
Hemp Seed Fermented by Aspergillus oryzae Attenuates Lipopolysaccharide-Stimulated Inflammatory Responses in N9 Microglial Cells


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
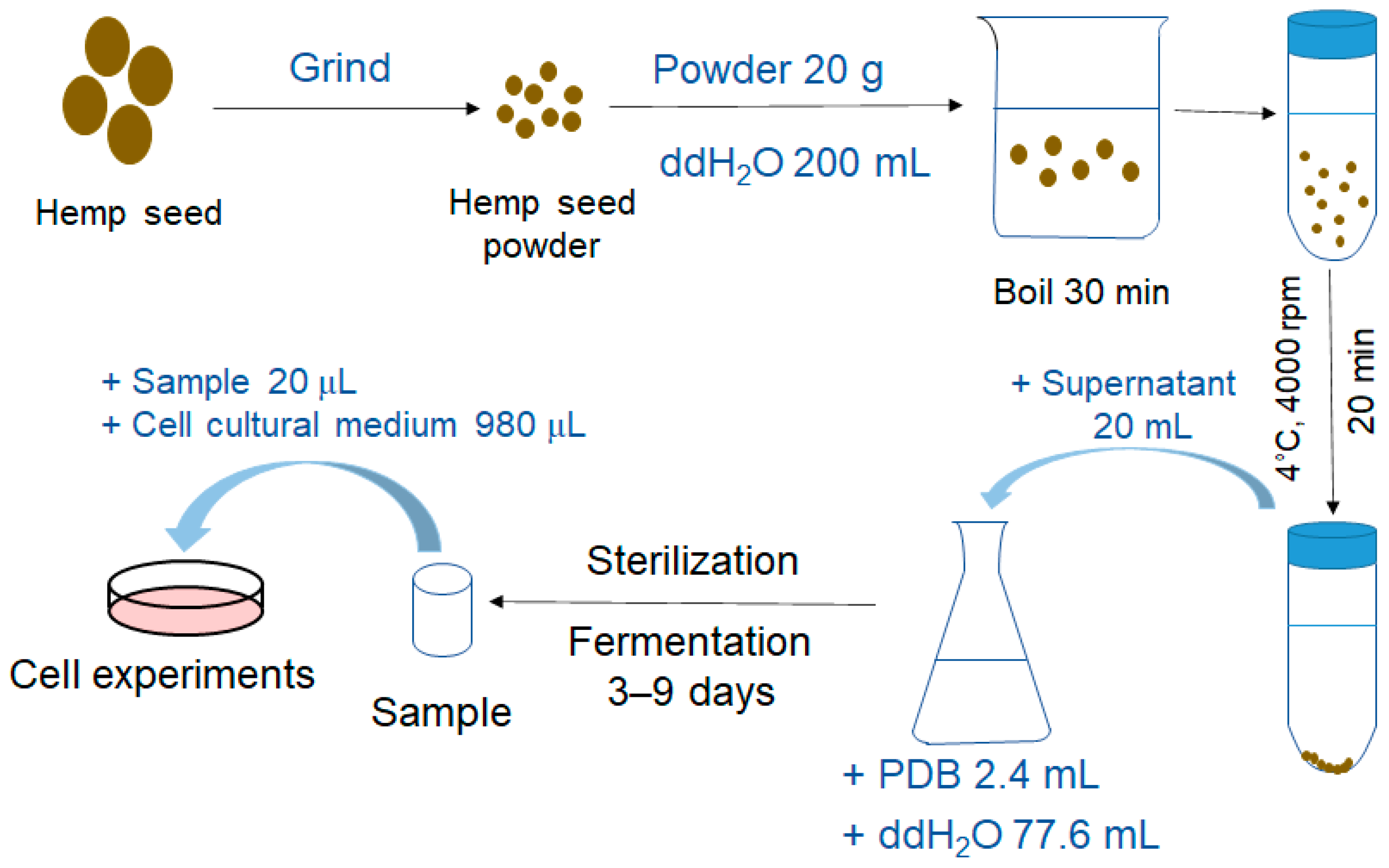
2.2. Extract Preparation and Fermentation
2.3. Cell Culture
2.4. MTT Assay
2.5. Bioassay for NO Production
2.6. Total RNA Extraction and Real-Time PCR
2.7. ELISA Assay
2.8. Western Blot Analysis
2.9. Detection of Intracellular ROS Production
2.10. Statistical Analysis
3. Results
3.1. Effects of FHS on Cell Cytotoxicity and Nitrite Production in LPS-Stimulated N9 Microglial Cells
3.2. Effects of FHS on iNOS mRNA and Protein Expression in LPS-Stimulated N9 Microglial Cells
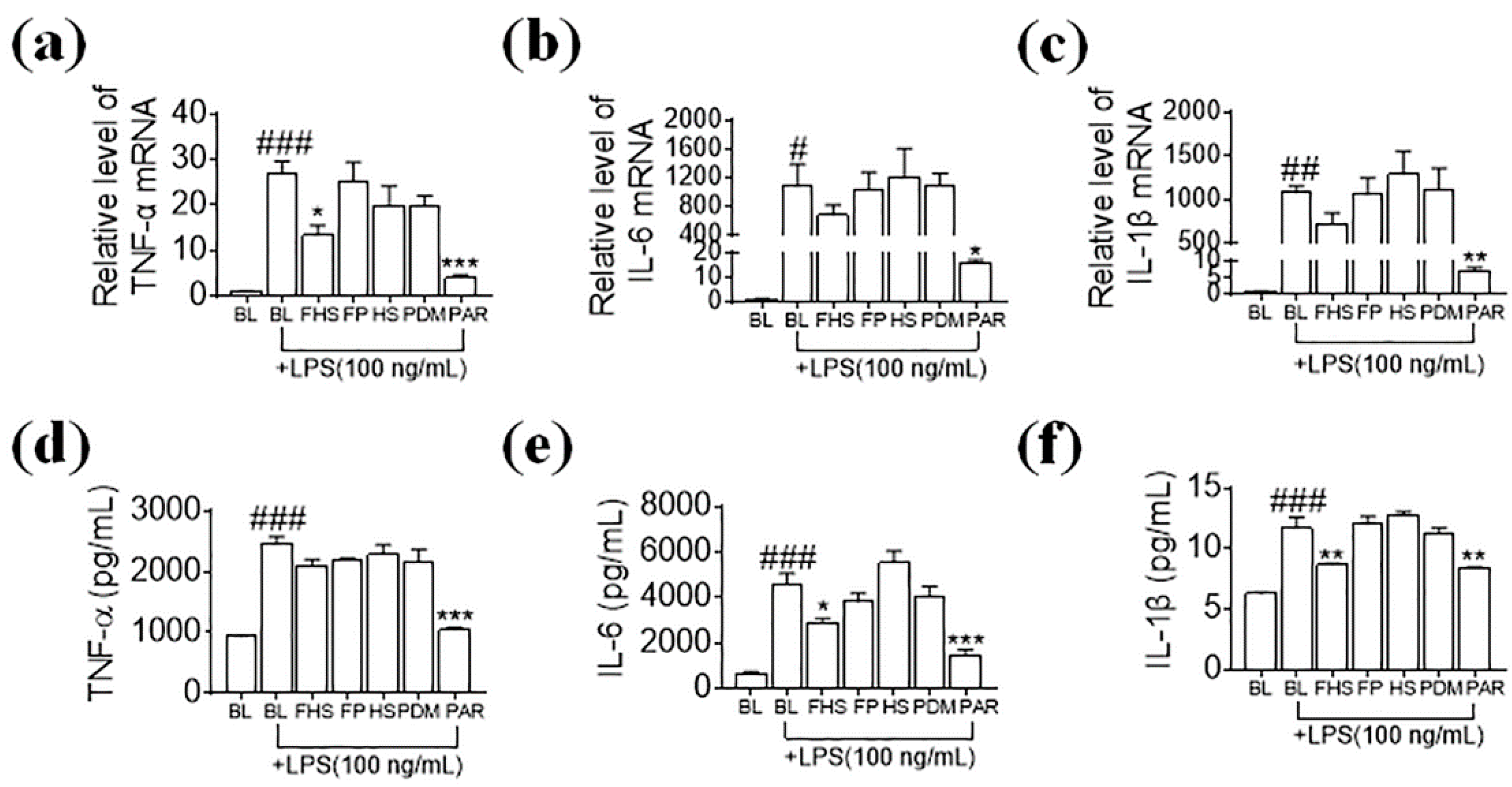
3.3. Effects of FHS on Pro-Inflammatory Cytokines in LPS-Stimulated N9 Microglial Cells
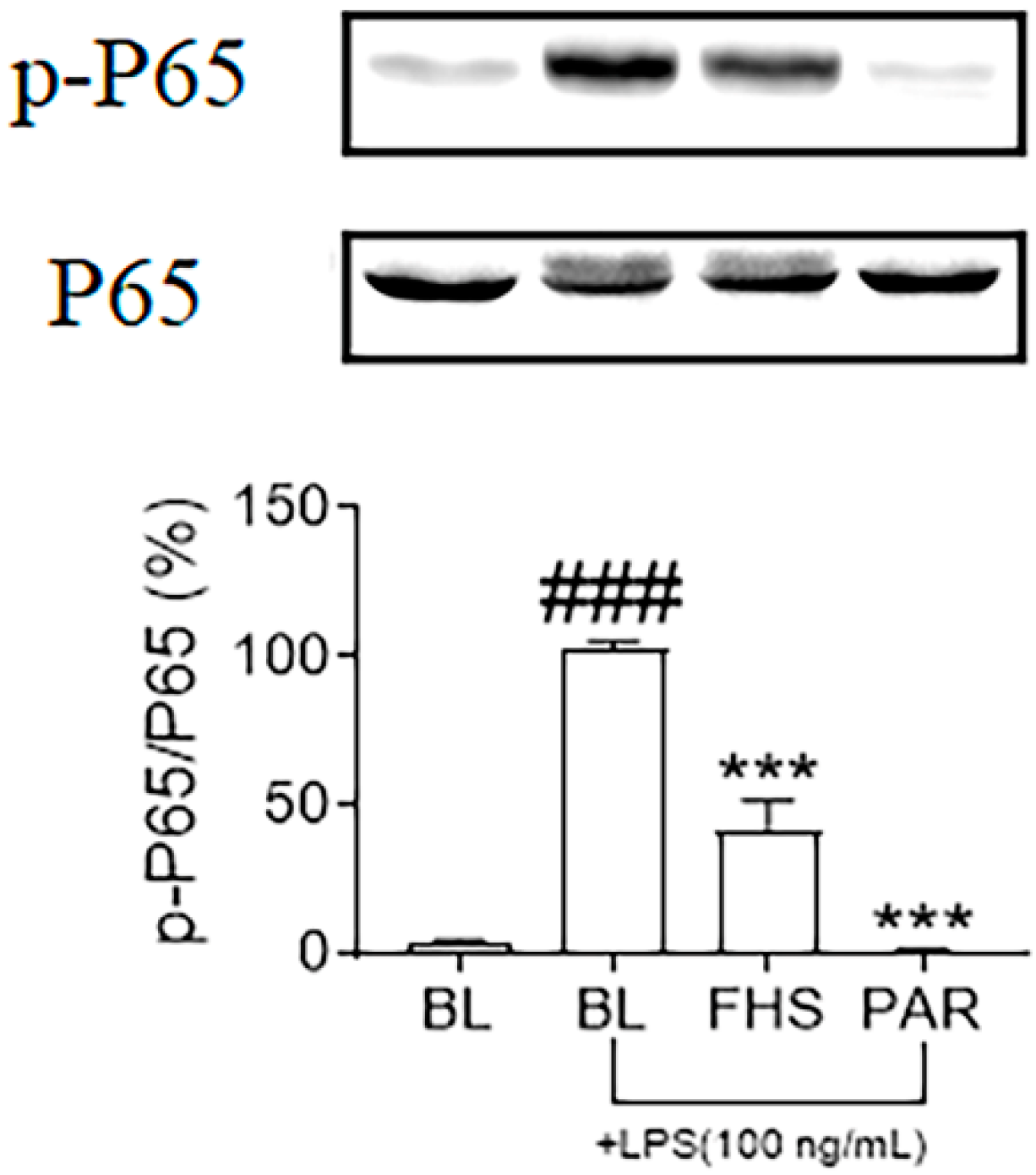
3.4. Effects of FHS on NF-κB Signaling Pathway in LPS-Stimulated N9 Microglial Cells
3.5. Effects of FHS on MAPKs and PI3K/Akt Signaling Pathways in LPS-Stimulated N9 Microglial Cells
3.6. Effects of FHS on ROS Production in LPS-Stimulated N9 Microglial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- González-Scarano, F.; Baltuch, G. Microglia as mediators of inflammatory and degenerative diseases. Annu. Rev. Neurosci. 1999, 22, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Harry, G.J. Microglia during development and aging. Pharmacol. Ther. 2013, 139, 313–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yirmiya, R.; Goshen, I. Immune modulation of learning, memory, neural plasticity and neurogenesis. Brain Behav. Immun. 2011, 25, 181–213. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Liu, Y.; Wang, T.; Wei, S.J.; Block, M.L.; Wilson, B.; Liu, B.; Hong, J.S. NADPH oxidase mediates lipopolysaccharide-induced neurotoxicity and proinflammatory gene expression in activated microglia. J. Biol. Chem. 2004, 279, 1415–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeinstra, E.; Wilczak, N.; De Keyser, J. Reactive astrocytes in chronic active lesions of multiple sclerosis express co-stimulatory molecules B7-1 and B7-2. J. Neuroimmunol. 2003, 135, 166–171. [Google Scholar] [CrossRef]
