In the study of Zouari et al. [49], the essential oil extracted from Tunisian T. algeriensis revealed a high antibacterial power with an IZD of 18.50 ± 0.50 mm and a MIC equal to 3.00 μL/mL. However, the MIC and MBC values of the Moroccan essential oil were 0.07% and 0.30%, respectively [155]. In the study by Messaoudi et al. [189], methanolic and ethanolic extracts of Algerian T. algeriensis also significantly inhibited the growth of E. faecalis ATCC49452 bacterial isolates. The MICs were equal to 80.00 µg/mL and 105.00 µg/mL, respectively [189]. Similarly, the aqueous extract (MICs >83.00 µg/mL) and essential oil (MIC = 140.00 µg/mL) of a Tunisian species exerted moderate inhibitory activity against E. faecalis strain ABC3 [50].
In vitro, T. algeriensis essential oil exerted significant antimicrobial effects on pathogenic Bacillus subtilis bacteria. The highest activity was observed by three Tunisian essential oils (Eo1, Eo2, and Eo3) from different regions of the B. subtilis 166 strains [48]. They exhibited antibacterial activity with significant IZD values ranging from 36 to 63 mm and MICs equal to 5.50, 4.00, and 4.50 mg/mL, respectively. Their antibacterial potential was compared to that of the tested antibiotics, chloramphenicol (IZD = 22 mm, MIC = 5 μg/μL) and streptomycin (IZD = 20 mm, MIC = 1 μg/μL). Their antibacterial potential was attributed to monoterpene alcohols such as terpinen-4-ol and linalool. Fatma et al. [57]. showed that a methanolic extract of T. algeriensis from another Tunisian region had an IZD equal to 24.00 mm [57]. In addition, the volatile Algerian oil showed an IZD of 42 mm and MIC of 0.50 µL/mL against B. subtilis ATCC6633.
The anti-infective activities of volatile oils extracted from the aerial parts of T. algeriensis from the north-eastern part of Morocco were evaluated on clinical isolates of Listeria monocytogenes 4b (CECT 935) and L. monocytogenes EGD-e [158]. The oils significantly inhibited bacteria with IZD of 33.70 ± 0.40 mm and 26.70 ± 2.30 mm, respectively. MICs and MBCs were < 0.5 µL/mL against both strains. According to the results, T. algeriensis showed the best bacteriostatic and bactericidal effect, followed by Eucalyptus globulus and Rosmarinus officinalis, which were also tested in the study. Indeed, its essential oil contained high amounts of borneol, linalool, camphene, or β-caryophyllene, and other components that could exert this antimicrobial effect [158].
Ben El Hadj Ali et al. [152] also evaluated the antibacterial activity of essential oils against L. monocytogene with Gentamycin as a positive control. T. algeriensis was collected at the vegetative stage from three different localities in the eastern region of Tunisia. The specimens belonged to the following different bioclimates: sub-humid, upper semi-arid, and lower semi-arid. The oils, when tested, had a strong inhibitory effect, with an IZDs between 20 and 45 mm and MICs between 2.00 and 7.50 mg/mL. However, Bukvicki et al. [244] investigation showed remarkable antibacterial activity of Lybian essential oils with MIC = 0.04 ± 0.00 mg/mL and MBC = 0.09 ± 0.02 mg/mL. The oil demonstrated better activity than the positive controls, ampicillin and streptomycin. Phytochemical screening isolated carvacrol and p-cymene and it was suggested that they were responsible for the observed antibacterial effects [244].
Table 4.
Summary of studies on antimicrobial activities carried on Maghreb Thymus algeriensis Boiss. and Reut (Algeria, Libya, Morocco, and Tunisia).
Nikolić et al. [247] reported the effect of T. algeriensis essential oil from Libya against Streptococcus mutans (IBR S001), S. pyogenes (IBR S004), S. salivarius (IBR S006), and S. sanguinis (IBR S002). The essential oil showed significant antibacterial activities, particularly against S. mutans, a known cariogenic species. The MICs and MBCs against all strains were 40 µL/mL and 80 µL/mL, respectively. In addition, T. algeriensis oil showed an antibacterial potential equal to that of Streptomycin but superior to that of Ampicillin on S. sanguinis. The study showed a correlation between the oil’s antimicrobial activity and chemical composition. It was suggested that the antibacterial activity could be attributed to the presence of the main constituent, thymol, described as an excellent antimicrobial agent in several studies [248]. It could also be related to the involvement of less abundant components in the oil.
The by-products of T. algeriensis, notably the essential oils, were also tested against the gram− bacteria Acinetobacter spp., Enterobacter cloacae, E. coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, Salmonella typhimurium and S. enterica (Table 4). The results show that some are less effective against this gram− bacteria than against gram+ bacteria. This can be due to the outer lipopolysaccharides layer or sophisticated efflux pumps or channels that release any substance foreign to the bacteria to the outside. These characteristics would thus influence the penetration capacity of the active compounds in the extracts or oil in gram–bacteria. Nevertheless, some products from T. algeriensis have proven efficacy against gram-negative bacteria. Essential oils of T. algeriensis from Laghouat, Media, Oum El Bouaghi, Souk Ahras, and Biskra regions of Algeria recorded MICs equal to 64.00 µg/mL, 5.00 µL/mL, 0.05%, 2.50 mg/mL, and 12.50 mg/mL, respectively against E. coli. On the other hand, petroleum ether, chloroform, and n-butanol extracts of T. algeriensis from Chelia Mountain, Batna (Algeria), showed a better efficacy, with MICs equal to 25.00 µg/mL [58]. By contrast, ethanolic extracts of the plant collected in Bechar (southwest of Algeria) [189] and Laghouat (Algerian Saharan Atlas) [132] showed MICs equal to 270.00 µg/mL and 256.00 µg/mL, respectively. For the latter, a MIC of 220.00 µg/mL was determined for its methanolic extract [132]. In Ghorbel et al. [50] study, the essential oil and aqueous extract of T. algeriensis from Mount Orbata of Gafsa (Tunisia) had MICs against E. coli equal to 140.00 µg/mL and > 166.00 µg/mL, respectively. Other oils extracted from plants from Korbous, Jdidi Jebel Mountain, and Hammem Sousse have MICs of 4.50 µL/mL, 3.25 µL/mL, and 5.00 µL/mL, respectively [152]. In addition, the oils from the Tunisian plants from Gafsa, Tamerza, and Kairouan originated IZDs of 30.00–50.00 mm and MICs of 1.80–4.00 mg/mL [48]. However, the Libyan plant gave better antibacterial results than the Tunisian plants, with a MIC equal to 0.08 ± 0.03 mg/mL and an MBC of 0.11 ± 0.07 mg/mL [244].
The essential oils and extracts of T. algeriensis that showed the best antibacterial activity against P. aeruginosa were those from the Tunisian (MIC > 140.00 µg/mL; MIC = 5.00 μL/mL) [49,50] and Algerian regions (MIC = 0.40%; MIC = 2.00 µL/mL) [136,245]. The same is observed for K. pneumoniae, responsible for Nosocomial Pneumonia; the extracts and oils that were most effective against it were Algerian (MIC = 256.00 µg/mL), Moroccan (0.07%), and Tunisian (6.00 μL/mL) origin [54,132,155].
4.5.2. Antifungal Effects
Screening tests of the antifungal activity of T. algeriensis were performed against seven different opportunistic fungi. They revealed that it had moderate to strong efficacy on these microorganisms (Table 4), and in some studies, these by-products were more effective than the drugs used as a reference in the tests.
In the study by Rezzoug et al. [132], moderate antifungal activity was associated with ethanolic extracts and essential oils of Algerian T. algeriensis against Candida glabrata (MIC = 128.00 µg/mL and 32.00 µg/mL, respectively) and C. albicans (MIC = 128.00 µg/mL and 64.00 µg/mL). In another study by Ouakouak et al. [61], the essential oil showed moderate to strong efficacy on fungi but was better than the reference drug (Itraconazole). It significantly reduced C. albicans (13.0 ± 0.40 mm), C. glabrata (18.0 ± 0.60 mm), and Saccharomyces cerevisiae (17.0 ± 0.50 mm) growth. In the study by Dob et al. [136], the growth of Mucor ramanniamus was completely inhibited by T. algeriensis oil at 0.5 µL/mL, while the growth of Fusarium oxysporum, F. ssp. albedinis, S. cerevisiae, and C. albicans was inhibited at 1 µL/mL. The same MIC was recorded by Tunisian essential oil against Fusarium solani [49].
Furthermore, Labiad et al. [155] evaluated the antifungal activity of two Moroccan essential oils against the three pathogenic fungi, Trichophyton violaceum, T. mentagrophytes, and Microsporum canis, and showed potent inhibition of fungal growth in a dose-dependent manner. A significant increase in the inhibition rate depended on increasing the concentration of the different essential oils of T. algeriensis. In the preliminary screening of the antimicrobial activity with the well-diffusion method, the essential oils showed significant efficacy in inhibiting the growth of the tested yeasts, with an inhibition percentage higher than 77% [155].
In the study by Bukvicki et al. [244], the antimicrobial activity of a Libyan essential oil was tested against eight fungi (Table 4), aiming to analyze T. algeriensis as a potential preservative in soft cheese. For antifungal activity, the oil was active at a content of 0.01–0.04 mg/mL, and a minimum fungicidal concentration of 0.01–0.04 mg/mL was determined. In the in situ treatment of sliced cheese with T. algeriensis, the essential oil showed antimicrobial potential against the foodborne pathogenic mold Penicillium aurantiogriseum in vitro and in real food systems (cheese). Indeed, the incidence of contamination of the fungus in sliced cheese decreased from 66 to 0.0% with increasing oil content from 20 to 25 µL/mL, respectively. In addition, the sensory evaluation results of the oil-sprayed cheese showed no change in the texture and color of the surface of the cheese slices after 30 days of storage at a temperature of 4 °C. The results indicated color increasing with the addition of oil, while the taste of the cheese decreased but remained above the limits of unappreciated products. With high antimicrobial capabilities, the essential oil of T. algeriensis could be recommended as a natural antimicrobial additive to extend the shelf life of soft cheese [244].
4.5.3. Antioxidant Activity
Many different assays are commonly used to assess the antioxidant activities of specific compounds or complex mixtures such as oils or extracts. The results of these assays are generally used to select the most effective samples for further investigation [50].
Several techniques have been used to assess the antioxidant activity of T. algeriensis by-products (Table 5), and the DPPH (2,2-diphenyl-1-picrylhydrazyl) radical colorimetric assay is the most widely used. The values usually obtained in the assays are IC50 values, indicating the sample concentration required to scavenge 50% of the DPPH radicals. The results show that the essential oil from a plant harvested in Tunisia is the most antioxidant compared to the other products tested (IC50 = 0.04 μg/mL) [50]. The aerial parts of this plant were collected during the spring season at the flowering stage in April, in the Mont Orbata (Jebel Orbata) of Gafsa, located west of Sfax, at an altitude of 1165 m. This plant is rich in oxygenated monoterpenes, which can act as radical scavengers [50].
Other by-products have also shown a high antioxidant capacity. The methanolic and n-butanol extracts of T. algeriensis from two different regions of the Batna province (Aures and Chelia Mountain) possess significant properties with IC50s of 1.60 ± 0.13 μg/mL and 5.05 ± 0.12 µg/mL, respectively [58,60]. In addition, Moroccan essential oil (province of Al Hoceima) (IC50 = 6.88 ± 0.05 µg/mL) and hydromethanolic extract of T. algeriensis from Bordj bou arreridj (Algeria) (IC50 = 7.40 ± 0.30 µg/mL) are also effective in scavenging free radicals [155,188]. The remaining by-products cannot be considered inferior because relevant phenolic or terpenoid contents were detected in these studies, which were sufficient to justify antioxidant activity (See Table 1 and Table 2).
Table 5 shows that the methanolic extracts tested by the DPPH method have higher activity than the essential oils, with IC50s ranging from 1.60 to 68.80 µg/mL [55,60,150,249,250]. Their potential is because they are polar preparations allowing a better solubility of phenolic compounds with redox properties, enabling them to act as reducing agents, hydrogen donors, and singlet oxygen quenchers [60,247]. Their antioxidant activity depends on the number and position of hydrogen-donating hydroxyl groups bound to aromatic rings. The phenolic compounds can trap free radicals and prevent the oxidation of biological molecules by converting the most numerous ROS into inactive species through donating hydrogen atoms [251].
The other most common method for assessing the antioxidant activity of T. algeriensis is the β-carotene bleaching test (BCB). The technique involves the use of an aqueous emulsion of linoleic acid and β-carotene, which is bleached by radicals generated by the spontaneous oxidation of the fatty acid promoted by thermal induction, usually at 50 °C [252,253]. The results are expressed as a percentage of bleaching inhibition (%), which determines the ability of antioxidants to inhibit lipid peroxidation in the initiation and propagation phases. Quantification is based on the rate change at which the absorbance of β-carotene decreases (~470 nm) in the presence of increasing concentrations of the antioxidant being evaluated [252,253].
For T. algeriensis, some extracts and oils had a clear capacity to protect β-carotene against bleaching (Table 5). In addition to showing the best anti-free radical capacity (which was even higher than that of the synthetic compound Trolox), the methanolic extracts of the Tunisian populations (Ta11, Ta12) also showed high efficacy in the BCB test [62]. The chemical specificity could explain their efficacy compared to the other by-products tested from T. algeriensis in the BCB assay. In more detail, the methanolic extracts analysis of Ta11 and Ta12 leaves (from the upper arid bioclimatic zones) revealed high levels of the phenolic monoterpene carvacrol (2222 and 1375 µg/mL for Ta11 and Ta12 extracts, respectively). These high levels contrast with their absence in the extracts of the other populations studied. Caffeoyl rosmarinic acid was also observed in populations Ta11 and Ta12 at 206.6 and 183 µg/mL of extract, respectively. In addition, the Ta11 population was rich in kaempferol-O-hexuronide (862.80 µg/mL extract) [62]. The hydroethanolic extract, infusion, and decoction of aerial parts of T. algeriensis from the Biskra province (arid Eastern arias of Algeria) also showed some anti-bleaching β-carotene activity (IC50 = 85.00 ± 3.00, 139.00 ± 4.00 and 149.00 ± 3.00 µg/mL, respectively) [183].
The reducing power test (RP) often assesses the electron-donating capacity of antioxidant molecules, i.e., the ability to reduce Fe3+ to Fe2+. During the reaction, the ferric cyanide complex (Fe3+) is reduced to ferrous cyanide (Fe2+), changing the solution medium to different shades of green to blue, depending on the reducing power of the test sample. The solution is Perl Prussian blue when a strong reducing compound is detected, which absorbs at 700 nm [254,255].
The reducing power of T. algeriensis has also been assessed (Table 5) in eight extracts and one oil. The n-butanol extract of the plant from Chelia Mountain, Batna (Algeria), was the best reducing agent, with an IC50 of 4.98 ± 0.48 μg/mL [58]. The others were chloroform extract (IC50 = 24.5 ± 0.52 μg/mL) and petroleum ether extract (IC50 = 25.25 ± 0.08µg/mL). In the study by Ziani et al. [183], decoction (IC50 = 49.80 ± 0.40 µg/mL), infusion (IC50 = 54.00 ± 0.50 µg/mL) as well as hydroethanolic extract (IC50 = 100.20 ± 0.50 µg/mL) of T. algeriensis from also showed a reducing capacity. However, Libyan essential oil also showed reduced power [247].
Known as the Trolox equivalent antioxidant capacity (TEAC) assay, the ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) test is also among the most commonly performed antioxidant studies. ABTS is a stable free radical frequently used to estimate total antioxidant capacity (TAC) [256]. During this test, the stable green-blue radical cationic chromophore, 2,2-azinobis-(3-ethylbenzothiazoline-6-sulfonate) (ABTS•+), is produced by oxidation [257]. It results from the reaction of a strong oxidising agent (e.g., potassium permanganate or potassium persulphate) with the ABTS salt. By adding a hydrogen-donating antioxidant to the reaction, there is a reduction of the blue-green ABTS radical and suppression of its characteristic long-wave absorption spectrum (734 nm) [256,258,259].
The scavenging activity of ABTS was measured for about ten extracts and oils obtained from T. algeriensis (Table 5). Their capacity to trap the ABTS•+ radical was expressed as IC50 (µg/mL). The maximum scavenging activity was found in the essential oil (IC50 = 6.96 ± 0.02 µg/mL) of a plant harvested in Morocco. In his study by Labiad et al. [155], the ABTS results agree with the DPPH test. They indicate that this plant was more effective in scavenging ABTS•+ cationic free radicals than T. broussonetii and T. vulgaris, also tested, but less effective than the synthetic antioxidant Trolox. The high antioxidant power found in this plant can be attributed to its high thymol (a phenolic compound) content (33.00%).
FRAP (ferric reducing antioxidant power) is another test adapted to quantify the ferric reducing antioxidant power of plant by-products. It is the only assay that directly measures antioxidants (or reductants) in a sample compared to other assays measuring inhibition of free radicals [260]. The reaction consists of reducing Fe3+-TPTZ (iron[III]-2,4,6-tripyridyl-S-triazine) to Fe2+-TPTZ in an acidic medium by the presence of an antioxidant. Tripyridyltriazine (TPTZ) is the most used iron-binding ligand in this assay. However, it can be replaced by ferrozine, for example. In the presence of an antioxidant, the reduction reaction of Fe3+-TPTZ results in the creation of a Prussian blue end product, whose color intensity and absorbance at 593 nm will determine the antioxidant power of the test sample [261]. Based on the FRAP assay, the total antioxidant capacity, expressed in mmol Fe2+/l of T. algeriensis by-products, was very significant. The most remarkable were the Tunisian plant’s methanolic extracts, Ta11 and Ta12 [62]. In fact, the determined values for Ta12 were 20.60 ± 0.20 mmol Fe2+/L and for Ta11 were 16.70 ± 0.10 mmol Fe2+/L, contrasting with those for the other samples (Ta1 to Ta10), which were below 7.00 mmol Fe2+/L. In addition, the hydromethanolic extract had the ability to reduce iron (5.30 ± 0.00 mM FeSO4/mg), although to a lesser degree than gallic acid (37.00 ± 0.00 mM FeSO4/mg), ascorbic acid (19.00 ± 0.00 mM FeSO4/mg), and butylated hydroxyanisole (BHA) (16.00 ± 0.00 mM FeSO4/mg), which were used as standard agents [188].
Table 5.
Antioxidant activities of Maghreb Thymus algeriensis (Algeria, Morocco, and Tunisia).
Furthermore, the total antioxidant capacity, expressed as IC50 (μg/mL), is very high in the Tunisian aqueous extract and essential oil (IC50 = 0.04 and 0.06 μg/mL, respectively) [50], followed by the ethanolic extract (IC50 = 0.90 ± 0.06 μg/mL) of T. algeriensis leaves from the Algerian Saharan Atlas (Laghouat region) [132].
The Thiobarbituric acid reactive substances (TBARS) assay is a method of choice for detecting one of the major oxidative stress indicators, lipid peroxidation. This procedure detects thiobarbituric acid reactive substances (TBARS), including lipid hydroperoxides and aldehydes, which increase following oxidative stress. It, therefore, measures the protection of a lipid substrate in the presence of an antioxidant. This assay measures malondialdehyde (MDA), which is the cleavage product of an unsaturated fatty acid endoperoxide resulting from the oxidation of lipid substrates. MDA reacts with thiobarbituric acid (TBA) to form a pink chromogen (TBARS) measured at 532–535 nm. The addition of an antioxidant agent to the test solution inhibits the oxidation process, and the reduced formation of chromogen indicates antioxidant capacity. In this test, an MDA standard is used to construct a standard curve against which unknown samples can be plotted [263].
