Abstract
The increasing incidence of food allergy is becoming a substantial public health concern. Increasing evidence suggests that alterations in the composition of the intestinal microbiota play a part in the development of food allergy. Additionally, the application of probiotics to correct gut microbiota imbalances and regulate food allergy has become a research hotspot. However, the mechanism by which the gut microbiota regulates food allergy and the efficacy of probiotics are still in the preliminary exploration stage, and there are no clear and specific conclusions. The aim of this review is to provide information regarding the immune mechanism underlying food allergy, the correlation between the intestinal microbiota and food allergy, a detailed description of causation, and mechanisms by which the intestinal microbiota regulates food allergy. Subsequently, we highlight how probiotics modulate the gut microbiome–immune axis to alleviate food allergy. This study will contribute to the dovetailing of bacterial therapeutics with immune system in allergic individuals to prevent food allergy and ameliorate food allergy symptoms.
1. Introduction
Food allergy is a repeatable atopic disorder resulting from exposure to specific food allergens that can lead to life-threatening allergic reactions. Nowadays, food allergy is becoming increasingly common and represents a serious public health issue which affects approximately 10% of children and 4% of adults worldwide [1]. The allergy often presents as atopic dermatitis, vomiting, diarrhea, and other adverse reactions, potentially leading to anaphylactic shock and death [2]. Therefore, the pathogenesis and treatment of food allergy have attracted more and more attention.
Food allergens are digested and absorbed in the gut, mainly in the small intestine, which contains symbiotic microbiota. With the co-evolution of humans and microbiota, symbiotic microbes inevitably have a considerable impact on human health. Thus, with its high level of microbial richness, the intestinal cavity presents a highly tolerant homeostatic environment in which immune cells respond favorably to food allergens. It has been proven that the human intestinal microbiota, especially the ileum and colon, plays an important role in the intestinal mucosal immunity, by promoting local homeostatic interactions and regulating the immune response to food allergens in the peripheral mucosa [3]. However, the type and number of symbiotic microbiota in the gut may vary and may be affected by the external environment [4]. For example, changes in the dietary patterns, antibiotics use, delivery modes, breastfeeding, vaccines, and pathogen exposure can modify the formation of the intestinal microbiota. Further experimental and epidemiological studies suggest that the gut microbiota composition is related to the clinical trajectory of food allergy and that early life (0–6 month) is a critical period for gut microbial colonization [5]. Several gut microbial metabolites, such as short-chain fatty acids (SCFAs), secondary bile acids (BAs), and amphoteric polysaccharide A (PSA), have been shown to directly or indirectly regulate peripheral induced regulatory T cells (Tregs) differentiation, most of which express the retinoic acid receptor (RAR)-associated orphan receptor γt (RORγt) [6]. RORγt is a master regulator of interleukin (IL)-22-producing immune cells in the gut and can enhance the intestinal epithelial barrier. Furthermore, Feehley et al. (2019) transplanted germ-free mice with feces from healthy or cow-milk-allergic (CMA) infants and found that those mice colonized with healthy infants’ microbiota were protective against allergic reactions to β-lactoglobulin (the major allergen in milk) [7]. Taken together, we can draw a conclusion that there is a strong relationship between the gut microbiota and food allergy, but the exact regulatory mechanism remains unclear. The purpose of this review is to provide information about the immune mechanisms underlying food allergy, the correlation between the gut microbiota and food allergy, and the mechanisms of gut microbiota in regulating food allergy. We also discuss how probiotics can alleviate food allergy by modulating the gut microbiota.
2. Food Allergy
According to the different causes, food allergy can generally be divided into two broad categories: immunoglobulin (Ig) E-mediated food allergy and non-IgE-mediated food allergy. In most cases, the immune mechanism of allergic reactions involves an adaptive T helper 2 (Th2)-type response, which subsequently leads to the production of allergens-specific IgE antibodies. The mechanisms of food allergic reaction are shown in Figure 1. The chain of events that occurs in an allergic reaction begins when the allergens are presented to the gastrointestinal immune system, resulting in the production of specific IgE antibodies and damage to the intestinal epithelium. When food allergens pass through the intestinal epithelium again, their infiltration is enhanced. Cytokines such as thymic stromal lymphopoietin, interleukin (IL)-25, and IL-33, which are generated by intestinal epithelial cells, induce OX40L expression in CD103+ dendritic cells (DCs) after injury, infection, and immunoactivation, which is beneficial for Th2 cells differentiation. In response, activated Th2 cells release cytokines, including IL-4, IL-5, and IL-13, thus promoting the immunoglobulin class switch of B cells and differentiating into IgE-secreting plasma cells [8].
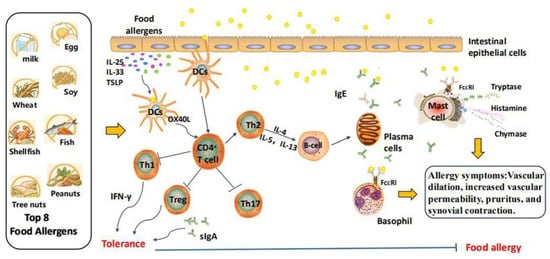
Figure 1.
The possible mechanism of food allergy. Food allergens are digested into peptides in the gut through the epithelial barrier, which are captured by DCS and presented to naive CD4+ T cells. Naive CD4+ T cells then differentiate into the Th-2 phenotype, producing IL-4, IL-5, and IL-13. Among them, IL-4 and IL-13 induce B cells to produce allergen-specific IgE antibodies, which bind to FcεRI receptors on the surface of basophils and mast cells, triggering degranulation and the release of chemical mediators such as histamine. TSLP: thymic stromal lymphopoietin; I-: inhibition.
Through the intestinal epithelium of allergic patients, food allergens are exposed to immune cells mainly located in the mucosa and blood vessels, such as mast cells and basophils, and bind to IgE and its high-affinity immunoglobulin gamma Fc receptor (FcεRI). These interactions lead to the activation of effector T cells and differentiation into Th0 cells, which are stimulated by IL-4 secreted by DCs and further differentiate into Th2 cells. On the surface of mast cells, FcεRI cross-linking triggers a signaling cascade starting from the tyrosine-protein kinase SYK and induces the extracellular secretion of hypersensitive mediators containing histamine, trypsin, and chymosin. These small molecules can induce physiological allergic reactions such as vascular dilatation, increased vascular permeability, pruritus, and synovial contraction, which manifest in the gastrointestinal tract as vascular edema, intestinal congestion, increased intestinal contractility and mucus secretion, acute diarrhea, etc. The factors that result in a deviation in the immune response may include symbiotic microbiota, intestinal epithelial damage, and allergens exposure in other body parts, especially the skin. For instance, many of the receptors that are reported to activate pathogen-associated molecular patterns (PAMPs), including Toll-like receptor 2 (TLR2), TLR5, TLR7, and TLR8, enhance the capacity of DCs to activate Th2 cells [9].
Despite high exposure to food allergens, only a few people experience adverse immunological reactions. The body’s immune failure to respond to food allergens is called immune tolerance. In general, oral tolerance is mainly mediated by CD103+ DCS, which can present allergens to primary T cells and induce Treg cells to differentiate and proliferate. Meanwhile, CD103+DCs can secrete IL-5 and IL-6 to promote the differentiation of B cells into IgA-producing plasma cells [10]. Notably, IgA enhances the barrier function of the intestinal mucosa and maintains the body’s tolerance.
Although genetic factors can cause a predisposition to food allergy, they cannot explain the rapid increase in the prevalence of food allergy; therefore, more and more attention has been paid to environmental factors, such as way of birth, breastfeeding, use of antibiotics, urban or rural settings, intake of junk food or low-fiber, high-fat diets, exposure to pets, etc. All these environmental factors mainly act to regulate the composition and function of gut microbiota. Growing evidence supports the possible role of the gut microbiota in the pathogenesis and progression of food allergy.
3. Effect of Intestinal Microbiota on Mucosal Immune System
The human gut contains a wide variety of microbiota, collectively referred to as intestinal microbiota, which includes bacteria, archaea, eukaryotes, and related viruses. Bacteroidetes and Firmicutes account for more than 90% of the total gut flora, among which obligate anaerobic bacteria is approximately 1000 times the number of aerobic bacteria [11]. The intestinal microbiota of healthy individuals is composed in a certain proportion, and the bacteria are mutually restricted and interdependent to maintain a certain amount and proportion of ecological balance, which plays an important role in the body’s immune response regulation. After birth, newborns are colonized by symbiotic microbiota from the mother’s skin, feces, vagina, and breast milk. Bifidobacterium and Lactobacillus from the vagina and breast milk are the main genera that colonize the gastrointestinal tract in the first 3 months of life, with Bifidobacterium present at the highest abundance [12]. Subsequently, the gut microbial diversity increases after weaning. In adulthood, the main gut bacteria are Firmicutes, Bacteroides, and Actinomycetes. With aging, the abundance of Firmicutes and Bifidobacterium decreases, while the abundance of Bacteroides and Proteus increases. The gastrointestinal tract provides a good environment for the growth, metabolism, and reproduction of various facultative and strict anaerobic bacteria [13]. These colonized or transient bacteria rely on nutrients in the gut tract and impact the renewal of gut tissue cells and the immune response [14].