- Depino, A.M.; Earl, C.; Kaczmarczyk, E.; Ferrari, C.; Besedovsky, H.; del Rey, A.; Pitossi, F.J.; Oertel, W.H. Microglial activation with atypical proinflammatory cytokine expression in a rat model of Parkinson’s disease. Eur. J. Neurosci. 2003, 18, 2731–2742. [Google Scholar] [CrossRef]
- Su, F.; Bai, F.; Zhang, Z. Inflammatory Cytokines and Alzheimer’s Disease: A Review from the Perspective of Genetic Polymorphisms. Neurosci. Bull. 2016, 32, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.Y.; Hwang, J.H.; Park, S.Y.; Jin, Y.J.; Ko, H.C.; Moon, S.W.; Kim, S.J. Fermented guava leaf extract inhibits LPS-induced COX-2 and iNOS expression in Mouse macrophage cells by inhibition of transcription factor NF-kappaB. Phytother. Res. 2008, 22, 1030–1034. [Google Scholar] [CrossRef]
- Chen, C.C.; Chyau, C.C.; Liao, C.C.; Hu, T.J.; Kuo, C.F. Enhanced anti-inflammatory activities of Monascus pilosus fermented products by addition of ginger to the medium. J. Agric. Food Chem. 2010, 58, 12006–12013. [Google Scholar] [CrossRef]
- Jung, H.J.; Choi, H.; Lim, H.W.; Shin, D.; Kim, H.; Kwon, B.; Lee, J.E.; Park, E.H.; Lim, C.J. Enhancement of anti-inflammatory and antinociceptive actions of red ginseng extract by fermentation. J. Pharm. Pharmacol. 2012, 64, 756–762. [Google Scholar] [CrossRef]
- Rupasinghe, H.P.V.; Davis, A.; Kumar, S.K.; Murray, B.; Zheljazkov, V.D. Industrial Hemp (Cannabis sativa subsp. sativa) as an Emerging Source for Value-Added Functional Food Ingredients and Nutraceuticals. Molecules 2020, 25, 4078. [Google Scholar]
- Montserrat-de la Paz, S.; Marín-Aguilar, F.; García-Giménez, M.D.; Fernández-Arche, M.A. Hemp (Cannabis sativa L.) seed oil: Analytical and phytochemical characterization of the unsaponifiable fraction. J. Agric. Food Chem. 2014, 62, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, Z.; Wu, H.; Tang, S.; Zhang, Y.; Yang, B.; Yang, H.; Huang, L. Therapeutic effect of Moringa oleifera leaves on constipation mice based on pharmacodynamics and serum metabonomics. J. Ethnopharmacol. 2022, 282, 114644. [Google Scholar] [CrossRef] [PubMed]