The results of TBARS analysis on T. algeriensis extracts and essential oils are expressed as EC or IC50, the concentration at which there is 50% inhibition of lipid damage (Table 5). The infusion (INF) and decoction (DEC) preparations, as well as hydroethanolic extracts (ETH) of Algerian plants, showed the most promising results compared to the other T. algeriensis by-products tested. The IC50 values for INF, DEC, and ETH were 26.30 ± 0.20 µg/mL, 22.70 ± 0.30 µg/mL, and 40.30 ± 0.30 µg/mL, respectively. In addition, Libyan essential oil was also evaluated and exhibited a higher efficiency (IC50 = 0.31 ± 0.01 μg/mL) compared to the positive control, Trolox (IC50 = 3.73 ± 1.90 μg/mL) [245].
The study by Hazzit et al. [135] showed that the three Algerian T. algeriensis oils tested have different levels of lipid damage inhibition. In fact, the essential oil from Chréa at 800 m showed a strong antioxidant effect (IC50 = 106.70 ± 8.40 µg/mL), superior to that of the BHT positive control (173.40 ± 4.00 µg/mL). The second oil tested was from Chréa at 1500 m and was not active. The last oil was from El-Asnam and showed only low activity (IC50 = 911.60 ± 7.40 µg/mL) [135]. In this study, T. algeriensis showed great chemical variation, even in samples collected in the same locality. This characteristic seems to be common to all essential oils of Thymus spp. [264], which could influence biological activities.
Cell culture is widely used to evaluate the antioxidant potential of plant products. In the study by Resq et al. [249], the cytotoxic effect of methanolic extract of T. algeriensis (from Algeria) was tested (10–100 µg/mL). After 48 h of incubation, the extract was fully biocompatible with the non-tumoral HaCaT cell model. Following this essay, the antioxidant effect of the extract was also analyzed. The cells were subjected to UVA irradiation (100 J/cm2). This caused an increase in the intracellular ROS content, which was subsequently analyzed by the DCFDA (2′,7′-dichlorofluorescein diacetate) test [249]. The analysis showed that incubating the cells with T. algeriensis extract before UVA exposure significantly changed ROS levels. Following these experiments, Resq et al. [249] investigated the involvement of the transcription factor Nrf-2 (nuclear factor erythroid 2) in the likely molecular mechanisms involved in the antioxidant effects of T. algeriensis methanolic extract. The cells were subjected to the extract treatment for 15–30 min, and the level of the transcription factor Nrf-2 was then analyzed by Western blot. This factor is a crucial regulator of the cellular antioxidant defense system and is highly expressed in epithelial cells, including keratinocytes. This factor is generally held in the cytoplasmic matrix by Kelch-like ECH-associated protein 1 (Keap-1), which subsequently directs it to the proteasome machinery for degradation. In a stressful environment and/or in the presence of antioxidants, the Keap-1 protein is released from Nrf-2. It is translocated into the nucleus and then activates the transcription process of antioxidant genes with sequences called ARE (antioxidant responsive elements) in their promoter region [249]. Following the cells treated with extract and incubation for 15 and 30 min, a significant increase in nuclear levels of Nrf-2 was observed.
4.5.4. Anti-Inflammatory Activity
The study of the anti-inflammatory effects of extracts or essential oils of medicinal plants involves various in vitro procedures [265]. Among these methods, one is the protein denaturation technique. Several inflammatory disorders, such as Parkinson’s disease, Alzheimer’s disease, Huntington’s disease, Amyloid Lateral Sclerosis, Type II diabetes, and cystic fibrosis, are associated with protein aggregation [266,267,268]. Therefore, the inhibition of this phenomenon has been associated with anti-inflammatory activity. The study by Mokhtari et al. [58] evaluated the in vitro anti-inflammatory effect of crude petroleum ether, chloroform, and n-butanol extracts of T. algeriensis (from Algeria) by the egg albumin denaturation method. According to the results, the extracts and standard drugs prevented the denaturation of the protein (albumin) in a dose-dependent manner. At 800 µg/mL, the chloroform extract showed the highest anti-inflammatory effect with inhibition of 45.27%, followed by the petroleum ether (30.26%) and n-butanol (26.03%) extracts. At the same content, the positive control, diclofenac sodium, exhibited the highest inhibition (99.23%) at 800 µg/mL.
Models for in vitro studying anti-inflammatory activity involve techniques in which enzymes and inflammatory mediators are directly exposed to the tested products. In vitro models involving direct monitoring of inflammatory mediator inhibition or activity are probably preferable in terms of sensitivity, reproducibility, and reliability. They allow a better understanding of the mechanisms of action and the inflammatory mediators involved [269]. The in vitro anti-inflammatory potential of T. algeriensis methanolic extract (from Morocco) is being evaluated for its effects on the main enzymes involved in the inflammation pathway, namely, the following: cyclooxygenase-1 (COX-1), cyclooxygenase-2 (COX-2), 5-lipoxygenase (5-LOX), and 5-lipoxygenase activating protein (FLAP) [209]. These enzymes, particularly COX-1 and COX-2, are known targets for various non-steroidal anti-inflammatory drugs. Indeed, the arachidonic acid cascade plays a central role in lipid mediators’ biosynthesis with pro- and anti-inflammatory properties [270,271]. They are collectively known as eicosanoids and possess potent biological activities and maintain normal hemostasis, regulating blood pressure, renal function, reproduction, and host defense. When formed in excess under pathological conditions, these molecules elicit pain, fever, and inflammation and play roles in many acute and chronic diseases [271]. The arachidonic acid is released upon phospholipase stimulation and can be converted to the above-referred lipid mediators by cyclooxygenases and lipoxygenases in the subsequent inflammation phase. Cyclooxygenases catalyze the initial step in forming prostaglandins and thromboxane A2 that trigger inflammation. On their hand, lipoxygenases produce hydroperoxyeicosatetraenoic acids, leukotrienes, and lipoxins [272].
In the study of Sobeh et al. [209], it was demonstrated that a T. algeriensis extract was more potent against COX-2 (IC50 = 0.05 ± 0.01µM) than COX-1 (IC50 = 12.4 ± 0.49µM). Furthermore, it was evidenced a potency similar to Diclofenac in inhibiting lipoxygenase in vitro (IC50 = 2.70 ± 0.23 µM) and to the reference 5-LOX inhibitor, zileuton (IC50 = 3.20 ± 0.15 µM). T. algeriensis extract showed higher selectivity for COX-2 than for COX-1, with a selectivity index similar to that of the selective COX-2 inhibitor Celecoxib (SI value = 248 and 266.2, respectively) and higher than Indomethacin and Diclofenac [209].
An in silico molecular docking study was also performed to evaluate the binding affinities of some compounds identified in T. algeriensis extract with these main enzymes. As a result, it was predicted that compounds have docking positions in the binding sites of the relevant inflammatory proteins similar to those observed for the co-crystallized ligands. The compounds salvianolic acid A, rosmarinic acid glucoside, apigenin 6,8-di-C-hexosides, and quercetin pentoside are the best candidates in this in silico study. For 5-LOX and FLAP, the compounds with the highest predicted potential were salvianolic acid A, apigenin 6,8 di-C-hexosides, and quercetin pentoside [209]. Salvianolic acid A interacts via hydrogen bonding with Gln413 and via pi interactions with Phe177 and Leu607. It also interacts hydrophobically with Ala410. Apigenin 6,8 di-C-hexoside interacts via pi-interactions with Phe177 and His367. It also interacts by a hydrogen bond with Asn554 and indirectly with Ala561 through nearby water molecules. The interactions of both compounds within the active site of 5-LOX suggest their strong potential as high-affinity inhibitors [209]. However, the top scoring compounds for the cyclooxygenases were salvianolic acid A, rosmarinic acid glucoside, and apigenin 6,8 di-C-hexoside. Salvianolic acid A binds to COX-1 through hydrogen bonding to Arg120 and hydrophobic interactions with Ser353. The interactions between apigenin 6,8-di-C-hexosides and COX-1 involve several amino acids, including hydrogen bonds with Arg120, Ser 516, Ile 517, and Gln192, in addition to the hydrophobic interactions with Phe518 as well as Ile523 [209]. The complex of rosmarinic acid glucoside and COX-2 was stabilized by hydrogen bond interactions with Ser530 and Tyr385; it was further strengthened by ionic exchange of the carboxylate oxygen with Arg513 as well as hydrophobic interactions with Phe518 and Val89 [209]. These candidates represent naturally occurring hits for developing safer anti-inflammatory alternatives to currently used drugs [209].
Researchers use numerous techniques to assess the in vivo anti-inflammatory activities of test plant products [269]. Anti-inflammatory effects of T. algeriensis in different doses were evaluated by utilizing different animal models representing various changes associated with inflammation, namely, carrageenan-induced paw oedema, total leukocyte count in paw fluid, and acetic acid-induced vascular permeability. In the study by El Ouahdani et al. [63], the effect was evaluated in vivo in a carrageenan-induced hind paw oedema model. This assay has been increasingly used to test new anti-inflammatory drugs. Indeed, the progression of carrageenan-induced oedema generally correlates with the early exudative stage of inflammation and is a valuable tool for studying systemic anti-inflammatory pathology. The essential oil of T. algeriensis (from Morocco) was administered orally, alone (a dose of 150 mg/Kg) or mixed with another oil of Artemesia herba-alba, against the positive group, which received Diclofenac (p.o., 1%). All groups received their treatments 1 h before the injection of 1% carrageenan, which was injected under the plantar fascia of the right hind paw. The pharmacological test showed that the essential oil of T. algeriensis at a dose of 150 mg/kg inhibited oedema by 83.33 ± 00%. However, when mixed with A. herba-alba oil in the same quantity, the oedema inhibition was better (89.99 ± 4.08%). Similarly, for Diclofenac, a maximum inhibition (88.57 ± 0.81%) under the same conditions and at the sixth hour after carrageenan injection [63].
In Sobeh et al. [209] study, the same in vivo model was used. The methanolic extract of T. algeriensis was tested at three doses (200, 400, and 600 mg/kg). Diclofenac (20 mg/kg) or dexamethasone (2 mg/kg) were tested as positive controls. The data revealed that rats injected one h earlier with T. algeriensis extract (200 and 400 mg/kg, p.o.) showed a slight reduction in oedema thickness compared to control rats. Increasing the dose of the extract to 600 mg/kg did not result in further inhibition of the measured oedema thickness. Diclofenac (20 mg/kg, p.o.) reduced paw thickness by 44% of the control values. Dexamethasone (2 mg/kg, p.o.) showed a 51% reduction compared to vehicle-treated rats [209]. In the study’s second phase, the effect of T. algeriensis extract on leukocyte recruitment in the peritoneal cavity of mice was evaluated [209]. In the essay, the mice were treated orally with 200, 400, and 600 mg/kg of T. algeriensis extract. Diclofenac (p.o., 20 mg/kg) and dexamethasone (p.o., 2 mg/kg) were used as reference standards. After four hours of intraperitoneal injection of carrageenan solution, the mice treated were euthanized. The peritoneal cavity was washed, and the total number of leukocytes was determined. Before the carrageenan stimulus, mice pre-treated with T. algeriensis extract (200, 400, and 600 mg/kg, p.o.) showed a dose-dependent reduction in total leukocyte count. The highest doses (400 and 600 mg/kg) reduced the dose by up to 52 and 62%, respectively, compared to the vehicle. Diclofenac or dexamethasone showed only 39 and 30% reductions in leukocyte counts, respectively. T. algeriensis methanolic extract may decrease leukocyte migration into the peritoneal cavity by inhibiting the expression of the chemotactic substance and/or suppressing adhesion molecule production [273]. This anti-inflammatory effect may be related to its main active constituents, rosmarinic acid (and rosmarinic glucoside), a potent anti-inflammatory agent that inhibits NF-κB activation [274].
The effect of a T. algeriensis methanolic extract on acetic acid-induced vascular permeability was also studied [209]. This in vivo model allows evaluating the extracts’ activity against the first inflammation phase. Increased vascular permeability is a hallmark of acute inflammation, during which endothelial cells contract and separate at their boundaries to expose the basement membrane, freely permeable to plasma proteins and fluids [273,275]. After an injury, phlogistic agents increase vascular permeability at different times. This can be induced chemically (acetic acid) and causes an immediate reaction lasting 24 h [276]. The inhibition of this phenomenon suggests that T. algeriensis extract administered can effectively suppress the exudative phase of acute inflammation. Experimental results showed that injection of acetic acid (0.6%, i.p.) in mice increased vascular permeability, as demonstrated by Evans’ blue absorbance. This effect was attenuated in mice pre-treated one hour before acetic acid injection with T. algeriensis extract (400 or 600 mg/kg, p.o.) by 63 and 58%, respectively. The exudates from mice treated with diclofenac (20 mg/kg) or dexamethasone (2 mg/kg) showed Evans blue readings of 73 and 60%, respectively. The anti-inflammatory effect of the extract on the acute inflammation phase could be attributed to the inhibition of inflammatory mediator release and the inhibition of vasodilation [209].
4.5.5. Anti-Pyretic and Anti-Nociceptive Activity
The effect of T. algeriensis methanolic extract on brewer’s yeast-induced pyrexia in mice was performed [209]. Moreover, called pathogenic fever, this model is triggered by increased prostaglandin E2 (PGE2) synthesis and release of the endogenous pyrogens IL-1β, IL-6, TNF-α, endothelin-1 (ET-1), corticotropin-releasing factor (CRF), bradykinin, and preformed pyrogenic factor (PFPF). PGE2-dependent fever results mainly from the action of cyclooxygenase enzymes activated in the preoptic area of the hypothalamus [277]. Inhibition of these pyrogens is therefore responsible for the antipyretic effect. In the present study, oral administration of T. algeriensis extract (200, 400, and 600 mg/kg, p.o.) did not significantly attenuate the rectal temperature of febrile yeast-induced mice. A trend towards a decrease in body temperature was observed at the highest dose used (600 mg/kg) [209].
The analgesic potential of T. algeriensis was evaluated using the abdominal contortion test induced by intraperitoneal injection of acetic acid in mice (the writhing test). This method is based on counting contortions and contractions caused by acetic acid. The model helps to screen for peripherally and centrally acting agents [278]. In addition, it allows the evaluation of anti-nociceptive responses to different types of drugs (i.e., anticholinergics and antihistamines). Acetic acid induction of spasm triggers the activation of various events that determine nociception, such as the release of mediators like histamine, bradykinin, and prostaglandins [279]. Their decrease suggests that the substance administered has an analgesic effect. In an in vivo study, Moroccan T. algeriensis essential oil was administered alone at 150 mg/Kg or mixed with the essential oil of Artemisia herba-alba to mice, and tramadol was used as a positive control [63]. One hour after T. algeriensis essential oil administration, the animals were given acetic acid intraperitoneally. The agitation numbers were recorded afterward. They are manifested by extension of the hind legs, constriction of the abdomen, or rotation of the trunk [209]. The T. algeriensis essential oil administered alone showed 52.40 ± 3.10 contractions, a slightly lower analgesic effect than that observed for Tramadol (42.00 ± 2.70) and the mixture with Artemisia (29.80 ± 1.92). This effect is also inferior to that of the methanolic extract of Algerian T. algeriensis [209]. At both doses tested (p.o., 200 and 400 mg/kg), the extract showed significant peripheral analgesic activity, represented by a dose-dependent decrease in acetic acid-induced writhes in mice. It showed a 94% reduction in writhing response at the highest dose (400 mg/kg). This effect was greater than that observed with diclofenac (20 mg/kg) or dexamethasone (2 mg/kg).
The hot plate method was used to evaluate the central anti-nociceptive mechanism of T. algeriensis methanolic extract in the production of acute analgesia [209]. The pain and hot plate response involves higher brain centers and is a supraspinal organized response [280]. The extract showed a significant analgesic effect, suggesting the presence of centrally acting anti-nociceptive components [209]. Animals pre-treated with the extract (200 and 400 mg/kg, i.p.) showed a longer response latency in a dose-dependent manner. Furthermore, at the high dose of 400 mg/kg, Algerian T. algeriensis showed a similar effect to nalbuphine. This centrally active narcotic analgesic was used as the reference standard in this experiment [209].
The analgesic activities of T. algeriensis were evaluated using two animal models [62,209]. According to the results, it may have potential clinical applications for treating inflammatory disorders. In the first model, acetic acid indirectly induces the release of endogenous substances that are involved in the modulation of nociception [281]. Resident macrophages and basophils release other factors such as the cytokines IL-1β, TNF, and IL-8 in the abdominal cavity [282,283]. The results of these studies lead to the hypothesis that the anti-nociceptive activity of T. algeriensis may result from direct or indirect inhibition of the pro-inflammatory mediator’s release as well as the participation of resident macrophages [62,249]. The hot plate test is the other model used to assess the central anti-nociceptive activity of T. algeriensis. The latter significantly increased the reaction time in the trial. The effect can be due to flavonoids in the extract, which are known to cross the blood-brain barrier and affect opioid and other receptor types in the CNS [284,285]. The antinociception exerted by the extract presumably relates to the opioid system [209].
4.5.6. Cytotoxic Activities
Five essential oils and six extracts of T. algeriensis (from Algeria, Libya, Tunisia, and Morocco) were tested on 21 normal and cancerous cell models. Interestingly, its by-products were found to have inhibitory activity in vitro on certain cancers.
In the study by Rezzoug et al. [132], the anticancer effects of ethanolic extract and essential oils of T. algeriensis (from the Algerian Saharan Atlas, Laghouat region) were evaluated on five human cancer cell lines. These were human breast (MCF-7 and MDA-MB-231), cervical (HeLa), prostatic (PC3), and leukaemia (K562) cancer cell lines. The MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide) assay was used to investigate the cytotoxic effect of both by-products [132]. The observed antiproliferative activities, presented in the LD50 form, were different between the two. In fact, essential oils (LD50 = 300–746 μg/mL) were much more effective in suppressing cancer cell growth than ethanolic extracts (> 10,000 μg/mL). The essential oil was most effective in inhibiting the growth of K562 cells (LD50 = 300.00 ± 13.00 μg/mL), followed by MCF-7 (647.00 ± 16.00 μg/mL), MDA-MB-231 (715.00 ± 22.00 μg/mL), HeLa (746.00 ± 19.00 μg/mL) and PC3 cells (1067.00 ± 96.00 μg/mL) [132]. On the other hand, the essential oil of the aerial parts of Libyan T. algeriensis showed a better effect against MCF7 (62.53 ± 1.88 μg/mL) and HeLa (64.79 ± 1.51 μg/mL) adenocarcinoma cell lines [245]. The same oil was also tested on the growth of the human tumor cell lines NCI-H460 (non-small cell lung cancer), HCT-15 (colon carcinoma), AGS (gastric adenocarcinoma cell line) as well as on normal PLP2 (porcine liver cells) [245]. The determined LC50s were approximately the same (63.94 ± 0.68 μg/mL, 64.13 ± 1.33 μg/mL, and 64.79 ± 1.51, respectively). However, it showed no efficacy against the non-tumor cell line PLP2 at concentrations above 400 μg/mL [245].