The intestinal mucosal barrier comprises the mucous layer, gut microbiota, and intestinal immune system, and is affected by the integrity of intestinal epithelial cells. The interaction between the intestinal microbiota and the gut barrier maintains relative stability. Notably, the intestinal microbiota establishes the gut immune response and natural defense system, with the latter playing an important role in maintaining the dynamic balance of the gut immune system. Previous studies have confirmed that the microbiota and its community structure affect human health and disease by acting on mucosal barriers. Jakobsson et al. (2015) found that the number of mucus-secreting goblet cells in the cecum of germ-free mice was significantly lower than that of normal mice, and the mucus layer was more unstable [15]. Subsequent studies showed that germ-free mice have abnormal gut morphology and structure, such as decreased total gut area, shortened ileum villi, and a decrease in the number of gut crypts, due to the lack of gut microbiota [16]. Additionally, intestinal microbiota can affect the expression levels of microRNAs in intestinal epithelial cells; however, the relevant mechanism remains unclear. Under normal physiological conditions, the intestinal microbiota promotes the development of the host immune system through specific components such as lipopolysaccharides (LPS), lipoproteins, and metabolites, which form a biological gut barrier [17]. Lactobacillus acidophilus mainly parasitizes the gastrointestinal tract of animals, adheres to intestinal epithelial cells, and forms a biological barrier on their surface. Moreover, it reduces the pH of the gut cavity by secreting lactic acid, acetic acid, propionic acid, and other metabolites to prevent the adhesion and reproduction of pathogenic bacteria. Additionally, sodium butyrate (a gut microbial derivative) significantly improved epithelial barrier function of the HT29-MTX-E12 human colonic cell line by increasing mucin 2 (MUC2) levels at concentrations of 1–10 mM. However, sodium butyrate has no positive effect on MUC2 expression at 50–100 mM, probably due to the induction of apoptosis by higher sodium butyrate concentrations [18]. Importantly, ingestion of a butyrate-rich diet increased the expression levels of MUC2 and occludin in the colon, confirming that butyrate facilitates intestinal barrier function [19]. Treatment with butyrate also results in the down-regulation of IL-1β levels, further suggesting that it may help protect against inflammation-induced intestinal barrier damage [20]. Tryptophan metabolites of the intestinal microbiota activate the transcription factor aromatic hydrocarbon receptor (AHR), which promotes immune-cell maturation, and inhibits pathogen colonization in the intestinal tract through IL-22 [21]. The synthesis of AHR ligands oragonists or dietary supplementation of bacterial strains enhances gut barrier function by inducing the secretion of incretin and glucagon-like peptide-1. Certain specialized structures in the gut microbes, such as flagella, pili, and capsules, are often used as antigens to induce an immune response. For example, stimulation of intestinal epithelial cells with salmonella flagellin triggers upregulation of CCL20 expression and chemotaxis of immature DCs [22]. Therefore, the intestinal microbiota substantially affects the intestinal mucosal immune system.
4. Influence of Dietary Intakes on Intestinal Microbiota Composition
Although colonization and host factors influence the intestinal microbiota composition, dietary intake also plays an important role, and the bacterial populations can be altered with specific dietary interventions.
Studies have shown that a high-fat diet (HFD) alters the composition of the intestinal microbiota. For example, HFD-fed mice showed increased abundances of Firmicutes, Proteobacteria and Actinobacteria, while decreased abundances of Bacteroidetes phylum, Bifidobacterium, and Akkermansia genera compared to normal mice [23]. Meanwhile, HFD consumption in humans resulted in increased abundances of Blautia, Alistipes, Bilophila, several genera of the group Gammaproteobacteria, and decreased abundances of Roseburia, Clostridium, and Bacteroidess spp. [24]. Interestingly, gender also influenced the changes in the microbiota after HFD intake with higher abundances of Campylobacter, Blautia, Flavonifractor and Erysipelatoclostridium, while the abundances of Anaerotruncus, Eisenbergiella, Clostridiales (FamilyXIIIUCG_001) and Lachnospiraceae were higher in males [25]. Protein is a necessary ingredient in dietary and plays an important role in maintaining host health. It is worth noting that a large number of studies have confirmed that dietary protein intake also regulated the diversity of intestinal microbiota, and there are close relationships between different dietary protein sources and intestinal microbiota profiles. As for plant proteins intake, the abundances of Bifidobacterium and Lactobacillus increased; However, with the intake of animal protein, the abundances of Bifdobacterium, Roseburia, and Eubacterium rectale decreased, and the levels of Bacteroides, Bilophila, and Alistipes markedly increased [26]. Among them, Roseburia spp. is the most important and abundant bacteria that are involved in butyrate production in the intestine; Bifidobacterium is commonly found in infants’ intestine and increases in these bacteria have been suggested to relieve symptoms of IgE or Th2 allergy [27]. Similarly, animal protein-based diets have been documented to result in increased relative abundances of Enterococcus, Streptococcus, Turicibater, Escherichia, Peptostreptococcaceae, as well as Ruminococcaceaea in mice; in contrast, plant proteins-based diets enriched Bifidobacteriaceae, Desulfovibrionaceae, and Coriobacteriaceae families in mice. Notably, diets based on both animal and plant proteins showed increased abundances of lactobacilli, Lachnospiraceae, and Erysipelotrichaceae [28]. Intake of a fiber-rich diet increased the abundances of Bifidobacterium, Prevotellaceae, and Lachnospiraceae in mice, while decreasing the abundances of Porphyromonadaceae and Lactobacilli [29]. Vitamins also have the potential to modulate the intestinal microbiota. A study of vitamin A supplementation in mice found that the proportion of Escherichia-Shigella could be reduced, while the abundance of Bacteroides was significantly increased [30]. Healthy individuals with a higher intake of vitamin D had greater fecal abundance of Prevotella, and decreased amount of Haemophilus and Veillonella [31]. In addition, a recent study by Sun et al. (2018) observed that polyphenols in tea samples could induce the proliferation of certain beneficial bacteria in the intestine and inhibit the growth of harmful bacteria such as Bacteroides-Prevotella and Clostridium histolyticum [32]. Similarly, a decrease in the amount of Firmicutes and Proteobacteria phyla was observed in the gut of mice fed cocoa-derived polyphenols, while an increase in the percentage of bacteria belonging to the Tenericutes and Cyanobacteria phyla [33]. Further studies have shown that dietary polyphenols intake was inversely associated with metabolic syndrome by modulating gut microbiota [34]. Taken together, these results suggest a strong association between specific dietary intakes and intestinal microbiota composition.
