
Influence of Atmospheric Cold Plasma Exposure on Naturally Present Fungal Spores and Physicochemical Characteristics of Sundried Tomatoes (Solanum lycopersicum L.)

 , , ,
, , ,  ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Plasma Treatments
2.2. Changes in the Natural Microbiota Associate to Sundried Tomatoes after ACP
Phenotypical and Molecular Identification of Filamentous Fungi
2.3. Preparation of Spore Suspensions
Spore Viability after ACP Treatment
2.4. Moisture, Water Activity and pH
2.5. Color Analysis
2.6. Lycopene Determination
2.7. Determination of the Antioxidant Capacity
2.8. Statistical Analysis
3. Results
3.1. Plasma Gas and Reactive Species Analysis
3.2. Effect of Atmospheric Cold Plasma on the Total Mesophilic Aerobic Bacteria and Filamentous Fungi Count Naturally Present in Sundried Tomatoes
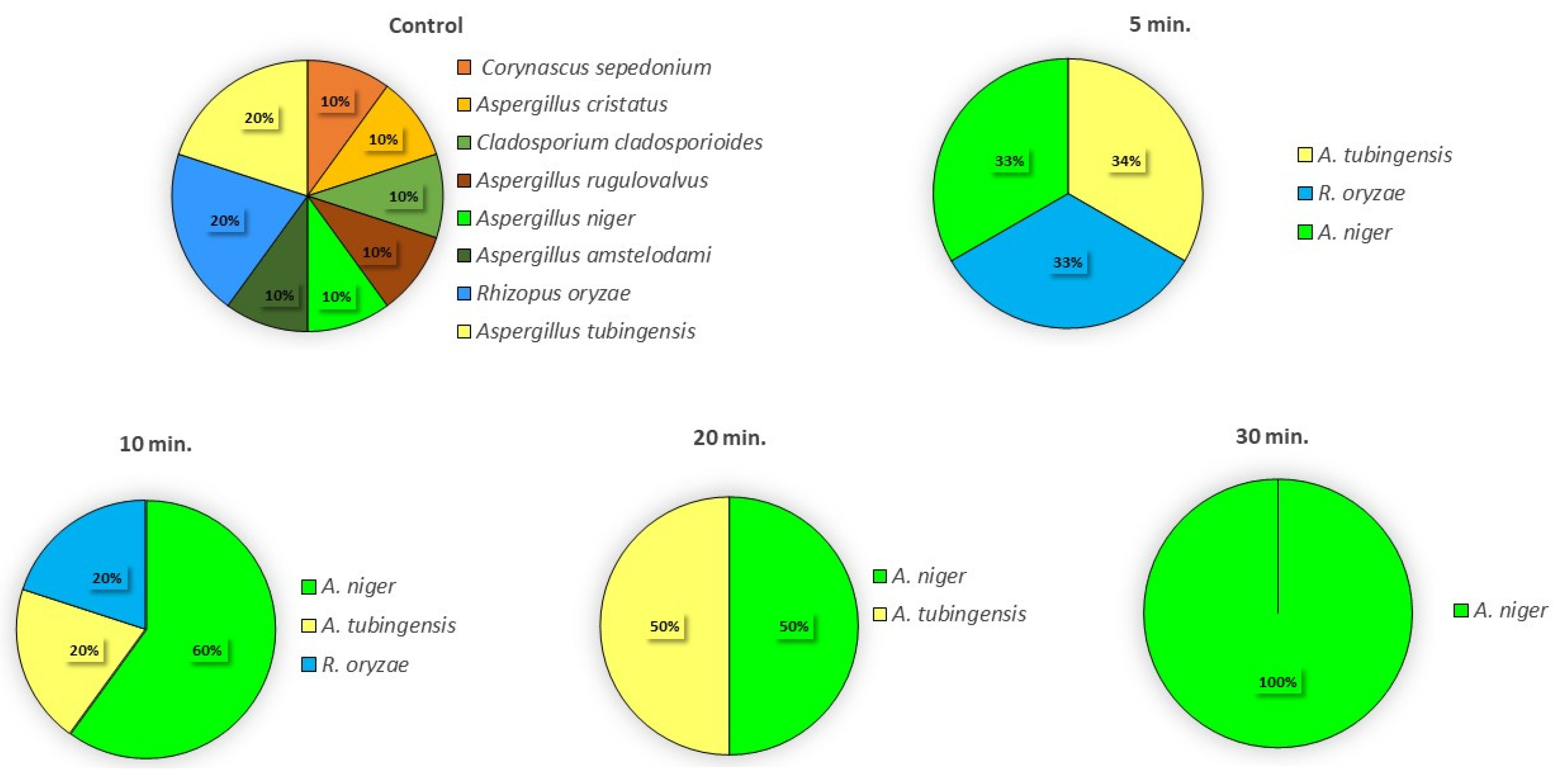
3.3. Effect of the CAP on the Fungal Species
3.4. Effects on Physicochemical Characteristics of Sundried Tomatoes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. News Archive 2011. Available online: https://www.fao.org/news/archive/news-by-date/2011/en/ (accessed on 1 October 2021).