In the study by Resq et al. [249], the cytotoxic effect of methanolic extract of T. algeriensis (from Algeria) was tested (10–100 µg/mL). Four cell lines were used, two cancerous (A431 and SVT2 cells) and two non-tumoral (HaCaT and BALB/c-3T3 cells). After 48 h of incubation, the extract was fully biocompatible with the two non-tumoral cell lines tested and was slightly toxic to the cancer cells, but only at the highest concentration tested.
In a study by Ouakouak et al. [61], the cytotoxic activity of the essential oil of T. algeriensis from the El-Guetfa region, M’sila province (Algeria), was evaluated on the following two human cell lines: HCT116 (colon) and HepG2 (hepatic). Cytotoxicity was evaluated for concentrations between 12.5 and 100 μg/mL of oil using an MTT assay. According to the results, the oil was moderately active against the HepG2 cell line (L50 > 100 μg/mL). On the other hand, it showed cytotoxic activity against HCT116 cells after 48 h of incubation, with LC50 = 39.8 μg/mL and LC90 = 59.6 μg/mL, respectively. At a 100 μg/mL concentration, the essential oil caused 100% inhibition of cells. Moreover, this efficacy is almost equal to that of the reference compound (Doxorubicin). However, in another study by Nikolic et al. [245], Lybian essential oil was tested on the same type of HCT15 cell line but only showed an LC50 equal to 64.13 ± 1.33 μg/mL.
The methanolic extract of the Tunisian T. algeriensis can act as a chemopreventive agent, bearing antioxidant properties [57]. In addition, it effectively protects against the in vitro proliferation of U266 cells in a dose-dependent manner. Indeed, it showed a potential to inhibit cell viability by significantly increasing apoptosis in tumor cells. These results suggest that treatment with methanolic extracts rich in phenolic acids may provide an enhanced therapeutic response in human multiple myeloma cells. However, 50 μg/mL did not affect tumor cell proliferation significantly. Furthermore, the induction of apoptosis in tumor cells was associated with the extract’s natural compounds [57].
Another study evaluated the antitumor activity of essential oil of T. algeriensis from Morocco against the tumor cell line P815 (mastocytoma) and human peripheral blood mononuclear cells (PBMC). Carvacrol was also tested [56]. The MTT results show that all products have a significant and dose-dependent cytotoxic effect against the P815 cell line. The essential oil was most cytotoxic at 0.0625% (v/v) with 100% cell lysis, and its LD50 was 0.01%. In addition, at a concentration capable of inducing cytotoxic activity against tumor cells (P815), no antiproliferative effect was observed against normal PBMC for the essential oil. However, on the contrary, a proliferative effect was observed [56]. This phenomenon was lower for carvacrol (115–133% viability after 48 h of treatment), while for the essential oil it was higher (200% viability). Probably, the oil efficiency on tumoral cells is due to the presence of carvacrol in high concentrations (76%). Indeed, the tested pure carvacrol can induce a very high cytotoxic activity, with an LD50 lower than 0.004% v/v [56]. These results agree with those from other investigations that report that carvacrol has a significant antitumor effect [286].
The cancer chemopreventive effect was studied by Guesmi et al. [153]. HCT116 cells were treated with Tunisian essential oil alone or with TRAIL (TNF-related apoptosis-inducing ligand), which has anticancer effects. The results showed a decrease in cell growth with an increasing oil dose. Its ability to enhance the anticancer effects of TRAIL has also been demonstrated. The essential oil was found to significantly improve the TRAIL-induced growth inhibition of human HCT116 cells in a dose-dependent manner [153].
The mechanisms of these effects were analyzed by different methods, namely, the live dead cell assay, caspase activation, and PARP cleavage [153]. The expression of the death receptor signaling pathway was also analyzed by Western blotting. HCT116 cells were treated with different concentrations of essential oil and analyzed under fluorescence microscopy showed that the volatile fractions induced dose-dependent cell death. Volatile thyme oil dose-dependently increased the number of apoptotic cells from 1.6 to ~ 92.13%. Furthermore, in another essay, the ability of the oil to enhance the apoptotic effect was evaluated. A 0.5 pg/mL concentration of the essential oil-induced moderate cell death. It was observed that 0.5 pg/mL of the oil and 25 ng/mL of TRAIL alone induced reasonable cell death [153]. However, the combination of T. algeriensis and TRAIL would significantly increase cell death (81%). The effect of the essential oil on the colony-forming ability of the colon cancer cell line was also performed. This assay would measure the ability of cultured cells to grow and divide into groups, a characteristic present in tumor cells in vivo [153]. According to the results, HCT116 colonies were susceptible to thyme oil and showed a dose-dependent inhibition of colony formation. T. algeriensis essential oil, even at 0.5 and 1 μg/mL, showed a ~30% and 100% reduction in colony forming ability, respectively [153]. Furthermore, further in vitro experiments showed that co-treatment of HCT116 cells with either the highest concentration (0.5 or 1 μg/mL) of the essential oil or TRAIL/Apo2L (25 ng/mL) alone significantly increased the activation of caspase 8 (5-fold), caspase 9 (2-fold) and PARP proteolysis (1.5-fold) in a dose and time-dependent manner. In addition, the results showed that oil increases the expression of death receptors (DR) and reduces the expression of TRAIL decoy receptors (DcR). It also up-regulates MAPK pathway signaling molecules (p38 kinase, ERK, and JNK), down-regulates c-FLIP, and over-expresses SP1 and CHOP [153]. The last part of this study was to determine the effect of T. algeriensis in the animal model with LPS-induced colon carcinoma [153]. Eight groups of animals (n = 6 mice/group) were used in the study. Two main treatment groups of T. algeriensis are organized, namely, the (1) group treated with T. algeriensis essential oil (oral 12.5 mg/mL) and the other (2) treated with T. algeriensis essential oil (oral 50 mg/mL). The others are (3) saline-treated sham group, (4) LPS-treated group (oral 10 µg/mL), (5) 5-FU-treated group (oral 20 mg/kg/day), (6) TS and LPS-treated group (oral 12.5 mg/mL and 10 µg/mL, respectively), (7) TS and LPS (50 mg/mL and 10 µg/mL, respectively), (8) 5-FU and LPS-treated group (20 mg/kg and 10 µg/mL, respectively). The essential oil and the comparator were administered intragastrically daily one hour before LPS treatment for one week. Observations of the mice’s spontaneous behavior and physical parameters were recorded daily before drug administration. The mice were sacrificed at the end of the experiments. The organ tissues (colon, liver, heart, spleen, kidney, and lung) were immediately collected and processed for macroscopic and microscopic analysis. The colon length of the different groups was also determined [153]. After seven days of treatment with a low dose, the mice showed no toxicity and no spontaneous behavioral changes. In contrast, groups of mice treated with LPS alone or in combination showed a loss of body weight and lower consumption of food and water. On the other hand, the mice treated with T. algeriensis essential oil gained weight. After macroscopic and microscopic study of the tissues, it was found that the LPS-treated colonies suffered damage and a shortening of length. In contrast, administration of T. algeriensis essential oil significantly inhibited colonie shortening. The appearance of the colon in the untreated group treated with T. algeriensis essential oil had a typical architecture. In contrast, the colon tissues in the LPS-induced group were significantly marked by inflammation, lesions, and carcinogenesis of tumor buds. Furthermore, it was observed that the lesions were reduced in the group treated with increasing doses of T. algeriensis essential oil [153].
4.5.7. Neuroprotective Effect
Acetylcholinesterase (AChE) is a key enzyme of the cholinergic nervous system. It is a serine hydrolase whose primary function is to stop neurotransmission by degrading acetylcholine [287,288]. Other non-neuronal functions are attributed to this enzyme, such as involvement in cell growth, apoptosis, drug resistance pathways, response to stress signals, and inflammation [288]. During the development of Alzheimer’s disease, cholinergic neurons in the forebrain become particularly vulnerable. They are accompanied by a progressive decrease in acetylcholine [289,290]. The main drugs developed for this disease are based on the improvement of this cholinergic function by AChE inhibition. However, their therapeutic effects are moderate and of relatively short duration [291,292]. Despite this and due to the lack of validated alternative mechanisms of action, the vast majority of studies are associated with treatments based on the inhibition of this enzyme. In this context, the use of medicinal plants represents one of the alternative strategies to conventional treatments. In fact, some natural compounds, notably phenolic derivatives, are known to modulate intracellular events involved in various neurodegenerative diseases, including the inhibition of AChE, a central therapeutic target in Alzheimer’s disease [48]. One of the strategies for screening potential inhibitors is to evaluate AChE activity in vitro and measure the associated enzyme kinetics. To our knowledge, only two studies have investigated the ability of T. algeriensis by-products to modulate AChE activity.
In the study by Jaouadi et al. (2019), twelve methanolic extracts (from Ta1 to Ta12) of T. algeriensis from Tunisia were tested. They showed a moderate ability to inhibit AChE with significant variations between populations. The lowest activity was observed for extracts from population Ta 3 (IC50 = 3.00 mg/mL) from the upper semi-arid bioclimatic zone. The intermediate activity was determined for populations Ta 1, 8, and 10, with IC50 values ranging from 1.00 to 1.20 mg/mL, while populations Ta 11 and Ta 12, from the upper arid zone, showed the highest activity (IC50 of 0.20 and 0.10 mg/mL, respectively). Nevertheless, the inhibitory activity was lower than that observed for donepezil (IC50 = 18.00 ± 0.10 μg/mL), which is a specific acetylcholinesterase inhibitor drug, used as a positive control [62].
Another study on anticholinesterase activity was carried out on methanolic extracts of an Algerian species [55]. The increase in inhibition percentages was remarkably noted with increasing concentrations of the extract. The inhibition rate exceeded 50%, and the IC50 of the extract was deduced to be 154.47 ± 3.55 μg/mL. The methanolic extract of T. algeriensis recorded an IC50 of 161.53 ± 22.65 μg/mL against butyrylcholinesterase. Again, the percentages of inhibition increased proportionally to the concentrations. From the concentration value of 25 μg/mL, the difference in the performance of the extract compared to the standard became significantly high [55].
An in vivo research work investigated the effect of T. algeriensis on neuropathic pain. According to the International Association for the Study of Pain (IASP), it is caused by injury or disease of the somatosensory nervous system. Primary injury, dysfunction, or transient peripheral or central nervous system disturbance are the leading causes of this pain. Chronic pain can also be caused by systemic diseases such as diabetes, viral infections, multiple sclerosis, and cancer. It is characterized by dysaesthesia (i.e., abnormal unpleasant sensations) and hyperalgesia (an increased response to painful stimuli). Pain in response to a stimulus that does not usually cause pain (allodynia) is also neuropathic pain [293,294]. Several conventional drug treatments for neuropathic pain are available. However, these drugs provide only partial relief in most patients, and specific side effects limit their use [294]. Therefore, further studies are needed to explore and develop new and better therapeutic strategies for treating this type of pain.
The Rezq et al. [249] study examined the possible protective effects of Algerian T. algeriensis, and the CCI-induced neuropathic pain model was used. The molecular mechanisms involved at the peripheral (sciatic nerve) and supraspinal (brainstem) levels were studied. In rats exposed to CCI, the mechanical hyperalgesia test (Pinprick test), the acetone drop test (cold allodynia of the paw), as well as the paintbrush test (dynamic mechanical allodynia) were performed. Oral administration of the Algerian T. algeriensis methanolic extract (200 mg/kg and 400 mg/kg) significantly altered the neuropathic pain behavior in the CCI model [249].
The effect of the extract on structural changes in the sciatic nerve and brainstem was investigated by performing a histopathological analysis of the tissues in all groups of CCI rats. Based on microscopic observations, structural disturbances of the sciatic nerve and brain stem were reduced after treatments with the methanolic extract. The result was comparable to that observed with pregabalin (positive control) at low doses and better than at high doses [249].
Following the histological analyses, an immunohistochemical study of brain stem tissue was performed. An apoptotic marker, caspase-3, was used to determine the apoptotic neurons in the different samples studied. T. algeriensis extract-enhanced synaptophysin expression in the brainstem and suppressed the apoptotic marker caspase-3 in the brainstem. In addition, the effect of T. algeriensis (p.o., 200–400 mg/kg) on synaptophysin (SYN) expression was also studied. The utilization of low or high doses of the extract improved sciatic nerve integrity (19.12 ± 0.55% and 16.04 ± 0.44%, respectively). The myelin sheath was also maintained in the low dose (53.22 ± 1.17%) and high dose (62.00 ± 1.67%) CCI rats compared to the normal nerve group (65.55 ± 1.59%) [249].
The effect of the extract on CCI-induced oxidative stress was investigated. The oxidative and nitrosative stress markers iNOS, NADPH oxidase (NOX1), and catalase (CAT) were analyzed. The CCI groups treated with both doses of T. algeriensis improved their oxidative status by a decrease in NOX1 levels and a dose-dependent increase in CAT activity compared to the values of the sham rats. Pro-inflammatory enzymes (COX-2 and LOX) and mediators (TNF-α, NF-κB, and PGE 2) were also assessed. They were reduced after administration of the extract to the brainstem and sciatic nerve of rats in the CCI model. Indeed, the extract (between 200 and 400 mg/kg) attenuated the inflammatory response after 14 days of treatment. Rats showed a significant reduction of COX2 (58 and 65%, respectively), LOX (40 and 52%, respectively), and PGE2 (17 and 53%, respectively) compared to the CCI control values. In addition, an increase in TNF-α (a pro-inflammatory cytokine) and NF-κB was observed in both brainstem and sciatic nerve on day 14 post-surgery, indicating a neuroinflammatory response. T. algeriensis abolished their increase at all dose levels tested [249].
According to all these results, T. algeriensis can effectively protect against painful peripheral neuropathy. The underlying mechanisms could be the suppression of oxidative stress-induced neuroinflammation and apoptosis. The methanolic extract of T. algeriensis can be a promising therapeutic option for managing neuropathic pain and related diseases [249].
4.5.8. Effect on the Gastrointestinal Tract
In the study by Beyi et al. [59], the aqueous extract of T. algeriensis was tested for its possible spasmolytic and relaxing effects in spontaneously contracting rabbit jejunum preparations. Quantification of relaxation is more accessible in this model than in rat jejunum. Animals were subjected to adrenergic and calcium channel blockade. The cholinergic receptor, the NO, and guanylate cyclase pathways were also tested to assess the extract’s spasmodic and relaxing effects. The results suggest that the activity of the aqueous extract of T. algeriensis is probably mediated by Ca++ and cholinergic receptor antagonism, and does not involve adrenergic receptors, NO, and guanylate cyclase. The relaxing and antispasmodic effects may be due to apigenin, luteolin, and quercitin, which are present in T. algeriensis aqueous extract [295,296]. These components could act alone or with other unidentified compounds [59].
The ulcer protective and antioxidant activity of T. Algeriensis essential oil on HCl/ethanol-induced ulcers in rats was also investigated. In addition, the influence of gender on changes in wound healing, gastric acid secretion, and blood flow at the ulcer margin was also studied. Male and female rat groups were treated with essential oil at a concentration of 54, 117, and 180 mL/kg, p.o. HCl/ethanol was administered after one hour to induce ulcers. Macroscopic evaluation of gastric lesions and measurement of mucus production were investigated. In addition, the gastroprotective effect of the essential oil was assessed by evaluating lipid peroxides and reduced glutathione. The activities of the antioxidant enzymes superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), and glutathione-S-transferase (GST) in the gastric mucosa were also assessed [150].
Oil administration to female rats reduced gastric ulceration induced by acidified ethanol. In fact, treatment with the essential oil (54–180 mg/kg, p.o.) resulted in a dose-dependent reduction of HCl/ethanol-induced gastric lesions, decreasing the ulcer index and the percentage of inhibition mainly at doses of 180 mg/kg for male rats (88.00%) and between 117 and 180 mg/kg for female rats (between 96.25 and 98.85%). In this study, omeprazole could protect the gastric mucosa against HCl/ethanol-induced ulceration (65.95%) [150].
The acidity of the gastric contents in experimental animals pre-treated with the essential oil (female rats) was significantly reduced compared to the ulcer control group. Mucus production of the gastric mucosa was also considerably increased in male rats treated with 180 mg/kg and in female rats (between 117 and 180 mg/kg) compared to the ulcer control group. The results showed protection of the gastric mucosa and inhibition of leukocyte infiltration in the gastric wall in oil-pretreated rats. These results were confirmed by histological observations, which revealed a marked reduction in gastric mucosal damage and cell influx. Some treatment-related histopathological abnormalities were observed in both sexes of rats [150].
The study results indicate that essential oil is protective against HCl/ethanol-induced ulcerogenesis in rats. Females showed higher resistance to ulcers, and gastric lesions occurred less often than in males. The SOD, CAT, GPx, GST, and GSH activities were significantly increased by the administration of the oil to treated rats, suggesting that it can restore these enzymes [150].
4.5.9. Insecticide and Phytotoxic Effects
Plant insecticides have been used to control pests for centuries [297], and they are the main alternative to synthetic insecticides due to their advantages over conventional products [298]. More than 283 plant species belonging to 44 plant families have been involved in habitat manipulation and biological control studies [299]. Fifteen of these plant families have species that have been exploited for their insecticidal properties. The Lamiaceae family is one of the families with the most significant number of species used for habitat manipulation and botanical insecticides [299]. Although plant substances are relatively less effective than modern synthetic substances, their relative safety has opened up a new avenue in plant insecticide research [300].
The insecticidal activity of T. algeriensis essential oils has been tested against different insect species to promote them as biopesticides. In the study by Adouane et al. [51], the essential oil of the Algerian species was tested against different development stages of the date moth Ectomyelois ceratoniae Zeller (Lepidoptera: Pyralidae). This insect species is known as the carob moth and is the primary pest of dates in Algeria, one of the world’s largest date-producing countries [301]. Its proliferation alters the quality of dates, making them unfit for human consumption and marketing [51]. This study involved the following three assays: contact ovicide toxicity test, fumigant toxicity test (against the adult form), and antifeedant toxicity (against the larval form).
Five doses of 0.5, 1, 1.5, 2, and 2.5 mg/mL of oil were tested on egg hatchlings. The hatching rate of the insects decreased with increasing oil content. The hatching rate was 15% when the eggs were exposed to T. algeriensis vapor at 2.5 mg/mL. However, most unhatched eggs had a dead embryo [51]. All applied concentrations of T. algeriensis showed a toxic effect on E. ceratoniae adults after 3, 6, 12, and 24 h of exposure. This toxic effect increased progressively as the content increased. Insect adults treated with T. algeriensis oil reached 80% mortality. In addition, the results of the LC50 bioassay also showed an efficacy of the oil of 0.19 mg/mL [51]. In the antifeedant toxicity bioassay, the results showed a significant effect. The ability of the oil to kill L1 first instar larvae was enhanced with increasing doses and exposure time. The essential oil of T. algeriensis showed moderate antifeedant functions, resulting in 63.33% efficacy after nine days and 66.67% after 12 days when applied at 10 mg/mL content mixed with the diet [51].
The ovicidal activity of T. algeriensis essential oil has been attributed to its monoterpenes. They exert a direct toxic effect on the nervous system of the developing embryo [302]. Essential oils also interfere with the egg hatching process by causing an alteration of oxygen and surface tension within the eggs [303]. Upon contact with adult insects, essential oils alter the physiology and behavior of the insects. They penetrate their respiratory system and cause asphyxiation, leading to final death [304]. In addition, the insecticidal effect of T. algeriensis can be due to its aromatic properties that disgust the insects from the food and reduce or stop feeding [305].