5. Relationship between the Intestinal Microbiota and Food Allergy
5.1. Changes in the Intestinal Microbiota in Patients with Food Allergy
Growing evidence suggests that gut dysbiosis, an imbalance in the intestinal microbiota, plays a decisive role in the development of food allergy. Briefly, the intestinal microbiota composition of people with food allergy differs significantly from that of healthy people, and these differences are more evident in infants and children (Table 1). Christmann et al. (2015) found that children with allergic diseases (including skin, respiratory, and food allergy) exhibited a lower IgG response to specific microbial antigen clusters than healthy children [35]. A study of 166 1-year-old infants with food allergy showed that 7.2% of the patients were sensitive to one or several food allergens, and the intestinal microbiota diversity of allergic infants was significantly lower than that of non-allergic infants [36]. Further studies have shown that the number of specific bacterial groups is linked with the development of food allergy. In particular, the abundance of Lactobacillus and Bifidobacterium species in 1-week-old infants exhibited a significant negative correlation with the risk of allergy after 5 years [37]. Using 16S rRNA sequencing to compare the fecal microbial composition of allergic and healthy children and found levels of Clostridium sensu stricto and Anaerobacter in children with food allergy increased and levels of Bacteroides and Clostridium XVIII decreased [38]. The diversity of total microbiota in children with food allergy was low. Among them, the abundance of Bacteroidetes was decreased significantly and the abundance of Firmicutes was increased significantly compared with healthy children [39]. Inoue et al. (2017) found that he abundances of Dorea and Akkermansia were significantly reduced and the abundance of Veronococcus was significantly increased in the gastrointestinal tract of infants with food allergy [40]. Lee et al. (2021) speculated that increased abundance of Ruminococcaceae UCG-002 (Firmicutes phylum), as well as other dominant microbial groups, may reshape the normal gut microbial ecosystem through methane and glycerolipid metabolism pathways into an imbalance state, thereby triggering host IgE-mediated allergic responses [41]. Furthermore, Fazlollahi et al. (2018) investigated the correlation between the intestinal microbiota composition in early life and egg allergy in 141 children and found that the intestinal microbiome diversity and genera of the Lachnospiraceae and Ruminococcaceae families were related to egg allergy [42]. Bunyavanich et al. (2016) also studied the intestinal microbiota of 226 subjects with milk-allergic subjects and found a significant association between the intestinal microbiota composition, predicted metagenomic function, and remission of milk allergy only in subjects aged 3 to 6 months, whereas Clostridia and Firmicutes were enriched [43]. This finding has been confirmed in animal experiments. A recent study suggested a correlation between ileal bacteria and genes with upregulated expression levels in the ileum of healthy mice or in the ileum of mice colonized with the CMA infant microbiota, and identified a Clostridium species, Anaerostipes caccae, which prevents allergic reactions to food [7]. Decreased Clostridiales and increased Bacteroidales were also found in the gut of mice with tree nut allergy [44]. It is noteworthy that Bifidobacterium infantis and Bifidobacterium lactis, which are commonly found in the intestinal tract of nursing mothers and infants, reduce the specific IgE activity in shellfish-sensitized mice [45]. In contrast, other studies have reported no correlation between intestinal microbiota diversity and food allergy [46].

Table 1.
Correlation of gut microbiota in human subjects’ association of food allergy (↓: decrease; ↑: increase; —: no change).
In summary, these findings suggest that the intestinal microbiota is critical for modulating food allergy and suggest that regulating the microbiota community composition may be therapeutically relevant for food allergy. So far, research on the characteristics of the intestinal microbiota in patients with food allergy is still in its infancy, and no specific bacterial taxa has been identified that may be associated with the occurrence of food allergy. Additionally, the main limitation of all these studies is that the number of patients with sensitization or food allergy is small, such that statistical analyses of the effects of potential confounding variables, such as delivery modes, breastfeeding, diet, antibiotic intake, and pets, have not been possible. In addition, several studies have focused on sensitization to food rather than evaluating people with a history of allergic reactions to food or a confirmatory food challenge coupled with skin and/or serum IgE food-specific testing. Moreover, it is recommended to consider the role of the microbiota in food allergy together with the interactions between different taxa and their metabolic effects, rather than just examining bacterial diversity.
5.2. Relationship between Intestinal Microbial Metabolites and Food Allergy
The intestinal microbiota is involved in developing and regulating host physiology and immunity by directly participating in the decomposition of dietary components to synthesize and re-synthesize metabolites. Dietary fiber, composed of indigestible carbohydrates extracted from plant polysaccharides and oligosaccharides, is the main source of nutrients for intestinal bacteria, and their fermentation leads to the production of SCFAs (mainly acetic, butyric, and propionic acid). SCFAs are key intermediaries regulating mucosal and systemic immune homeostasis. Intestinal IgA, regulated by the intestinal microbiota, also plays an important role in maintaining intestinal homeostasis and normal function. The SCFAs metabolite, acetate, has been proved to promote intestinal IgA responses mediated by the “metabolite-sensing” receptor, GPR43 [47]. Treatment of mice with the SCFA, propionate, resulted in bone marrow hematopoietic alterations characterized by enhanced generation of macrophages and DC precursors via GPR41, which inhibits Th2 effector cells, thus alleviating allergic inflammation [48]. Moreover, GPR109a is required for butyrate-mediated Treg cells induction and IL-18 in the formation of immune tolerance to food allergens [49]. In addition, SCFAs may also be used as histone deacetylase (HDAC) inhibitors, to increase the acetylation of S6 kinase (S6K), and the phosphorylation of ribosomal protein S6, to regulate the mammalian target of rapamycin (mTOR) pathway, thereby promoting T cells to Th1, Th17, and Treg cell differentiation [50]. Valerate intake increases mTOR activity and production of the anti-inflammatory factor IL-10 [51]. Other studies have shown that acetate increases acetylation of the Foxp3 promoter by inhibiting HDAC9 and accelerates the production of Treg cells to alleviate allergic diseases [52]. Moreover, SCFAs, combined with G-protein coupled receptors (GPRs), can act on intestinal epithelial cells and activate NLRP3 inflammatory bodies to produce IL-18 [53]. Notably, butyric acid induces protective mechanisms, including the production of mucins and antimicrobial peptides, and increased expression levels of tight junction proteins that strengthen the intestinal epithelial barrier [54].
Emerging evidence has shown that microbiota-derived tryptophan and bile-acid metabolites also play vital roles in food allergy. Tryptophan is an essential amino acid that is degraded to serotonin, kynurenine, or indole in the gut. Supplementation of D-tryptophan produced by Bifidobacterium and Lactobacillus has been shown to inhibit allergic inflammation in the lungs by increasing intestinal microbial diversity and promoting Treg cell production [55]. Another study suggested that the occurrence of food allergy is associated with an increase in serum indolepropionic acid concentrations, while the presence of multiple food allergies is characterized by a decrease in serum kynurenine and serotonin concentrations [56]. Furthermore, indole 3-propionic acid can effectively modulate the intestinal mucosal system in mice [57]. Secondary BAs produced by intestinal bacteria have also been confirmed to play a key role in regulating intestinal mucosal tolerance by inducing Treg cell differentiation through suppression of the immunostimulatory properties of DCs. Other studies have shown that derivatives of secondary BAs are involved in regulating specific aspects of T cell responses, for example, the reduction in the number of RORγt+ Treg cells observed in food-allergic subjects is the result of insufficient microbiota production of secondary BAs metabolites, while these secondary BAs metabolites are associated with RORγt+ Treg cell differentiation [58]. Moreover, PSA promotes cytokine production in T cells by activating TLR expression on the surface of DCs and increasing cytokine production by directly binding to receptors on Tregs, thereby maintaining gut homeostasis and health [59].
To sum up, these findings suggest that SCFAs modulate food allergy by activating GPRs, inhibiting HDAC activity, as well as strengthening the protection of the intestinal epithelium barrier (Table 2).
Table 2.
Summary of proposed mechanistic actions of the effect that the gut microbial metabolites SCFAs on alleviating the food allergy.
5.3. Potential Mechanisms of Intestinal Microbiota in Regulating Food Allergy
Although the occurrence of food allergy is related to the composition of the intestinal microbiota, the exact mechanism is not clear. In this section, we describe in detail the possible mechanisms by which the intestinal microbiota modulates food allergy (Figure 2): (1) regulation of allergen uptake and (2) regulation of signaling molecules involved in immune responses.
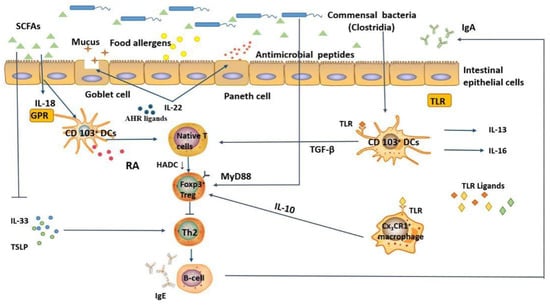
Figure 2.
Potential mechanisms of intestinal microbiota and its metabolites in regulating food allergy, (1) regulation of allergen uptake and (2) regulation of signaling molecules involved in immune responses. Under normal circumstances, strengthening the intestinal mucosal barrier driven by commensal bacteria can reduce the entry of food allergens into the systemic circulation, thereby alleviating the symptoms of food allergy. When the balance between sensitization and oral tolerance is disrupted, signals (such as TLR, AHR) can be received by intestinal epithelial cells and DCs, enhancing the epithelial barrier, promoting Foxp3+ Treg cell induction, and preventing the generation of food-specific Th2 effector populations. SCFAs, short-chain fatty acids; TLR, toll-like receptor; GPR, G-protein coupled receptor; HDAC, histone deacetylase; AHR, aryl hydrocarbon receptor.