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Hegazy, E.M. Mycotoxin and fungal contamination of fresh and dried tomato. Annu. Res. Rev. Biol. 2017, 17, 1–9. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Statistics. Available online: https://www.fao.org/food-agriculture-statistics/en/ (accessed on 15 October 2021).
- FAOSTAT. The State of Food and Agriculture. Available online: https://www.fao.org/3/ca6030en/ca6030en.pdf (accessed on 15 October 2021).
- Sanzani, S.M.; Gallone, T.; Garganese, F.; Caruso, A.G.; Amenduni, M.; Ippolito, A. Contamination of fresh and dried tomato by Alternaria toxins in southern Italy. Food Addit. Contam. Part A 2019, 36, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Doymaz, I. Air-drying characteristics of tomatoes. J. Food Eng. 2007, 78, 1291–1297. [Google Scholar] [CrossRef]
- Kakde, U.B.; Kakde, H.U. Incidence of post-harvest disease and airborne fungal spores in a vegetable market. Acta Bot. Croat. 2012, 71, 147–157. [Google Scholar] [CrossRef][Green Version]
- Sephton-Clark, P.C.S.; Muñoz, J.F.; Ballou, E.R.; Cuomo, C.A.; Voelz, K. Pathways of Pathogenicity: Transcriptional Stages of Germination in the Fatal Fungal Pathogen Rhizopus delemar. mSphere 2018, 3, e00403-18. [Google Scholar] [CrossRef] [PubMed]
- Dwivedy, A.K.; Prakash, B.; Chanotiya, C.S.; Bisht, D.; Dubey, N.K. Chemically characterized Mentha cardiaca L. essential oil as plant based preservative in view of efficacy against biodeteriorating fungi of dry fruits, aflatoxin secretion, lipid peroxidation and safety profile assessment. Food Chem. Toxicol. 2017, 106, 175–184. [Google Scholar] [CrossRef]
- Ndagijimana, M.; Chaves-López, C.; Corsetti, A.; Tofalo, R.; Sergi, M.; Paparella, A.; Guerzoni, M.E.; Suzzi, G. Growth and metabolites production by Penicillium brevicompactum in yoghurt. Int. J. Food Microbiol. 2008, 127, 276–283. [Google Scholar] [CrossRef]
- Cappato, L.P.; Ferreira, M.V.S.; Guimaraes, J.T.; Portela, J.B.; Costa, A.L.R.; Freitas, M.Q.; Cunha, R.L.; Oliveira, C.A.F.; Mercali, G.D.; Marzack, L.D.F.; et al. Ohmic heating in dairy processing: Relevant aspects for safety and quality. Trends Food Sci. Technol. 2017, 62, 104–112. [Google Scholar] [CrossRef]
- Amaral, G.V.; Silva, E.K.; Cavalcanti, R.N.; Cappato, L.P.; Guimaraes, J.T.; Alvarenga, V.O.; Esmerino, E.A.; Portela, J.B.; Sant’ Ana, A.S.; Freitas, M.Q.; et al. Dairy processing using supercritical carbon dioxide technology: Theoretical fundamentals, quality and safety aspects. Trends Food Sci. Technol. 2017, 64, 94–101. [Google Scholar] [CrossRef]
- Alvarez, I.; Niemira, B.A.; Fan, X.; Sommers, C.H. Inactivation of Salmonella serovars in liquid whole egg by heat following irradiation treatments. J. Food Prot. 2006, 69, 2066–2074. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Olanya, O.M.; Niemira, B.A.; Phillips, J.G. Effects of gamma irradiation on the survival of Pseudomonas fluorescens inoculated on romaine lettuce and baby spinach. LWT 2015, 62, 55–61. [Google Scholar] [CrossRef]
