
Lactic Acid Bacteria (LAB): Autochthonous and Probiotic Microbes for Meat Preservation and Fortification

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Bacteriocins and Their Classification
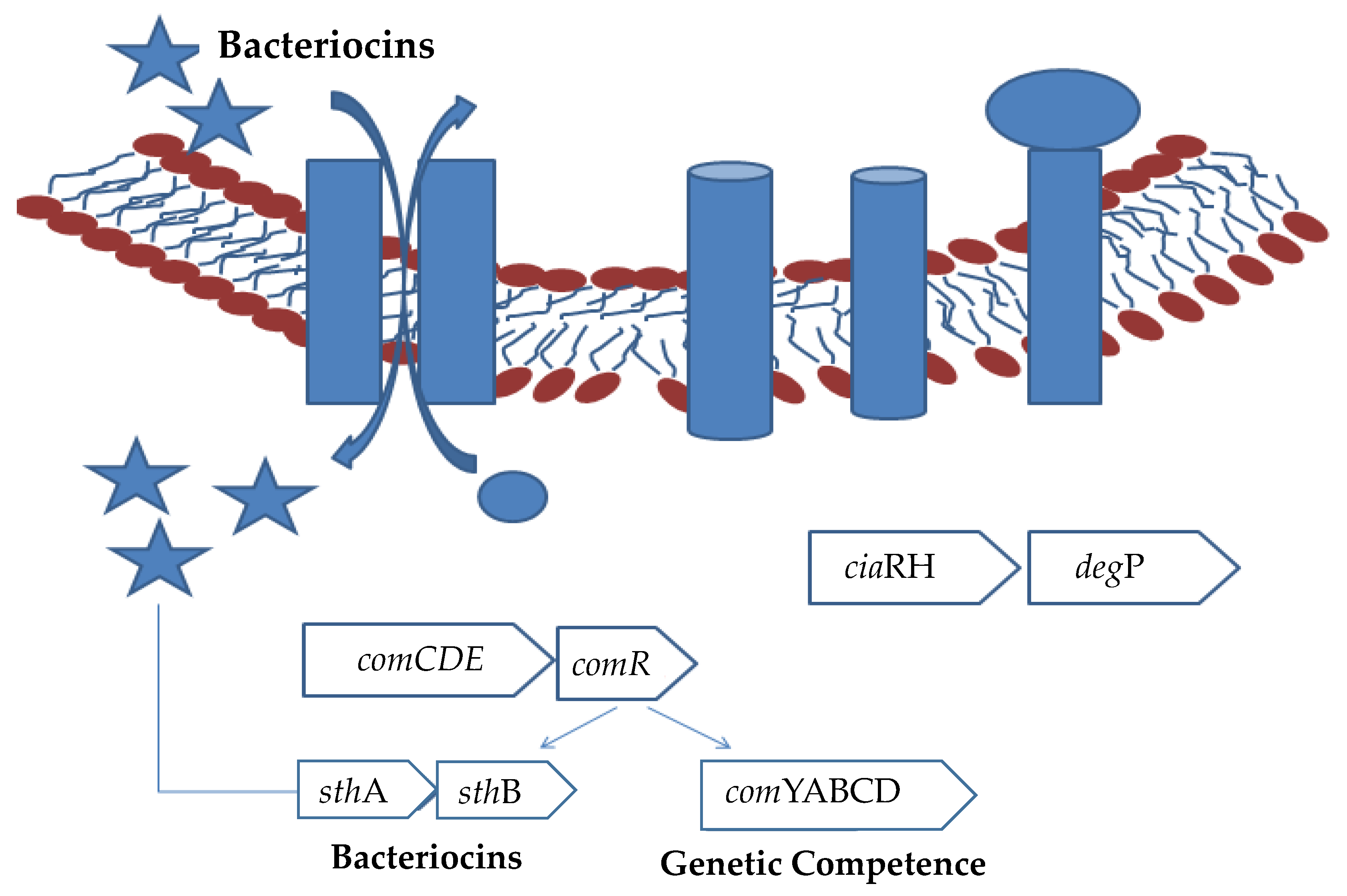
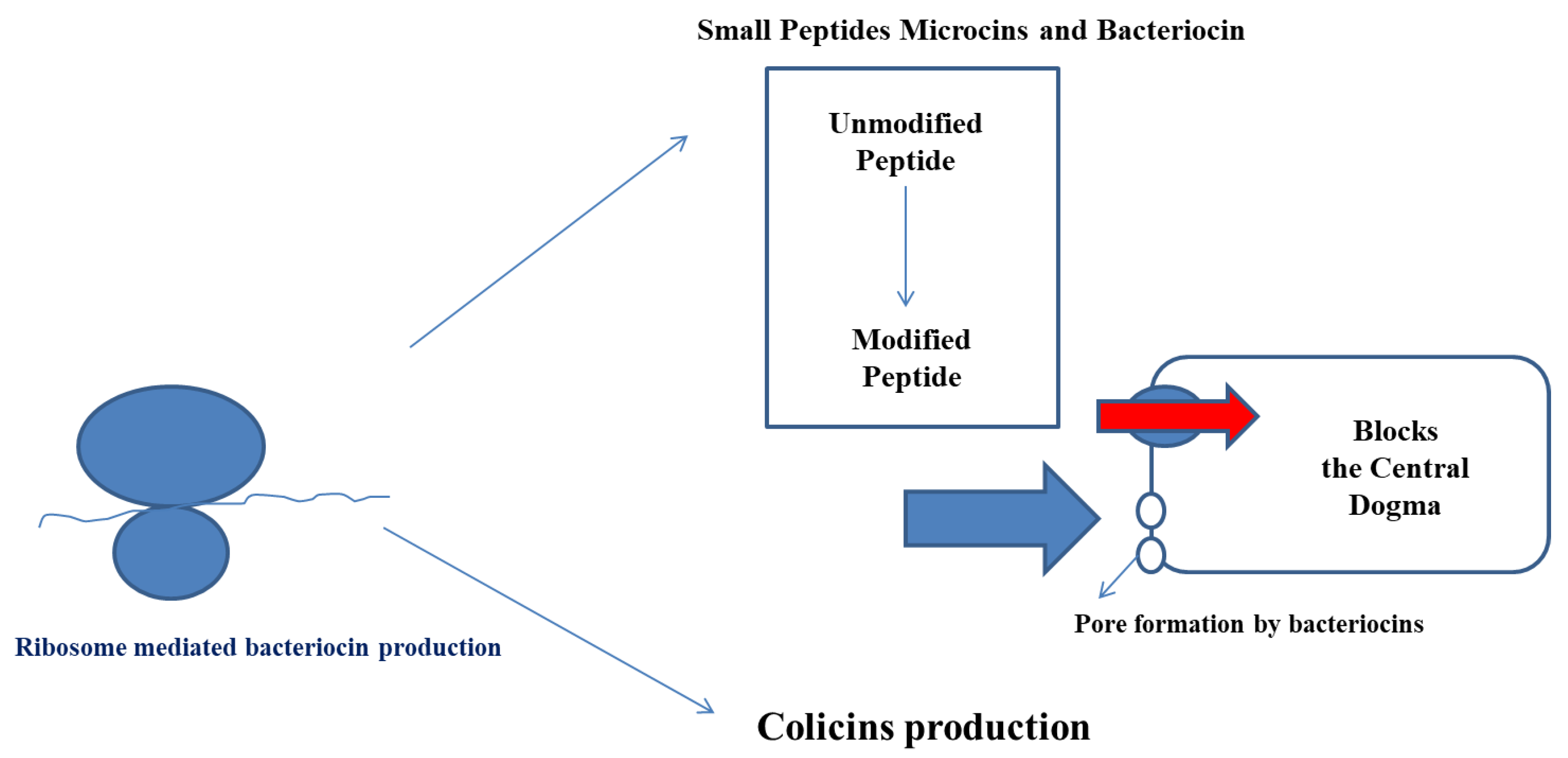
3. Genes Responsible for Bacteriocin Production
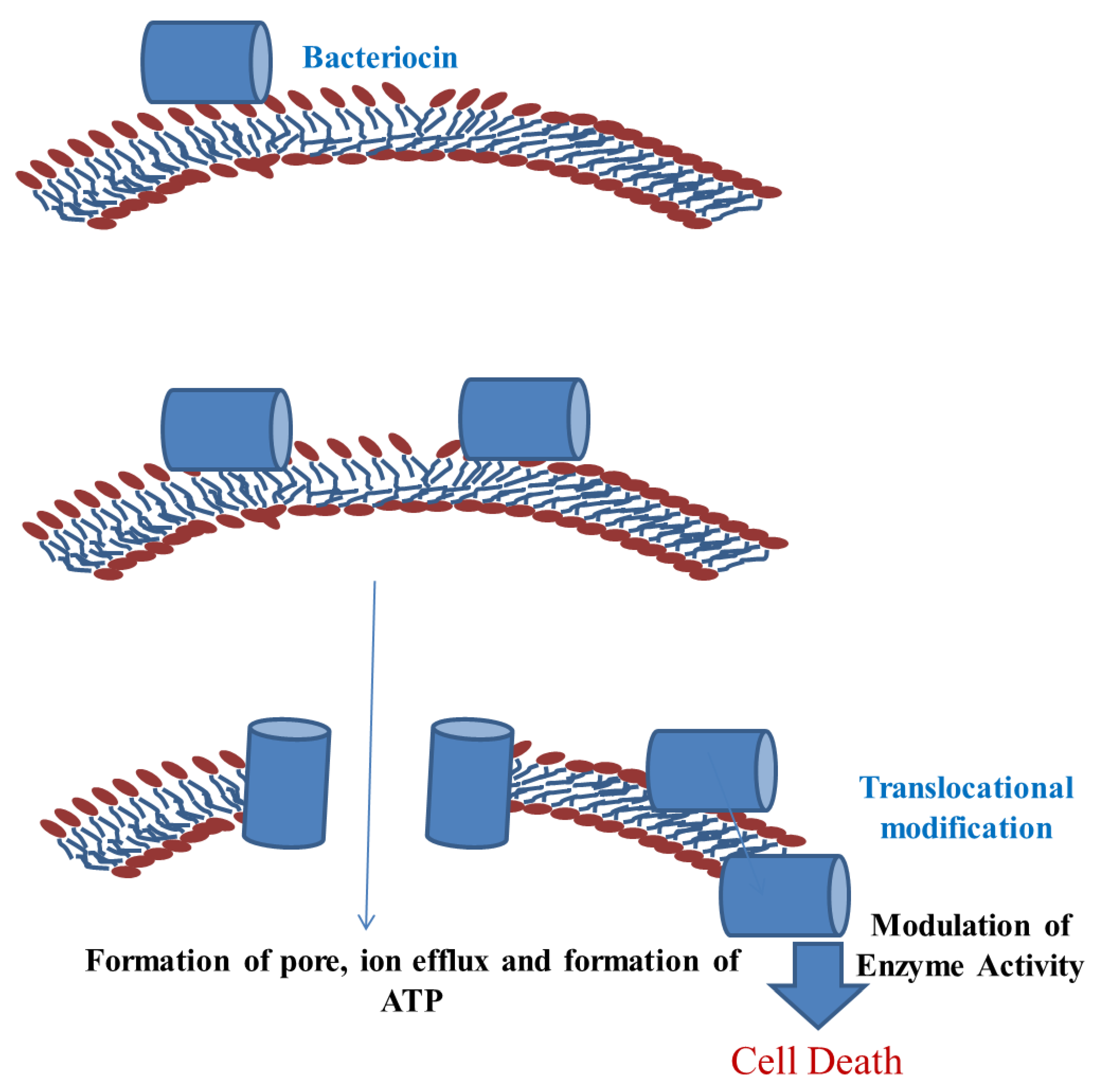
4. Mechanisms of Action of Bacteriocin
4.1. PMF (Proton Motive Force) Depletion: Microorganism Bears an ATPase in Their Plasma Membrane
4.2. Membrane Insertion and Pore Formation
- Membrane Interaction—Gram-positive bacteria are known for their abundance of anionic lipid present in their membrane. Because of the cationic nature of the bacteriocin, the anionic lipid binding is enhanced. Nisin interacts to the anionic liposome of the Gram-positive bacteria. This interaction is strong because nisin binds slowly to other liposomes and the fragments of the nisin helps in identification of the region suitable for binding. Similarly, the class II bacteriocins initially follow the anionic phospholipid membrane interaction [12].