5.3.1. Regulation of Allergen Uptake
The intestine contains hundreds of bacterial and various food allergens. Intestinal epithelial cells are physical barriers and primary sites for interactions between the body and the microbiota. Increasing evidence suggests that interactions between the intestinal microbiota and intestinal epithelial cells (mainly microfold cells, goblet cells, and Paneth cells) regulate allergen uptake and maintain the mucosal immune balance. Briefly, the gut microbiota directly interacts with intestinal epithelial cells, promotes the production of defensins, and strengthens the gut barrier function. For example, microfold cells present allergens to DCs and other antigen-presenting cells when stimulated by microbial signals [60]. In addition, the microbiota regulates allergen uptake by inducing Paneth cells to produce antimicrobial peptides and promoting goblet cells to secrete mucus [61]. It is noteworthy that CX3CR1+ cells, a suspected macrophage cell type, are generally located in the lamina propria of the intestine and are mainly involved in the uptake of allergens from the intestinal lumen. However, when the structure of the intestinal microbiota changes, these cells migrate to other sites, thereby reducing the uptake of allergens. Meanwhile, the intestinal microbiota can interact with DCs, innate immune cells, and macrophages to indirectly enhance the intestinal barrier function and regulate the uptake of food allergens through the production of IL-22.
5.3.2. Regulation of Signaling Molecules Involved in Immune Responses
Intestinal microbiota metabolites interact with host and microbiota, transmit signals to the host, and regulate the immune response. Previous studies showed that SCFAs act on DCs through GPRs and promote Th0 cells to differentiate into Foxp3+ Treg cells. Microarray analysis of mouse intestinal epithelial cells revealed that Clostridia species modulate innate lymphoid cell function and intestinal epithelial permeability to prevent allergen sensitization [62]. Foxp3+ Tregs are one of the major components involved in the body’s immune tolerance system, which can inhibit inflammatory responses and maintain oral tolerance through IL-10, IL-35, and TGF-β [63]. Moreover, Foxp3+ Treg cells also promote B cells to secrete more IgA/IgG4 into the gut lumen, thereby inhibiting Th2 cells, decreasing the concentration of Th2-type cytokines and antigen-specific IgE [64].
Recent studies of pattern recognition receptors (PRRs), such as toll-like receptors (TLRs), have elucidated the mechanism of action of the microbiota in the immune regulation of food allergy. PRRs are important part of the innate immune system of the body and exists in various forms. It is not only expressed on the cell membrane, but also widely distributed in the endosomal membrane, lysosomal membrane and cytoplasm. Their role is to monitor the presence of viral molecules, initiate the body’s inflammatory response and antiviral immune signaling pathways, and protect the host from infection. Once PRRs are activated, intestinal epithelial cells produce antimicrobial peptides and mucus to eliminate pathogens and maintain intestinal barrier function. Conversely, the TLR signaling pathway activates gut CD103+ DCs, CX3CR1+ macrophages, and RORt+ innate immune cells to secrete cytokines, promote the differentiation of Tregs, and protect against allergy [60]. A lack of TLR-mediated signaling molecules, such as TNF receptor-associated factor 6 (TRAF6), a critical downstream molecule of the TLR signaling pathway, that promotes differentiation of Th2 cells and impaired Treg cells transformation in response to allergens [65]. MyD88 is a downstream molecule involved in TLR signal transduction and inhibits food allergy through the MyD88/RORt+ pathway of regulatory T cells. However, a recent study confirmed the vital role of AHR in regulating and responding to the microbiota. One of the most prominent roles of AHR is the regulation of IL-22 expression. For example, in the absence of AHR, only small amounts IL-22 are produced, immune responses to gut pathogens are reduced, and loss of AHR signaling leads to a disbalance of T cell subsets in the gut, which subsequently changes the composition of the microbiota [66]. Likewise, other studies have demonstrated that AHR activation decreases the number of effector T cells and activates the CD103+ DC population to promote oral tolerance to food allergens [67].
Collectively, these findings indicate that various signals from the microbiota must be integrated by various cell subsets in the intestine to alter the balance between sensitization and oral tolerance. When this balance is disrupted, sensitization occurs. TLR and AHR signaling can be received by intestinal epithelial cells, DCs, macrophages, and innate lymphoid cells, enhancing the epithelial barrier, promoting Foxp3+ Treg cell induction, and preventing the generation of food-specific Th2 effector populations.
6. Research Progress on the Role of Probiotics in Relieving Allergy
Probiotics are active microbiota that positively impact human health when administered appropriately. They regulate the microbiological balance, inhibit the growth of harmful bacteria, lower blood lipids and cholesterol concentrations, maintain normal immunity of the gut mucosa, and regulate blood pressure [68]. Probiotics mainly include lactic acid bacteria, Bifidobacteria, and yeast, with lactic acid bacteria being the primary probiotic bacteria. The probiotics with the greatest potential in promoting the development of the immune system are those belonging to the genus Lactobacillus and Bifidobacterium. At present, the use of probiotics for the prevention and treatment of allergic diseases mainly focuses on maintaining the Th1/Th2 cell balance, improving intestinal barrier function, and maintaining the balance of intestinal microbiota (Table 3). Reportedly, the addition of Lacticaseibacillus rhamnosus, but not Lacticaseibacillus casei or Bifidobacterium, has been reported to be effective in accelerating the induction of oral tolerance in individuals with milk allergy [69]. Other studies have shown that supplementation with Lacticaseibacillus rhamnosus promote the abundance of butyrate-producing strains such as Lachnospiraceae and Ruminoccaceae, suggesting that probiotics can increase the abundances of tolerance-promoting microbes and promote immune tolerance to food allergens [70].
Table 3.
Summary of proposed mechanistic actions of the effect that the probiotics has on food allergy.
Cell models provide the basis for animal and clinical experiments on probiotics. Many probiotics have been shown to have potential immunomodulatory effects in immune cell models. Although in vitro cell models have some limitations, they have played a crucial role in the preliminary screening of the effects of different bacteria and metabolic components on the immune response. Cell models used to assess the immunomodulatory effects of probiotics include macrophages, human peripheral blood mononuclear cells (PBMCs), mouse spleen lymphocytes, and gut-associated lymphoid tissue. López et al. (2010) tested the specific immune activation properties of different Bifidobacterium strains (B. longum, B. breve, B. bifidum, and B. animalis subsp) co-incubated with PBMCs in vitro and found that all strains tested induced full DC maturation. However, there were differences in cytokine production levels, especially that of IL-12, IL-10, TNFα, and IL-1β. Probiotic bacteria, cytoplasm, surface extract, and fermentation supernatant also showed immunomodulatory activity, promoting CD8+ T cell activation-induced Treg differentiation [83]. In addition to immune cells, certain intestinal epithelial cells, such as Caco-2, T84, and HT-29 cells, have been widely used to study the immunomodulatory effects of probiotics. Caco-2 and HT-29 cells differ from specialized immune cells in that they are cell types that are directly exposed to the gut environment and play important roles in initiating interactions between probiotics and the host. Price et al. (2014) observed that peanut allergens altered intestinal barrier permeability and the tight junction localization in Caco-2 cell cultures [84]. Chichlowski et al. (2012) provided evidence for a specific relationship between human milk oligosaccharide-grown Bifidobacteria and intestinal epithelial cells through studies of Caco-2 and HT-29 cells [85]. Moreover, PBMCs from food allergic individuals showed decreased T84 cell barrier integrity compared to those from healthy controls [86].
Fermentation can improve the nutritional value of food by increasing the bioavailability of nutrients and reducing the amount of antinutritional factors. Moreover, fermented food can be used as a functional ingredient with high protein digestibility and probiotics. Several studies have shown that fermentation with probiotics may reduce the immunoreactivity of food allergens. Bu et al. (2010) found that Lactobacillus helveticus and Streptococcus thermophilus was the most effective combination for reducing the antigenicity of α-lactalbumin and β-lactoglobulin [71]. Other studies have shown that Lactobacillus-fermented milk interrupts allergen epitopes, produces bioactive peptides, and regulates the immune system [72]. In addition, lactic acid bacteria fermentation is more conducive to degrade IgE epitopes in wheat allergenic proteins [73]. Consistent with this finding, De Angelis et al. (2007) demonstrated that the probiotic, VSL#3, decreases the number of sensitizing proteins that cannot be degraded by pepsin and trypsin and reduces the potential allergen risk of wheat protein to a certain extent [74].