- Berrios-Rodriguez, A.; Olanya, O.M.; Annous, B.A.; Cassidy, J.M.; Orellana, L.; Niemira, B.A. Survival of Salmonella Typhimurium on soybean sprouts following treatments with gaseous chlorine dioxide and biocontrol Pseudomonas bacteria. Food Sci. Biotechnol. 2017, 26, 513–520. [Google Scholar] [CrossRef]
- Misra, N.N.; Pankaj, S.K.; Segat, A.; Ishikawa, K. Cold plasma interactions with enzymes in foods and model systems. Trends Food Sci. Technol. 2016, 55, 39–47. [Google Scholar] [CrossRef]
- Misra, N.N.; Jo, C. Applications of cold plasma technology for microbiological safety in meat industry. Trends Food Sci. Technol. 2017, 64, 74–86. [Google Scholar] [CrossRef]
- Pankaj, S.K.; Keener, K.M. Cold plasma: Background, applications and current trends. Curr. Opin. Food Sci. 2017, 16, 49–52. [Google Scholar] [CrossRef]
- Munitz, M.S.; Garrido, C.E.; Gonzalez, H.H.L.; Resnik, S.L.; Salas, P.M.; Montti, M.I.T. Mycoflora and Potential Mycotoxin Production of Freshly Harvested Blueberry in Concordia, Entre Ríos Province, Argentina. Int. J. Fruit Sci. 2013, 13, 312–325. [Google Scholar] [CrossRef]
- Delgado-Ospina, J.; Molina-Hernandez, J.B.; Maggio, F.; Fernández-Daza, F.; Sciarra, P.; Paparella, A.; Chaves-López, C. Advances in understanding the enzymatic potential and production of ochratoxin A of moulds isolated from cocoa fermented beans. Food Microbiol. 2022, in press. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Makhlouf, J.; Carvajal-Campos, A.; Querin, A.; Tadrist, S.; Puel, O.; Lorber, S.; Oswald, I.P.; Hamze, M.; Bailly, J.D.; Bailly, S. Morphologic, molecular and metabolic characterization of Aspergillus section Flavi in spices marketed in Lebanon. Sci. Rep. 2019, 9, 5263. [Google Scholar] [CrossRef]
- Ali, A.; Muhammad, M.T.M.; Sijam, K.; Siddiqui, Y. Potential of chitosan coating in delaying the postharvest anthracnose (Colletotrichum gloeosporioides Penz.) of Eksotika II papaya. Int. J. Food Sci. Technol. 2010, 45, 2134–2140. [Google Scholar] [CrossRef]
- Molina-Hernandez, J.B.; Aceto, A.; Bucciarelli, T.; Paludi, D.; Valbonetti, L.; Zilli, K.; Scotti, L.; Chaves-López, C. The membrane depolarization and increase intracellular calcium level produced by silver nanoclusters are responsible for bacterial death. Sci. Rep. 2021, 11, 21557. [Google Scholar] [CrossRef]
- Horwitz, W. Official Methods of Analysis International, Agricultural Chemicals, Contaminants, Drugs, 15th ed.; AOAC 925.10; AOAC: Rockville, MD, USA, 1990. [Google Scholar]
- Salder, G.; Davis, J.; Dezman, D. Rapid Extraction of Lycopene and β-Carotene from Reconstituted Tomato Paste and Pink Grapefruit Homogenates. J. Food Sci. 1990, 55, 1460–1461. [Google Scholar] [CrossRef]
- Souza, A.L.R.; Hidalgo-Chávez, D.W.; Pontes, S.M.; Gomes, F.S.; Cabral, L.M.C.; Tonon, R.V. Microencapsulation by spray drying of a lycopene-rich tomato concentrate: Characterization and stability. LWT—Food Sci. Technol. 2018, 91, 286–292. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A.J. Colorimetry to total phenolics with phosphomolybdic acid reagents. Am. J. Enol. Vinic. 1965, 16, 144–158. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Min Yang, A.; Rice-Evans, C. Antioxidant activity applying an improved abts radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Shimizu, T.; Sakiyama, Y.; Graves, D.B.; Zimmermann, J.L.; Morfill, G.E. The dynamics of ozone generation and mode transition in air surface micro-discharge plasma at atmospheric pressure. New J. Phys. 2012, 14, 103028. [Google Scholar] [CrossRef]
