
Effects of Cinnamon Essential Oil on Oxidative Damage and Outer Membrane Protein Genes of Salmonella enteritidis Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Antioxidant Activity Assay
2.2.1. Determination of ROS
2.2.2. O2− Free Radical Determination
2.2.3. Fluorescence Microscopy to Observe Changes in ROS Content
2.3. Enzyme Activity Determination
2.3.1. SOD Activity Determination
2.3.2. CAT, POD and MDA Activity Determination
2.4. Determination of Protein Carbonylation
2.5. The Effect of CEO on Omp Gene Transcription Level
2.5.1. PCR Amplification
2.5.2. Agarose Gel Electrophoresis
2.5.3. Preparation of Salmonella enteritidis cDNA
2.5.4. q-PCR Test
2.6. Data Analysis
3. Results and Analysis
3.1. Antioxidant Activity Results
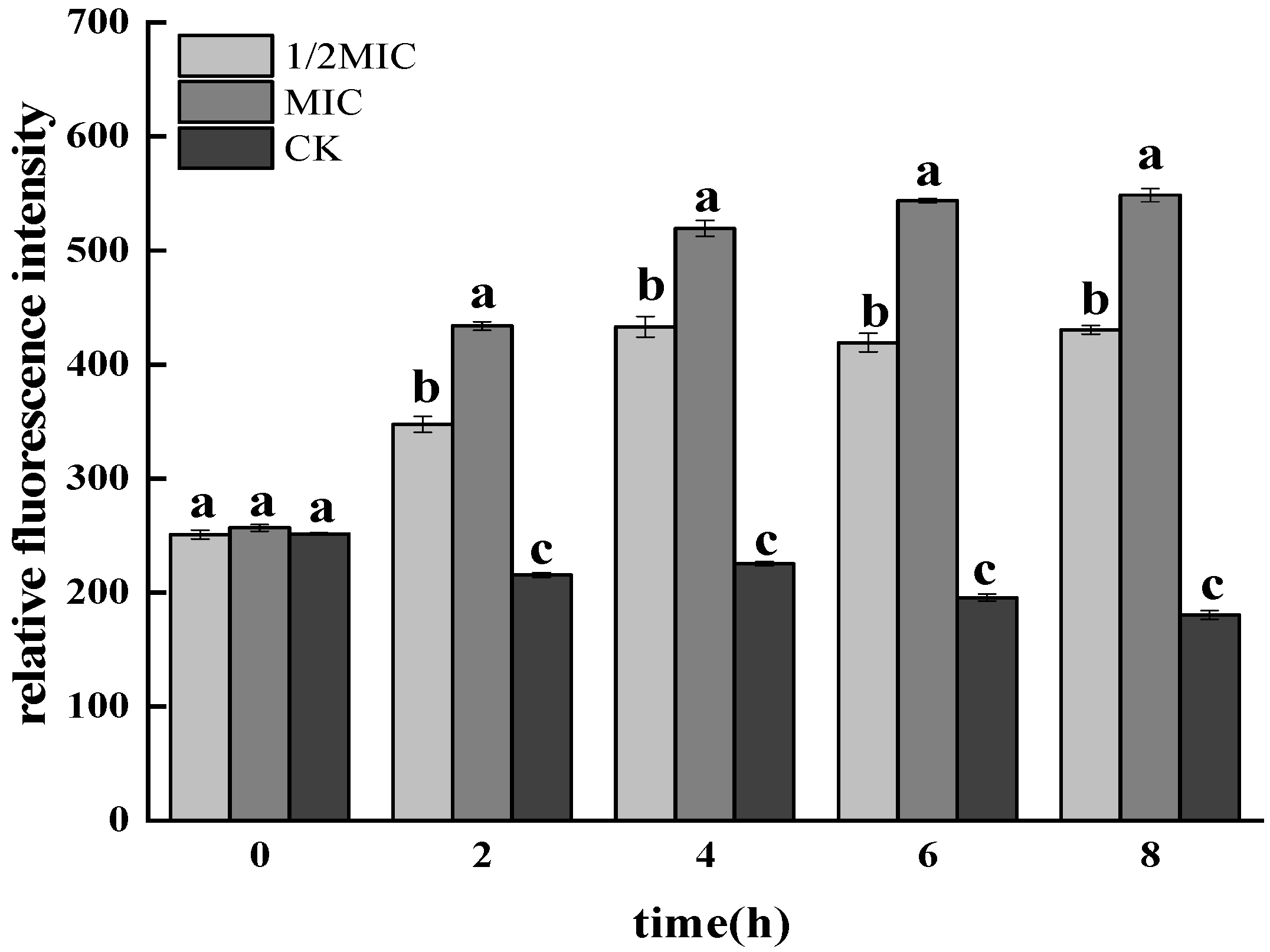
3.1.1. Determination of ROS
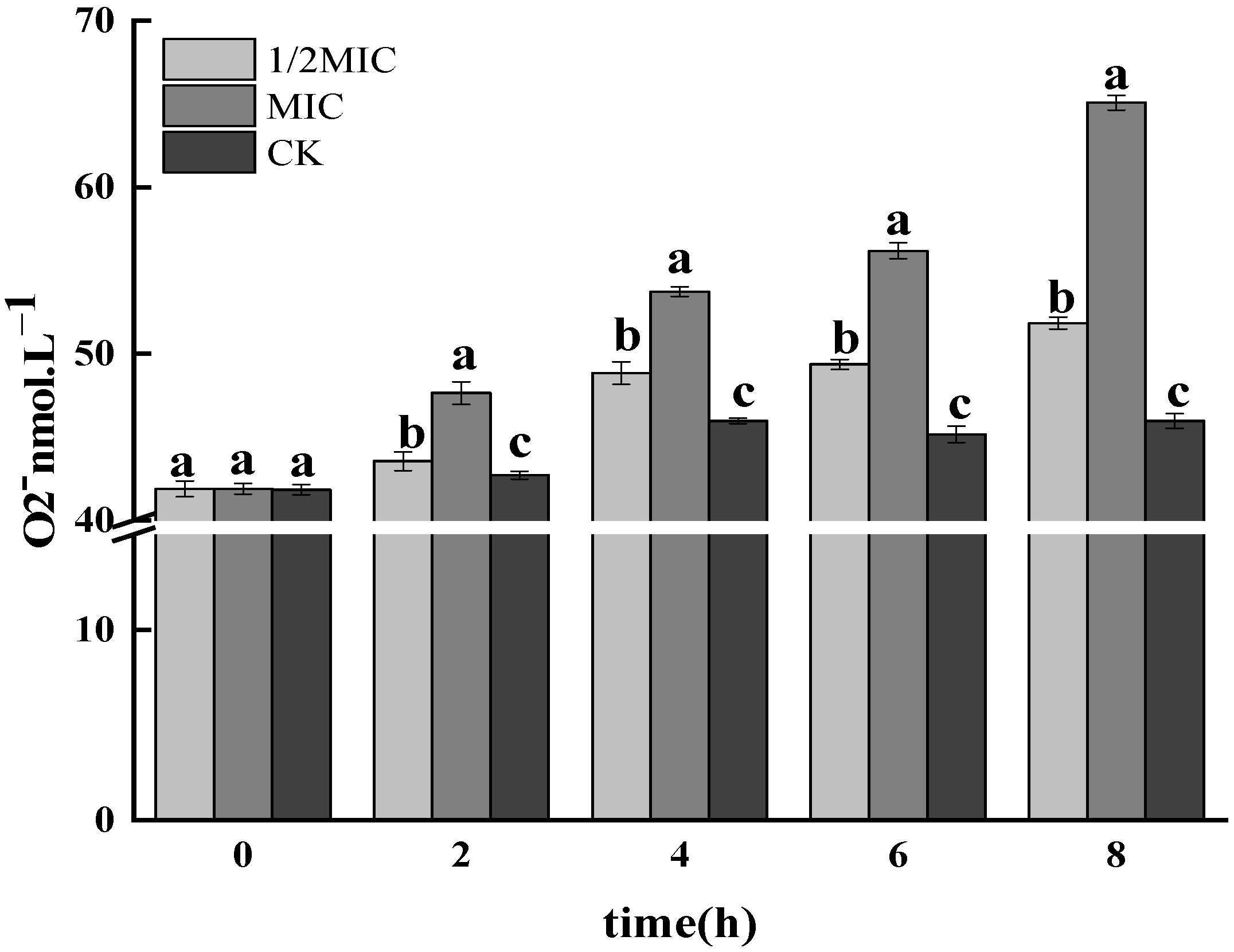
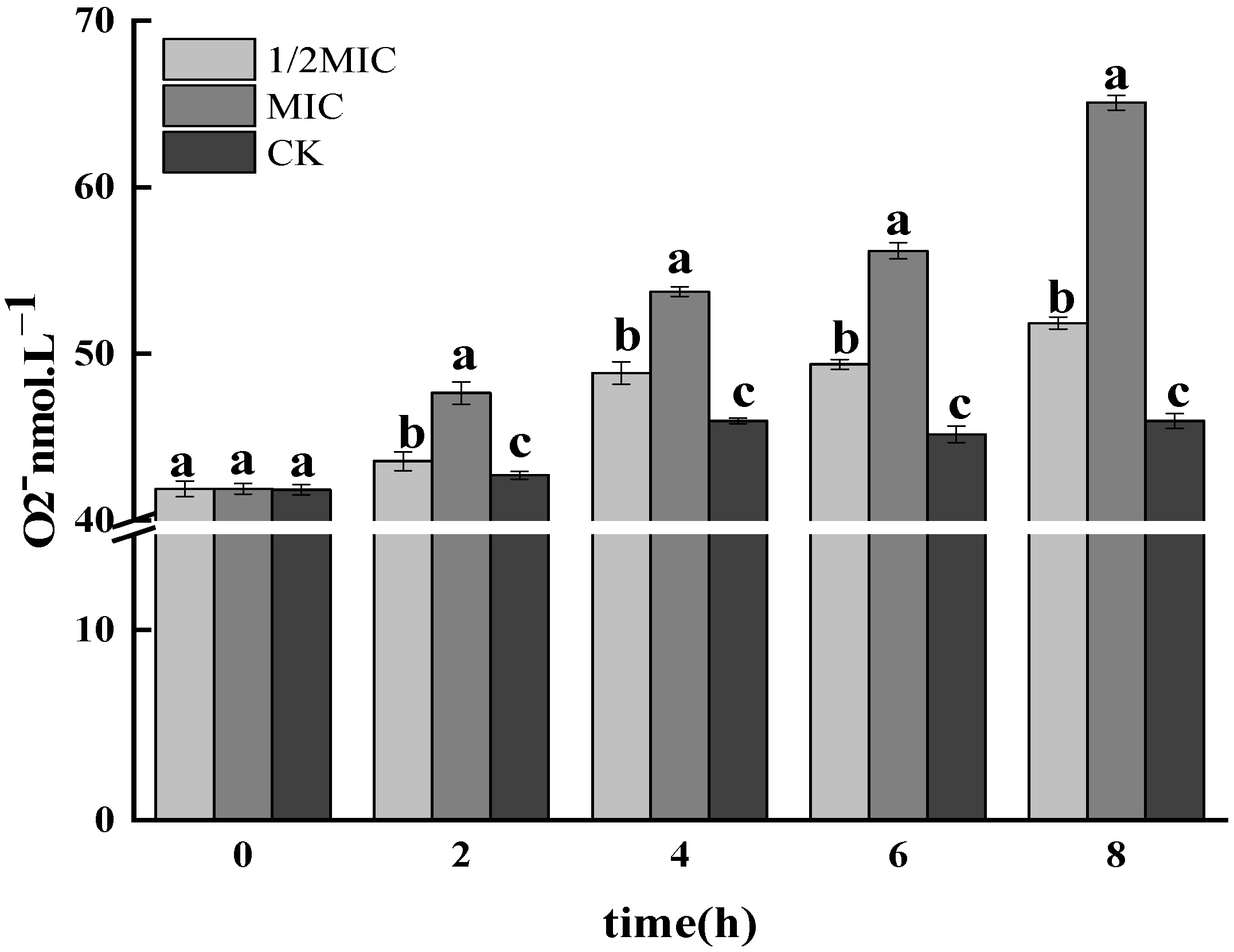
3.1.2. Determination of O2− Free Radicals
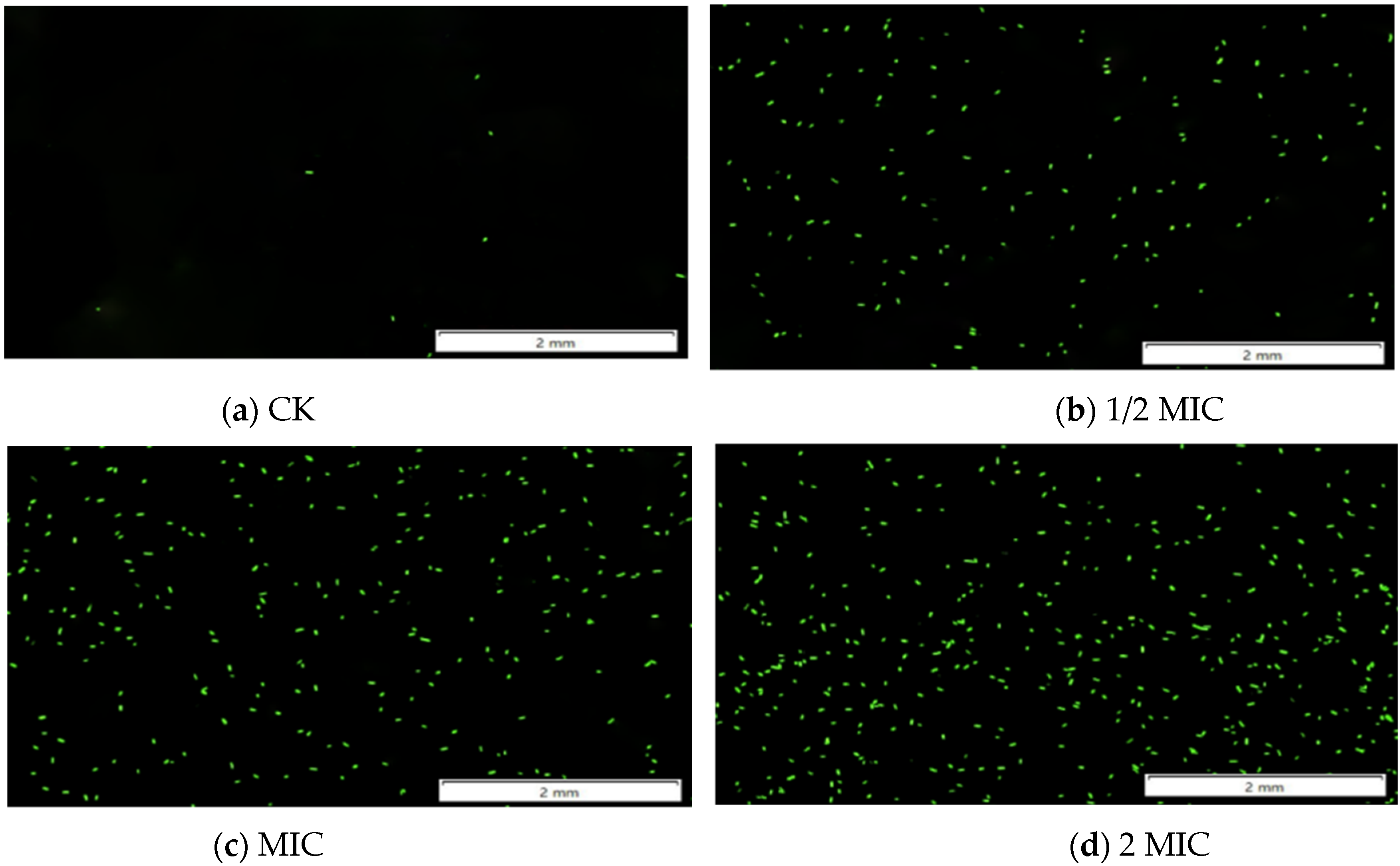
3.1.3. Fluorescence Microscopy to Observe the Changes of ROS
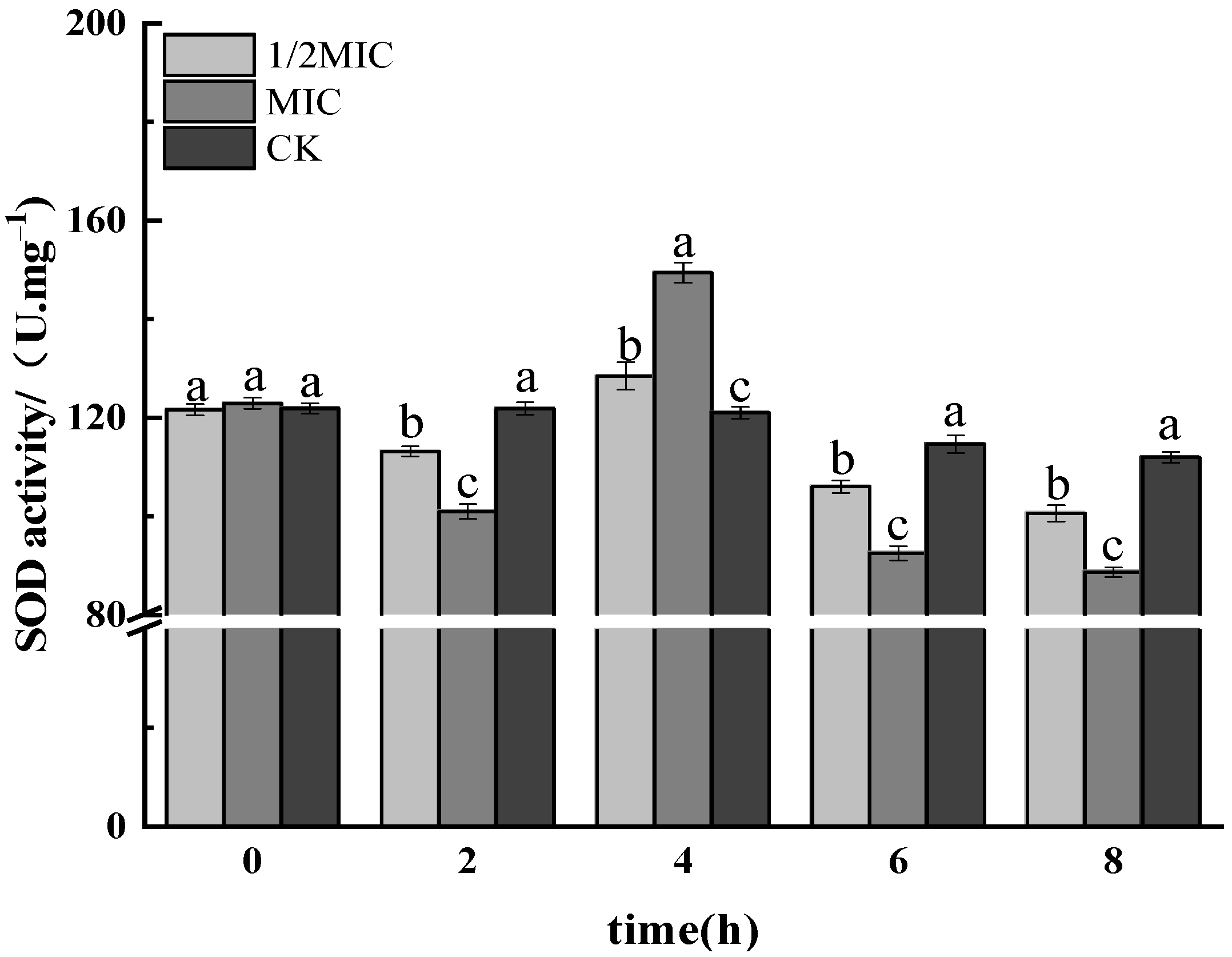
3.1.4. SOD Activity Assay
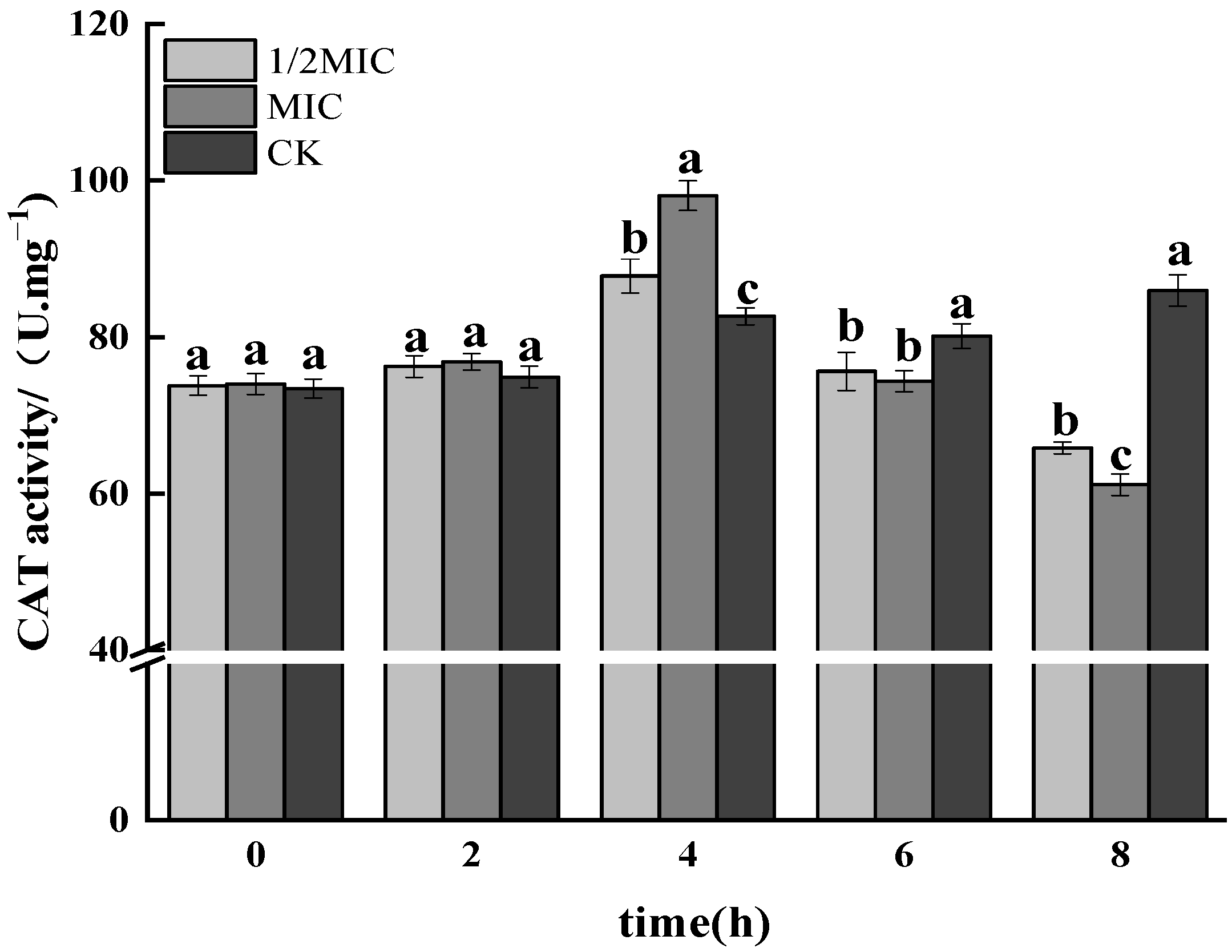
3.1.5. CAT Activity Assay
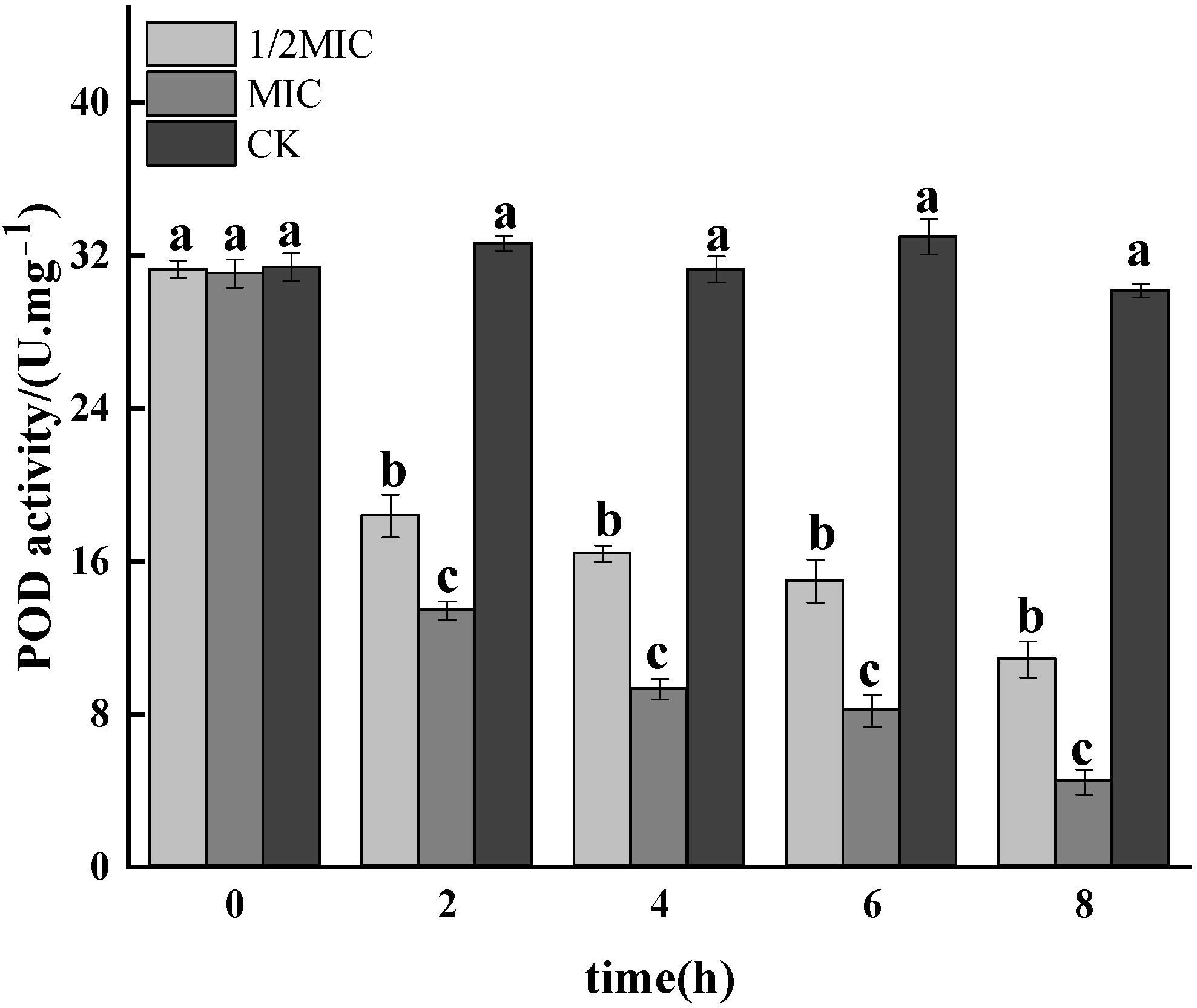
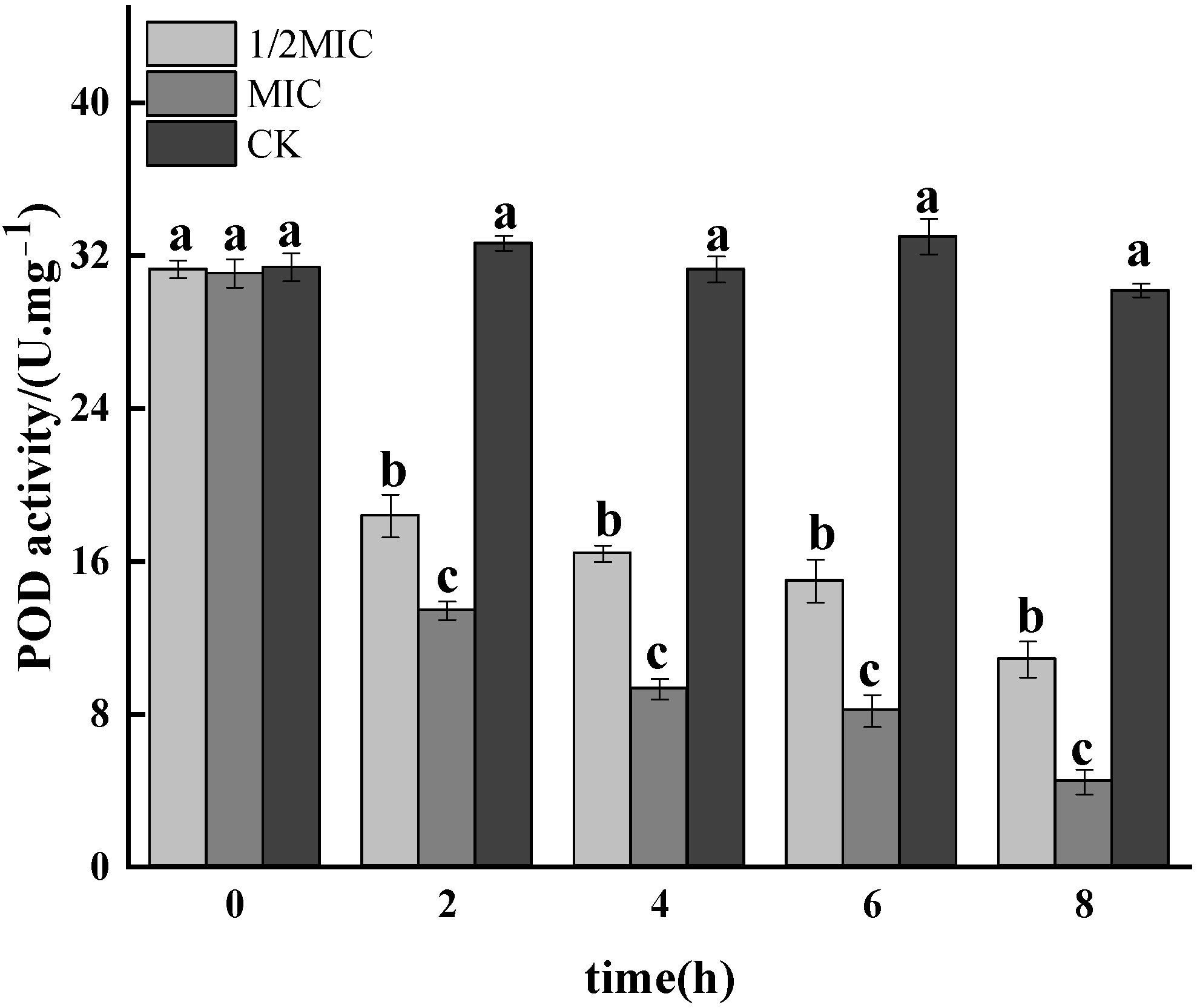
3.1.6. POD Activity Assay
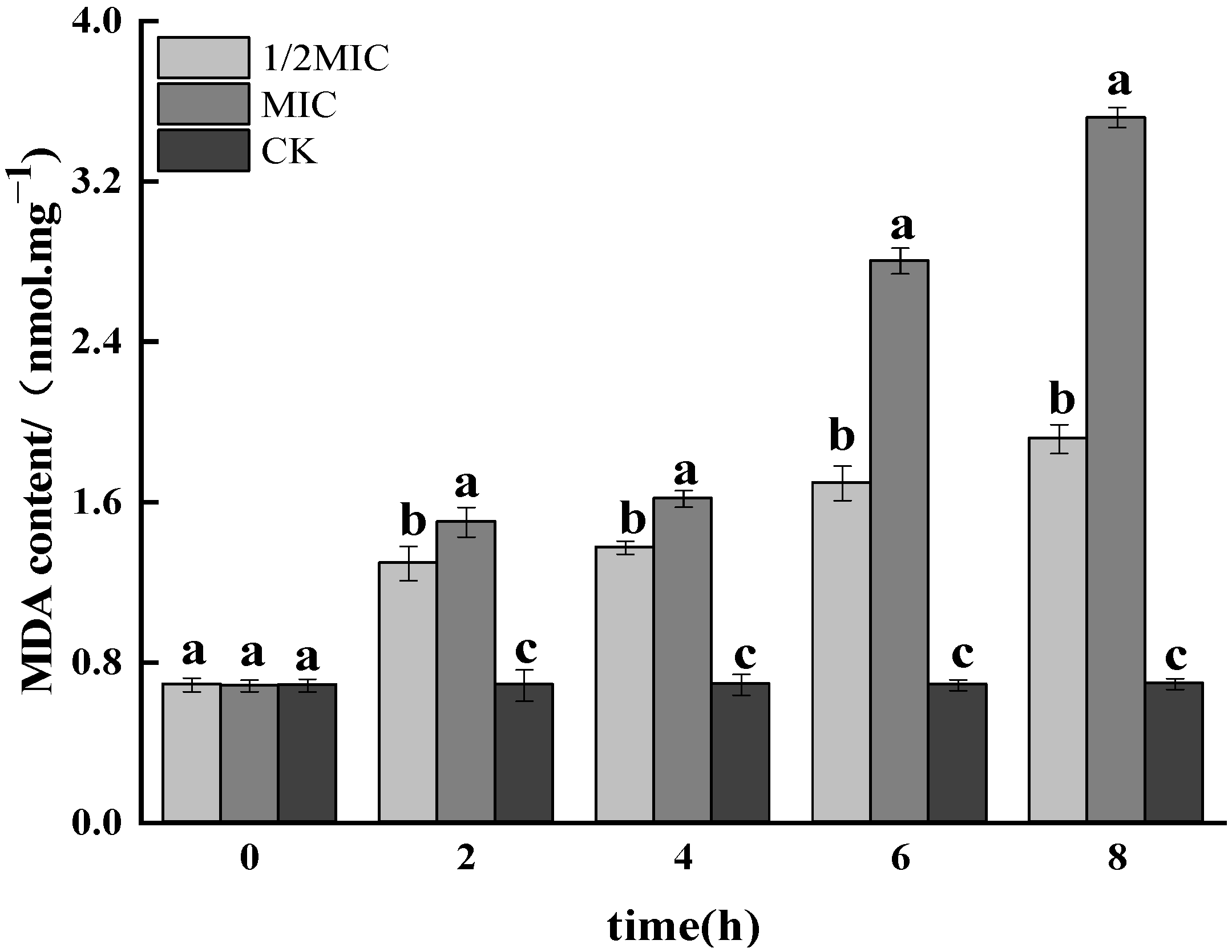
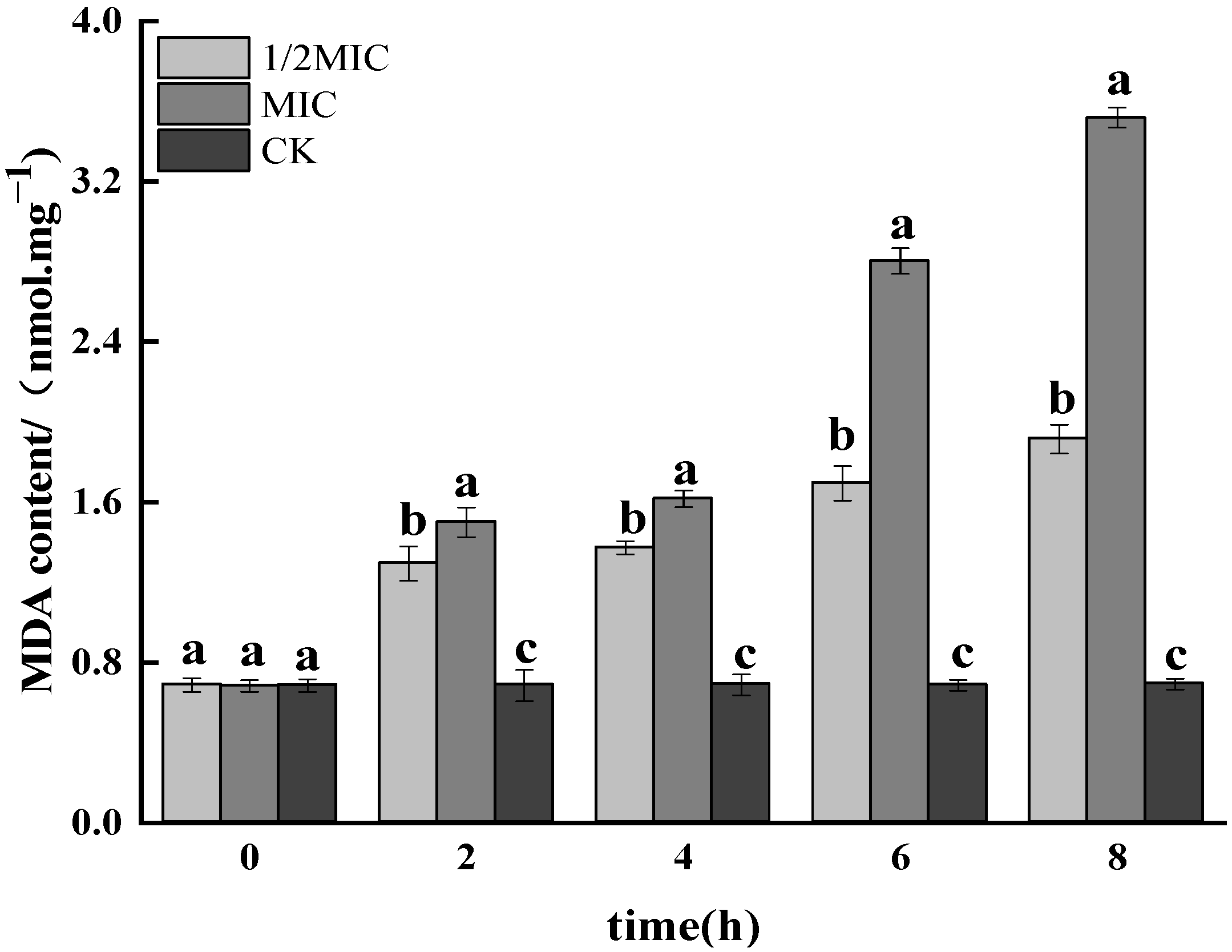
3.1.7. Determination of MDA Content
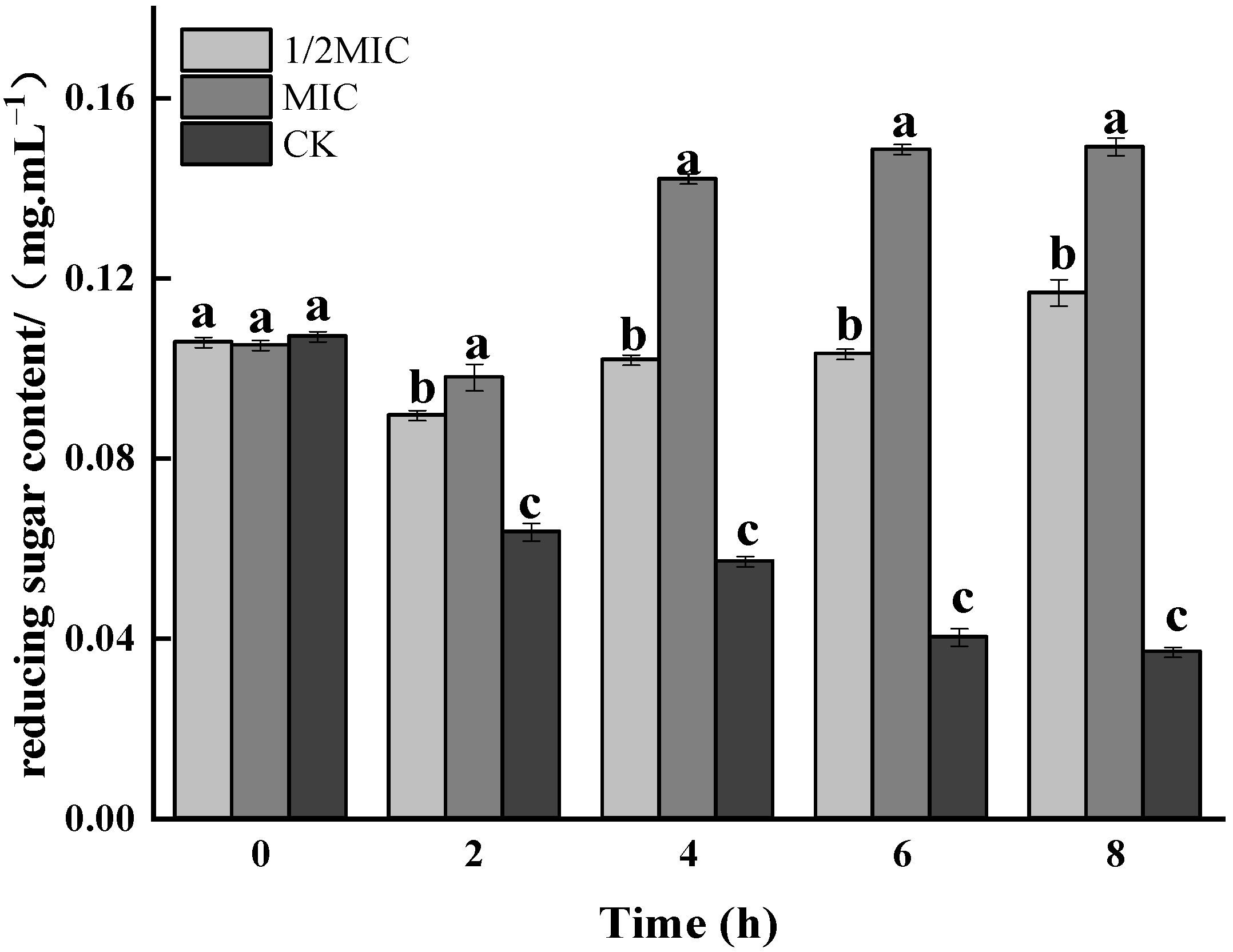
3.2. Determination of Protein Carbonylation
3.3. The EFFECT of CEO on Omp Gene Transcription Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pande, M.V. Review of egg-related salmonellosis and reduction strategies in United States, Australia, United Kingdom and New Zealand. Crit. Rev. Microbiol. 2018, 44, 290–303. [Google Scholar]
- Kumar, D.; Sudha, M.; Tarai, B.; Das, P. Evaluation of mean monocyte volume in septicemia caused by Salmonella species. J. Lab. Physicians 2018, 10, 397–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, L.; Moraes, A.; Costa, K.; Marcos, J.; Faria, L. Bioactive Natural Compounds and Antioxidant Activity of Essential Oils from Spice Plants: New Findings and Potential Applications. Biomolecules 2020, 10, 988. [Google Scholar] [CrossRef]
- Ortiz, C. Effect of Essential Oils on the Inhibition of Biofilm and Quorum Sensing in Salmonella enteritidis 13076 and Salmonella typhimurium 14028. Antibiotics 2021, 10, 1191. [Google Scholar]