Many animal experiments and clinical studies have shown that probiotics can be used as an effective tool for allergy prevention and alleviation. Bifidobacterium species have been shown to prevent and treat egg allergic reactions by promoting the synthesis of Tregs and inhibiting Th2 cells, to increase the number of CD103+ DCs in gut-associated lymphoid tissue CD103+ DCs [75]. Fu et al. (2017) found that an oral probiotic, Bifidobacterium infantis 14.518 (Binf), effectively suppresses shrimp tropomyosin-induced anaphylaxis in mice via both preventive and therapeutic strategies [76]. Briefly, Binf was found to stimulate DCs maturation and accumulation of CD103+ tolerogenic DCs in gut-associated lymphoid tissues, which subsequently induced the differentiation of regulatory T cells and inhibited Th2-biased responses. Neau et al. (2016) identified 3 probiotics with anti-allergic properties, from a total of 31 strains, using PBMCs and Th2-skewed murine splenocytes, and these 3 strains showed a protective effect against sensitization, with decreased concentrations of β-lactoglobulin-specific IgE, IgG1, IgG2a, and mast cell protease [77]. Kozkova et al. (2014) used a mixed gavage of three Lactobacillus strains to enhance the gut barrier function; increase sIgA and TGF-β concentrations; reduce gut permeability and decrease the serum concentrations of Bet v 1-specific IgE, IgG1, IgG2a, and alleviate allergic reactions in mice allergic to birch pollen [78]. In addition, Lacticaseibacillus rhamnosus GG remarkedly improves the ovalbumin induced food allergy symptoms, most likely by reducing the ratio of IL-4/IFN-γ [79]. The probiotic, Dahi (Lactococcus lactis ssp. cremoris NCDC-86 and Lactococcus lactis ssp. lactis biovar diacetylactis NCDC-60), has been shown to skew the Th2-specific immune response toward Th1 and decrease IgE concentration in a whey protein allergy mouse model [80]. Another study showed that Bifidobacterium brevis M-16 V alters the intestinal microbiota via IL-33/ST2 signaling pathway and relieves allergic symptoms [81]. Notably, new studies have confirmed that intragastric administration of a probiotic strain, Lacticaseibacillus casei BL23, induces Foxp3+ and RORγt+ Tregs in mice [82]. A similar phenomenon was found in OVA-LPS-induced of allergic asthma inflammation [87]. Obviously, experimental animals are very different from humans in key aspects, such as the gut microbiota composition, immune function or metabolism. Thus, extrapolating the results obtained from animal models to humans may be ineffective and should always be considered as preliminary or limited.
In a clinical study, Capurso (2019) found that administration of Lacticaseibacillus rhamnosus GG or a Bifidobacterium complete hydrolysis formula significantly improves the condition of skin and ameliorates inflammatory indicators in infants with atopic eczema [88]. Jerzynska et al. (2016) confirmed clinical and immunologic effects of probiotic and vitamin D supplementation for sublingual immunotherapy, for example, in children with allergic rhinitis, where probiotic supplementation demonstrated better clinical and immunotherapeutic effects [89]. Birch pollen allergy has been associated with changes in the fecal microbiota composition, and a probiotic combination of Lactobacillus acidophilus NCFMTM (ATCC 700396) and Bifidobacterium lactis Bl-04 (ATCC SD5219) has been shown to prevent the pollen-induced eosinophil infiltration into the nasal mucosa and promote a tendency to reduce nasal symptoms [90]. However, the long-term efficacy of probiotics in the prevention and treatment of allergic disease needs to be further evaluated. Plummer et al. (2020) found that postnatal administration of probiotics had no effect on the incidence of allergic disease or atopic sensitization in preterm children during the first 2 years [91]. Moreover, West et al. (2013) reported that Lactobacillus paracasei ssp paracasei F19 had no effect on any diagnosed allergic disease, airway inflammation, or IgE sensitization [92]. It is worth noting that the results vary from study to study due to multiple factors (dose, probiotics supplement, and environment).
Taken together, many mouse models and human studies have confirmed that single probiotics, mixed probiotics, or some probiotic components have specific preventive and therapeutic effects against food allergy. These findings preliminarily demonstrate the potential of probiotics to regulate food allergy. However, the results of clinical trials of probiotics in patients with food allergy have been controversial. Although most studies have shown that probiotics help prevent food allergy, some of the experimental results of these studies are invalid, and the results have been inconsistent and unstable. In addition, most studies of probiotics have focused on the bacteria themselves, rather than host–bacteria interactions, and the mechanism of action of probiotics on food allergies remains unclear. Furthermore, most data on probiotics for the treatment of food allergy are based on animal experiments. The small number of clinical studies in this field has been limited to observational studies, and there are few studies on the mechanism of probiotics in the treatment of food allergy. Moreover, medical studies suggest that, although uncommon, the use of probiotics may have adverse effects such as the spread of inappropriate resistance genes in the gut microbial populations, virulence factors in probiotic strains, transfer to tissues and blood, inflammation reactions, and infections. Therefore, further studies are needed to explore the role and mechanism of intestinal microbes in food allergy, to select more effective strains for treatment and prevention, and promote the establishment of new methods for regulating food allergy based on the gut microecology.
7. Conclusions
Food allergy has become a food safety and public health problem globally, and as such, it has garnered increasing research attention. In recent years, numerous animal and human studies have shown that the intestinal microbiota is strongly related to food allergy. Briefly, the intestinal microbiota composition of people with food allergy differs significantly from that of healthy people, and these differences are more evident in infants and children. Interactions between the microorganisms and microbial metabolites (SCFAs, tryptophan, and bile-acid) and the host immune response are probably the mechanism by which the microorganisms influence food allergy. Studies have shown that SCFAs modulate food allergy by activating GPRs, inhibiting HDAC activity, and strengthening the protection of the intestinal epithelium barrier. The application of probiotics to correct intestinal microbiota imbalances and regulate food allergy has become a research hotspot, and the combination of probiotics with other dietary interventions has been shown to have significant effects on preventing and treating disease. However, the mechanisms by which the intestinal microbiota regulates food allergy and the efficacy of probiotics are still in the preliminary exploration stage, and there are no clear and specific conclusions. In addition, the efficacy of probiotics also depends on the composition and activity of the host intestinal flora, as well as the derived metabolites and by-products of probiotics synthesis. Thus, further investigation is warranted in this field. With advances in the study of intestinal microbiota and its involvement in food allergy, we will have a greater understanding of food allergy, and it is expected that we will discover new strategies for the prevention and treatment of food allergy.
Author Contributions
Conceptualization, R.W. and J.W.; writing—original draft preparation, H.Y., Y.Q., Y.G. and S.S.; writing—review and editing, H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Science Foundation of China (grant number 32001641), China Postdoctoral Science Foundation funded project (grant number 2021M702298), Shenyang City Youth Science and Technology Innovation Leading Talent Project (RC200495), Shenyang Science and technology innovation platform project (21-103-0-14, 21-104-0-28), and Liaoning Provincial Department of Education Project (LJKZ0651).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Martinis, M.; Sirufo, M.M.; Suppa, M.; Ginaldi, L. New Perspectives in Food Allergy. Int. J. Mol. Sci. 2022, 21, 1474. [Google Scholar] [CrossRef]