- Pavlovich, M.J.; Clarck, D.S.; Graves, D.B. Quantification of air plasma chemistry for surface disinfection. Plasma Sources Sci. Technol. 2014, 23, 065036. [Google Scholar] [CrossRef]
- Eliasson, B.; Hirth, M.; Kogelschatz1, U. Ozone synthesis from oxygen in dielectric barrier discharges. J. Phys. D Appl. Phys. 1987, 20, 1421. [Google Scholar] [CrossRef]
- Kogelschatz, U.; Eliasson, B.; Hirth, M. Ozone generation from oxygen and air: Discharge physics and reactions mechanisms. Ozone Sci. Eng. 1987, 10, 367–368. [Google Scholar] [CrossRef]
- Finn, S.; Condell, O.; McClure, P.; Amézquita, A.; Fanning, S. Mechanisms of survival, responses, and sources of salmonella in low-moisture environments. Front. Microbiol. 2013, 4, 331. [Google Scholar] [CrossRef] [PubMed]
- Puligundla, P.; Mok, C. Inactivation of spores by nonthermal plasmas. World J. Microbiol. Biotechnol. 2018, 34, 143. [Google Scholar] [CrossRef]
- Dobrynin, D.; Fridman, G.; Mukhin, Y.V.; Wynosky-Dolfi, M.A.; Rieger, J.; Rest, R.F.; Gutsol, A.F.; Fridman, A. Cold plasma inactivation of Bacillus cereus and Bacillus anthracis (anthrax) spores. IEEE Trans. Plasma Sci. 2010, 38, 1878–1884. [Google Scholar] [CrossRef]
- Hertwig, C.; Reineke, K.; Rauh, C.; Schlüter, O. Factors involved in Bacillus spore’s resistance to cold atmospheric pressure plasma. Innov. Food Sci. Emerg. Technol. 2017, 43, 173–181. [Google Scholar] [CrossRef]
- Young, S.B.; Setlow, P. Mechanisms of Bacillus subtilis spore resistance to and killing by aqueous ozone. J. Appl. Microbiol. 2004, 96, 1133–1142. [Google Scholar] [CrossRef]
- Patil, S.; Moiseev, T.; Misra, N.N.; Cullen, P.J.; Mosnier, J.P.; Keener, K.M.; Bourke, P. Influence of high voltage atmospheric cold plasma process parameters and role of relative humidity on inactivation of Bacillus atrophaeus spores inside a sealed package. J. Hosp. Infect. 2014, 88, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Gavahian, M.; Peng, H.J.; Chu, Y.H. Efficacy of cold plasma in producing Salmonella-free duck eggs: Effects on physical characteristics, lipid oxidation, and fatty acid profile. J. Food Sci. Technol. 2019, 56, 5271–5281. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, A.; Niemira, B.A.; Gurtler, J.B.; Fan, X.; Sites, J.; Boyd, G.; Chen, H. Atmospheric cold plasma inactivation of aerobic microorganisms on blueberries and effects on quality attributes. Food Microbiol. 2015, 46, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Ulbin-Figlewicz, N.; Jarmoluk, A. Effect of low-pressure plasma treatment on the color and oxidative stability of raw pork during refrigerated storage. Food Sci. Technol. Int. 2016, 22, 313–324. [Google Scholar] [CrossRef]