- Valdivieso-Ugarte, M.; Plaza-Díaz, J.; Gomez-Llorente, C.; Gómez, E.L.; Gil, N. In vitro examination of antibacterial and immunomodulatory activities of cinnamon, white thyme, and clove essential oils. J. Funct. Foods 2021, 81, 104436. [Google Scholar] [CrossRef]
- Yang, Y.L.; Al-Mahdy, D.A.; Wu, M.L.; Zheng, X.T.; Ge, Y.W. LC-MS-based identification and antioxidant evaluation of small molecules from the cinnamon oil extraction waste. Food Chem. 2021, 366, 130576. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Sun, F.; Wang, S.; Wang, W.; Dong, J.; Gao, F. Enhanced extraction of essential oil from Cinnamomum cassia bark by ultrasound assisted hydrodistillation. Chin. J. Chem. Eng. 2020, 36, 38–46. [Google Scholar] [CrossRef]
- Pima, B.; Jk, P.B.; Mad, B. Intensified approach towards isolation of cinnamon oil using microwave radiation: Parametric, optimization and comparative studies. Ind. Crops Prod. 2021, 173, 114088. [Google Scholar]
- Mortazavi, N.; Aliakbarlu, J. Antibacterial effects of ultrasound, cinnamon essential oil, and their combination against listeria monocytogenes and salmonella typhimurium in milk. J. Food Sci. 2019, 84, 3700–3706. [Google Scholar] [CrossRef]
- Somrani, M.; Debbabi, H.; Abidi, F.; Palop, A. Garlic, onion, and cinnamon essential oil anti-biofilms′ effect against listeria monocytogenes. Foods 2020, 9, 567. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.J.; Kim, H.E.; Han, S.J.; Choi, J.S. Antibacterial and antibiofilm activities of cinnamon essential oil nanoemulsion against multi-species oral biofilms. Sci. Rep. 2021, 11, 5911. [Google Scholar] [CrossRef]
- He, J.; Wu, D.; Zhang, Q.; Chen, H.; Li, H.; Han, Q. Efficacy and mechanism of cinnamon essential oil on inhibition of colletotrichum acutatum isolated from ‘hongyang′ kiwifruit. Front Microbiol. 2018, 9, 1288. [Google Scholar] [CrossRef] [Green Version]
- Elcocks, E.R.; Spencer-Phillips, P.T.N.; Adukwu, E.C. Rapid bactericidal effect of cinnamon bark essential oil against Pseudomonas aeruginosa. J. Appl. Microbiol. 2020, 128, 1025–1037. [Google Scholar] [CrossRef]
- Hicksa, J.M.; Halkerstonb, R.; Silmanb, N.; Jacksonc, S.K.; Aylotta, J.W.; Rawson, F.J. Real-time bacterial detection with an intracellular ROS sensing platform. J. Biosens. Bioelectron. 2019, 141, 111430. [Google Scholar] [CrossRef]