In Morocco, the main pests of stored foodstuffs belong to the Coleoptera order, and these are Sitophilus oryzae (L.), Rhyzopertha dominica (Fab.), Trogoder magranarium (Everts), and Tribolium castaneum (Hbst.). In the study by Labiad et al. [155], the following two of these species were used: R. dominica and S. oryzae. A volume of 3 μL, 12 μL, and 50 μL, respectively, of essential oil, was tested on adults of both species [155]. According to the results, T. algeriensis proved to be potent against both beetle species within the first 24 h for all volumes of oil used (3, 12, and 50 μL) [155]. The main constituents of the tested T. algeriensis oil, such as thymol, γ-terpinene, and p-cymene, were considered responsible for the insecticidal function [306]. A synergistic effect of thymol and carvacrol on the Coleoptera and Lepidoptera orders has been reported in some studies [307,308]. They cause disorganization of the cell membrane of the target insects, resulting in a loss of permeability [309]. In addition, other minor constituents of the oil may also participate in this efficacy through critical synergistic effects.
The insecticidal activity of three T. algeriensis essential oils from Tunisia was also tested on the larvae of the cotton caterpillar, Spodoptera littoralis Boisd. (Lepidoptera: Noctuidae) [152]. It was studied against the third larval stage by a biological fumigation test. The three essential oils of T. algeriensis (Eo1, Eo2, and Eo3), at different essential oil concentrations of 0, 25, 50, 100, and 200 μL/L, were highly toxic. The study showed that as the concentration of essential oils increased, the percentage of mortality increased. However, the lowest concentration tested, 25 μL/L of air, gave 0% mortality for Eo1 (Korbous region) and Eo3 (Hammem Sousse region) and 13% mortality for Eo2 (Dj. Jdidi region) [152]. The mortality percentages increased to 100% when the concentration increased to 100 μL/L of air for Eo2 and Eo3. The LC50 value ranged from 44.25 μL/L air for Eo3 (with limits of 41.75 and 46.5 μL/L air for the lower and upper limits, respectively) to 112.75 μL/L air for Eo1 (with limits of 86.5 and 131 μL/L air for the lower and upper limits, respectively) [152]. The LC90 values were 157.00, 74.25, and 56.25 μL/L of air for Eo1, EO2, and Eo3, respectively. The insecticidal activity of Tunisian T. algeriensis oils is probably related to the following main constituents: α-pinene, 1,8-cineole, caryophyllene oxide, camphor, linalool, camphene, and p-eugenol, which were associated with toxicity in many investigations [300,310,311]. However, the insecticide activity is inherently variable between oils for several reasons, such as plant age, plant tissues or organs, interactions between structural components, etc. [152].
In the same study [152], phytotoxic activity was also investigated. The inhibitory potential of T. algeriensis oils on seed germination, hypocotyl and root length, and dry weight of Medicago sativa L. and Triticum æstivum L. seedlings was investigated. Different essential oil contents (0.10, 0.25, 0.50, and 1.00 mg/mL) were tested, and their allelopathic activity on the germination of M. sativa L. and T. æstivum L. seeds was found to be locally variable. The oils significantly inhibited the germination of M. sativa seeds. The germination percentage varied between 62.00 and 0.00% on the seventh day of germination. Maximum inhibition of seed germination was 100% at 1 mg/mL. At a low content of 0.10 mg/mL, they were also effective with 20.50, 27.00, and 27.50% for Eo3, Eo2, and Eo1, respectively. Rootlet growth is also inhibited. Inhibition ranged from 36.5 (at 0.10 mg/mL; EO3) to 100% (at 1.00 mg/mL, all oils) and hypocotyl from 28.8 to 100%. Biomass production was slightly inhibited in the presence of different oils at 0.10 mg/mL of oil, and the dry weight of seedlings treated with 1.00 mg/mL of different samples was strongly reduced (100%).
4.5.10. Other Effects
The acaricidal effect of the essential oil of T. algeriensis was tested on the Varroa destructor [134]. The acaricide treatment used was spraying the oil at different doses (0.1, 0.3, and 0.5%, v/v). It was applied to the top of the hive frames using sprayers to ensure contact with the treatment with Varroa mites in the four batches of hives (A, B, C, and D). The results of the treatments showed that the oils did not have a negative effect on the activity of the bee colony and the egg laying of the queen. On the other hand, applying the oils reduced the infestation of the different batches infected by V. destructor. The mortality rates obtained are A (4.1%), B (24.0%), C (32.4%), and D (32.6%). The average number of dead mites was for lot A (173), lot B (435), lot C (1274), and lot D (1366) [134].
The antileishmanial activity of T. algeriensis was evaluated in a cytotoxic model involving murine macrophage cells RAW264.7 [151]. Two species of Leishmania were used, L. major and L. infantum, and the classical thiazolyl blue tetrazolium bromide [3-(4,5-dimethyl thiazol-2-yl2,5-diphenyltetrazolium) bromide] MTT test was applied. T. algeriensis essential oil was effective against parasites and non-cytotoxic to cells after 24 h of incubation. It had a leishmanicidal activity against both Leishmania species, with an IC50 value equal to 0.43 µg/mL for L. major and 0.25 µg/mL for L. infantum. In addition, the oil showed low cytotoxicity when incubated with macrophages [151].
Inhibition of angiotensin I-converting enzyme (ACE, EC 3.4.15.1) activity is a practical therapeutic approach to combat hypertensive disorders. The activity of T. algeriensis against this enzyme was tested as it is traditionally used in Tunisian folk medicine to treat hypertension [49]. The essential oil showed a dose-dependent ACE inhibitory activity (50, 100, 150, or 200 μg/mL). The IC50 value was 150 μg/mL, suggesting that T. algeriensis could be used as an ACE inhibitor to prevent and remedy hypertension.
Using the erythrocyte osmotic fragility model, all extracts (petroleum ether, chloroform, and n-butanol) of T. algeriensis showed an anti-haemolytic effect in a concentration-dependent manner [58]. The highest protective effect was noted for the n-butanol extract, with a 50% haemolysis inhibitory concentration value of 322.85 ± 0.87 µg/mL, followed by the chloroform extract at 443.25 ± 0.52 µg/mL. Petroleum ether extract showed the lowest activity with 19.51 ± 0.17% at 800 µg/mL.
4.5.11. Toxicity
Extracts and essential oils of T. algeriensis have been studied for various pharmacological activities; however, data on their potential toxicity are limited.
In a 14-day acute toxicity study in adult male and female Wistar rats, the essential oil collected orally (between 300 and 500 mg/kg body weight) showed no toxicity. Anatomical results showed no abnormal organ damage in the rats. Neither female nor male rats showed toxicity or mortality, and there were no abnormal physiological or behavioral changes or alterations in body weight at any time during the 14 days of observation. Histological examination of the liver and kidneys showed no difference from the control group [150].
In another study, acute oral toxicity was evaluated in two-month-old mice. After 14 days of treatment with T. algeriensis essential oil (150 mg/kg body weight) or combined with Artemisia herba-alba, no abnormal behavior (signs of toxicity) or death of the mice was observed. All the animals survived after the test. Their body weight remained almost stable over time, and there was no significant difference in the body weight variation between treated and control mice over 14 days [63].
5. Conclusions
The botanical characteristics, traditional uses, chemical composition, and pharmacological properties of Thymus algeriensis Boiss. and Reut have been summarised in this review. It can be seen that this plant is rich in various phenolic and terpenic compounds. So far, all these extracts and oils have antitumor, anti-inflammatory, antioxidant, antibacterial, and other pharmacological activities. It is widely used in popular medicine in Northwest Africa, precisely in the Maghreb, where it is endemic. This region is represented by Algeria, Libya, Morocco, Mauritania, and Tunisia. It is used there traditionally as an astringent, expectorant, stimulant, hypertonic, etc. Most of the reported research was carried out with various extracts of different parts of the plant, and it confirms some of its traditional uses. Nevertheless, this extensive knowledge of this species still has some gaps, and further investigations are needed. First of all, given its particular endemism in the Maghreb region, this species should be explored in more detail. This is particularly the case for the two Moroccan and Libyan species, whose phytochemistry has only been examined by two investigations for Morocco and none for Libya. As for the pharmacological research, both species have been evaluated only for their antimicrobial (six studies in total), anti-inflammatory (one study for the Moroccan, none for the Libyan), antitumor/cytotoxic (two studies), antioxidant (six studies), and insecticide (only one study for the Moroccan species) activities. Only one toxicological report and two investigations into the effect of Moroccan T. algeriensis on the intestinal tract (spasmolytic and gastroprotective effects) have been conducted. Therefore, future research should focus on these two so far side-lined species, which in combination with the high degree of endemism, the probability of finding new active molecules is quite high. Secondly, this species has only been extensively assessed for its in vitro antimicrobial activity, whereas a study of its antiviral and antiparasitic activities could provide new interesting bioactive compounds. Thirdly, several traditional claims for T. algeriensis remain unexplored. Indeed, while its anti-inflammatory, antipyretic, antinociceptive, and neuroprotective uses have been scientifically investigated, its use against diabetes and cardiovascular disease has not. Therefore, future studies should be conducted to demonstrate the usefulness of this species against these types of diseases. Fourthly, despite the pharmacological activities performed, little has been performed to progress the various co-products of T. algeriensis toward clinical trials. Studies on some biological activities have been conducted in vitro/in vivo, and some data have not been more thoroughly analyzed. Thus, more research is needed on the phytochemical, pharmacological, and toxicological properties of the co-products of this species.
In conclusion, Thymus algeriensis Boiss. and Reut can be considered as a natural product with potential for future use as medicine, but further investigations are needed. Numerous pharmacological studies have highlighted its therapeutic potential as an antimicrobial, antioxidant, anti-inflammatory, anticancer, antipyretic, antinociceptive, etc., which could therefore serve as a basis for further research. However, future studies should be carried out based on traditional uses. They should be developed on different cell lines and animal models with appropriate controls and doses, for pharmacological and toxicological evaluations, as well as elucidation of the mechanisms of action of the identified bioactive compounds.
Author Contributions
Conceptualization, L.R.S. and R.A.L., methodology L.R.S., P.S., A.C.G., G.A. and R.A.L.; software, R.A.L. and N.S., writing—original draft preparation, R.A.L., N.S. and L.R.S.; writing—review and editing, R.A.L. and L.R.S., supervision, M.I.I., S.S. and J.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tzima, K.; Brunton, N.P.; Rai, D.K. Qualitative and quantitative analysis of polyphenols in Lamiaceae plants—A review. Plants 2018, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Nieto, G. Biological activities of three essential oils of the Lamiaceae family. Medicines 2017, 4, 63. [Google Scholar] [CrossRef] [PubMed]
- Harley, R.M.; Atkins, S.; Budantsev, A.L.; Cantino, P.D.; Conn, B.J.; Grayer, R.; Harley, M.M.; De Tok, R.; Krestovskaja, T.; Morales, R.; et al. Labiatae. In VII Flowering Plants Dicotyledons, Lamiales (Except Acanthaceae Including Avicenniaceae); Kadereit, J., Ed.; The Families and Genera of Vascular Plants; Springer: Berlin/Heidelberg, Germany, 2004; Volume 6, pp. 167–275. [Google Scholar]
- Wordfloraonline Lamiaceae Martinov. Available online: http://www.worldfloraonline.org/taxon/wfo-7000000318 (accessed on 25 June 2022).
- Raja, R.R. Medicinally potential plants of Labiatae (Lamiaceae) family: An overview. Res. J. Med. Plant 2012, 6, 203–213. [Google Scholar] [CrossRef]
- Stankovic, M. (Ed.) Lamiaceae Species: Biology, Ecology and Practical Uses; MDPI: Basel, Switzerland, 2020. [Google Scholar] [CrossRef]
- Carović-Stanko, K.; Petek, M.; Grdiša, M.; Pintar, J.; Bedeković, D.; Ćustić, M.H.; Satovic, Z. Medicinal plants of the family Lamiaceae as functional foods—A review. Czech J. Food Sci. 2016, 34, 377–390. [Google Scholar] [CrossRef]
- Hedge, I. A global survey of the biogeography of the Labiatae. In Advances in Labiatae Science; Harley, R.M., Reynolds, T., Eds.; The Royal Botanical Garden, Kew: London, UK, 1992; pp. 7–17. [Google Scholar]
- Kaufmann, M.; Wink, M. Molecular systematics of the Nepetoideae (Family Labiatae): Phylogenetic implications from RbcL Gene Sequences. Z. Naturforsch. C J. Biosci. 1994, 49, 635–645. [Google Scholar] [CrossRef]
- Damerdji, A. Malacological diversity on Some Lamiaceae in the region of Tlemcen (Northwest Algeria). J. Life Sci. 2013, 7, 856–861. [Google Scholar] [CrossRef][Green Version]
- Michel, J.; Abd Rani, N.Z.; Husain, K. A review on the potential use of medicinal plants from Asteraceae and Lamiaceae plant family in cardiovascular diseases. Front. Pharmacol. 2020, 11, 852. [Google Scholar] [CrossRef]
- Trivellini, A.; Lucchesini, M.; Maggini, R.; Mosadegh, H.; Villamarin, T.S.S.; Vernieri, P.; Mensuali-Sodi, A.; Par-dossi, A. Lamiaceae phenols as multifaceted compounds: Bioactivity, industrial prospects and role of “positive-stress”. Ind. Crops Prod. 2016, 83, 241–254. [Google Scholar] [CrossRef]
- Lee, C.J.; Chen, L.G.; Chang, T.L.; Ke, W.M.; Lo, Y.F.; Wang, C.C. The correlation between skin-care effects and phytochemical contents in Lamiaceae plants. Food Chem. 2011, 124, 833–841. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Ziaee, M.; Palla, F. Essential oils extracted from different species of the Lamiaceae plant family as prospective bioagents against several detrimental pests. Molecules 2020, 25, 1556. [Google Scholar] [CrossRef]
- De Elguea-Culebras, G.O.; Bravo, E.M.; Sánchez-Vioque, R. Potential sources and methodologies for the recovery of phenolic compounds from distillation residues of mediterranean aromatic plants. An approach to the valuation of by-products of the essential oil market—A review. Ind. Crops Prod. 2022, 175, 114261. [Google Scholar] [CrossRef]
- Food and Drug Administration Code of Federal Regulations (CFR). Title 21: Food and Drugs Chapter I—Food and Drug Administration, Department of Health and Human Services, Subchapter B—Food for Human Consumption, Part182—Substances Generally Recognized as Safe (GRAS). Available online: https://www.zotero.org/didalalou/search/FDA/titleCreatorYear/items/3PIJLFG6/item-list (accessed on 25 June 2022).
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A Review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Khazdair, M.R.; Gholamnezhad, Z.; Rezaee, R.; Boskabady, M.H. Immuno-modulatory and anti-inflammatory effects of Thymus Vulgaris, Zataria multiflora, and Portulaca oleracea and their constituents. Pharmacol. Res.—Mod. Chin. Med. 2021, 1, 100010. [Google Scholar] [CrossRef]
- Elufioye, T.O.; Habtemariam, S. Hepatoprotective effects of rosmarinic acid: Insight into its mechanisms of action. Biomed. Pharmacother. 2019, 112, 108600. [Google Scholar] [CrossRef] [PubMed]
- Mouhid, L.; Gómez De Cedrón, M.; Vargas, T.; García-Carrascosa, E.; Herranz, N.; García-Risco, M.; Reglero, G.; Fornari, T.; de Molina, A.R. Identification of antitumoral agents against human pancreatic cancer cells from Asteraceae and Lamiaceae plant extracts. BMC Complement. Altern. Med. 2018, 18, 254. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Dantas, F.G.; de Castilho, P.F.; de Almeida-Apolonio, A.A.; de Araújo, R.P.; Oliveira, K.M.P. Mutagenic potential of medicinal plants evaluated by the Ames Salmonella/microsome assay: A systematic review. Mutat. Res. Rev. Mutat. Res. 2020, 786, 108338. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.M.; Adamczak, A.; Ożarowski, M. Radioprotective effects of plants from the Lamiaceae family. Anticancer. Agents Med. Chem. 2022, 22, 4–19. [Google Scholar] [CrossRef]
- Nabavi, S.M.; Marchese, A.; Izadi, M.; Curti, V.; Daglia, M.; Nabavi, S.F. Plants belonging to the genus Thymus as antibacterial agents: From farm to pharmacy. Food Chem. 2015, 173, 339–347. [Google Scholar] [CrossRef]
- Waller, S.B.; Cleff, M.B.; Serra, E.F.; Silva, A.L.; dos Reis Gomes, A.; de Mello, J.R.B.; de Faria, R.O.; Meireles, M.C.A. Plants from Lamiaceae family as source of antifungal molecules in humane and veterinary medicine. Microb. Pathog. 2017, 104, 232–237. [Google Scholar] [CrossRef]
- Bekut, M.; Brkić, S.; Kladar, N.; Dragović, G.; Gavarić, N.; Božin, B. Potential of selected Lamiaceae plants in an-ti(retro)viral therapy. Pharmacol. Res. 2018, 133, 301–314. [Google Scholar] [CrossRef]
- Koulivand, P.H.; Khaleghi Ghadiri, M.; Gorji, A. Lavender and the nervous system. Evid.-Based Complement. Altern. Med. 2013, 2013, 681304. [Google Scholar] [CrossRef]
- Nematolahi, P.; Mehrabani, M.; Karami-Mohajeri, S.; Dabaghzadeh, F. Effects of Rosmarinus officinalis L. on memory performance, anxiety, depression, and sleep quality in university students: A randomized clinical trial complement. Ther. Clin. Pract. 2018, 30, 24–28. [Google Scholar] [CrossRef]
- Uritu, C.M.; Mihai, C.T.; Stanciu, G.D.; Dodi, G.; Alexa-Stratulat, T.; Luca, A.; Leon-Constantin, M.M.; Stefanescu, R.; Bild, V.; Melnic, S.; et al. Medicinal plants of the family Lamiaceae in pain therapy: A review. Pain Res. Manag. 2018, 2018, 7801543. [Google Scholar] [CrossRef]
- Mohammed, A.; Tajuddeen, N. Antidiabetic compounds from medicinal plants traditionally used for the treatment of diabetes in Africa: A review update (2015–2020). S. Afr. J. Bot. 2022, 146, 585–602. [Google Scholar] [CrossRef]
- Hamza, N.; Berke, B.; Umar, A.; Cheze, C.; Gin, H.; Moore, N. A review of Algerian medicinal plants used in the treatment of diabetes. J. Ethnopharmacol. 2019, 238, 111841. [Google Scholar] [CrossRef]
- The Euro+Med PlantBase—The Information Resource for Euro-Mediterranean Plant Diversity. Available online: https://ww2.bgbm.org/EuroPlusMed/PTaxonDetail.asp?NameId=111505&PTRefFk=8000000 (accessed on 25 June 2022).
- WCSP World Checklist of Selected Plant of Lamiaceae Familly: Royal Botanic Gardens, Kew. Available online: https://wcsp.science.kew.org/reportbuilder.do (accessed on 17 June 2022).