- Richard, M.N.; Ganguly, R.; Steigerwald, S.N.; Al-Khalifa, A.; Pierce, G.N. Dietary hempseed reduces platelet aggregation. J. Thromb. Haemost. 2007, 5, 424–425. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Park, S.H.; Han, J.H.; Hong, Y.K.; Hwang, S.; Lee, S.; Kim, D.; Han, S.Y.; Kim, E.S.; Cho, K.S. The effects of hempseed meal intake and linoleic acid on Drosophila models of neurodegenerative diseases and hypercholesterolemia. Mol. Cells 2011, 31, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Yan, X.; Bobrovskaya, L.; Ji, M.; Yuan, H.; Lou, H.; Fan, P. Anti-neuroinflammatory effects of grossamide from hemp seed via suppression of TLR-4-mediated NF-κB signaling pathways in lipopolysaccharide-stimulated BV2 microglia cells. Mol. Cell. Biochem. 2017, 428, 129–137. [Google Scholar] [CrossRef]
- Shankar, S.K.; Mulimani, V.H. Alpha-galactosidase production by Aspergillus oryzae in solid-state fermentation. Bioresour. Technol. 2007, 98, 958–961. [Google Scholar] [CrossRef]
- Lin, C.H.; Wei, Y.T.; Chou, C.C. Enhanced antioxidative activity of soybean koji prepared with various filamentous fungi. Food Microbiol. 2006, 23, 628–633. [Google Scholar] [CrossRef]
- Chandel, A.K.; Narasu, M.L.; Chandrasekhar, G.; Manikyam, A.; Rao, L.V. Use of Saccharum spontaneum (wild sugarcane) as biomaterial for cell immobilization and modulated ethanol production by thermotolerant Saccharomyces cerevisiae VS3. Bioresour. Technol. 2009, 100, 2404–2410. [Google Scholar] [CrossRef]
- Wu, L.; Fan, Y.; Fan, C.; Yu, Y.; Sun, L.; Jin, Y.; Zhang, Y.; Ye, R.D. Licocoumarone isolated from Glycyrrhiza uralensis selectively alters LPS-induced inflammatory responses in RAW 264.7 macrophages. Eur. J. Pharmacol. 2017, 801, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.R.; Park, Y.; Li, M.; Kim, Y.K.; Lee, S.; Son, S.Y.; Lee, S.; Lee, J.S.; Lee, C.H.; Park, H.H.; et al. Anti-inflammatory effect of Ailanthus altissima (Mill.) Swingle leaves in lipopolysaccharide-stimulated astrocytes. J. Ethnopharmacol. 2022, 286, 114258. [Google Scholar] [CrossRef] [PubMed]