- Chen, Q.; Yang, G. Signal Function Studies of ROS, Especially RBOH-Dependent ROS, in Plant Growth, Development and Environmental Stress. J. Plant Growth Regul. 2019, 39, 157–171. [Google Scholar] [CrossRef]
- Nantapong, N.; Murata, R.; Trakulnaleamsai, S.; Kataoka, N.; Yakushi, T.; Matsushita, K. The effect of reactive oxygen species (ros) and ros-scavenging enzymes, superoxide dismutase and catalase, on the thermotolerant ability of corynebacterium glutamicum. Appl. Microbiol Biot. 2019, 103, 5355–5366. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, E. Antimicrobials functioning through ros-mediated mechanisms: Current insights. Microorganisms 2021, 10, 61. [Google Scholar]
- Hu, S.; Yu, Y.; Lv, Z.; Shen, J.; Ke, Y.; Xiao, X. Proteomics study unveils ROS balance in acid-adapted Salmonella Enteritidis. Food Micro. 2020, 92, 103585. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, K.; Shi, Y.; Shao, C. The tango of ros and p53 in tissue stem cells. Cell Death Differ. 2018, 25, 639–641. [Google Scholar] [CrossRef] [Green Version]
- Franklin, M.W.; Nepomnyachyi, S.; Feehan, R.; Ben-Tal, N.; Slusky, J.S. Evolutionary pathways of repeat protein topology in bacterial outer membrane proteins. Elife 2018, 7, e40308. [Google Scholar] [CrossRef]
- Rocker, A.; Lacey, J.A.; Belousoff, M.J.; Wilksch, J.J.; Strugnell, R.A.; Davies, M.R.; Lithgow, T. Global Trends in Proteome Remodeling of the Outer Membrane Modulate Antimicrobial Permeability in Klebsiella pneumoniae. mBio 2020, 11, e00603–e00620. [Google Scholar] [CrossRef] [Green Version]
- Futoma-Kołoch, B.; Bugla-Płoskonska, G.; Dudek, B.; Dorotkiewicz-Jach, A.; Drulis-Kawa, Z.; Gamian, A. Outer membrane proteins of salmonella as potential markers of resistance to serum, antibiotics and biocides. Curr. Med. Chem. 2018, 26, 1960–1978. [Google Scholar] [CrossRef]
- He, W.; Yu, G.; Li, T.; Bai, L.; Yang, Y.; Xue, Z.; Pang, Y.; Reichmann, D.; Hiller, S.; He, L.; et al. Chaperone Spy Protects Outer Membrane Proteins from Folding Stress via Dynamic Complex Formation. mBio 2021, 12, e0213021. [Google Scholar] [CrossRef]