- Fakchich, J.; Elachouri, M. An overview on ethnobotanico-pharmacological studies carried out in Morocco, from 1991 to 2015: Systematic review (part 1). J. Ethnopharmacol. 2021, 267, 113200. [Google Scholar] [CrossRef]
- Bouafia, M.; Amamou, F.; Gherib, M.; Benaissa, M.; Azzi, R.; Nemmiche, S. Ethnobotanical and ethnomedicinal analysis of wild medicinal plants traditionally used in Naâma, Southwest Algeria. Vegetos 2021, 34, 654–662. [Google Scholar] [CrossRef]
- Karous, O.; Jilani, I.B.H.; Ghrabi-Gammar, Z. Ethnobotanical study on plant used by semi-nomad descendants’ community in Ouled Dabbeb-Southern Tunisia. Plants 2021, 10, 642. [Google Scholar] [CrossRef]
- Kachmar, M.R.; Naceiri Mrabti, H.; Bellahmar, M.; Ouahbi, A.; Haloui, Z.; el Badaoui, K.; Bouyahya, A.; Chakir, S. Traditional knowledge of medicinal plants used in the northeastern part of Morocco. Evid.-Based Complement. Altern. Med. 2021, 2021, 6002949. [Google Scholar] [CrossRef]
- El-Gharbaoui, A.; Benítez, G.; González-Tejero, M.R.; Molero-Mesa, J.; Merzouki, A. Comparison of Lamiaceae medicinal uses in eastern Morocco and Eastern Andalusia and in Ibn Al-Baytar’s compendium of simple medicaments. (13th Century CE). J. Ethnopharmacol. 2017, 202, 208–224. [Google Scholar] [CrossRef]
- Nafis, A.; Iriti, M.; Ouchari, L.; el Otmani, F.; Marraiki, N.; Elgorban, A.M.; Syed, A.; Mezrioui, N.; Hassani, L.; Custódio, L. New insight into the chemical composition, antimicrobial and synergistic effects of the Moroccan endemic Thymus atlanticus (Ball) Roussine essential oil in combination with conventional antibiotics. Molecules 2021, 26, 5850. [Google Scholar] [CrossRef]
- Silva, A.M.; Martins-Gomes, C.; Souto, E.B.; Schäfer, J.; Santos, J.A.; Bunzel, M.; Nunes, F.M. Thymus zygis Subsp. Zygis an endemic Portuguese plant: Phytochemical profiling, antioxidant, anti-proliferative and anti-inflammatory activities. Antioxidants 2020, 9, 482. [Google Scholar] [CrossRef]
- Valerio, F.; Mezzapesa, G.N.; Ghannouchi, A.; Mondelli, D.; Logrieco, A.F.; Perrino, E.V.; Ghannouchi, G.N.; Mondelli, A.; Logrieco, D.; Perrino, A.F.; et al. Characterization and antimicrobial properties of essential oils from four wild taxa of Lamiaceae family growing in Apulia. Agronomy 2021, 11, 1431. [Google Scholar] [CrossRef]
- Perrino, E.V.; Valerio, F.; Jallali, S.; Trani, A.; Mezzapesa, G.N. Ecological and biological properties of Satureja cuneifolia Ten. and Thymus spinulosus Ten.: Two wild officinal species of conservation concern in Apulia (Italy). A preliminary survey. Plants 2021, 10, 1952. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P. Crop wild relatives (CWRs) threatened and endemic to Italy: Urgent actions for protection and use. Biology 2022, 11, 193. [Google Scholar] [CrossRef]
- African Plant Database. Available online: https://africanplantdatabase.ch/en/nomen/145270 (accessed on 21 June 2022).
- EMPB. The Euro+Med Plantbase Project, the Information Resource for Euro-Mediterranean Plant Diversity: Lamiaceae Occurring in Algeria. Available online: https://www.emplantbase.org/home.html (accessed on 17 June 2022).
- Quézel, P.; Santa, S. Nouvelle Flore de l’Algérie et des Régions Désertiques Méridionales; CNRS: France, Paris, 1962; Volume 2, pp. 565–1170. [Google Scholar]
- Kew Science Thymus, L. Plants of the World Online. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:30002942-2 (accessed on 16 June 2022).
- Miladi, H.; Mili, D.; ben Slama, R.; Zouari, S.; Ammar, E.; Bakhrouf, A. Antibiofilm formation and anti-adhesive property of three Mediterranean essential oils against a foodborne pathogen Salmonella Strain. Microb. Pathog. 2016, 93, 22–31. [Google Scholar] [CrossRef]
- Fatma, G.; Mouna, B.F.; Mondher, M.; Ahmed, L. In-Vitro Assessment of antioxidant and antimicrobial activities of methanol extracts and essential oil of Thymus hirtus sp. algeriensis. Lipids Health Dis. 2014, 13, 114. [Google Scholar] [CrossRef]
- Zouari, N.; Fakhfakh, N.; Zouari, S.; Bougatef, A.; Karray, A.; Neffati, M.; Ayadi, M.A. Chemical composition, Angiotensin I-Converting enzyme inhibitory, antioxidant and antimicrobial activities of essential oil of Tunisian Thymus algeriensis Boiss. & Reut. (Lamiaceae). Food Bioprod. Process. 2011, 89, 257–265. [Google Scholar] [CrossRef]
- Ghorbel, A.; Fakhfakh, J.; Brieudes, V.; Halabalaki, M.; Fontanay, S.; Duval, R.E.; Mliki, K.; Sayadi, S.; Allouche, N. Chemical composition, antibacterial activity using micro-broth dilution method and antioxidant activity of essential oil and water extract from aerial part of Tunisian Thymus algeriensis Boiss. & Reut. J. Essent. Oil-Bear. Plants 2022, 24, 1349–1364. [Google Scholar] [CrossRef]
- Adouane, S.; Mehaoua, M.S.; Bouatrous, Y.; Tudela, J.; Flamini, G.; Mechaala, S. Natural insecticides from native plants of the mediterranean basin and their activity for the control of the date moth Ectomyelois ceratoniae (Zeller) (Lepidoptera: Pyralidae). J. Plant Dis. Prot. 2022, 129, 775–782. [Google Scholar] [CrossRef]
- Amarti, F.; El Ajjouri, M.; Ghanmi, M.; Satrani, B.; Aafi, A.; Farah, A.; Khia, A.; Guedira, A.; Rahouti, M.; Chaouch, A. Composition chimique et activité antimicrobienne des huiles essentielles de Thymus algeriensis Boiss. & Reut. et Thymus ciliatus (Desf.) Benth. du Maroc. Biotechnol. Agron. Soc. Environ. 2010, 14, 141–148. [Google Scholar]
- Giordani, R.; Hadef, Y.; Kaloustian, J. Compositions and antifungal activities of essential oils of some Algerian aromatic plants. Fitoterapia 2008, 79, 199–203. [Google Scholar] [CrossRef]
- Sara, M.; Yamina, B.; Ramazan, E.; Mesut, G.; Selma, A. Dietary risk of BlaESBL producing multidrug resistant enterobacteriaceae and their inhibition by Artemisia herba-alba and Thymus Algeriensis essential oils. J. Essent. Oil-Bear. Plants 2021, 24, 658–670. [Google Scholar] [CrossRef]
- Zohra, B.F.; Amar, Z.; Chawki, B. Chemical analysis, antioxidant, anti-alzheimer and anti-diabetic effect of two endemic plants from Algeria: Lavandula Antineae and Thymus Algeriensis. Jordan J. Biol. Sci. 2021, 14, 551–558. [Google Scholar] [CrossRef]
- Jaafari, A.; Mouse, H.A.; Rakib, E.M.; M’Barek, L.A.; Tilaoui, M.; Benbakhta, C.; Boulli, A.; Abbad, A.; Zyad, A. Chemical composition and antitumor activity of different Wild varieties of Moroccan Thyme. Rev. Bras. Farmacogn. 2007, 17, 477–491. [Google Scholar] [CrossRef]
- Fatma, G.; Issam, S.; Rawya, S.; Najla, H.; Ahmed, L. Antioxidant potential of four species of natural product and therapeutic strategies for cancer through suppression of viability in the human multiple myeloma cell line U266. Biomed. Environ. Sci. 2019, 32, 22–33. [Google Scholar] [CrossRef]
- Mokhtari, M.; Chabani, S.; Mouffouk, S.; Aberkane, M.C.; Dibi, A.; Benkhaled, M.; Haba, H. Phytochemicals, antihemolytic, anti-inflammatory, antioxidant, and antibacterial activities from Thymus algeriensis. J. Herbs Spices Med. Plants 2021, 27, 253–266. [Google Scholar] [CrossRef]
- Beyi, L.; Zrouri, H.; Makrane, H.; Mekhfi, H.; Ziyyat, A.; Bnouham, M.; Aziz, M. Relaxant and antispasmodic activities of aqueous extract from Thymus algeriensis Boiss. And Reut. J. Nat. Rem. 2021, 21, 165–171. [Google Scholar] [CrossRef]
- Ouakouak, H.; Benarfa, A.; Messaoudi, M.; Begaa, S.; Sawicka, B.; Benchikha, N.; Simal-Gandara, J. Biological properties of essential oils from Thymus Algeriensis Boiss. Plants 2021, 10, 786. [Google Scholar] [CrossRef]
- Jaouadi, R.; Silva, A.M.S.; Boussaid, M.; Yahia, I.B.H.; Cardoso, S.M.; Zaouali, Y. Differentiation of phenolic composition among Tunisian Thymus Algeriensis Boiss. et Reut. (Lamiaceae) populations: Correlation to bioactive activities. Antioxidants 2019, 8, 515. [Google Scholar] [CrossRef]
- El Ouahdani, K.; Es-Safi, I.; Mechchate, H.; Al-Zahrani, M.; Qurtam, A.A.; Aleissa, M.; Bari, A.; Bousta, D. Thymus Algeriensis and Artemisia herba-Alba essential oils: Chemical analysis, antioxidant potential and in vivo anti-inflammatory, analgesic activities, and acute toxicity. Molecules 2021, 26, 6780. [Google Scholar] [CrossRef] [PubMed]
- Bouguerra, A.; Djebili, S.; Zouaoui, N.; Barkat, M. Evaluation of phenolic contents and antioxidant activities of some medicinal plants growing in Algerian Aurès Mountains. Acta Nat. Sci. 2020, 7, 15–30. [Google Scholar] [CrossRef]
- Rey, C. Selection of Thyme for extreme areas (of Switzerland). Acta Hortic. 1992, 306, 66–70. [Google Scholar] [CrossRef]
- Morales, R. The History, Botany and Taxonomy of the Genus Thymus. In Thyme the Genus Thymus; Stahl-Biskup, E., Saez, F., Eds.; CRC Press: London, UK, 2002; pp. 1–43. [Google Scholar]
- Anne, M. Healing with Flowers: The Power of Floral Medicine; Aeon Books: London, UK, 2022; p. 488. ISBN 9781913504793. [Google Scholar]
- Evelyn, K. Thyme & Oregano, Healing and Cooking Herbs, and More than 30 Ways to Use Them; Kindle: Athens, Greece, 2014; ISBN 9781312662186. [Google Scholar]
- Richard, H.; Benjilali, B.; Banquour, N.; Baritaux, O. Etude de diverses huiles essentielles de Thym du Maroc—Institut National de Recherche En Agriculture, Alimentation et Environnement. Lebensm-Wiss.+ Technol. 1985, 18, 105–110. [Google Scholar]
- Beloued, A. Plantes Médicinales d’Algérie; Offices Des Publications Universitaires: Ben Aknoun, Algeria, 2005; ISBN 9789961003046. [Google Scholar]
- Morales, R. Synopsis of the Genus Thymus L. in Mediterranean area. Lagascalia 1997, 19, 249–262. [Google Scholar]
- Karaca, M.; İnce, A.G.; Aydin, A.; Elmasulu, S.Y.; Turgut, K. Microsatellites for genetic and taxonomic research on Thyme (Thymus L.). Turk. J. Biol. 2015, 39, 147–159. [Google Scholar] [CrossRef]
- Bartolucci, F.; Peruzzi, L.; Passalacqua, N. Typification of names and taxonomic notes within the genus Thymus L. (Lamiaceae). Taxon 2013, 62, 1308–1314. [Google Scholar] [CrossRef]
- Bower, A.; Marquez, S.; de Mejia, E.G. The health benefits of selected culinary herbs and spices found in the traditional Mediterranean Diet. Crit. Rev. Food Sci. Nutr. 2016, 56, 2728–2746. [Google Scholar] [CrossRef]
- Amrouche, T.A.; Yang, X.; Capanoglu, E.; Huang, W.; Chen, Q.; Wu, L.; Zhu, Y.; Liu, Y.; Wang, Y.; Lu, B. Contribution of edible flowers to the Mediterranean diet: Phytonutrients, bioactivity evaluation and applications. Front. Nutr. 2022, 9, 1–39. [Google Scholar] [CrossRef]
- Nieto, G. A Review on applications and uses of Thymus in the food industry. Plants 2020, 9, 961. [Google Scholar] [CrossRef]
- Sánchez-Mata, D.; Morales, R. Mediterranean Wild Edible Plants; Sánchez-Mata, M., Tardío, J., Eds.; Springer: New York, NY, USA, 2016; pp. 15–31. [Google Scholar] [CrossRef]
- Tassin, C. Paysages Végétaux du Domaine Méditerranéen: Bassin Méditerranéen, Californie, Chili Central, Afrique du Sud, Australie Méridionale; IRD (Institut de Recherche pour le Développement): Paris, France, 2012; p. 1135. ISBN -10 2709917319. [Google Scholar] [CrossRef]
- Eflora Maghreb Thymus Algeriensis Boiss. & Reut. Available online: https://efloramaghreb.org/specie/145270 (accessed on 21 June 2022).
- El Ajjouri, M.; Ghanmi, M.; Satrani, B.; Amarti, F.; Rahouti, M.; Aafi, A.; Rachid Ismaili, M.; Farah, A. Composition chimique et activité antifongique des huiles essentielles de Thymus Algeriensis Boiss. & Reut. et Thymus Ciliatus (Desf.) Benth. contre les champignons de pourriture du bois. Acta Bot. Gall. 2010, 157, 285–294. [Google Scholar] [CrossRef][Green Version]
- Morales, R. Studies on the genus Thymus. Lamiales Newsl. 1996, 4, 6–8. [Google Scholar]
- Stahl-Biskup, E.; Sáez, F. Thyme: The Genus Thymus, 1st ed.; CRC Press: Boca Raton, FL, USA, 2002; ISBN 9780429218651. [Google Scholar] [CrossRef]
- INaturalist Site, Species Observed in Algeria and Tunisia by Larbi Afoutni and Khaled Bessrour, Occurrence 3455238505. Available online: https://www.gbif.org/occurrence/3455238505 (accessed on 22 July 2022).