- Peters, R.L.; Mavoa, S.; Koplin, J.J. An Overview of Environmental Risk Factors for Food Allergy. Int. J. Environ. Res. Public Health 2022, 19, 722. [Google Scholar] [CrossRef]
- Khan, M.; Sori, N. Diet-Gut Microbiota-Brain Axis and IgE-Mediated Food Allergy. Microb. Gut Brain Axis 2022, 2022, 153–168. [Google Scholar] [CrossRef]
- Peroni, D.G.; Nuzzi, G.; Trambusti, I.; Di Cicco, M.E.; Comberiati, P. Microbiome Composition and Its Impact on the Development of Allergic Diseases. Front. Immunol. 2020, 11, 700. [Google Scholar] [CrossRef]
- Joseph, C.L.; Sitarik, A.R.; Kim, H.; Huffnagle, G.; Fujimura, K.; Yong, G.J.M.; Levin, A.M.; Zoratti, E.; Lynch, S.; Ownby, D.R.; et al. Infant gut bacterial community composition and food-related manifestation of atopy in early childhood. Pediatr. Allergy Immunol. 2022, 33, e13704. [Google Scholar] [CrossRef]
- Kreft, L.; Hoffmann, C.; Ohnmacht, C. Therapeutic Potential of the Intestinal Microbiota for Immunomodulation of Food Allergies. Front. Immunol. 2020, 11, 1853. [Google Scholar] [CrossRef]
- Feehley, T.; Plunkett, C.H.; Bao, R.; Hong, S.M.C.; Culleen, E.; Belda-Ferre, P.; Campbell, E.; Aitoro, R.; Nocerino, R.; Paparo, L.; et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat. Med. 2019, 25, 448–453. [Google Scholar] [CrossRef]
- Pratap, K.; Taki, A.C.; Johnston, E.B.; Lopata, A.L.; Kamath, S.D. A Comprehensive Review on Natural Bioactive Compounds and Probiotics as Potential Therapeutics in Food Allergy Treatment. Front. Immunol. 2020, 11, 996. [Google Scholar] [CrossRef]
- De Martinis, M.; Sirufo, M.M.; Viscido, A.; Ginaldi, L. Food Allergy Insights: A Changing Landscape. Arch. Immunol. Ther. Exp. 2020, 68, 1–15. [Google Scholar] [CrossRef]
- Tordesillas, L.; Berin, M.C. Mechanisms of Oral Tolerance. Clin. Rev. Allergy Immunol. 2018, 55, 107–117. [Google Scholar] [CrossRef]
- Sirufo, M.M.; De Pietro, F.; Catalogna, A.; Ginaldi, L.; De Martinis, M. The Microbiota-Bone-Allergy Interplay. Int. J. Environ. Res. Public Health 2022, 19, 282. [Google Scholar] [CrossRef]
- Stephen-Victor, E.; Crestani, E.; Chatila, T.A. Dietary and Microbial Determinants in Food Allergy. Immunity 2020, 53, 277–289. [Google Scholar] [CrossRef]
- Al-Sahlany, S.T.G.; Khassaf, W.H.; Niamah, A.K.; Al-Manhel, A.J.A. Date juice addition to bio-yogurt: The effects on physicochemical and microbiological properties during storage, as well as blood parameters in vivo. J. Saudi Soc. Agric. Sci. 2022, in press. [Google Scholar] [CrossRef]
- Martens, E.C.; Neumann, M.; Desai, M.S. Interactions of commensal and pathogenic microorganisms with the intestinal mucosal barrier. Nat. Rev. Genet. 2018, 16, 457–470. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Rodríguez-Piñeiro, A.M.; Schütte, A.; Ermund, A.; Boysen, P.; Bemark, M.; Sommer, F.; Bäckhed, F.; Hansson, G.C.; Johansson, M.E.V. The composition of the gut microbiota shapes the colon mucus barrier. EMBO Rep. 2015, 16, 164–177. [Google Scholar] [CrossRef]
- Kwon, O.; Han, T.-S.; Son, M.-Y. Intestinal Morphogenesis in Development, Regeneration, and Disease: The Potential Utility of Intestinal Organoids for Studying Compartmentalization of the Crypt-Villus Structure. Front. Cell Dev. Biol. 2020, 8, 593969. [Google Scholar] [CrossRef]
- Rotkiewicz, T.; Rotkiewicz, Z.; Depta, A.; Kander, M. Effect of Lactobacillus acidophilus and Bifidobacterium sp. on the course of Cryptosporidium parvum invasion in new-born piglets. Bull.-Vet. Inst. Pulawy 2001, 45, 187–196. [Google Scholar]
- Nielsen, D.S.G.; Jensen, B.B.; Theil, P.K.; Nielsen, T.S.; Knudsen, K.E.B.; Purup, S. Effect of butyrate and fermentation products on epithelial integrity in a mucus-secreting human colon cell line. J. Funct. Foods 2018, 40, 9–17. [Google Scholar] [CrossRef]
- Knudsen, K.E.B.; Lærke, H.N.; Hedemann, M.S.; Nielsen, T.S.; Ingerslev, A.K.; Nielsen, D.S.G.; Theil, P.K.; Purup, S.; Hald, S.; Schioldan, A.G.; et al. Impact of Diet-Modulated Butyrate Production on Intestinal Barrier Function and Inflammation. Nutrients 2018, 10, 1499. [Google Scholar] [CrossRef]
- Diao, H.; Jiao, A.R.; Yu, B.; Mao, X.B.; Chen, D.W. Gastric infusion of short-chain fatty acids can improve intestinal barrier function in weaned piglets. Genes Nutr. 2019, 14, 1–16. [Google Scholar] [CrossRef]
- Sonner, J.K.; Keil, M.; Falk-Paulsen, M.; Mishra, N.; Rehman, A.; Kramer, M.; Deumelandt, K.; Röwe, J.; Sanghvi, K.; Wolf, L.; et al. Dietary tryptophan links encephalogenicity of autoreactive T cells with gut microbial ecology. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Liu, X.; Fang, Y.; Zhou, P.; Zhang, Y.; Wang, Y. Flagellin as a vaccine adjuvant. Expert Rev. Vaccines 2018, 17, 335–349. [Google Scholar] [CrossRef]
- Coelho, O.G.L.; Cândido, F.G.; Alfenas, R.D.C.G. Dietary fat and gut microbiota: Mechanisms involved in obesity control. Crit. Rev. Food Sci. Nutr. 2019, 59, 3045–3053. [Google Scholar] [CrossRef]
- Agans, R.; Gordon, A.; Kramer, D.L.; Perez-Burillo, S.; Rufián-Henares, J.A.; Paliy, O. Dietary Fatty Acids Sustain the Growth of the Human Gut Microbiota. Appl. Environ. Microbiol. 2018, 84, e01525-18. [Google Scholar] [CrossRef]
- Bailén, M.; Bressa, C.; Martínez-López, S.; González-Soltero, R.; Lominchar, M.G.M.; Juan, C.S.; Larrosa, M. Microbiota Features Associated With a High-Fat/Low-Fiber Diet in Healthy Adults. Front. Nutr. 2020, 7, 583608. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Yao, J.; Wan, Q.; Guo, J.; Pu, F.; Shi, L.; Hu, W.; Yang, Y.; Li, L.; Li, M.; et al. Oral administration of Bifidobacterium bifidum TMC3115 to neonatal mice may alleviate IgE-mediated allergic risk in adulthood. Benef. Microbes 2018, 9, 815–828. [Google Scholar] [CrossRef]
- Kostovcikova, K.; Coufal, S.; Galanova, N.; Fajstova, A.; Hudcovic, T.; Kostovcik, M.; Prochazkova, P.; Jiraskova Zakostelska, Z.; Cermakova, M.; Sediva, B.; et al. Diet Rich in Animal Protein Promotes Pro-inflammatory Macrophage Response and Exacerbates Colitis in Mice. Front. Immunol. 2019, 10, 919. [Google Scholar] [CrossRef]
- Hashemi, Z.; Fouhse, J.; Im, H.S.; Chan, C.B.; Willing, B.P. Dietary Pea Fiber Supplementation Improves Glycemia and Induces Changes in the Composition of Gut Microbiota, Serum Short Chain Fatty Acid Profile and Expression of Mucins in Glucose Intolerant Rats. Nutrients 2017, 9, 1236. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Li, Q.; Hu, K.; He, Y.; Ai, Q.; Hu, L.; Yu, J. Vitamin A and Retinoic Acid Exhibit Protective Effects on Necrotizing Enterocolitis by Regulating Intestinal Flora and Enhancing the Intestinal Epithelial Barrier. Arch. Med. Res. 2018, 49, 1–9. [Google Scholar] [CrossRef]
- Luthold, R.V.; Fernandes, G.R.; Franco-De-Moraes, A.C.; Folchetti, L.G.; Ferreira, S.R.G. Gut microbiota interactions with the immunomodulatory role of vitamin D in normal individuals. Metabolism 2017, 69, 76–86. [Google Scholar] [CrossRef]
- Sun, H.; Chen, Y.; Cheng, M.; Zhang, X.; Zheng, X.; Zhang, Z. The modulatory effect of polyphenols from green tea, oolong tea and black tea on human intestinal microbiota in vitro. J. Food Sci. Technol. 2018, 55, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Camps-Bossacoma, M.; Pérez-Cano, F.J.; Franch, A.; Castell, M. Gut Microbiota in a Rat Oral Sensitization Model: Effect of a Cocoa-Enriched Diet. Oxidative Med. Cell. Longev. 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Gowd, V.; Karim, N.; Shishir, M.R.I.; Xie, L.; Chen, W. Dietary polyphenols to combat the metabolic diseases via altering gut microbiota. Trends Food Sci. Technol. 2019, 93, 81–93. [Google Scholar] [CrossRef]
- Christmann, B.S.; Abrahamsson, T.R.; Bernstein, C.N.; Duck, L.W.; Mannon, P.J.; Berg, G.; Björkstén, B.; Jenmalm, M.C.; Elson, C.O. Human seroreactivity to gut microbiota antigens. J. Allergy Clin. Immunol. 2015, 136, 1378–1386.e5. [Google Scholar] [CrossRef]
- Azad, M.B.; Konya, T.; Guttman, D.S.; Field, C.; Sears, M.R.; HayGlass, K.T.; Mandhane, P.J.; Turvey, S.; Subbarao, P.; Becker, A.B.; et al. Infant gut microbiota and food sensitization: Associations in the first year of life. Clin. Exp. Allergy 2015, 45, 632–643. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, M.; Dong, P. Association Between Food Allergy and Microbiome Status in 1-Year-Old Infants in STRONG Kids 2 Cohort; Illinois Library: Urbana, IL, USA, 2018; Available online: https://hdl.handle.net/2142/99865 (accessed on 1 April 2018).