- Guan, Q.; Bhowmick, B.; Upadhyay, A.; Han, Q. Structure and functions of bacterial outer membrane protein A, a potential therapeutic target for bacterial infection. Curr. Top Med. Chem. 2021, 21, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Radünz, M.; Trindade, M.L.M.D.; Camargo, T.M.; Radünz, A.L.; Helbig, E. Antimicrobial and antioxidant activity of unencapsulated and encapsulated clove (syzygium aromaticum, l.) essential oil. Food Chem. 2018, 276, 180–186. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Liu, J.; Wang, Y. Effects of oxygen and water content on microbial distribution in the polyurethane foam cubes of a biofilter for SO2 removal. J. Environ. Sci. 2018, 63, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, M.A.; Roeschlin, R.A.; Favaro, M.A.; Uviedo, F.; Campos-Beneyto, L.; D′Andrea, R.; Marano, M.R. Plant responses underlying nonhost resistance of Citrus limon against Xanthomonas campestris pv. campestris. Mol. Plant Pathol. 2019, 20, 254–269. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.A.; Tan, B.L.U.; Nonato, M.G.; An, S.S.A. Neuroprotective effects on amyloid-beta induced cytotoxicity of Pandanus clementis Merr. 3 Biotech 2021, 11, 330. [Google Scholar] [CrossRef]
- Ramazani, E.; Yazdfazeli, M.; Emami, S.A.; Mohtashami, L.; Tayarani-Najaran, Z. Protective effects of cinnamomum verum, cinnamomum cassia and cinnamaldehyde against 6-ohda-induced apoptosis in pc12 cells. Mol. Biol. Rep. 2020, 47, 2437–2445. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.J.; Kasote, D.M.; Jayaprakasha, G.K.; Avila, C.A.; Patil, B.S. Effect of production system and inhibitory potential of aroma volatiles on polyphenol oxidase and peroxidase activities of tomatoes. J. Sci. Food Agric. 2020, 101, 307–314. [Google Scholar] [CrossRef]
- Zepeda-Arce, R.; Rojas-Garcfa, A.E.; Benitez-Trinidad, A.; Herrera-Moreno, J.F.; Medina-Diaz, I.M.; Barron-Vivanco, B.S.; Villegas, G.P.; Hernandez-Ochoa, I.; Heredia, M.d.J.S.; Bernal-Hernandez, Y.Y. Oxidative stress and genetic damage among workers exposed primarily to organophosphate and pyrethroid pesticides. Environ. Toxicol. 2017, 32, 1754–1764. [Google Scholar] [CrossRef]
- Mohammad, M.O.; Omid, F.; Khadijeh, M.; Akram, J.; Mohammadreza, A.; Sirus, H.; Negar, A.; Hossein, N.; Reza, H. Betaine supplementation mitigates intestinal damage and decreases serum bacterial endotoxin in cirrhotic rats—Sciencedirect. Pharmanutrition 2020, 12, 100179. [Google Scholar]