- Ali, I.B.E.H.; Zaouali, Y.; Bejaoui, A.; Boussaid, M. Variation of the chemical composition of essential oils in Tunisian populations of Thymus Algeriensis Boiss. et Reut. (Lamiaceae) and implication for conservation. Chem. Biodivers. 2010, 7, 1276–1289. [Google Scholar] [CrossRef]
- Tarayre, M.; Thompson, J.D. Population genetic structure of the Gynodioecious Thymus vulgaris L. (Labiatae) in Southern France. J. Evol. Biol. 1997, 10, 157–174. [Google Scholar] [CrossRef]
- Ben El Hadj Ali, I.; Guetat, A.; Boussaid, M. Chemical and genetic variability of Thymus Algeriensis Boiss. et Reut. (Lamiaceae), a North African endemic species. Ind. Crops Prod. 2012, 40, 277–284. [Google Scholar] [CrossRef]
- Guesmi, F.; Saidi, I.; Bouzenna, H.; Hfaiedh, N.; Landoulsi, A. Phytocompound variability, antioxidant and an-tibacterial activities, anatomical features of glandular and aglandular hairs of Thymus hirtus Willd. ssp. algeriensis Boiss. and Reut. over developmental stages. S. Afr. J. Bot. 2019, 127, 234–243. [Google Scholar] [CrossRef]
- Baba Aïssa, F. Les Plantes Medicinales en Algerie; Bouchéne-ad Diwan: Algiers, Algeria, 1991. [Google Scholar]
- Benkiniouar, R.; Rhouati, S.; Touil, A.; Seguin, E.; Chosson, E. Flavonoids from Thymus algeriensis. Chem. Nat. Compd. 2007, 43, 321–322. [Google Scholar] [CrossRef]
- Mechaala, S.; Bouatrous, Y.; Adouane, S. Traditional knowledge and diversity of Wild medicinal plants in El Kantara’s Area (Algerian Sahara Gate): An ethnobotany survey. Acta Ecol. Sin. 2022, 42, 33–45. [Google Scholar] [CrossRef]
- Hachi, M.; Ouafae, B.; Hachi, T.; Mohamed, E.B.; Imane, B.; Atmane, R.; Zidane, L. Contribution to ethnobotanical study of antidiabetic medicinal plants of the central middle Atlas region (Morocco). Lazaroa 2016, 37, 135–144. [Google Scholar] [CrossRef]
- Sijelmassi, A. Les Plantes Médicinales du Maroc; Le Fennec: Casablanca, Maroco, 1993; Volume 1, p. 285. ISBN -10 9981838020. [Google Scholar]
- Bellakhdar, J. Pharmacopée Marocaine traditionnelle: Médecine Arabe Ancienne et Savoirs Populaires; Paris Ibiss Press: Paris, France, 1997; p. 766. [Google Scholar]
- Ismaili, H.; Milella, L.; Fkih-Tetouani, S.; Ilidrissi, A.; Camporese, A.; Sosa, S.; Altinier, G.; della Loggia, R.; Aquino, R. In Vivo topical anti-inflammatory and in vitro antioxidant activities of two extracts of Thymus satureioides Leaves. J. Ethnopharmacol. 2004, 91, 31–36. [Google Scholar] [CrossRef]
- Ismaili, H.; Sosa, S.; Brkic, D.; Fkih-Tetouani, S.; Ilidrissi, A.; Touati, D.; Aquino, R.P.; Tubaro, A. Topical antiinflammatory activity of extracts and compounds from Thymus broussonettii. J. Pharm. Pharmacol. 2002, 54, 1137–1140. [Google Scholar] [CrossRef] [PubMed]
- Ismaili, H.; Tortora, S.; Sosa, S.; Fkih-Tetouani, S.; Ilidrissi, A.; Della Loggia, R.; Tubaro, A.; Aquino, R. Topical Anti-inflammatory activity of Thymus willdenowii. J. Pharm. Pharmacol. 2001, 53, 1645–1652. [Google Scholar] [CrossRef] [PubMed]
- Pottier-Alapetite, G.A. Flore de la Tunisie: Angiosperme–Dicotylédones: Gamopétales; Ministère de l’Enseignement Supérieur et de la Recherche Scientifique et le Ministère de l’Agriculture: Tunis, Tunisia, 1981; Volume 2. [Google Scholar]
- Le Floc’h, É. Contribution à une Étude Ethnobotanique de la Flore Tunisienne; Ministère de l’Enseignement Supérieur et de la Recherche Scientifique: Tunis, Tunisia, 1983. [Google Scholar]
- Guo, K.; Liu, Y.; Li, S.H. The untapped potential of plant Sesterterpenoids: Chemistry, biological activities and biosynthesis. Nat. Prod. Rep. 2021, 38, 2293–2314. [Google Scholar] [CrossRef] [PubMed]
- Milevskaya, V.V.; Prasad, S.; Temerdashev, Z.A. Extraction and chromatographic determination of phenolic compounds from medicinal herbs in the Lamiaceae and Hypericaceae Families: A review. Microchem. J. 2019, 145, 1036–1049. [Google Scholar] [CrossRef]
- Lichman, B.R.; Godden, G.T.; Buell, C.R. Gene and genome duplications in the evolution of chemodiversity: Perspectives from studies of Lamiaceae. Curr. Opin. Plant Biol. 2020, 55, 74–83. [Google Scholar] [CrossRef]
- Boachon, B.; Buell, C.R.; Crisovan, E.; Dudareva, N.; Garcia, N.; Godden, G.; Henry, L.; Kamileen, M.O.; Kates, H.R.; Kilgore, M.B.; et al. Phylogenomic mining of the mints reveals multiple mechanisms contributing to the evolution of chemical diversity in Lamiaceae. Mol. Plant 2018, 11, 1084–1096. [Google Scholar] [CrossRef]
- Trindade, H.; Pedro, L.G.; Figueiredo, A.C.; Barroso, J.G. Chemotypes and terpene synthase genes in Thymus genus: State of the art. Ind. Crops Prod. 2018, 124, 530–547. [Google Scholar] [CrossRef]
- Perrino, E.V.; Valerio, F.; Gannouchi, A.; Trani, A.; Mezzapesa, G. Ecological and plant community implication on essential oils composition in useful wild officinal species: A Pilot case study in Apulia (Italy). Plants 2021, 10, 574. [Google Scholar] [CrossRef]
- Pelvan, E.; Karaoğlu, Ö.; Önder Fırat, E.; Betül Kalyon, K.; Ros, E.; Alasalvar, C. Immunomodulatory effects of selected medicinal herbs and their essential oils: A comprehensive review. J. Funct. Foods 2022, 94, 105108. [Google Scholar] [CrossRef]
- Masyita, A.; Mustika Sari, R.; Dwi Astuti, A.; Yasir, B.; Rahma Rumata, N.; bin Emran, T.; Nainu, F.; Si-mal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in hu-man health and potential application as natural food preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef]
- Sarkic, A.; Stappen, I. Essential oils and their single compounds in cosmetics—A critical review. Cosmetics 2018, 5, 11. [Google Scholar] [CrossRef]
- Sharmeen, J.B.; Mahomoodally, F.M.; Zengin, G.; Maggi, F. Essential oils as natural sources of fragrance compounds for cosmetics and cosmeceuticals. Molecules 2021, 26, 666. [Google Scholar] [CrossRef] [PubMed]
- Yeshi, K.; Wangchuk, P. Essential oils and their bioactive molecules in healthcare. herbal biomolecules in healthcare applications. In Herbal Biomolecules in Healthcare Applications; Academic Press: Cambridge, MA, USA, 2022; pp. 215–237. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential oils’ chemical characterization and investigation of some biological activities: A critical review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Berger, R.G. Flavours and Fragrances: Chemistry, Bioprocessing and Sustainability; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–648. [Google Scholar] [CrossRef]
- Hüsnü, K.; Baśer, C.; Demirci, F. Chemistry of essential oils. In Flavours and Fragrances: Chemistry, Bioprocessing and Sustainability; Springer: Berlin/Heidelberg, Germany, 2007; pp. 43–86. [Google Scholar] [CrossRef]
- Venditti, A.; Bianco, A.; Frezza, C.; Conti, F.; Bini, L.M.; Giuliani, C.; Bramucci, M.; Quassinti, L.; Damiano, S.; Lupidi, G.; et al. Essential oil composition, polar compounds, glandular trichomes and biological activity of Hyssopus officinalis Subsp. Aristatus (Godr.) Nyman from Central Italy. Ind. Crops Prod. 2015, 77, 353–363. [Google Scholar] [CrossRef]
- Delaney, K.J.; Breza-Boruta, B.; Grzegorz, L.; Bocianowski, J. Maize. VOC Induction after infection by the bacterial pathogen, Pantoea ananatis, alters neighbouring plant voc emissions. J. Plant Dis. Prot. 2015, 122, 125–132. [Google Scholar] [CrossRef]
- Kong, H.G.; Song, G.C.; Sim, H.J.; Ryu, C.M. Achieving similar root microbiota composition in neighbouring plants through airborne signalling. ISME J. 2020, 15, 397–408. [Google Scholar] [CrossRef]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Dicke, M.; van Loon, J.J.A. Multitrophic effects of herbivore-induced plant volatiles in an evolutionary context. Entomol. Exp. Appl. 2000, 97, 237–249. [Google Scholar] [CrossRef]
- Dicke, M.; Bruin, J. Chemical Information Transfer between Plants: Back to the Future. Biochem. Syst. Ecol. 2001, 29, 981–994. [Google Scholar] [CrossRef]
- Agrawal, A.A. Mechanisms, Ecological Consequences and Agricultural Implications of Tri-Trophic Interactions. Curr. Opin. Plant Biol. 2000, 3, 329–335. [Google Scholar] [CrossRef]
- Birkett, M.A.; Campbell, C.A.M.; Chamberlain, K.; Guerrieri, E.; Hick, A.J.; Martin, J.L.; Matthes, M.; Napier, J.A.; Pettersson, J.; Pickett, J.A.; et al. New roles for cis-jasmone as an insect semiochemical and in plant defense. Proc. Natl. Acad. Sci. USA 2000, 97, 9329–9334. [Google Scholar] [CrossRef] [PubMed]
- Karban, R.; Baldwin, I.T.; Baxter, K.J.; Laue, G.; Felton, G.W. Communication between plants: Induced resistance in wild Tobacco plants following clipping of neighboring sagebrush. Oecologia 2000, 125, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.I.; Ozawa, R.; Shimoda, T.; Nishloka, T.; Boland, W.; Takabayashi, J. Herbivory-induced volatiles elicit defence genes in lima bean leaves. Nature 2000, 406, 512–515. [Google Scholar] [CrossRef]
- Fahn, A. Structure and Function of Secretory Cells. Adv. Bot. Res. 2000, 31, 37–75. [Google Scholar] [CrossRef]
- Sadgrove, N.; Jones, G. A Contemporary introduction to essential oils: Chemistry, bioactivity and prospects for australian agriculture. Agriculture 2015, 5, 48–102. [Google Scholar] [CrossRef]
- Abu-Asab, M.; Cantino, P.D. Phylogenetic implications of leaf anatomy in sub-tribe Melittidinae (Labiatae) and Related Taxa. J. Arnold Arbor. 1987, 68, 1–34. [Google Scholar] [CrossRef]
- Seyedi, Z.; Salmaki, Y. Trichome Morphology and Its Significance in the Systematics of Phlomoides (Lamiaceae; Lamioideae; Phlomideae). Flora Morphol. Distrib. Funct. Ecol. Plants 2015, 213, 40–48. [Google Scholar] [CrossRef]
- Bhatt, A.; Naidoo, Y.; Nicholas, A. The foliar trichomes of Plectranthus laxiflorus Benth: An important medicinal plant. N. Z. J. Bot. 2010, 48, 55–61. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; del Mar Contreras, M.; Segura-Carretero, A.; Fathi, H.; Nasrabadi, N.N.; Kobarfard, F.; Sharifi-Rad, J. Thymol, Thyme, and other plant sources: Health and potential uses. Phytother. Res. 2018, 32, 1688–1706. [Google Scholar] [CrossRef]
- Salehi, B.; Abu-Darwish, M.S.; Tarawneh, A.H.; Cabral, C.; Gadetskaya, A.V.; Salgueiro, L.; Hosseinabadi, T.; Ra-jabi, S.; Chanda, W.; Sharifi-Rad, M.; et al. Thymus spp. Plants—Food applications and phytopharmacy properties. Trends Food Sci. Technol. 2019, 85, 287–306. [Google Scholar] [CrossRef]
- Barra, A. Factors affecting chemical variability of essential oils: A review of recent developments. Nat. Prod. Commun. 2009, 8, 1147–1154. [Google Scholar] [CrossRef]
- Elhouiti, F.; Benabed, K.H.; Tahri, D.; Ouinten, M.; Yousfi, M. Antioxidant and antifungal activities of essential oils from algerian spontaneous plants against five strains of Fusarium spp. Hell. Plant Prot. J. 2022, 15, 30–39. [Google Scholar] [CrossRef]
- Zouaoui, N.; Chenchouni, H.; Bouguerra, A.; Massouras, T.; Barkat, M. Characterization of volatile organic compounds from six aromatic and medicinal plant species growing wild in North African drylands. NFS J. 2020, 18, 19–28. [Google Scholar] [CrossRef]
- Rezzoug, M.; Bakchiche, B.; Gherib, A.; Roberta, A.; Guido, F.; Kilinçarslan, Ö.; Mammadov, R.; Bardaweel, S.K. Chemical composition and bioactivity of essential oils and ethanolic extracts of Ocimum basilicum L. and Thymus algeriensis Boiss. & Reut. from the Algerian Saharan Atlas. BMC Complement. Med. Ther. 2019, 19, 146. [Google Scholar] [CrossRef]
- Mehalaine, S.; Chenchouni, H. Effect of climatic factors on essential oil accumulation in two Lamiaceae species from Algerian semiarid lands. Jordan J. Biol. Sci. 2019, 12, 57–60. [Google Scholar] [CrossRef]
- Kouache, B.; Brada, M.; Saadi, A.; Fauconnier, M.L.; Lognay, G.; Heuskin, S. Chemical composition and acaricidal activity of Thymus Algeriensis essential oil against Varroa destructor. Nat. Prod. Commun. 2017, 12, 135–138. [Google Scholar] [CrossRef]
- Hazzit, M.; Baaliouamer, A.; Veríssimo, A.R.; Faleiro, M.L.; Miguel, M.G. Chemical composition and biological activities of Algerian Thymus oils. Food Chem. 2009, 116, 714–721. [Google Scholar] [CrossRef]
- Dob, T.; Dahmane, D.; Benabdelkader, T.; Chelghoum, C. Studies on the essential oil composition and antimicrobial activity of Thymus Algeriensis Boiss. et Reut. Int. J. Aromather. 2006, 16, 95–100. [Google Scholar] [CrossRef]
- Houmani, Z.; Azzoudj, S.; Naxakis, G.; Skoula, M. The essential oil composition of Algerian Zaâtar: Origanum spp. and Thymus spp. J. Herbs Spices Med. Plants 2002, 9, 275–280. [Google Scholar] [CrossRef]
- Hamza, O.; Fahima, A.; Hassani, A.A. Chemical composition, antioxidant activity of the essential oil of Thymus algeriensis Boiss, North Algeria. Int. Lett. Chem. Phys. Astron. 2015, 59, 72–80. [Google Scholar] [CrossRef]
- Mehalaine, S.; Belfadel, O.; Menasria, T.; Messaili, A. Chemical composition and antibacterial activity of essential oils of three medicinal plants from Algerian semi-arid climatic zone. Phytothérapie 2018, 16, S155–S163. [Google Scholar] [CrossRef]
- Kebbi, S.; Fadel, H.; Chalchat, J.; Figueredo, G.; Chalard, P.; Hazmoune, H.; Benayache, F.; Benayache, S.; Seghiri, R. Chemical composition of Algerian Thymus algeriensis Boiss. & Reut. and Marrubium vulgare L. (Lamiaceae) essential oils from the Aures Region. Acta Nat. Sci. 2020, 7, 1–14. [Google Scholar] [CrossRef]
- Jamali, C.A.; Kasrati, A.; Bekkouche, K.; Hassani, L.; Wohlmuth, H.; Leach, D.; Abbad, A. Phenological changes to the chemical composition and biological activity of the essential oil from Moroccan endemic Thyme (Thymus maroccanus Ball). Ind. Crops Prod. 2013, 49, 366–372. [Google Scholar] [CrossRef]
- Goyal, S.; Pathak, R.; Pandey, H.K.; Kumari, A.; Tewari, G.; Bhandari, N.S.; Bala, M. Comparative study of the volatile constituents of Thymus Serpyllum L. grown at different altitudes of Western Himalayas. SN Appl. Sci. 2020, 2, 1208. [Google Scholar] [CrossRef]
- Zouari, N.; Ayadi, I.; Fakhfakh, N.; Rebai, A.; Zouari, S. Variation of chemical composition of essential oils in Wild populations of Thymus Algeriensis Boiss. et Reut., a North African endemic species. Lipids Health Dis. 2012, 11, 28. [Google Scholar] [CrossRef]
- Tátrai, Z.A.; Sanoubar, R.; Pluhár, Z.; Mancarella, S.; Orsini, F.; Gianquinto, G. Morphological and physiological plant responses to drought stress in Thymus citriodorus. J. Agron. 2016, 2016, 4165750. [Google Scholar] [CrossRef]
- Rustaiee, A.R.; Yavari, A.; Nazeri, V.; Shokrpour, M.; Sefidkon, F.; Rasouli, M. Genetic diversity and chemical polymorphism of some Thymus Species. Chem. Biodivers. 2013, 10, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, A.; Moradi, P.; Mastinu, A. Variation in terpene profiles of Thymus vulgaris in water deficit stress response. Molecules 2020, 25, 1091. [Google Scholar] [CrossRef]
- Ćavar Zeljković, S.; Smékalová, K.; Kaffková, K.; Štefelová, N. Influence of post-harvesting period on quality of Thyme and Spearmint essential oils. J. Appl. Res. Med. Aromat. Plants 2021, 25, 100335. [Google Scholar] [CrossRef]
- Rowshan, V.; Bahmanzadegan, A.; Saharkhiz, M.J. Influence of storage conditions on the essential oil composition of Thymus daenensis Celak. Ind. Crops Prod. 2013, 49, 97–101. [Google Scholar] [CrossRef]
- Gavahian, M.; Farahnaky, A.; Javidnia, K.; Majzoobi, M. Comparison of ohmic-assisted hydrodistillation with traditional hydrodistillation for the extraction of essential oils from Thymus vulgaris L. Innov. Food Sci. Emerg. Technol. 2012, 14, 85–91. [Google Scholar] [CrossRef]
- Guesmi, F.; ben Ali, M.; Barkaoui, T.; Tahri, W.; Mejri, M.; Ben-Attia, M.; Bellamine, H.; Landoulsi, A. Effects of Thymus Hirtus sp. algeriensis Boiss. & Reut. (Lamiaceae) essential oil on healing gastric ulcers according to sex. Lipids Health Dis. 2014, 13, 138. [Google Scholar] [CrossRef] [PubMed]
- Ben Hadj Ahmed, S.; Sghaier, R.M.; Guesmi, F.; Kaabi, B.; Mejri, M.; Attia, H.; Laouini, D.; Smaali, I. Evaluation of antileishmanial, cytotoxic and antioxidant activities of essential oils extracted from plants issued from the Leishmaniasis-endemic region of Sned (Tunisia). Nat. Prod. Res. 2011, 25, 1195–1201. [Google Scholar] [CrossRef]
- Ben El Hadj Ali, I.; Chaouachi, M.; Bahri, R.; Chaieb, I.; Boussaïd, M.; Harzallah-Skhiri, F. Chemical composition and antioxidant, antibacterial, allelopathic and insecticidal activities of essential oil of Thymus Algeriensis Boiss. & Reut. Ind. Crops Prod. 2015, 77, 631–639. [Google Scholar] [CrossRef]
- Guesmi, F.; Prasad, S.; ben Ali, M.; Ismail, I.A.; Landoulsi, A. Thymus hirtus sp. algeriensis Boiss. & Reut. volatile oil enhances TRAIL/Apo2L induced apoptosis and inhibits colon carcinogenesis through upregulation of death receptor pathway. Aging 2021, 13, 21975. [Google Scholar] [CrossRef] [PubMed]
- Touhami, A.; Chefrour, A.; Khellaf, N.; Bukhari, A.; Fadel, I. phytochemical characterization of the essential oils obtained from Mediterranean Thymus spp. (Lamiaceae) harvested at different stages of growth. J. Pharm. Pharmacol. 2017, 5, 37–45. [Google Scholar] [CrossRef]
- Labiad, M.H.; Belmaghraoui, W.; Ghanimi, A.; El-Guezzane, C.; Chahboun, N.; Harhar, H.; Egea-Gilabert, C.; Zarrouk, A.; Tabyaoui, M. Biological properties and chemical profiling of essential oils of Thymus (vulgaris, algeriensis and broussonettii) grown in Morocco. Chem. Data Collect. 2022, 37, 100797. [Google Scholar] [CrossRef]
- Salhi, A.; Bouyanzer, A.; El Mounsi, I.; Bendaha, H.; Hamdani, I.; El Ouariachi, E.; Chetouani, A.; Chahboun, N.; Hammouti, B.; Desjobert, J.M.; et al. Chemical composition, anti-oxidant and anticorrosive activities of Thymus algeriensis. J. Mater. Environ. Sci. 2016, 7, 3949–3960. [Google Scholar]