- Membrane Insertion—The insertion with the lipid monolayer has been established. As per the research study, the capability of the nisin to interact with the lipid layer present in the membrane is enhanced by the presence of strong antimicrobial activity. It has been stated that a low anti-microbial activity results in a decreased lipid interaction of the bacteriocin nisin and vice versa [12].
- Cell Wall Passage and Receptors—The class I bacteriocin Lantibiotics can form ion-conducting pores in the black lipid membranes of the Gram-positive bacteria. This results in the interaction with the peptidoglycan precursor lipid 2 in the membrane. The presence of the lipid 2 precursor enhances the ability of the bacteriocin to depolarize the electrical potential of the membrane. These data support the fact that lipid 2 acts as the docking molecule or the membrane nucleus, which controls all the activities. The Gram-positive cell wall allows the passage of the bacteriocins as enhanced by the Lipid 2. The polymers on the cell surface, lipoic acid and lipoteichoic acid, play a crucial role in the initial interaction of the bacterial cell wall and the bacteriocins. However, the receptor plays a role in this whole process and their implications have been explained in many research studies on specific narrow targets [13].
4.3. Bacteriocin Affecting DNA Synthesis
5. Spoilage of Meat and Meat Products
6. Development of Biofilm by LAB
7. Role of LAB in the Preservation of Meat and Meat Products
Mechanism of Protection of Meat and Meat Products by LAB
8. Fortification of Meat Products by LAB
9. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barcenilla, C.; Ducic, M.; López, M.; Prieto, M.; Álvarez-Ordóñez, A. Application of lactic acid bacteria for the biopreservation of meat products: A systematic review. Meat Sci. 2022, 183, 108661. [Google Scholar] [CrossRef] [PubMed]
- Woraprayote, W.; Malila, Y.; Sorapukdee, S.; Swetwiwathana, A.; Benjakul, S.; Visessanguan, W. Bacteriocins from lactic acid bacteria and their applications in meat and meat products. Meat Sci. 2016, 120, 118–132. [Google Scholar] [CrossRef] [PubMed]
- da Costa, R.J.; Voloski, F.L.S.; Mondadori, R.G.; Duval, E.H.; Fiorentini, Â.M. Preservation of Meat Products with Bacteriocins Produced by Lactic Acid Bacteria Isolated from Meat. J. Food Qual. 2019, 2019, 4726510. [Google Scholar] [CrossRef]
- Koort, J.; Murros, A.; Coenye, T.; Eerola, S.; Vandamme, P.; Sukura, A.; Björkroth, J. Lactobacillus oligofermentans sp. nov., associated with spoilage of modified-atmosphere-packaged poultry products. Appl. Environ. Microbiol. 2005, 71, 4400–4406. [Google Scholar] [CrossRef] [PubMed]
- Björkroth, J. Microbiological ecology of marinated meat products. Meat Sci. 2005, 70, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.J.; Scott, R.W. New horizons for host defense peptides and lantibiotics. Curr. Opin. Pharmacol. 2012, 12, 545–550. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Karczewski, J.; Krasucki, S.P.; Asare-Okai, P.N.; Diehl, C.; Friedman, A.; Brown, C.M.; Maezato, Y.; Streatfield, S.J. Isolation, Characterization and Structure Elucidation of a Novel Lantibiotic From Paenibacillus sp. Front. Microbiol. 2020, 11, 598789. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; Abriouel, H.; López, R.L.; Omar, N. Ben Bacteriocin-based strategies for food biopreservation. Int. J. Food Microbiol. 2007, 120, 51–70. [Google Scholar] [CrossRef]