- Kleinert, H.; Schwarz, P.M.; Förstermann, U. Regulation of the expression of inducible nitric oxide synthase. Biol. Chem. 2003, 384, 1343–1364. [Google Scholar] [CrossRef] [PubMed]
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wu, Z.; Cao, X.; Ding, L.; Wen, Z.; Bian, J.S. HNO suppresses LPS-induced inflammation in BV-2 microglial cells via inhibition of NF-kappaB and p38 MAPK pathways. Pharmacol. Res. 2016, 111, 885–895. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Chen, B.; Kang, X.; Zhang, R.; Guo, Y.; Zhao, J.; Yang, H. Neuroprotective effects of natural compounds on LPS-induced inflammatory responses in microglia. Am. J. Transl. Res. 2020, 12, 2353–2378. [Google Scholar]
- Fan, C.; Wu, L.H.; Zhang, G.F.; Xu, F.; Zhang, S.; Zhang, X.; Sun, L.; Yu, Y.; Zhang, Y.; Ye, R.D. 4’-Hydroxywogonin suppresses lipopolysaccharide-induced inflammatory responses in RAW 264.7 macrophages and acute lung injury mice. PLoS ONE 2017, 12, e0181191. [Google Scholar]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-mediated neuroinflammation in neurodegenerative diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef]
- Hickman, S.; Izzy, S.; Sen, P.; Morsett, L.; El Khoury, J. Microglia in neurodegeneration. Nat. Neurosci. 2018, 21, 1359–1369. [Google Scholar] [CrossRef]
- Scott, M.C.; Bedi, S.S.; Olson, S.D.; Sears, C.M.; Cox, C.S. Microglia as therapeutic targets after neurological injury: Strategy for cell therapy. Expert Opin. Ther. Targets 2021, 25, 365–380. [Google Scholar] [CrossRef]
- Shang, A.; Gan, R.Y.; Xu, X.Y.; Mao, Q.Q.; Zhang, P.Z.; Li, H.B. Effects and mechanisms of edible and medicinal plants on obesity: An updated review. Crit. Rev. Food Sci. Nutr. 2021, 61, 2061–2077. [Google Scholar] [CrossRef]
- Xia, W.; Zhou, X.; Ma, J.; Li, T.; Zhang, X.; Li, J.; Xue, Y. A Review on a Medicinal and Edible Plant: Aralia elata (Miq.) Seem. Mini Rev. Med. Chem. 2021, 21, 2567–2583. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Tang, J.; dos Santos Passos, C.; Nurisso, A.; Simoes-Pires, C.A.; Ji, M.; Lou, H.; Fan, P. Characterization of Lignanamides from Hemp (Cannabis sativa L.) Seed and Their Antioxidant and Acetylcholinesterase Inhibitory Activities. J. Agric. Food Chem. 2015, 63, 10611–10619. [Google Scholar] [CrossRef] [PubMed]