- Ziuzina, D.; Patil, S.; Cullen, P.J.; Keener, K.M.; Bourke, P. Atmospheric cold plasma inactivation of Escherichia coli, Salmonella enterica serovar Typhimurium and Listeria monocytogenes inoculated on fresh produce. Food Microbiol. 2014, 42, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Butscher, D.; Zimmermann, D.; Schuppler, M.; von Rohr, P.R. Plasma inactivation of bacterial endospores on wheat grains and polymeric model substrates in a dielectric barrier discharge. Food Control 2016, 60, 636–645. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, D.U.; Min, S.C. Microbial decontamination of red pepper powder by cold plasma. Food Microbiol. 2014, 38, 128–136. [Google Scholar] [CrossRef]
- Iseki, S.; Ohta, T.; Aomatsu, A.; Ito, M.; Kano, H.; Higashijima, Y.; Hori, M. Rapid inactivation of Penicillium digitatum spores using high-density nonequilibrium atmospheric pressure plasma. Appl. Phys. Lett. 2010, 96, 153704. [Google Scholar] [CrossRef]
- Yaguchi, T.; Horie, Y.; Tanaka, R.; Matsuzawa, T.; Ito, J.; Nishimura, K. Molecular phylogenetics of multiple genes on Aspergillus section Fumigati isolated from clinical specimens in Japan. Jpn. J. Med. Mycol. 2007, 48, 37–46. [Google Scholar] [CrossRef]
- Trompeter, F.J.; Neff, W.J.; Franken, O.; Heise, M.; Neiger, M.; Liu, S.; Pietsch, G.J.; Saveljew, A.B. Reduction of Bacillus Subtilis and Aspergillus Niger spores using nonthermal atmospheric gas discharges. IEEE Trans. Plasma Sci. 2002, 30, 1416–1423. [Google Scholar] [CrossRef]
- Ott, L.C.; Appleton, H.J.; Shi, H.; Keener, K.; Mellata, M. High voltage atmospheric cold plasma treatment inactivates Aspergillus flavus spores and deoxynivalenol toxin. Food Microbiol. 2021, 95, 103669. [Google Scholar] [CrossRef]
- Oliveira, B.R.; Marques, A.P.; Ressurreição, M.; Moreira, C.J.S.; Pereira, C.S.; Crespo, M.T.B.; Pereira, V.J. Inactivation of Aspergillus species in real water matrices using medium pressure mercury lamps. J. Photochem. Photobiol. B Biol. 2021, 221, 112242. [Google Scholar] [CrossRef] [PubMed]
- Misra, N.N.; Yadav, B.; Roopesh, M.S.; Jo, C. Cold Plasma for Effective Fungal and Mycotoxin Control in Foods: Mechanisms, Inactivation Effects, and Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.J.B.; Casadevall, A. Functions of fungal melanin beyond virulence. Fungal Biol. Rev. 2017, 31, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Montie, T.C.; Kelly-Wintenberg, K.; Roth, J.R. An overview of research using the one atmosphere uniform glow discharge plasma (OAUGDP) for sterilization of surfaces and materials. IEEE Trans. Plasma Sci. 2000, 28, 41–50. [Google Scholar] [CrossRef]
- Hopke, A.; Brown, A.J.P.; Hall, R.A.; Wheeler, R.T. Dynamic Fungal Cell Wall Architecture in Stress Adaptation and Immune Evasion. Trends Microbiol. 2018, 26, 284–295. [Google Scholar] [CrossRef]
- Ambrico, P.F.; Šimek, M.; Rotolo, C.; Morano, M.; Minafra, A.; Ambrico, M.; Pollastro, S.; Gerin, D.; Faretra, F.; De Miccolis Angelini, R.M. Surface Dielectric Barrier Discharge plasma: A suitable measure against fungal plant pathogens. Sci. Rep. 2020, 10, 3673. [Google Scholar] [CrossRef]