- Condon, S. Responses of lactic acid bacteria to oxygen. FEMS Microbiol. Rev. 1987, 3, 269–280. [Google Scholar] [CrossRef]
- Erkmen, O.; Bozoglu, T. Spoilage of Meat and Meat Products. Food Microbiol. Princ. into Pract. 2016, 279–295. [Google Scholar]
- De Martinis, E.C.P.; Alves, V.F.; Franco, B.D.G.M. Fundamentals and perspectives for the use of bacteriocins produced by lactic acid bacteria in meat products. Food Rev. Int. 2002, 18, 191–208. [Google Scholar] [CrossRef]
- de Azevedo, P.O.S.; Mendonça, C.M.N.; Seibert, L.; Domínguez, J.M.; Converti, A.; Gierus, M.; Oliveira, R.P.S. Bacteriocin-like inhibitory substance of Pediococcus pentosaceus as a biopreservative for Listeria sp. control in ready-to-eat pork ham. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2020, 51, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Smaoui, S.; Ben Hsouna, A.; Lahmar, A.; Ennouri, K.; Mtibaa-Chakchouk, A.; Sellem, I.; Najah, S.; Bouaziz, M.; Mellouli, L. Bio-preservative effect of the essential oil of the endemic Mentha piperita used alone and in combination with BacTN635 in stored minced beef meat. Meat Sci. 2016, 117, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Cho, A.R.; Han, J. Antioxidant and antimicrobial activities of leafy green vegetable extracts and their applications to meat product preservation. Food Control 2013, 29, 112–120. [Google Scholar] [CrossRef]
- EFSA The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6462. [CrossRef]
- Cosansu, S.; Geornaras, I.; Ayhan, K.; Sofos, J.N. Control of Listeria monocytogenes by bacteriocin-producing Pediococcus acidilactici 13 and its antimicrobial substance in a dry fermented sausage sucuk and in turkey breast. J. Food Nutr. Res. 2010, 49, 206–214. [Google Scholar]
- Pisoschi, A.M.; Pop, A.; Georgescu, C.; Turcuş, V.; Olah, N.K.; Mathe, E. An overview of natural antimicrobials role in food. Eur. J. Med. Chem. 2018, 143, 922–935. [Google Scholar] [CrossRef]
- Borch, E.; Kant-Muermans, M.-L.; Blixt, Y. Bacterial spoilage of meat and cured meat products. Int. J. Food Microbiol. 1996, 33, 103–120. [Google Scholar] [CrossRef]
- Ajaykumar, V.J.; Mandal, P.K. Chapter 18—Modern concept and detection of spoilage in meat and meat products. In Meat Quality Analysis; Biswas, A.K., Mandal, P.K., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 335–349. ISBN 978-0-12-819233-7. [Google Scholar]
- Lahiri, D.; Dash, S.; Dutta, R.; Nag, M. Elucidating the effect of anti-biofilm activity of bioactive compounds extracted from plants. J. Biosci. 2019, 44, 52. [Google Scholar] [CrossRef]
- Stiles, M.E.; Holzapfel, W.H. Lactic acid bacteria of foods and their current taxonomy. Int. J. Food Microbiol. 1997, 36, 1–29. [Google Scholar] [CrossRef]
- Terraf, M.C.L.; Juárez Tomás, M.S.; Nader-Macías, M.E.F.; Silva, C. Screening of biofilm formation by beneficial vaginal lactobacilli and influence of culture media components. J. Appl. Microbiol. 2012, 113, 1517–1529. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Verhoeven, T.L.A.; Perea Vélez, M.; Vanderleyden, J.; De Keersmaecker, S.C.J. Impact of environmental and genetic factors on biofilm formation by the probiotic strain Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 2007, 73, 6768–6775. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.N.; Cabral, M.E.S.; Noseda, D.; Bosch, A.; Yantorno, O.M.; Valdez, J.C. Antipathogenic properties of Lactobacillus plantarum on Pseudomonas aeruginosa: The potential use of its supernatants in the treatment of infected chronic wounds. Wound Repair Regen. 2012, 20, 552–562. [Google Scholar] [CrossRef]