- Prociuk, M.A.; Edel, A.L.; Richard, M.N.; Gavel, N.T.; Ander, B.P.; Dupasquier, C.M.; Pierce, G.N. Cholesterol-induced stimulation of platelet aggregation is prevented by a hempseed-enriched diet. Can. J. Physiol. Pharmacol. 2008, 86, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Farinon, B.; Molinari, R.; Costantini, L.; Merendino, N. The seed of industrial hemp (Cannabis sativa L.): Nutritional Quality and Potential Functionality for Human Health and Nutrition. Nutrients 2020, 12, 1935. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, S.; Lou, H.; Fan, P. Chemical constituents of hemp (Cannabis sativa L.) seed with potential anti-neuroinflammatory activity. Phytochem. Lett. 2018, 23, 57–61. [Google Scholar] [CrossRef]
- Wang, S.; Luo, Q.; Zhou, Y.; Fan, P. CLG from Hemp Seed Inhibits LPS-Stimulated Neuroinflammation in BV2 Microglia by Regulating NF-κB and Nrf-2 Pathways. ACS Omega 2019, 4, 16517–16523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wang, S.; Ji, J.; Lou, H.; Fan, P. Hemp (Cannabis sativa L.) Seed Phenylpropionamides Composition and Effects on Memory Dysfunction and Biomarkers of Neuroinflammation Induced by Lipopolysaccharide in Mice. ACS Omega 2018, 3, 15988–15995. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.Y.; Liu, C.W.; Lin, W.; Ding, Y.; Bian, Z.Y.; Huang, L.; Huang, H.; Yu, K.H.; Chen, S.B.; Sun, Y.; et al. Extract of Fructus Cannabis Ameliorates Learning and Memory Impairment Induced by D-Galactose in an Aging Rats Model. Evid. Based Complementary Altern. Med. 2017, 2017, 4757520. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Kim, H.S. Korean traditional natural herbs and plants as immune enhancing, antidiabetic, chemopreventive, and antioxidative agents: A narrative review and perspective. J. Med. Food 2014, 17, 21–27. [Google Scholar] [CrossRef]
- Nissen, L.; di Carlo, E.; Gianotti, A. Prebiotic potential of hemp blended drinks fermented by probiotics. Food Res. Int. 2020, 131, 109029. [Google Scholar] [CrossRef]