- Ling, Z.; Li, Z.; Liu, X.; Cheng, Y.; Luo, Y.; Tong, X.; Yuan, L.; Wang, Y.; Sun, J.; Li, L.; et al. Altered Fecal Microbiota Composition Associated with Food Allergy in Infants. Appl. Environ. Microbiol. 2014, 80, 2546–2554. [Google Scholar] [CrossRef]
- Chen, C.; Chen, K.-J.; Kong, M.; Chang, H.; Huang, J. Alterations in the gut microbiotas of children with food sensitization in early life. Pediatr. Allergy Immunol. 2016, 27, 254–262. [Google Scholar] [CrossRef]
- Inoue, R.; Sawai, T.; Sawai, C.; Nakatani, M.; Romero-Pérez, G.A.; Ozeki, M.; Nonomura, K.; Tsukahara, T. A preliminary study of gut dysbiosis in children with food allergy. Biosci. Biotechnol. Biochem. 2017, 81, 2396–2399. [Google Scholar] [CrossRef]
- Lee, K.; Guo, J.; Song, Y.; Ariff, A.; O’Sullivan, M.; Hales, B.; Mullins, B.; Zhang, G. Dysfunctional Gut Microbiome Networks in Childhood IgE-Mediated Food Allergy. Int. J. Mol. Sci. 2021, 22, 2079. [Google Scholar] [CrossRef]
- Fazlollahi, M.; Chun, Y.; Grishin, A.; Wood, R.A.; Burks, A.W.; Dawson, P.; Jones, S.M.; Leung, D.Y.M.; Sampson, H.A.; Sicherer, S.H.; et al. Early-life gut microbiome and egg allergy. Allergy 2018, 73, 1515–1524. [Google Scholar] [CrossRef]
- Bunyavanich, S.; Shen, N.; Grishin, A.; Wood, R.; Burks, W.; Dawson, P.; Jones, S.M.; Leung, D.Y.; Sampson, H.; Sicherer, S.; et al. Early-life gut microbiome composition and milk allergy resolution. J. Allergy Clin. Immunol. 2016, 138, 1122–1130. [Google Scholar] [CrossRef]
- Hua, X.; Goedert, J.J.; Pu, A.; Yu, G.; Shi, J. Allergy associations with the adult fecal microbiota: Analysis of the American Gut Project. eBioMedicine 2016, 3, 172–179. [Google Scholar] [CrossRef]
- Liu, Q.; Jing, W.; Wang, W. Bifidobacterium lactis Ameliorates the Risk of Food Allergy in Chinese Children by Affecting Relative Percentage of Treg and Th17 Cells. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 4561038. [Google Scholar] [CrossRef]
- Savage, J.H.; Lee-Sarwar, K.A.; Sordillo, J.; Bunyavanich, S.; Zhou, Y.; O’Connor, G.; Sandel, M.; Bacharier, L.B.; Zeiger, R.; Sodergren, E.; et al. A prospective microbiome-wide association study of food sensitization and food allergy in early childhood. Allergy 2018, 73, 145–152. [Google Scholar] [CrossRef]
- Wu, W.; Sun, M.; Chen, F.; Cao, A.T.; Liu, H.; Zhao, Y.; Huang, X.; Xiao, Y.; Yao, S.; Zhao, Q.; et al. Microbiota metabolite short-chain fatty acid acetate promotes intestinal IgA response to microbiota which is mediated by GPR43. Mucosal Immunol. 2017, 10, 946–956. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, Receptor for Niacin and the Commensal Metabolite Butyrate, Suppresses Colonic Inflammation and Carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR–S6K pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef]
- Michaudel, C.; Sokol, H. The Gut Microbiota at the Service of Immunometabolism. Cell Metab. 2020, 32, 514–523. [Google Scholar] [CrossRef]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.Y.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef] [PubMed]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; McKenzie, C.l.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wang, Y.; Wang, P.; Huang, Y.; Wang, F. Short-Chain Fatty Acids Manifest Stimulative and Protective Effects on Intestinal Barrier Function Through the Inhibition of NLRP3 Inflammasome and Autophagy. Cell. Physiol. Biochem. 2018, 49, 190–205. [Google Scholar] [CrossRef] [PubMed]
- Kepert, I.; Fonseca, J.; Müller, C.; Milger, K.; Hochwind, K.; Kostric, M.; Fedoseeva, M.; Ohnmacht, C.; Dehmel, S.; Nathan, P.; et al. D-tryptophan from probiotic bacteria influences the gut microbiome and allergic airway disease. J. Allergy Clin. Immunol. 2017, 139, 1525–1535. [Google Scholar] [CrossRef]
- Crestani, E.; Harb, H.; Charbonnier, L.-M.; Leirer, J.; Motsinger-Reif, A.; Rachid, R.; Phipatanakul, W.; Kaddurah-Daouk, R.; Chatila, T.A. Untargeted metabolomic profiling identifies disease-specific signatures in food allergy and asthma. J. Allergy Clin. Immunol. 2020, 145, 897–906. [Google Scholar] [CrossRef]
- Venkatesh, M.; Mukherjee, S.; Wang, H.; Li, H.; Sun, K.; Benechet, A.P.; Qiu, Z.; Maher, L.; Redinbo, M.R.; Phillips, R.S.; et al. Symbiotic Bacterial Metabolites Regulate Gastrointestinal Barrier Function via the Xenobiotic Sensor PXR and Toll-like Receptor 4. Immunity 2014, 41, 296–310. [Google Scholar] [CrossRef]
- Song, X.; Sun, X.; Oh, S.F.; Wu, M.; Zhang, Y.; Zheng, W.; Geva-Zatorsky, N.; Jupp, R.; Mathis, D.; Benoist, C.; et al. Microbial bile acid metabolites modulate gut RORγ+ regulatory T cell homeostasis. Nature 2020, 577, 410–415. [Google Scholar] [CrossRef]
- Quraishi, M.N.; Shaheen, W.; Oo, Y.H.; Iqbal, T.H. Immunological mechanisms underpinning faecal microbiota transplantation for the treatment of inflammatory bowel disease. Clin. Exp. Immunol. 2020, 199, 24–38. [Google Scholar] [CrossRef]
- Feehley, T.; Nagler, C.R. Cellular and molecular pathways through which commensal bacteria modulate sensitization to dietary antigens. Curr. Opin. Immunol. 2014, 31, 79–86. [Google Scholar] [CrossRef]
- Kulkarni, D.H.; Gustafsson, J.K.; Knoop, K.A.; McDonald, K.G.; Bidani, S.S.; Davis, J.E.; Floyd, A.N.; Hogan, S.P.; Hsieh, C.-S.; Newberry, R.D. Goblet cell associated antigen passages support the induction and maintenance of oral tolerance. Mucosal Immunol. 2020, 13, 271–282. [Google Scholar] [CrossRef]
- Stefka, A.T.; Feehley, T.; Tripathi, P.; Qiu, J.; McCoy, K.; Mazmanian, S.K.; Tjota, M.Y.; Seo, G.-Y.; Cao, S.; Theriault, B.R.; et al. Commensal bacteria protect against food allergen sensitization. Proc. Natl. Acad. Sci. USA 2014, 111, 13145–13150. [Google Scholar] [CrossRef] [PubMed]
- Sardecka-Milewska, I.; Łoś-Rycharska, E.; Gawryjołek, J.; Kowalska, E.T.; Krogulska, A. Role of FOXP3 Expression and Serum Vitamin D and C Concentrations When Predicting Acquisition of Tolerance in Infants With Cow’s Milk Allergy. J. Investig. Allergy Clin. Immunol. 2019, 30, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Vuillermin, P.J.; Goverse, G.; Vinuesa, C.G.; Mebius, R.E.; Macia, L.; Mackay, C.R. Dietary Fiber and Bacterial SCFA Enhance Oral Tolerance and Protect against Food Allergy through Diverse Cellular Pathways. Cell Rep. 2016, 15, 2809–2824. [Google Scholar] [CrossRef] [PubMed]
- Michels, K.R.; Lukacs, N.W.; Fonseca, W. TLR Activation and Allergic Disease: Early Life Microbiome and Treatment. Curr. Allergy Asthma Rep. 2018, 18, 61. [Google Scholar] [CrossRef]
- Pernomian, L.; Duarte-Silva, M.; Cardoso, C.R.D.B. The Aryl Hydrocarbon Receptor (AHR) as a Potential Target for the Control of Intestinal Inflammation: Insights from an Immune and Bacteria Sensor Receptor. Clin. Rev. Allergy Immunol. 2020, 59, 382–390. [Google Scholar] [CrossRef]