- Bujňáková, D.; Kmeť, V. Functional properties of Lactobacillus strains isolated from dairy products. Folia Microbiol. 2012, 57, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Verhoeven, T.L.A.; Francius, G.; Schoofs, G.; Lambrichts, I.; Dufrêne, Y.; Vanderleyden, J.; De Keersmaecker, S.C.J. Identification of a Gene Cluster for the Biosynthesis of a Long, Galactose-Rich Exopolysaccharide in Lactobacillus rhamnosus GG and Functional Analysis of the Priming Glycosyltransferase. Appl. Environ. Microbiol. 2009, 75, 3554–3563. [Google Scholar] [CrossRef]
- Aoudia, N.; Rieu, A.; Briandet, R.; Deschamps, J.; Chluba, J.; Jego, G.; Garrido, C.; Guzzo, J. Biofilms of Lactobacillus plantarum and Lactobacillus fermentum: Effect on stress responses, antagonistic effects on pathogen growth and immunomodulatory properties. Food Microbiol. 2016, 53, 51–59. [Google Scholar] [CrossRef]
- Sakaridis, I.; Soultos, N.; Dovas, C.I.; Papavergou, E.; Ambrosiadis, I.; Koidis, P. Lactic acid bacteria from chicken carcasses with inhibitory activity against Salmonella spp. and Listeria monocytogenes. Anaerobe 2012, 18, 62–66. [Google Scholar] [CrossRef]
- Gonzalez, H.M.I.; Yien, W.; Castrillon, V.J.A.; Ortega, P.A. Addition of Carnobacterium maltaromaticum CB1 in vacuum packaged chorizo and morcilla, to inhibit the growth of Listeria monocytogenes. Vitae 2013, 20, 23–29. [Google Scholar]
- Gelinski, J.M.L.N.; Baratto, C.M.; Casagrande, M.; de Oliveira, T.P.; Megiolaro, F.; de Martini Soares, F.A.S.; de Souza, E.M.B.; Vicente, V.A.; Fonseca, G.G. Control of pathogens in fresh pork sausage by inclusion of Lactobacillus sakei BAS0117. Can. J. Microbiol. 2019, 65, 831–841. [Google Scholar] [CrossRef]
- Arief, I.I.; Jenie, B.S.L.; Suryati, T.; Ayuningtyas, G.; Fuziawan, A. Antimicrobial activity of bacteriocin from indigenous Lactobacillus plantarum 2C12 and its application on beef meatball as biopreservative. J. Indones. Trop. Anim. Agric. 2012, 37, 90–96. [Google Scholar] [CrossRef]
- Castellano, P.; González, C.; Carduza, F.; Vignolo, G. Protective action of Lactobacillus curvatus CRL705 on vacuum-packaged raw beef. Effect on sensory and structural characteristics. Meat Sci. 2010, 85, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.Y.; Balay, D.; Hu, Y.; McMullen, L.M.; Gänzle, M.G. Effect of chitosan, and bacteriocin—Producing Carnobacterium maltaromaticum on survival of Escherichia coli and Salmonella Typhimurium on beef. Int. J. Food Microbiol. 2019, 290, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Olaoye, O.A.; Onilude, A.A. Investigation on the potential application of biological agents in the extension of shelf life of fresh beef in Nigeria. World J. Microbiol. Biotechnol. 2010, 26, 1445–1454. [Google Scholar] [CrossRef]
- Kamiloğlu, A.; Kaban, G.; Kaya, M. Effects of autochthonous Lactobacillus plantarum strains on Listeria monocytogenes in sucuk during ripening. J. Food Saf. 2019, 39, e12618. [Google Scholar] [CrossRef]
- Kumar, Y.; Kaur, K.; Shahi, A.K.; Kairam, N.; Tyagi, S.K. Antilisterial, antimicrobial and antioxidant effects of pediocin and Murraya koenigii berry extract in refrigerated goat meat emulsion. LWT—Food Sci. Technol. 2017, 79, 135–144. [Google Scholar] [CrossRef]