- Hyeon, J.Y.; Mann, D.A.; Wang, J.; Kim, W.K.; Deng, X. Rapid detection of salmonella in poultry environmental samples using real-time pcr coupled with immunomagnetic separation and whole genome amplification. Poult. Sci. 2019, 98, 6973–6979. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.T.; Wang, S.X.; Pei, D.; Qu, L.J.; Li, Y.; Chen, J.J.; Di, D.L.; Gao, K. Antibacterial Activity of Hydroxytyrosol Acetate from Olive Leaves (Olea Europaea L.). Nat. Prod. Commun. 2018, 32, 1967–1970. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R. ROS-induced ROS Release in Plant and Animal Cells. Free Radic. Bio. Med. 2017, 122, 21–27. [Google Scholar] [CrossRef]
- Thais, D.S.A.; João, M.A.R.D.C.; Fabio, D.O.S.R.; Antonia, C.D.J.O.; Jhones, D.N.D.; Alyne, R.D.A.; Ayslan, B.B.; Manoel, D.P.B.; Taiane, M.D.O.; Miguel Peixoto, D.A.; et al. Nanoemulsion of cashew gum and clove essential oil (Ocimum gratissimum Linn) potentiating antioxidant and antimicrobial activity. Int. J. Biol. Macromol. 2021, 193, 100–108. [Google Scholar]
- Bhattacharya, P.; Dey, A.; Neogi, S. An insight into the mechanism of antibacterial activity by magnesium oxide nanoparticles. J. Mater. Chem. B 2021, 9, 5329–5339. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zha, Y.; Zhong, Z.; Ruan, Y.; Hou, S. Improved detection of reactive oxygen species by DCFH-DA: New insight into self-amplification of fluorescence signal by light irradiation. Sen. Actua. B Chem. 2021, 339, 129878. [Google Scholar] [CrossRef]
- Das, S.; Horváth, B.; Afranko, S.; Joki, S.; Széchenyi, A.; Kőszegi, T. Antimicrobial activity of chamomile essential oil: Effect of different formulations. Molecules 2019, 24, 4321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timoumi, R.; Salem, I.B.; Amara, I.; Annabi, E.; Abid-Essefi, S. Protective effects of fennel essential oil against oxidative stress and genotoxicity induced by the insecticide triflumuron in human colon carcinoma cells. Environ. Sci. Pollut. Res. 2020, 27, 7957–7966. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Cai, Z.; Ba, L.; Qin, Y.; Su, X.; Luo, D.; Shan, W.; Kuang, J.; Lu, W.; Li, L.; et al. Maintenance of Postharvest Quality and Reactive Oxygen Species Homeostasis of Pitaya Fruit by Essential Oil p-Anisaldehyde Treatment. Foods 2021, 10, 2434. [Google Scholar] [CrossRef]
- Sandra, B.; Elisabetta, C.; Alessia, T.; Angela, G.; Consuelo, M.; Luigi, P.; Giacomo, M.; Lorena, U.; Carla, E.; Brunella, T. Effect of Curcumin on Protein Damage Induced by Rotenone in Dopaminergic PC12 Cells. Int. J. Mol. Sci. 2021, 21, 2761. [Google Scholar]