- Caminero, A.; Meisel, M.; Jabri, B.; Verdu, E.F. Mechanisms by which gut microorganisms influence food sensitivities. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 7–18. [Google Scholar] [CrossRef]
- Al-Sahlany, S.T.; Niamah, A.K. Bacterial viability, antioxidant stability, antimutagenicity and sensory properties of onion types fermentation by using probiotic starter during storage. Nutr. Food Sci. 2022, 52, 901–916. [Google Scholar] [CrossRef]
- Berni Canani, R.; Di Costanzo, M.; Bedogni, G.; Amoroso, A.; Cosenza, L.; Di Scala, C.; Granata, V.; Nocerino, R. Extensively hydrolyzed casein formula containing Lactobacillus rhamnosus GG reduces the occurrence of other allergic manifestations in children with cow’s milk allergy: 3-year randomized controlled trial. J. Allergy Clin. Immunol. 2017, 139, 1906–1913.e4. [Google Scholar] [CrossRef]
- Berni Canani, R.; Sangwan, N.; Stefka, A.; Nocerino, R.; Paparo, L.; Aitoro, R.; Calignano, A.; Khan, A.A.; Gilbert, J.A.; Nagler, C.R. Lactobacillus rhamnosus GG-supplemented formula expands butyrate-producing bacterial strains in food allergic infants. ISME J. 2016, 10, 742–750. [Google Scholar] [CrossRef]
- Bu, G.; Luo, Y.; Zhang, Y.; Chen, F. Effects of fermentation by lactic acid bacteria on the antigenicity of bovine whey proteins. J. Sci. Food Agric. 2010, 90, 2015–2020. [Google Scholar] [CrossRef]
- Bertrand-Harb, C.; Ivanova, I.; Dalgalarrondo, M.; Haertllé, T. Evolution of β-lactoglobulin and α-lactalbumin content during yoghurt fermentation. Int. Dairy J. 2003, 13, 39–45. [Google Scholar] [CrossRef]
- Rizzello, C.G.; De Angelis, M.; Coda, R.; Gobbetti, M. Use of selected sourdough lactic acid bacteria to hydrolyze wheat and rye proteins responsible for cereal allergy. Eur. Food Res. Technol. 2006, 223, 405–411. [Google Scholar] [CrossRef]
- De Angelis, M.; Rizzello, C.G.; Scala, E.; De Simone, C.; Farris, G.A.; Turrini, F.; Gobbetti, M. Probiotic Preparation Has the Capacity To Hydrolyze Proteins Responsible for Wheat Allergy. J. Food Prot. 2007, 70, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Xiao, L.; Liu, S.; Liu, X.; Luo, Y.; Ji, Q.; Yang, P.; Liu, Z. Exploration of the effect of probiotics supplementation on intestinal microbiota of food allergic mice. Am. J. Transl. Res. 2017, 9, 376–385. [Google Scholar] [PubMed]
- Fu, L.; Song, J.; Wang, C.; Fu, S.; Wang, Y. Bifidobacterium infantis Potentially Alleviates Shrimp Tropomyosin-Induced Allergy by Tolerogenic Dendritic Cell-Dependent Induction of Regulatory T Cells and Alterations in Gut Microbiota. Front. Immunol. 2017, 8, 1536. [Google Scholar] [CrossRef] [PubMed]
- Neau, E.; Delannoy, J.; Marion, C.; Cottart, C.-H.; Labellie, C.; Holowacz, S.; Butel, M.-J.; Kapel, N.; Waligora-Dupriet, A.-J. Three Novel Candidate Probiotic Strains with Prophylactic Properties in a Murine Model of Cow’s Milk Allergy. Appl. Environ. Microbiol. 2016, 82, 1722–1733. [Google Scholar] [CrossRef] [PubMed]
- Kozakova, H.; Schwarzer, M.; Srutkova, D.; Hudcovic, T.; Cukrowska, B. Colonisation of germ-free mice with probiotic lactobacilli mitigated allergic sensitisation in murine model of birch pollen allergy. Clin. Transl. Allergy 2014, 4, P26. [Google Scholar] [CrossRef]
- Fang, S.; Zhang, Y.; Zhang, Y.; Zhu, X.; Yie, B. Lactobacillus rhamnosus GG improves symptoms and its mechnism in mice with ovalbumin-induced food allergy. Chin. J. Cell. Mol. Immunol. 2017, 33, 597–600. [Google Scholar]
- Shandilya, U.K.; Sharma, A.; Kapila, R.; Kansal, V.K. Probiotic Dahi containing Lactobacillus acidophilus and Bifidobacterium bifidum modulates immunoglobulin levels and cytokines expression in whey proteins sensitised mice. J. Sci. Food Agric. 2016, 96, 3180–3187. [Google Scholar] [CrossRef]
- Li, N.; Yu, Y.; Chen, X.; Gao, S.; Zhang, Q.; Xu, C. Bifidobacterium breve M-16V alters the gut microbiota to alleviate OVA-induced food allergy through IL-33/ST2 signal pathway. J. Cell. Physiol. 2020, 235, 9464–9473. [Google Scholar] [CrossRef]
- Cortes-Perez, N.; Lozano-Ojalvo, D.; Maiga, M.; Hazebrouck, S.; Adel-Patient, K. Intragastric administration of Lactobacillus casei BL23 induces regulatory FoxP3+RORγt+ T cells subset in mice. Benef. Microbes 2017, 8, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Suarez, P.; Gueimonde, M.; Margolles, A.; Suarez-Diaz, A.M. Distinct Bifidobacterium strains drive different immune responses in vitro. Int. J. Food Microbiol. 2010, 138, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Price, D.B.; Ackland, M.L.; Burks, W.; Knight, M.I.; Suphioglu, C. Peanut Allergens Alter Intestinal Barrier Permeability and Tight Junction Localisation in Caco-2 Cell Cultures. Cell. Physiol. Biochem. 2014, 33, 1758–1777. [Google Scholar] [CrossRef] [PubMed]
- Chichlowski, M.; De Lartigue, G.; German, J.B.; Raybould, H.E.; Mills, D.A. Bifidobacteria Isolated From Infants and Cultured on Human Milk Oligosaccharides Affect Intestinal Epithelial Function. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 321–327. [Google Scholar] [CrossRef] [PubMed]
- de Kivit, S.; Tobin, M.C.; DeMeo, M.T.; Fox, S.; Garssen, J.; Forsyth, C.B.; Keshavarzian, A.; Landay, A.L. In vitro evaluation of intestinal epithelial TLR activation in preventing food allergic responses. Clin. Immunol. 2014, 154, 91–99. [Google Scholar] [CrossRef]
- Wu, Z.; Nasab, E.M.; Arora, P.; Athari, S.S. Study effect of probiotics and prebiotics on treatment of OVA-LPS-induced of allergic asthma inflammation and pneumonia by regulating the TLR4/NF-kB signaling pathway. J. Transl. Med. 2022, 20, 130. [Google Scholar] [CrossRef]
- Capurso, L. Thirty Years of Lactobacillus rhamnosus GG. J. Clin. Gastroenterol. 2019, 53, S1–S41. [Google Scholar] [CrossRef]
- Jerzynska, J.; Stelmach, W.; Balcerak, J.; Woicka-Kolejwa, K.; Rychlik, B.; Blauz, A.; Wachulec, M.; Stelmach, P.; Majak, P.; Stelmach, I. Effect of Lactobacillus rhamnosus GG and vitamin D supplementation on the immunologic effectiveness of grass-specific sublingual immunotherapy in children with allergy. Allergy Asthma Proc. 2016, 37, 324–334. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Nermes, M.; Collado, M.C.; Rautonen, N.; Salminen, S.; Isolauri, E. Specific probiotics alleviate allergic rhinitis during the birch pollen season. World J. Gastroenterol. 2009, 15, 3261–3268. [Google Scholar] [CrossRef]
- Plummer, E.L.; Lozinsky, A.C.; Tobin, J.M.; Uebergang, J.B.; Axelrad, C.; Garland, S.M.; Jacobs, S.E.; Tang, M.L.K.; ProPrems Study Group. Postnatal probiotics and allergic disease in very preterm infants: Sub-study to the ProPrems randomized trial. Allergy 2020, 75, 127–136. [Google Scholar] [CrossRef]
- West, C.E.; Hammarström, M.-L.; Hernell, O. Probiotics in primary prevention of allergic disease—Follow-up at 8–9 years of age. Allergy 2013, 68, 1015–1020. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).