- Wijnker, J.J.; Weerts, E.A.W.S.; Breukink, E.J.; Houben, J.H.; Lipman, L.J.A. Reduction of Clostridium sporogenes spore outgrowth in natural sausage casings using nisin. Food Microbiol. 2011, 28, 974–979. [Google Scholar] [CrossRef]
- García-Díez, J.; Patarata, L. Influence of salt level, starter culture, fermentable carbohydrates, and temperature on the behaviour of L. monocytogenes in sliced chouriço during storage. Acta Aliment. 2017, 46, 206–213. [Google Scholar] [CrossRef]
- Parolin, C.; Croatti, V.; Laghi, L.; Giordani, B.; Tondi, M.R.; De Gregorio, P.R.; Foschi, C.; Vitali, B. Lactobacillus Biofilms Influence Anti-Candida Activity. Front. Microbiol. 2021, 12, 750368. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and Inputs From Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters during Food-Animal Production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef]
- Kröckel, L. Evaluation of a novel agar medium for the detection of hydrogen peroxide producing lactic acid bacteria. Die Fleischwirtschaft 2011, 91, 97–101. [Google Scholar]
- Santos, M.H.S. Biogenic amines: Their importance in foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Schirone, M.; Esposito, L.; D’Onofrio, F.; Visciano, P.; Martuscelli, M.; Mastrocola, D.; Paparella, A. Biogenic Amines in Meat and Meat Products: A Review of the Science and Future Perspectives. Foods 2022, 11, 788. [Google Scholar] [CrossRef] [PubMed]
- Khorshidian, N.; Khanniri, E.; Mohammadi, M.; Mortazavian, A.M.; Yousefi, M. Antibacterial Activity of Pediocin and Pediocin-Producing Bacteria Against Listeria monocytogenes in Meat Products. Front. Microbiol. 2021, 12, 709959. [Google Scholar] [CrossRef]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of bacteriocins and protective cultures in dairy food preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Rivas, F.P.; Cayré, M.E.; Campos, C.A.; Castro, M.P. Natural and artificial casings as bacteriocin carriers for the biopreservation of meats products. J. Food Saf. 2018, 38, e12419. [Google Scholar] [CrossRef]
- Vijayakumar, P.P.; Muriana, P.M. Inhibition of Listeria monocytogenes on ready-to-eat meats using bacteriocin mixtures based on mode-of-action. Foods 2017, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- de Macedo, R.E.F.; Pflanzer, S.; Gomes, C. Probiotic Meat Products. In Probiotic in Animals; Rigobelo, E., Ed.; IntechOpen: London, UK, 2012. [Google Scholar]
- Heenan, C.N.; Adams, M.C.; Hosken, R.W.; Fleet, G.H. Growth Medium for Culturing Probiotic Bacteria for Applications in Vegetarian Food Products. LWT—Food Sci. Technol. 2002, 35, 171–176. [Google Scholar] [CrossRef]
- Arihara, K.; Ota, H.; Itoh, M.; Kondo, Y.; Sameshima, T.; Yamanaka, H.; Akimoto, M.; Kanai, S.; Miki, T. Lactobacillus acidophilus Group Lactic Acid Bacteria Applied to Meat Fermentation. J. Food Sci. 1998, 63, 544–547. [Google Scholar] [CrossRef]
- Vermeiren, L.; Devlieghere, F.; Debevere, J. Evaluation of meat born lactic acid bacteria as protective cultures for the biopreservation of cooked meat products. Int. J. Food Microbiol. 2004, 96, 149–164. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name of LAB | Function of the Biofilm | Reference |
|---|---|---|
| Lactobacillus rhamnosus | The development of the biofilm helps in the inhibition of the biofilm formed by Salmonella sp. H9812 and Escherichia coli | [25] |