- Seixas, A.F.; Quendera, A.P.; Sousa, J.P.; Silva, A.F.Q.; Arraiano, C.M.; Andrade, J.M. Bacterial Response to Oxidative Stress and RNA Oxidation. Front Genet. 2022, 12, 821535. [Google Scholar] [CrossRef]
- Luo, K.Y.; Zhao, P.Y.; He, Y.F.; Kang, S.G.; Shen, C.Y.; Wang, S.; Guo, M.X.; Wang, L.H.; Shi, C. Antibacterial Effect of Oregano Essential Oil against Vibrio vulnificus and Its Mechanism. Foods 2002, 11, 2304–8158. [Google Scholar] [CrossRef]
- Akagawa, M. Protein carbonylation: Molecular mechanisms, biological implications, and analytical approaches. Free. Radic. Res. 2020, 55, 31–37. [Google Scholar] [CrossRef]
- Rodríguez-García, A.; García-Vicente, R.; Morales, M.L.; Ortiz-Ruiz, A.; Linares, M. Protein Carbonylation and Lipid Peroxidation in Hematological Malignancies. Antioxid. Basel 2020, 9, 1212. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, X.; Su, Y.; Xu, W.; Wang, J. Dimethyl phthalate damaged the cell membrane of Escherichia coli K12. Ecotox. Environ. Safe 2019, 180, 208–214. [Google Scholar] [CrossRef]
- Hill, R.L.; Singh, I.N.; Brelsfoard, J.; Hall, E.D. Pharmacological inhibition of lipid peroxidative damage by the 21-aminosteroid U-74389G improves cortical mitochondrial function following traumatic brain injury in young adult male rats. Neuropharmacology 2022, 170, 108023. [Google Scholar] [CrossRef]
- Chen, Z.W.; Song, S.F.; Wen, Y.Z.; Zou, Y.Q.; Liu, H.J. Toxicity of Cu (II) to the green alga Chlorella vulgaris: A perspective of photosynthesis and oxidant stress. Environ. Sci. Pollut. R 2016, 23, 17910–17918. [Google Scholar] [CrossRef]
- Guest, R.L.; Raivio, T.L. Role of the Gram-Negative Envelope Stress Response in the Presence of Antimicrobial Agents. Trends Microbiol. 2016, 24, 377–390. [Google Scholar] [CrossRef]
- Jozef, K.; Petr, B.; Josef, H.; Olga, K.; Ivo, P. Physiology and methodology of chromium toxicity using alga Scenedesmus quadricauda as model object. Chemosphere 2015, 120, 23–30. [Google Scholar]
- Niu, L.Y.; Zhang, Y.L.; Jie, M.S.; Cheng, Y.X.; Xiang, Q.S.; Zhang, Z.J.; Bai, Y.H. Synergetic effect of petit-high pressure carbon dioxide combined with cinnamon (Cinnamomum cassia) essential oil against Salmonella typhimurium. Int. J. Food Sci. Tech. 2022, 57, 2954–2967. [Google Scholar] [CrossRef]