- Bao, Y.; Reddivari, L.; Huang, J.Y. Development of cold plasma pretreatment for improving phenolics extractability from tomato pomace. Innov. Food Sci. Emerg. Technol. 2020, 65, 102445. [Google Scholar] [CrossRef]
- Starek, A.; Pawłat, J.; Chudzik, B.; Kwiatkowski, M.; Terebun, P.; Sagan, A.; Andrejko, D. Evaluation of selected microbial and physicochemical parameters of fresh tomato juice after cold atmospheric pressure plasma treatment during refrigerated storage. Sci. Rep. 2019, 9, 8407. [Google Scholar] [CrossRef]
- Ramazzina, I.; Berardinelli, A.; Rizzi, F.; Tappi, S.; Ragni, L.; Sacchetti, G.; Rocculi, P. Effect of cold plasma treatment on physico-chemical parameters and antioxidant activity of minimally processed kiwifruit. Postharvest Biol. Technol. 2015, 107, 55–65. [Google Scholar] [CrossRef]
- Tappi, S.; Ramazzina, I.; Rizzi, F.; Sacchetti, G.; Ragni, L.; Rocculi, P. Effect of plasma exposure time on the polyphenolic profile and antioxidant activity of fresh-cut apples. Appl. Sci. 2018, 8, 1939. [Google Scholar] [CrossRef]
- Dasan, B.G.; Boyaci, I.H. Effect of Cold Atmospheric Plasma on Inactivation of Escherichia coli and Physicochemical Properties of Apple, Orange, Tomato Juices, and Sour Cherry Nectar. Food Bioprocess Technol. 2018, 11, 334–343. [Google Scholar] [CrossRef]
- Li, X.; Li, M.; Ji, N.; Jin, P.; Zhang, J.; Zheng, Y.; Zhang, X.; Li, F. Cold plasma treatment induces phenolic accumulation and enhances antioxidant activity in fresh-cut pitaya (Hylocereus undatus) fruit. LWT 2019, 115, 108447. [Google Scholar] [CrossRef]
- Alothman, M.; Bhat, R.; Karim, A.A. UV radiation-induced changes of antioxidant capacity of fresh-cut tropical fruits. Innov. Food Sci. Emerg. Technol. 2009, 10, 512–516. [Google Scholar] [CrossRef]
- Ozen, E.; Singh, R.K. Atmospheric cold plasma treatment of fruit juices: A review. Trends Food Sci. Technol. 2020, 103, 144–151. [Google Scholar] [CrossRef]
- Hoffmann, A.M.; Noga, G.; Hunsche, M. High blue light improves acclimation and photosynthetic recovery of pepper plants exposed to UV stress. Environ. Exp. Bot. 2015, 109, 254–263. [Google Scholar] [CrossRef]
- Sarangapani, C.; O’Toole, G.; Cullen, P.J.; Bourke, P. Atmospheric cold plasma dissipation efficiency of agrochemicals on blueberries. Innov. Food Sci. Emerg. Technol. 2017, 44, 235–241. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Gene Target | Length bp | Primer | Sequences (5′ → 3′) | Reference |
|---|---|---|---|---|
| Internal transcribed Spacer (ITS) | 420–825 | ITS1 (F) | 5′TCCGTAGGTGAACCTGCGG3′ | [22] |
| ITS4 (R) | 5′TCCTCCGCTTATTGATATGC3′ | |||
| β-tubulin (BenA) | 1125 | β-tub 2a (F) | 5′GGTAACCAAATCGGTGCTTTC3′ | [22] |
| β-tub 2b (R) | 5′ACCCTCAGTGTAGTGACCCTTGGC3′ | [23] | ||
| Calmodulin (CaM) | 543 | Cmd5 (F) | 5′-CCGAGTACAAGGAGGCCTTC-3′ | [23] |
| Cmd6 (R) | 5′-CCGATAGAGGTCATAACGTGG-3′ |
| Time of Treatment (min.) | Mesophilic Bacteria (log UFC/g) | Filamentous Fungi (log CFU/g) | ||
|---|---|---|---|---|
| t0 | t22 | t0 | t22 | |
| Control | 3.06 ± 0.78 a | 3.12 ±0.54 a | 1.79 ± 0.82 a | 1.56 ± 0.75 a |