| welE gene helps in the production of the exoplysaccharide (EPS) there by helping in the adherence with various surfaces | [26] | |
| Lactiplantibacillus plantarum | Helps in the eradication of organisms like Salmonella enterica serovar Enteritidis, Staphyloccocus aureus, and Listeria monocytogenes | [27] |
| Lactobacillus reuteri | Inhibits the growth of Escherichia coli, | [27] |
| Lactobacillus fermentum | Helps in inhibiting the growth of S. Enteritidis and Escherichia coli | [27] |
| Preserved Meat | Type of LAB Used | Type of Targeted Organism | Observation | Reference |
|---|---|---|---|---|
| Raw beef packed in vacuum packs | L. curvatus-mediated production of lactocin | B. thermosphacta | Effective viability reduction of B. thermosphacta was observed | [32] |
| Pork-ham ready to eat packs | P. pentosaceus-mediated production of bacteriocin-like substance and nisin | L. seeligeri | It helps in bringing about log 1.7 times inhibition of L. seeligeri | [12] |
| Meat balls made up of beef | L. plantarum-mediated production of bacteriocin | E. coli and Salmonella enterica serovar Typhimurium | It helps in the significant reduction of the pathogenic organism | [31] |
| Beef slices | C. maltaromaticum-mediated production of bateriocin | S. Typhimurium and E. coli | It brings about marked reduction of the targeted organism | [33] |
| Fresh samples of beef | P. acidilactici and P. pentosaceus | S. Typhimurium and L. monocytogenes | Brings about two-fold reduction in the growth of the pathogenic organisms those are associated with the degradation of meat | [34] |
| Sausages of meat | P. acidilactici-associated production of bacteriocin | L. monocytogenes | Three-fold reduction in the targeted organism | [16] |
| Sucuk sausages | L. plantarum-mediated production of bacteriocin | L. monocytogenes | Brings about marked reduction in the growth of the microorganims | [35] |
| Emulsion of goat meat | Murraya koenigii andb P. pentosaceus-mediated pediocin production | L. innocua | Brings about 2- to 3-fold reduction in the targeted organism | [36] |
| Natural casings of sheep | Bacteriocins produced by LAB | Clostridium sporogene | Brings about marked reduction in the Clostridium sp. | [37] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lahiri, D.; Nag, M.; Sarkar, T.; Ray, R.R.; Shariati, M.A.; Rebezov, M.; Bangar, S.P.; Lorenzo, J.M.; Domínguez, R. Lactic Acid Bacteria (LAB): Autochthonous and Probiotic Microbes for Meat Preservation and Fortification. Foods 2022, 11, 2792. https://doi.org/10.3390/foods11182792
Lahiri D, Nag M, Sarkar T, Ray RR, Shariati MA, Rebezov M, Bangar SP, Lorenzo JM, Domínguez R. Lactic Acid Bacteria (LAB): Autochthonous and Probiotic Microbes for Meat Preservation and Fortification. Foods. 2022; 11(18):2792. https://doi.org/10.3390/foods11182792
Chicago/Turabian StyleLahiri, Dibyajit, Moupriya Nag, Tanmay Sarkar, Rina Rani Ray, Mohammad Ali Shariati, Maksim Rebezov, Sneh Punia Bangar, José M. Lorenzo, and Rubén Domínguez. 2022. "Lactic Acid Bacteria (LAB): Autochthonous and Probiotic Microbes for Meat Preservation and Fortification" Foods 11, no. 18: 2792. https://doi.org/10.3390/foods11182792
APA StyleLahiri, D., Nag, M., Sarkar, T., Ray, R. R., Shariati, M. A., Rebezov, M., Bangar, S. P., Lorenzo, J. M., & Domínguez, R. (2022). Lactic Acid Bacteria (LAB): Autochthonous and Probiotic Microbes for Meat Preservation and Fortification. Foods, 11(18), 2792. https://doi.org/10.3390/foods11182792