- Rouco, L.; Alvario, R.; Alfonso, A.; Romero, M.J.; Pedrido, R.; Maneiro, M. Neuroprotective effects of fluorophore-labelled manganese complexes: Determination of ROS production, mitochondrial membrane potential and confocal fluorescence microscopy studies in neuroblastoma cells. J. Inorg. Biochem. 2022, 227, 111670. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhang, L.L.; Li, J.; He, X.J.; Cai, J.C. Bioaccumulation and tolerance characteristics of a submerged plant (Ceratophyllum demersum L.) exposed to toxic metal lead. Ecotox Environ. Safe 2015, 122, 313–321. [Google Scholar] [CrossRef]
- Gao, M.; Feng, L.; Jiang, T. Browning inhibition and quality preservation of button mushroom (Agaricus bisporus) by essential oils fumigation treatment. Food Chem. 2014, 149, 107–113. [Google Scholar] [CrossRef]
- An, P.P.; Yang, X.B.; Yu, J.; Qi, J.R.; Ren, X.Y. α-terpineol and terpene-4-ol, the critical components of tea tree oil, exert antifungal activities in vitro and in vivo against Aspergillus niger in grapes by inducing morphous damage and metabolic changes of fungus. Food Control. 2019, 98, 42–53. [Google Scholar] [CrossRef]
- Blaskó, Á.; Zoltán, G.; Pál, G.; Gábor, M.; Szilvia, S.; Judit, K.; Csaba, V.; Lilla, M.; Miklós, P. Effects of clary sage oil and its main components, linalool and linalyl acetate, on the plasma membrane of Candida albicans: An in vivo EPR study. Apoptosis 2017, 22, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Medvedeva, N.; Zaytseva, T.; Kuzikova, I. Cellular responses and bioremoval of nonylphenol by the bloom-forming cyanobacterium Planktothrix agardhii 1113. J. Mar. Syst. 2017, 171, 120–128. [Google Scholar] [CrossRef]
- Mandras, N.; Nostro, A.; Roana, J.; Scalas, D.; Banche, G.; Ghisetti, V.; Del Re, S.; Fucale, G.; Cuffini, A.M.; Tullio, V. Liquid and vapour-phase antifungal activities of essential oils against Candida albicans and non-albicans Candida. BMC Complement. Altern. Med. 2016, 16, 330. [Google Scholar] [CrossRef]
- Cao, J.; Liu, H.; Wang, Y.; He, X.; Jiang, H.; Yao, J.; Xia, F.; Zhao, Y.; Chen, X. Antimicrobial and antivirulence efficacies of citral against foodborne pathogen Vibrio parahaemolyticus RIMD2210633. Food Control 2020, 120, 107507. [Google Scholar] [CrossRef]