- El Ouariachi, E.M.; Hamdani, I.; Bouyanzer, A.; Hammouti, B.; Majidi, L.; Costa, J.; Paolini, J.; Chetouani, A. Chemical composition and antioxidant activity of essential oils of Thymus broussonetii Boiss. and Thymus algeriensis Boiss. from Morocco. Asian Pac. J. Trop. Dis 2014, 4, 281–286. [Google Scholar] [CrossRef]
- Ait-Ouazzou, A.; Lorán, S.; Bakkali, M.; Laglaoui, A.; Rota, C.; Herrera, A.; Pagán, R.; Conchello, P. Chemical composition and antimicrobial activity of essential oils of Thymus Algeriensis, Eucalyptus globulus and Rosmarinus officinalis from Morocco. J. Sci. Food Agric. 2011, 91, 2643–2651. [Google Scholar] [CrossRef]
- Croteau, R.; Gershenzon, J. Genetic control of monoterpene biosynthesis in Mints (Mentha: Lamiaceae). In Genetic Engineering of Plant Secondary Metabolism; Ellis, B.E., Kuroki, G.W., Stafford, H.A., Eds.; Recent Advances in Phytochemistry; Springer: Boston, MA, USA, 1994; Volume 28, pp. 193–229. [Google Scholar] [CrossRef]
- Osbourn, A.; Lanzotti, V. Plant-Derived Natural Products, Synthesis, Function, and Application; Osbourn, A.E., Lanzotti, V., Eds.; Springer: New York, NY, USA, 2009; p. 597. ISBN 978-0-387-85497-7 158. [Google Scholar] [CrossRef]
- Ifuku, O. Botanical ingredients. In Cosmetic Science and Technology: Theoretical Principles and Applications; Kazutami, S., Robert, Y.L., Howard, I.M., Yuji, Y., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 305–320. [Google Scholar] [CrossRef]
- Saito, K.; Matsuda, F. Metabolomics for functional genomics, systems biology, and biotechnology. Annu. Rev. Plant Biol. 2010, 61, 463–489. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; The International Natural Product Sciences Taskforce; Claudiu, T.S. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.A.; de Souza, L.P.; Serag, A.; Fernie, A.R.; Farag, M.A.; Ezzat, S.M.; Alseekh, S. Metabolomics in the con-text of plant natural products research: From sample preparation to metabolite analysis. Metabolites 2020, 10, 37. [Google Scholar] [CrossRef]
- Scossa, F.; Fernie, A.R. The evolution of metabolism: How to test evolutionary hypotheses at the genomic level. Comput. Struct. Biotechnol. J. 2020, 18, 482–500. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, C.; Borsotto, P. Essential Oils: Market and Legislation. In Potential of Essential Oils; El-Shemy, H.A., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Ivanišová, E.; Kačániová, M.; Savitskaya, T.A.; Grinshpan, D.D. Medicinal Herbs: Important Source of Bioactive Compounds for Food Industry. In Herbs and Spices—New Processing Technologies; Ahmad, R.S., Ed.; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Waqas, S.; Akram, M.; Panda, A.K.; Elbossaty, W.F.; Hegazil, A.G.; Ghasemian, A.; Aharwal, R.P.; Chelladurai, G.; Mandal, S.K.; Mbaye, E.H.S.; et al. Current trends and future prospect of medicinal plants de-rived nutraceuticals: A review. Curr. Trends Pharm. Pharm. Chem. 2022, 4, 30–34. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Al-Rabadi, G.J.; Tranchant, C.C.; Almajwal, A.; Kubow, S.; Alli, I. Occurrence, types, properties and interactions of phenolic compounds with other food constituents in oil-bearing plants. Crit. Rev. Food Sci. Nutr. 2018, 58, 3209–3218. [Google Scholar] [CrossRef]
- Pereira, O.R.; Cardoso, S.M. Overview on Mentha and Thymus polyphenols. Curr. Anal. Chem. 2013, 9, 382–396. [Google Scholar] [CrossRef]
- Leal, F.; Taghouti, M.; Nunes, F.; Silva, A.; Coelho, A.C.; Matos, M. Thymus plants: A review—Micropropagation, molecular and antifungal activity. In Active Ingredients from Aromatic and Medicinal Plants; El-Shemy, H.A., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Frezza, C.; Venditti, A.; Serafini, M.; Bianco, A. Phytochemistry, chemotaxonomy, ethnopharmacology, and nutraceutics of Lamiaceae. Stud. Nat. Prod. Chem. 2019, 62, 125–178. [Google Scholar] [CrossRef]
- Parr, A.J.; Bolwell, G.P. Phenols in the plant and in man. the potential for possible nutritional enhancement of the diet by modifying the phenols content or profile. J. Sci. Food Agric. 2000, 80, 985–1012. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Babotă, M.; Frumuzachi, O.; Gâvan, A.; Iacoviță, C.; Pinela, J.; Barros, L.; Ferreira, I.C.F.R.; Zhang, L.; Lucini, L.; Rocchetti, G.; et al. Optimized ultrasound-assisted extraction of phenolic compounds from Thymus comosus Heuff. Ex Griseb. et Schenk (Wild Thyme) and their bioactive potential. Ultrason. Sonochem. 2022, 84, 105954. [Google Scholar] [CrossRef] [PubMed]
- Barbouti, A.; Goulas, V. Dietary antioxidants in the Mediterranean diet. Antioxidants 2021, 10, 1213. [Google Scholar] [CrossRef] [PubMed]
- Küçükaydın, S.; Çayan, F.; Tel-Çayan, G.; Duru, M.E. HPLC-DAD Phytochemical profiles of Thymus cariensis and T. cilicicus with antioxidant, cytotoxic, anticholinesterase, anti-urease, anti-tyrosinase, and antidiabetic activities. S. Afr. J. Bot. 2021, 143, 155–163. [Google Scholar] [CrossRef]
- Sarfaraz, D.; Rahimmalek, M.; Saeidi, G. Polyphenolic and molecular variation in Thymus species using HPLC and SRAP Analyses. Sci. Rep. 2021, 11, 5019. [Google Scholar] [CrossRef]
- Martins-Gomes, C.; Taghouti, M.; Schäfer, J.; Bunzel, M.; Silva, A.M.; Nunes, F.M. Chemical characterization and bioactive properties of decoctions and hydroethanolic extracts of Thymus carnosus Boiss. J. Funct. Foods 2018, 43, 154–164. [Google Scholar] [CrossRef]
- Silva, A.M.; Félix, L.M.; Teixeira, I.; Martins-Gomes, C.; Schäfer, J.; Souto, E.B.; Santos, D.J.; Bunzel, M.; Nunes, F.M. Orange Thyme: Phytochemical profiling, in vitro bioactivities of extracts and potential health benefits. Food Chem. 2021, 12, 100171. [Google Scholar] [CrossRef]
- Boga, M.; Ozkan, E.E.; Ersoy, E.; Tuncay, E.; Canturk, Y.Y.; Cinar, E.; Kara, E.M.; Zengin, G. Identification and quantification of phenolic and volatile constituents in five different Anatolian Thyme species using LC–MS/MS and GC-MS, with biological activities. Food Biosci. 2021, 43, 101141. [Google Scholar] [CrossRef]
- Taghouti, M.; Martins-Gomes, C.; Schäfer, J.; Félix, L.M.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Thymus pulegioides L. as a rich source of antioxidant, anti-proliferative and neuroprotective phenolic compounds. Food Funct. 2018, 9, 3617–3629. [Google Scholar] [CrossRef]
- Ziani, B.E.C.; Heleno, S.A.; Bachari, K.; Dias, M.I.; Alves, M.J.; Barros, L.; Ferreira, I.C.F.R. Phenolic compounds characterization by LC-DAD- ESI/MSn and bioactive properties of Thymus Algeriensis Boiss. & Reut. and Ephedra alata Decne. Int. Food Res. J. 2019, 116, 312–319. [Google Scholar] [CrossRef]
- Boutaoui, N.; Zaiter, L.; Benayache, F.; Benayache, S.; Carradori, S.; Cesa, S.; Giusti, A.M.; Campestre, C.; Menghini, L.; Innosa, D.; et al. Qualitative and quantitative phytochemical analysis of different extracts from Thymus algeriensis aerial parts. Molecules 2018, 23, 463. [Google Scholar] [CrossRef]
- Mena, P.; Cirlini, M.; Tassotti, M.; Herrlinger, K.A.; Dall’Asta, C.; del Rio, D. Phytochemical profiling of flavonoids, phenolic acids, terpenoids, and volatile fraction of a Rosemary (Rosmarinus officinalis L.) extract. Molecules 2016, 21, 1576. [Google Scholar] [CrossRef]
- Yin, H.; Zou, L.; Sheng, Y.; Bai, X.; Liu, Q.; Yan, B. Rapid HPLC analytical method development for herbal med-icine formulae based on retention rules acquired from the constituting herbs. Anal. Sci. 2018, 34, 207–214. [Google Scholar] [CrossRef]
- Watson, R. (Ed.) Polyphenols in Plants: Isolation, Purification and Extract Preparation; Academic Press: Cambridge, MA, USA, 2018; p. 442. ISBN 9780128137697. [Google Scholar] [CrossRef]
- Righi, N.; Boumerfeg, S.; Fernandes, P.A.R.; Deghima, A.; Baali, F.; Coelho, E.; Cardoso, S.M.; Coimbra, M.A.; Baghiani, A. Thymus algeriensis Bioss & Reut: Relationship of phenolic compounds composition with in vitro/in vivo antioxidant and antibacterial activity. Int. Food Res. J. 2020, 136, 109500. [Google Scholar] [CrossRef]
- Messaoudi, M.; Benreguieg, M.; Merah, M.; Messaoudi, Z.A. Antibacterial effects of Thymus algeriensis extracts on some pathogenic bacteria. Acta Sci. Biol. Sci. 2019, 41, 48548. [Google Scholar] [CrossRef]
- Benabdallah, F.Z.; Zellagui, A.; Demirtas, I. Chemical composition of essential oils and antioxidant activities of extracts of two endemic plants from Algeria. Int. J. Pharm. Sci. Res. 2017, 13, 244–250. [Google Scholar]
- Boulanouar, B.; Abdelaziz, G.; Aazza, S.; Gago, C.; Miguel, M.G. Antioxidant activities of eight Algerian plant extracts and two essential oils. Ind. Crops Prod. 2013, 46, 85–96. [Google Scholar] [CrossRef]
- Dias, M.I.; Sousa, M.J.; Alves, R.C.; Ferreira, I.C.F.R. Exploring plant tissue culture to improve the production of phenolic compounds: A review. Ind. Crops Prod. 2016, 82, 9–22. [Google Scholar] [CrossRef]
- Yu, J.; Vasanthan, T.; Temelli, F. Analysis of phenolic acids in barley by high-performance liquid chromatography. J. Agric. Food Chem. 2001, 49, 4352–4358. [Google Scholar] [CrossRef]
- Robbins, R.J. Phenolic acids in foods: An overview of analytical methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef]
- Gross, G.G. Biosynthesis and metabolism of phenolic acids and monolignols. In Biosynthesis and Biodegradation of Wood Components; Takayoshi, H., Ed.; Academic Press: Kyoto, Japan, 1985; pp. 229–271. [Google Scholar] [CrossRef]
- Zampelas, A.; Micha, R. (Eds.) Antioxidants in Health and Disease; CRC Press: Boca Raton, FL, USA, 2015; 341p. [Google Scholar] [CrossRef]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Appl. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Lorigooini, Z.; Jamshidi-Kia, F.; Hosseini, Z. Analysis of aromatic acids (phenolic acids and hydroxycinnamic acids). In Recent Advances in Natural Products Analysis; Ana, S.S., Seyed, F.N., Mina, S., Seyed, M.N., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 199–219. [Google Scholar] [CrossRef]
- Al Jitan, S.; Alkhoori, S.A.; Yousef, L.F. Chapter 13—Phenolic acids from plants: Extraction and application to human health. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; Volume 58, pp. 389–417. [Google Scholar]
- Clifford, M.N. Review Chlorogenic acids and other cinnamates-nature, occurrence and dietary burden. J. Sci. Food Agric. 1999, 79, 362–372. [Google Scholar] [CrossRef]
- De, P.; Baltas, M.; Bedos-Belval, F. Cinnamic Acid Derivatives as Anticancer Agents—A Review. Curr. Med. Chem. 2011, 18, 1672–1703. [Google Scholar] [CrossRef]
- Ait Kaki, F.; Benkiniouar, R.; Demirtas, I.; Merzoug, A.; Touil, A. Phytochemical study of two Algerian plants Origanum vulgare L. Sbsp. Glandulosum (Desf) Ietswaart and Thymus algeriensis (Boiss. and Reut). Asian J. Chem. 2019, 31, 1105–1109. [Google Scholar] [CrossRef]
- Herrero, M.; Plaza, M.; Cifuentes, A.; Ibáñez, E. Extraction techniques for the determination of phenolic compounds in food. In Comprehensive Sampling and Sample Preparation; Pawliszyn, J., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; Volume 4, pp. 159–180. [Google Scholar] [CrossRef]
- Mekinić, I.G.; Skroza, D.; Šimat, V.; Hamed, I.; Čagalj, M.; Perković, Z.P. Phenolic content of brown Algae (Phe-ophyceae) species: Extraction, identification, and quantification. Biomolecules 2019, 9, 244. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, H.; Seeram, N.P. Development and UFLC-MS/MS characterization of a product-specific standard for phenolic quantification of maple-derived foods. J. Agric. Food Chem. 2016, 64, 3311–3317. [Google Scholar] [CrossRef]
- Michiu, D.; Socaciu, M.I.; Fogarasi, M.; Jimborean, A.M.; Ranga, F.; Mureşan, V.; Semeniuc, C.A. Implementation of an analytical method for spectrophotometric evaluation of total phenolic content in essential oils. Molecules 2022, 27, 1345. [Google Scholar] [CrossRef]
- Chen, H.; Song, X.; Huang, X. Development of magnetism-assisted in-tube solid phase microextraction of phenolic acids in fruit juices prior to high-performance liquid chromatography quantification. J. Sep. Sci. 2021, 44, 3418–3428. [Google Scholar] [CrossRef]
- Sobeh, M.; Rezq, S.; Cheurfa, M.; Abdelfattah, M.A.O.; Rashied, R.M.H.; El-Shazly, A.M.; Yasri, A.; Wink, M.; Mahmoud, M.F. Thymus algeriensis and Thymus fontanesii: Chemical composition, in vivo antiinflammatory, pain killing and antipyretic activities: A comprehensive comparison. Biomolecules 2020, 10, 599. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.; Valentão, P.; Pereira, J.A.P.; Andrade, P.B. Phenolics: From chemistry to biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- Kalinowska, M.; Gołębiewska, E.; Świderski, G.; Męczyńska-Wielgosz, S.; Lewandowska, H.; Pietryczuk, A.; Cudowski, A.; Astel, A.; Świsłocka, R.; Samsonowicz, M.; et al. Plant-derived and dietary hydroxybenzoic acids—A comprehensive study of structural, anti-/pro-oxidant, lipophilic, antimicrobial, and cytotoxic activity in Mda-Mb-231 and Mcf-7 Cell Lines. Nutrients 2021, 13, 3107. [Google Scholar] [CrossRef] [PubMed]
- El-Seedi, H.R.; El-Said, A.M.A.; Khalifa, S.A.M.; Göransson, U.; Bohlin, L.; Borg-Karlson, A.K.; Verpoorte, R. Biosynthesis, natural sources, dietary intake, pharmacokinetic properties, and biological activities of hydroxycinnamic acids. J. Agric. Food Chem. 2012, 60, 10877–10895. [Google Scholar] [CrossRef] [PubMed]
- Fatma, G.; Sami, B.H.A.; Ahmed, L.; Fatma, G.; Sami, B.H.A.; Ahmed, L. Investigation of extracts from Tunisian ethnomedicinal plants as antioxidants, cytotoxins, and antimicrobials. Biomed. Environ. Sci. 2017, 30, 811–824. [Google Scholar] [CrossRef]
- Sova, M.; Saso, L. Natural sources, pharmacokinetics, biological activities and health benefits of hydroxycinnamic acids and their metabolites. Nutrients 2020, 12, 2190. [Google Scholar] [CrossRef]
- Coman, V.; Vodnar, D.C. Hydroxycinnamic Acids and Human Health: Recent Advances. J. Sci. Food Agric. 2020, 100, 483–499. [Google Scholar] [CrossRef]
- Taofiq, O.; González-Paramás, A.M.; Barreiro, M.F.; Ferreira, I.C.F.R.; McPhee, D.J. Hydroxycinnamic acids and their derivatives: Cosmeceutical significance, challenges and future perspectives, a review. Molecules 2017, 22, 281. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Hossain, H.; Hossain, M.; Reza, H.M.; Rahman, M.M.; Ullah, M.O. Hydroxycinnamic acid derivatives: A potential class of natural compounds for the management of lipid metabolism and obesity. Nutr. Metab. 2016, 13, 27. [Google Scholar] [CrossRef]
- Menezes, J.C.J.M.D.S.; Edraki, N.; Kamat, S.P.; Khoshneviszadeh, M.; Kayani, Z.; Mirzaei, H.H.; Miri, R.; Erfani, N.; Nejati, M.; Cavaleiro, J.A.S.; et al. Long Chain alkyl esters of hydroxycinnamic acids as promising anticancer agents: Selective induction of apoptosis in cancer cells. J. Agric. Food Chem. 2017, 65, 7228–7239. [Google Scholar] [CrossRef]
- Rocha, L.D.; Monteiro, M.C.; Teodoro, A.J. Anticancer Properties of Hydroxycinnamic Acids—A Review. Cancer Clin. Oncol. 2012, 1, 109. [Google Scholar] [CrossRef]
- Markham, K.R.; Andersen, O.M. Flavonoids. Chemistry, Biochemistry and Applications; Markham, K.R., Andersen, O.M., Eds.; CRC Press: Boca Raton, FL, USA, 2006; p. 1256. ISBN 9780849320217. [Google Scholar] [CrossRef]
- Dixon, R.A.; Dey, P.M.; Lamb, C.J. Phytoalexins: Enzymology and molecular biology. In Advances in Enzymology and Related Areas of Molecular Biology; John Wiley & Sons: Hoboken, NJ, USA, 1983; Volume 55, pp. 1–136. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef]
- Dixon, R.A.; Pasinetti, G.M. Flavonoids and isoflavonoids: From plant biology to agriculture and neuroscience. Plant Physiol. 2010, 154, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Spencer, J.P.E.; Rice-Evans, C. Flavonoids: Antioxidants or signalling molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Vermerris, W.; Nicholson, R. Phenolic Compound Biochemistry; Springer: Dordrecht, The Netherlands, 2006; pp. 1–276. [Google Scholar] [CrossRef]
- Martens, S.; Preuß, A.; Matern, U. Multifunctional flavonoid dioxygenases: Flavonol and anthocyanin biosynthesis in Arabidopsis thaliana L. Phytochemistry 2010, 71, 1040–1049. [Google Scholar] [CrossRef]
- Rababah, T.M.; Banat, F.; Rababah, A.; Ereifej, K.; Yang, W. Optimization of extraction conditions of total phenolics, antioxidant activities, and anthocyanin of oregano, thyme, terebinth, and pomegranate. J. Food Sci. 2010, 75, C626–C632. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Mulinacci, N.; Innocenti, M.; Bellumori, M.; Giaccherini, C.; Martini, V.; Michelozzi, M. Storage method, drying processes and extraction procedures strongly affect the phenolic fraction of Rosemary leaves: An HPLC/DAD/MS study. Talanta 2011, 85, 167–176. [Google Scholar] [CrossRef]
- Złotek, U.; Mikulska, S.; Nagajek, M.; Świeca, M. The Effect of different solvents and number of extraction steps on the polyphenol content and antioxidant capacity of Basil leaves (Ocimum basilicum L.) extracts. Saudi J. Biol. Sci. 2016, 23, 628–633. [Google Scholar] [CrossRef]
- Costa, D.C.; Costa, H.S.; Albuquerque, T.G.; Ramos, F.; Castilho, M.C.; Sanches-Silva, A. Advances in phenolic compounds analysis of aromatic plants and their potential applications. Trends Food Sci. Technol. 2015, 45, 336–354. [Google Scholar] [CrossRef]