- Pontonio, E.; Verni, M.; Dingeo, C.; Diaz-de-Cerio, E.; Pinto, D.; Rizzello, C.G. Impact of Enzymatic and Microbial Bioprocessing on Antioxidant Properties of Hemp (Cannabis sativa L.). Antioxidants 2020, 9, 1258. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Jun, S.C.; Han, K.H.; Hong, S.B.; Yu, J.H. Diversity, Application, and Synthetic Biology of Industrially Important Aspergillus Fungi. Adv. Appl. Microbiol. 2017, 100, 161–202. [Google Scholar] [PubMed]
- Behera, B.C. Citric acid from Aspergillus niger: A comprehensive overview. Crit. Rev. Microbiol. 2020, 46, 727–749. [Google Scholar] [CrossRef]
- Jin, F.J.; Hu, S.; Wang, B.T.; Jin, L. Advances in Genetic Engineering Technology and Its Application in the Industrial Fungus Aspergillus oryzae. Front. Microbiol. 2021, 12, 644404. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, H.; Jantan, I.; Haque, M.A.; Kumolosasi, E. Anti-Inflammatory Effects of Hypophyllanthin and Niranthin Through Downregulation of NF-kappaB/MAPKs/PI3K-Akt Signaling Pathways. Inflammation 2018, 41, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cai, J.; Fu, Q.; Cheng, L.; Wu, L.; Zhang, W.; Zhang, Y.; Jin, Y.; Zhang, C. Anti-Inflammatory Activities of Compounds Isolated from the Rhizome of Anemarrhena asphodeloides. Molecules 2018, 23, 2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, L.J.; Surles-Zeigler, M.C.; Li, Y.; Ford, G.D.; Newman, G.D.; Ford, B.D. Regulation of inflammatory responses by neuregulin-1 in brain ischemia and microglial cells in vitro involves the NF-kappa B pathway. J. Neuroinflamm. 2016, 13, 237. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Zhang, L.; Lian, G.; Wang, X.; Zhang, H.; Yao, X.; Yang, J.; Wu, C. Sildenafil attenuates LPS-induced pro-inflammatory responses through down-regulation of intracellular ROS-related MAPK/NF-κB signaling pathways in N9 microglia. Int. Immunopharmacol. 2011, 11, 468–474. [Google Scholar] [CrossRef]
- Akira, S. Toll-like receptor signaling. J. Biol. Chem. 2003, 278, 38105–38108. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, C.; Wang, J.; Zhao, S.; Zhang, K.; Wang, J.; Zhang, W.; Wu, C.; Yang, J. Pseudoginsenoside-F11 (PF11) exerts anti-neuroinflammatory effects on LPS-activated microglial cells by inhibiting TLR4-mediated TAK1/IKK/NF-κB, MAPKs and Akt signaling pathways. Neuropharmacology 2014, 79, 642–656. [Google Scholar] [CrossRef]