- Xiao, M.; Yong, L.; Jian, S.; Chen, G.; Yan, A. Transcriptional Regulation of the Outer Membrane Porin Gene ompW Reveals its Physiological Role during the Transition from the Aerobic to the Anaerobic Lifestyle of Escherichia coli. Front Microbiol. 2016, 7, 799. [Google Scholar] [CrossRef] [Green Version]
- Jasmin, R.; Baker, M.A.; Zhu, Y.; Han, M.L.; Velkov, T. A Comparative Study of Outer Membrane Proteome between Paired Colistin-Susceptible and Extremely Colistin-Resistant Klebsiella pneumoniae Strains. ACS Infect Dis. 2018, 4, 1692–1704. [Google Scholar]
- Marisol, P.T.; Nuriban, V.P.; Rodolfo, P.P.; Cristina, G.C.; Christian, P.S.; Moreno-Eutimio, M.A.; Ingeborg, B.; Mayra, P.T.S.; Lourdes, A.P.; Cunningham, A.F. Salmonella Typhi Porins OmpC and OmpF Are Potent Adjuvants for T-Dependent and T-Independent Antigens. Front Immunol. 2017, 8, 230. [Google Scholar]
- Vasconcelos, N.G.; Croda, J.; Simionatto, S. Antibacterial mechanisms of cinnamon and its constituents: A review. Microb Pathogenesis 2018, 120, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, G.K.; de Oliveira, T.R.; Maia, F.C.; de Feiria, S.B.; Barbosa, J.P.; Joia, F.; Boni, G.C.; Höfling, J.F. Efficacy of true cinnamon (Cinnamomum verum) leaf essential oil as a therapeutic alternative for Candida biofilm infections. Iran J. Basic Med. Sci. 2021, 24, 787–795. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhao, Y.; Chen, X.; Li, W.; Li, W.; Du, J.; Wang, L. Effects of Cinnamon Essential Oil on Oxidative Damage and Outer Membrane Protein Genes of Salmonella enteritidis Cells. Foods 2022, 11, 2234. https://doi.org/10.3390/foods11152234
Zhang Z, Zhao Y, Chen X, Li W, Li W, Du J, Wang L. Effects of Cinnamon Essential Oil on Oxidative Damage and Outer Membrane Protein Genes of Salmonella enteritidis Cells. Foods. 2022; 11(15):2234. https://doi.org/10.3390/foods11152234
Chicago/Turabian StyleZhang, Zhen, Yuanyuan Zhao, Xueqin Chen, Wei Li, Wen Li, Jianming Du, and Li Wang. 2022. "Effects of Cinnamon Essential Oil on Oxidative Damage and Outer Membrane Protein Genes of Salmonella enteritidis Cells" Foods 11, no. 15: 2234. https://doi.org/10.3390/foods11152234
APA StyleZhang, Z., Zhao, Y., Chen, X., Li, W., Li, W., Du, J., & Wang, L. (2022). Effects of Cinnamon Essential Oil on Oxidative Damage and Outer Membrane Protein Genes of Salmonella enteritidis Cells. Foods, 11(15), 2234. https://doi.org/10.3390/foods11152234