- Ganie, S.H.; Upadhyay, P.; Das, S.; Prasad Sharma, M. Authentication of medicinal plants by dna markers. Plant Gene 2015, 4, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition and antioxidant potential of grain legume seeds: A review. Int. Food Res. J. 2017, 101, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Perez-Vizcaino, F.; Fraga, C.G. Research trends in flavonoids and health. Arch. Biochem. Biophys. 2018, 646, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Pandey, V.; Vadnere, G.P.; Lodhi, S. Role of flavonoids in management of inflammatory disorders. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases; Ronald, R.W., Victor, R.P., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 293–322. [Google Scholar] [CrossRef]
- Ribeiro, D.; Freitas, M.; Lima, J.L.F.C.; Fernandes, E. Proinflammatory pathways: The modulation by flavonoids. Med. Res. Rev. 2015, 35, 877–936. [Google Scholar] [CrossRef]
- Celik, S.; Cakir, E.; Akyuz, S.; Ozel, A.E. Flavonoids: Their Anticarcinogenic Effects and Molecular Modeling Studies. In Handbook of Research on Natural Products and Their Bioactive Compounds as Cancer Therapeutics; IGI Global: Hershey, PA, USA, 2022; pp. 265–296. [Google Scholar] [CrossRef]
- Amarowicz, R.; Pegg, R.B. Natural antioxidants of plant origin. Adv. Food Nutr. Res. 2019, 90, 1–81. [Google Scholar] [CrossRef] [PubMed]
- Benavente-García, O.; Castillo, J. Update on uses and properties of citrus flavonoids: New findings in anti-cancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef] [PubMed]
- Hoensch, H.P.; Oertel, R. The value of flavonoids for the human nutrition: Short review and perspectives. Clin. Nutr. Exp. 2015, 3, 8–14. [Google Scholar] [CrossRef]
- Wright, B.; Spencer, J.P.E.; Lovegrove, J.A.; Gibbins, J.M. Insights into dietary flavonoids as molecular templates for the design of anti-platelet drugs. Cardiovasc. Res. 2013, 97, 13–22. [Google Scholar] [CrossRef]
- Gherairia, N.; Boukerche, S.; Chouikh, A.; Khoudir, S.; Chefrour, A. Antibacterial activity of essential oils from two species of genus Thymus growing in different sites of north-eastern Algerian. An. Univ. Din Oradea Fasc. Biol. 2019, 26, 100–104. [Google Scholar]
- Bukvicki, D.; Giweli, A.; Stojkovic, D.; Vujisic, L.; Tesevic, V.; Nikolic, M.; Sokovic, M.; Marin, P.D. Cheese supplemented with thymus algeriensis oil, a potential natural food preservative. Int. J. Dairy Sci. 2018, 101, 3859–3865. [Google Scholar] [CrossRef]
- Nikolić, M.; Glamočlija, J.; Ferreira, I.C.F.R.; Calhelha, R.C.; Fernandes, Â.; Marković, T.; Marković, D.; Giweli, A.; Soković, M. Chemical composition, antimicrobial, antioxidant and antitumor activity of Thymus serpyllum L., Thymus algeriensis Boiss. and Reut and Thymus vulgaris L. essential oils. Ind. Crops Prod. 2014, 52, 183–190. [Google Scholar] [CrossRef]
- Nikolić, M.; Jovanović, K.K.; Marković, T.; Marković, D.; Gligorijević, N.; Radulović, S.; Soković, M. Chemical composition, antimicrobial, and cytotoxic properties of five Lamiaceae essential oils. Ind. Crops Prod. 2014, 61, 225–232. [Google Scholar] [CrossRef]
- Kachur, K.; Suntres, Z. The Antibacterial properties of phenolic isomers, carvacrol and thymol. Crit. Rev. Food Sci. Nutr. 2019, 60, 3042–3053. [Google Scholar] [CrossRef] [PubMed]
- Rezq, S.; Alsemeh, A.E.; D’Elia, L.; El-Shazly, A.M.; Monti, D.M.; Sobeh, M.; Mahmoud, M.F. Thymus algeriensis and Thymus fontanesii exert neuroprotective effect against chronic constriction injury-induced neuropathic pain in rats. Sci. Rep. 2020, 10, 20559. [Google Scholar] [CrossRef] [PubMed]
- Ben Haj Yahia, I.; Jaouadi, R.; Trimech, R.; Boussaid, M.; Zaouali, Y. Variation of chemical composition and antioxidant activity of essential oils of Mentha x rotundifolia (L.) Huds. (Lamiaceae) collected from different bioclimatic areas of Tunisia. Biochem. Syst. Ecol. 2019, 84, 8–16. [Google Scholar] [CrossRef]
- Ďuračková, Z. Some current insights into oxidative stress. Physiol. Res. 2010, 59, 459–469. [Google Scholar] [CrossRef]
- Marco, G.J. A Rapid method for evaluation of antioxidants. J. Am. Oil Chem. Soc. 1968, 45, 594–598. [Google Scholar] [CrossRef]
- Prieto, M.A.; Rodríguez-Amado, I.; Vázquez, J.A.; Murado, M.A. β-Carotene assay revisited. Application to characterize and quantify antioxidant and prooxidant activities in a microplate. J. Agric. Food Chem. 2012, 60, 8983–8993. [Google Scholar] [CrossRef]
- Yildirim, A.; Mavi, A.; Oktay, M.; Kara, A.A.; Algur, O.F.; Bilaloglu, V. Comparison of antioxidant and antimi-crobial activities of Tilia (Tilia argentea Desf Ex DC), Sage (Salvia triloba l.), and Black Tea (Camellia sinensis) extracts. J. Agric. Food Chem. 2000, 48, 5030–5034. [Google Scholar] [CrossRef]
- Ferreira, I.C.F.R.; Baptista, P.; Vilas-Boas, M.; Barros, L. Free-radical scavenging capacity and reducing power of wild edible mushrooms from Northeast Portugal: Individual cap and stipe activity. Food Chem. 2007, 100, 1511–1516. [Google Scholar] [CrossRef]
- Dong, J.-W.; Cai, L.; Xing, Y.; Yu, J.; Ding, Z.-T. Re-evaluation of ABTS•+ assay for total antioxidant capacity of natural products. Nat. Prod. Commun. 2015, 10, 1934578X1501001239. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. standardized methods for the determination of antioxidant capacity and phe-nolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Opitz, S.E.W.; Smrke, S.; Goodman, B.A.; Yeretzian, C. Methodology for the measurement of antioxidant capacity of coffee: A validated platform composed of three complementary antioxidant assays. In Processing and Impact on Antioxidants in Beverages; Preedy, V., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 253–264. [Google Scholar] [CrossRef]
- Ratnavathi, C.V.; Komala, V.V. Sorghum grain quality. In Sorghum Biochemistry: An Industrial Perspective; Ratnavathi, C.V., Patil, J.V., Chavan, U.D., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 1–61. [Google Scholar] [CrossRef]
- Rezzoug, M.; Bakchiche, B.; Gherib, A.; Elasri, O. Phytochemical screening and antioxidant activities of different organic extracts of three Algerian plants. J. Drug Deliv. Ther. 2020, 10, 75–79. [Google Scholar] [CrossRef]
- Halvorsen, B.; Holte, K.; Mari, C.W.; Barikmo, I.; Hvattum, E.; Fagertun, R.S.; Wold, A.B.; Haffner, K.; Baugerød, H.; Andersen, L.F. A Systematic Screening of Total Antioxidants in Dietary Plants. J. Nutr. 2002, 132, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Shahidi, F. Methods for the assessment of antioxidant activity in foods. In Handbook of Antioxidants for Food Preservation; Shahidi, F., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 287–333. [Google Scholar] [CrossRef]
- Kumar, S.; Krishna Chaitanya, R.; Preedy, V.R. Assessment of antioxidant potential of dietary components. In HIV/AIDS: Oxidative Stress and Dietary Antioxidants; Preedy, V., Watson, R., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 239–253. [Google Scholar] [CrossRef]
- Naghdi Badi, H.; Yazdani, D.; Ali, S.M.; Nazari, F. Effects of spacing and harvesting time on herbage yield and quality/quantity of oil in Thyme, Thymus vulgaris L. Ind. Crops Prod. 2004, 19, 231–236. [Google Scholar] [CrossRef]
- Sarveswaran, R.; Jayasuriya, W.; Suresh, T. In vitro assays to investigate the anti-inflammatory activity of herbal extracts a review. World J. Pharm. Res. 2017, 6, 131. [Google Scholar] [CrossRef]
- Candelise, N.; Scaricamazza, S.; Salvatori, I.; Ferri, A.; Valle, C.; Manganelli, V.; Garofalo, T.; Sorice, M.; Misasi, R. Protein aggregation landscape in neurodegenerative diseases: Clinical relevance and future applications. Int. J. Mol. Sci. 2021, 22, 6016. [Google Scholar] [CrossRef]
- Horwich, A. Protein aggregation in disease: A role for folding intermediates forming specific multimeric interactions. J. Clin. Investig. 2002, 110, 1221–1232. [Google Scholar] [CrossRef]
- Chiti, F.; Dobson, C.M. Protein misfolding, functional amyloid, and human disease. Annu. Rev. Biochem. 2006, 75, 333–366. [Google Scholar] [CrossRef]
- Eze, F.I.; Uzor, P.F.; Ikechukwu, P.; Obi, B.C.; Osadebe, P.O. In vitro and in vivo models for anti-inflammation: An evaluative review. INNOSC Theranost. Pharmacol. Sci. 2019, 2, 755. [Google Scholar] [CrossRef]
- Harizi, H.; Corcuff, J.B.; Gualde, N. Arachidonic-acid-derived eicosanoids: Roles in biology and immuno-pathology. Trends Mol. Med. 2008, 14, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Haeggström, J.Z.; Funk, C.D. Lipoxygenase and leukotriene pathways: Biochemistry, biology, and roles in disease. Chem. Rev. 2011, 111, 5866–5896. [Google Scholar] [CrossRef] [PubMed]
- Garscha, U.; Romp, E.; Pace, S.; Rossi, A.; Temml, V.; Schuster, D.; König, S.; Gerstmeier, J.; Liening, S.; Werner, M.; et al. Pharmacological profile and efficiency in vivo of Diflapolin, the first dual inhibitor of 5-lipoxygenase-activating protein and soluble epoxide hydrolase. Sci. Rep. 2017, 7, 9398. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, E.R.; Toliver-Kinsky, T. Mechanisms of the inflammatory response. Best Pract. Res. Clin. Anaesthesiol. 2004, 18, 385–405. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Chen, J.; Hu, Y.; Lu, W.; Zhang, X.; Wang, R.; Chu, K. Rosmarinic Acid Mitigates lipopolysaccharide induced neuroinflammatory responses through the inhibition of TLR4 and CD14 Expression and NF-ΚB and NLRP3 inflammasome activation. Inflammation 2018, 41, 732–740. [Google Scholar] [CrossRef]
- Opal, S.M.; van der Poll, T. Endothelial barrier dysfunction in septic shock. J. Intern. Med. 2015, 277, 277–293. [Google Scholar] [CrossRef]
- Kumar, T.; Jain, V. Antinociceptive and anti-inflammatory activities of Bridelia retusa methanolic fruit extract in experimental animals. Sci. World J. 2014, 2014, 890151. [Google Scholar] [CrossRef]
- Do Malvar, D.C.; Aguiar, F.A.; Vaz, A.d.L.L.; Assis, D.C.R.; de Melo, M.C.C.; Jabor, V.A.P.; Kalapothakis, E.; Ferreira, S.H.; Clososki, G.C.; de Souza, G.E.P.; et al. Dipyrone metabolite 4-MAA induces hypothermia and inhibits PGE2-dependent and -independent Fever While 4-AA Only Blocks PGE2-dependent Fever. Br. J. Pharmacol. 2014, 171, 3666–3679. [Google Scholar] [CrossRef]
- Le Bars, D.; Gozariu, M.; Cadden, S.W. Animal models of nociception. Pharmacol. Rev. 2001, 53, 597–652. [Google Scholar] [CrossRef]
- Koster, R.; Anderson, M.; De Beer, E.J. Acetic Acid for Analgesic Screening. Fed. Proc. 1959, 18, 412–417. [Google Scholar]
- Chapman, C.R.; Casey, K.L.; Dubner, R.; Foley, K.M.; Gracely, R.H.; Reading, A.E. Pain measurement: An overview. Pain 1985, 22, 1–31. [Google Scholar] [CrossRef]
- Witkin, L.B.; Heubner, C.F.; Galdi, F.; O’Keefe, E.; Spitaletta, P.; Plummer, A.J. Pharmacology of 2-amino-indane hydrochloride (SU-8629): A potent non-narcotic analgesic. J. Pharmacol. Exp. Ther. 1961, 133, 400–408. [Google Scholar]
- Ribeiro, R.A.; Vale, M.L.; Thomazzi, S.M.; Paschoalato, A.B.P.; Poole, S.; Ferreira, S.H.; Cunha, F.Q. Involvement of resident macrophages and mast cells in the writhing nociceptive response induced by Zymosan and Acetic Acid in mice. Eur. J. Pharmacol. 2000, 387, 111–118. [Google Scholar] [CrossRef]
- Liu, J.A.; Yu, J.; Cheung, C.W. Immune actions on the peripheral nervous system in pain. Int. J. Mol. Sci. 2021, 22, 1448. [Google Scholar] [CrossRef]
- Velásquez-Jiménez, D.; Corella-Salazar, D.A.; Zuñiga-Martínez, B.S.; Domínguez-Avila, J.A.; Montiel-Herrera, M.; Salazar-López, N.J.; Rodrigo-Garcia, J.; Villegas-Ochoa, M.A.; González-Aguilar, G.A. Phenolic compounds that cross the blood–brain barrier exert positive health effects as central nervous system antioxidants. Food Funct. 2021, 12, 10356–10369. [Google Scholar] [CrossRef]
- Jäger, A.K.; Saaby, L. Flavonoids and the CNS. Molecules 2011, 16, 1471. [Google Scholar] [CrossRef]
- Sampaio, L.A.; Pina, L.T.S.; Serafini, M.R.; dos Santos Tavares, D.; Guimarães, A.G. Antitumor effects of carvacrol and thymol: A systematic review. Front. Pharmacol. 2021, 12, 1673. [Google Scholar] [CrossRef]
- García-Ayllón, M.S.; Small, D.H.; Avila, J.; Sáez-Valero, J. Revisiting the role of Acetylcholinesterase in Alzheimer’s disease: Cross-talk with β-Tau and p-Amyloid. Front. Mol. Neurosci. 2011, 4, 22. [Google Scholar] [CrossRef]
- Richbart, S.D.; Merritt, J.C.; Nolan, N.A.; Dasgupta, P. Acetylcholinesterase and human cancers. Adv. Cancer Res. 2021, 152, 1–66. [Google Scholar] [CrossRef]
- Davies, P.; Maloney, A.J.F. Selective loss of central cholinergic neurons in Alzheimer’s disease. Lancet 1976, 308, 1403. [Google Scholar] [CrossRef]
- Perry, E.K.; Perry, R.H.; Blessed, G.; Tomlinson, B.E. Necropsy evidence of central cholinergic deficits in senile dementia. Lancet 1977, 309, 189. [Google Scholar] [CrossRef]
- Kaduszkiewicz, H.; Zimmermann, T.; Beck-Bornholdt, H.P.; van Bussche, H. Cholinesterase inhibitors for patients with Alzheimer’s disease: Systematic review of randomised clinical trials. BMJ 2005, 331, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Giacobini, E. Long-term stabilizing effect of cholinesterase inhibitors in the therapy of Alzheimer’ disease. J. Neural Transm. Suppl. 2002, 62, 181–187. [Google Scholar] [CrossRef]
- Cohen, S.; Mao, J. Neuropathic pain: Mechanisms and their clinical implications. BMJ 2014, 348, f7656. [Google Scholar] [CrossRef]
- Bannister, K.; Sachau, J.; Baron, R.; Dickenson, A.H. Neuropathic pain: Mechanism-based therapeutics. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 257–274. [Google Scholar] [CrossRef]
- Modzelewska, B.; Drygalski, K.; Kleszczewski, T.; Chomentowski, A.; Koryciński, K.; Kiełczewska, A.; Pawłusze-wicz, P.; Razak Hady, H. Quercetin relaxes human gastric smooth muscles directly through ATP-Sensitive potassium channels and not depending on the nitric oxide pathway. Neurogastroenterol. Motil. 2021, 33, e14093. [Google Scholar] [CrossRef]
- Sadraei, H.; Ghanadian, M.; Asghari, G.; Sekhavati, N. Antispasmodic activity of apigenin and luteolin, two components of Dracocephalum kotschyi extract, on rat ileum contractions. J. HerbMed Pharmacol. 2018, 7, 100–105. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef]
- Chengala, L.; Singh, N. Botanical pesticides—A Major alternative to chemical pesticides: A Review. Int. J. Life Sci. 2017, 5, 722–729. [Google Scholar]
- Amoabeng, B.W.; Johnson, A.C.; Gurr, G.M. Natural enemy enhancement and botanical insecticide source: A review of dual use companion plants. Appl. Entomol. Zool. 2019, 54, 1–19. [Google Scholar] [CrossRef]
- Souto, A.L.; Sylvestre, M.; Tölke, E.D.; Tavares, J.F.; Barbosa-Filho, J.M.; Cebrián-Torrejón, G. Plant-derived pesticides as an alternative to pest management and sustainable agricultural production: Prospects, applications and challenges. Molecules 2021, 26, 4835. [Google Scholar] [CrossRef] [PubMed]
- Mehaoua, M.S.; Hadjeb, A.; Lagha, M.; Bensalah, M.K.; Ouakid, M.L. Study of the toxicity of Azadira chtinon larval mortality and fertility of Carob Moth’s Female Ectomyelois ceratoniae (Lepidoptera, Pyralidae) under Controlled. Am.-Eurasian J. Sustain. Agric. 2013, 7, 1–9. [Google Scholar]
- Credland, P.F. The structure of bruchid eggs may explain the ovicidal effect of oils. J. Stored Prod. Res. 1992, 28, 1–9. [Google Scholar] [CrossRef]
- Abdullahi, N.; Majeed, Q.; Oyeyi, T.I. Studies on the efficacy of Vittallaria paradoxa seed oil on the oviposition, hatchability of eggs and emergence of Callasobruchus maculatus (F.) (Coleoptera: Bruchidae) on treated cow-pea seed. J. Entomol. 2011, 8, 391–397. [Google Scholar] [CrossRef][Green Version]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Arasu, M.V.; Al-Dhabi, N.A.; Saritha, V.; Duraipandiyan, V.; Muthukumar, C.; Kim, S.J. Antifeedant, larvicidal and growth inhibitory bioactivities of novel polyketide metabolite isolated from Streptomyces sp. AP-123 against Helicoverpa armigera and Spodoptera litura. BMC Microbiol. 2013, 13, 105. [Google Scholar] [CrossRef]
- Das Graças Freire De Medeiros, M.; da Silva, A.C.; das Graças Lopes Citó, A.M.; Borges, A.R.; de Lima, S.G.; Lopes, J.A.D.; Figueiredo, R.C.B.Q. In vitro antileishmanial activity and cytotoxicity of essential oil from Lippia sidoides Cham. Parasitol. Int. 2011, 60, 237–241. [Google Scholar] [CrossRef]
- Karin Lima, R.; das Graças Cardoso, M.; Campos Moraes, J.; Malfitano Carvalho, S.; Gregório Rodrigues, V.; Gustavo Lima Guimarães, L. Chemical composition and fumigant effect of essential oil of Lippia sidoides Cham. and monoterpenes against Tenebrio molitor (L.) (Coleoptera: Tenebrionidae). Ciênc. Agrotec. 2011, 35, 664–671. [Google Scholar] [CrossRef]
- Rice-evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic. Res. 1995, 22, 375–383. [Google Scholar] [CrossRef]
- Pavela, R. Essential oils for the development of eco-friendly mosquito larvicides: A review. Ind. Crops Prod. 2015, 76, 174–187. [Google Scholar] [CrossRef]
- Liu, J.; Hua, J.; Qu, B.; Guo, X.; Wang, Y.; Shao, M.; Luo, S. Insecticidal terpenes from the essential oils of Artemisia nakaii and their inhibitory effects on Acetylcholinesterase. Front. Plant Sci. 2021, 12, 720816. [Google Scholar] [CrossRef] [PubMed]
- Scalerandi, E.; Flores, G.A.; Palacio, M.; Defagó, M.T.; Carpinella, M.C.; Valladares, G.; Bertoni, A.; Palacios, S.M. Understanding synergistic toxicity of terpenes as insecticides: Contribution of metabolic detoxification in Musca domestica. Front. Plant Sci. 2018, 871, 1579. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).