| 5 | 2.95 ± 0.45 a | 3.06 ±0.34 a | 1.68 ± 0.4 a | 1.54 ± 0.56 a |
| 10 | 2.30 ± 0.50 b | 2.40 ±0.10 b | 0.77 ± 0.45 a | 0.47 ± 0.52 a |
| 20 | 2.15 ± 0.28 b | 2.30 ±0.19 b | 0.62 ± 0.6 a | 0.67 ± 0.58 a |
| 30 | 2.20 ± 0.56 b | 2.10 ±0.64 b | 0.44 ± 0.52 b | 0.51 ± 0.61 a |
| Time of Treatment (min.) | pH | Moisture Content % | aw | Lycopene (g/g dw) | FC (mg GAE/g dw) | TEAC (µmol TE/g dw) |
|---|---|---|---|---|---|---|
| 0 | 4.36 ± 0.059 a | 34.25 ± 1.57 a | 0.608 ± 0.006 a | 3.22 ± 0.05 d | 3.35 ± 0.16 a | 8.35 ± 0.55 ab |
| 5 | 4.34 ± 0.053 a | 33.29 ± 1.37 a | 0.610 ± 0.005 a | 3.58 ± 0.21 c | 3.58 ± 0.24 a | 7.58 ± 0.42 b |
| 10 | 4.30 ± 0.075 a | 29.00 ± 0.28 b | 0.610 ± 0.004 a | 4.13 ± 0.26 b | 3.57 ± 0.24 a | 9.28 ± 0.66 a |
| 20 | 4.25 ± 0.079 a | 29.86 ± 0.29 b | 0.611 ± 0.004 a | 4.57 ± 0.03 a | 3.35 ± 0.05 a | 7.51 ± 0.91 b |
| 30 | 4.29 ± 0.076 a | 32.43 ± 0.36 a | 0.613 ± 0.007 a | 4.14 ± 0.11 b | 3.48 ± 0.13 a | 7.89 ± 0.71 ab |
| Time of Treatment (min) | L* | a* | b* | C* | h° |
|---|---|---|---|---|---|
| 0 | 29.20± 3.64 a | 19.74 ± 3.75 a | 15.20 ± 3.29 a | 24.94 ± 4.81 a | 37.48 a ± 3.14 a |
| 5 | 26.03 ± 5.79 b | 16.98 ± 4.76 a | 14.49 ± 5.25 a | 22.37 ± 6.92 a | 39.54 a ±4.47 a |
| 10 | 25.10 ± 3.27 b | 18.23 ± 4.68 a | 14.38 ± 3.94 a | 23.27 ± 5.92 a | 38.16 a ± 3.79 a |
| 20 | 24.07 ± 4.52 b | 17.45 ± 2.94 a | 12.91 ± 2.69 a | 21.71 a ± 3.93 a | 36.35 a ± 1.83 a |
| 30 | 25.92 ± 4.76 b | 18.63 ± 2.46 a | 14.72 ± 3.27 a | 23.78 a ± 3.81 a | 38.01 a ± 3.42 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molina-Hernandez, J.B.; Laika, J.; Peralta-Ruiz, Y.; Palivala, V.K.; Tappi, S.; Cappelli, F.; Ricci, A.; Neri, L.; Chaves-López, C. Influence of Atmospheric Cold Plasma Exposure on Naturally Present Fungal Spores and Physicochemical Characteristics of Sundried Tomatoes (Solanum lycopersicum L.). Foods 2022, 11, 210. https://doi.org/10.3390/foods11020210
Molina-Hernandez JB, Laika J, Peralta-Ruiz Y, Palivala VK, Tappi S, Cappelli F, Ricci A, Neri L, Chaves-López C. Influence of Atmospheric Cold Plasma Exposure on Naturally Present Fungal Spores and Physicochemical Characteristics of Sundried Tomatoes (Solanum lycopersicum L.). Foods. 2022; 11(2):210. https://doi.org/10.3390/foods11020210
Chicago/Turabian StyleMolina-Hernandez, Junior Bernardo, Jessica Laika, Yeimmy Peralta-Ruiz, Vinay Kumar Palivala, Silvia Tappi, Filippo Cappelli, Antonella Ricci, Lilia Neri, and Clemencia Chaves-López. 2022. "Influence of Atmospheric Cold Plasma Exposure on Naturally Present Fungal Spores and Physicochemical Characteristics of Sundried Tomatoes (Solanum lycopersicum L.)" Foods 11, no. 2: 210. https://doi.org/10.3390/foods11020210
APA StyleMolina-Hernandez, J. B., Laika, J., Peralta-Ruiz, Y., Palivala, V. K., Tappi, S., Cappelli, F., Ricci, A., Neri, L., & Chaves-López, C. (2022). Influence of Atmospheric Cold Plasma Exposure on Naturally Present Fungal Spores and Physicochemical Characteristics of Sundried Tomatoes (Solanum lycopersicum L.). Foods, 11(2), 210. https://doi.org/10.3390/foods11020210