- Jabs, T. Reactive oxygen intermediates as mediators of programmed cell death in plants and animals. Biochem. Pharmacol. 1999, 57, 231–245. [Google Scholar] [CrossRef]
- Rosanna, D.P.; Salvatore, C. Reactive oxygen species, inflammation, and lung diseases. Curr. Pharm. Des. 2012, 18, 3889–3900. [Google Scholar] [CrossRef] [PubMed]
- Mignolet-Spruyt, L.; Xu, E.; Idanheimo, N.; Hoeberichts, F.A.; Muhlenbock, P.; Brosche, M.; Van Breusegem, F.; Kangasjarvi, J. Spreading the news: Subcellular and organellar reactive oxygen species production and signalling. J. Exp. Bot. 2016, 67, 3831–3844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxidative Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parajuli, B.; Horiuchi, H.; Mizuno, T.; Takeuchi, H.; Suzumura, A. CCL11 enhances excitotoxic neuronal death by producing reactive oxygen species in microglia. Glia 2015, 63, 2274–2284. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Wu, L.; Fu, D.; Zhang, Y.; Zhang, C. Hemp Seed Fermented by Aspergillus oryzae Attenuates Lipopolysaccharide-Stimulated Inflammatory Responses in N9 Microglial Cells. Foods 2022, 11, 1689. https://doi.org/10.3390/foods11121689
Wang Z, Wu L, Fu D, Zhang Y, Zhang C. Hemp Seed Fermented by Aspergillus oryzae Attenuates Lipopolysaccharide-Stimulated Inflammatory Responses in N9 Microglial Cells. Foods. 2022; 11(12):1689. https://doi.org/10.3390/foods11121689
Chicago/Turabian StyleWang, Zeyuan, Lehao Wu, Dongmei Fu, Yan Zhang, and Chunzhi Zhang. 2022. "Hemp Seed Fermented by Aspergillus oryzae Attenuates Lipopolysaccharide-Stimulated Inflammatory Responses in N9 Microglial Cells" Foods 11, no. 12: 1689. https://doi.org/10.3390/foods11121689
APA StyleWang, Z., Wu, L., Fu, D., Zhang, Y., & Zhang, C. (2022). Hemp Seed Fermented by Aspergillus oryzae Attenuates Lipopolysaccharide-Stimulated Inflammatory Responses in N9 Microglial Cells. Foods, 11(12), 1689. https://doi.org/10.3390/foods11121689