
Production of Innovative Essential Oil-Based Emulsion Coatings for Fungal Growth Control on Postharvest Fruits

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials and Chemicals
2.2. Preparation of Pathogenic Fungal Isolates
2.3. BCNCs/GelA-CEO Emulsion Preparation
2.4. In Vitro Antifungal Activity of EOs and BCNC/GelA-CEO Emulsions
2.5. Antifungal Index
2.6. In vivo Antifungal Activity of EOs and BCNC/GelA-CEO Emulsions
2.7. Statistical Analysis
3. Results
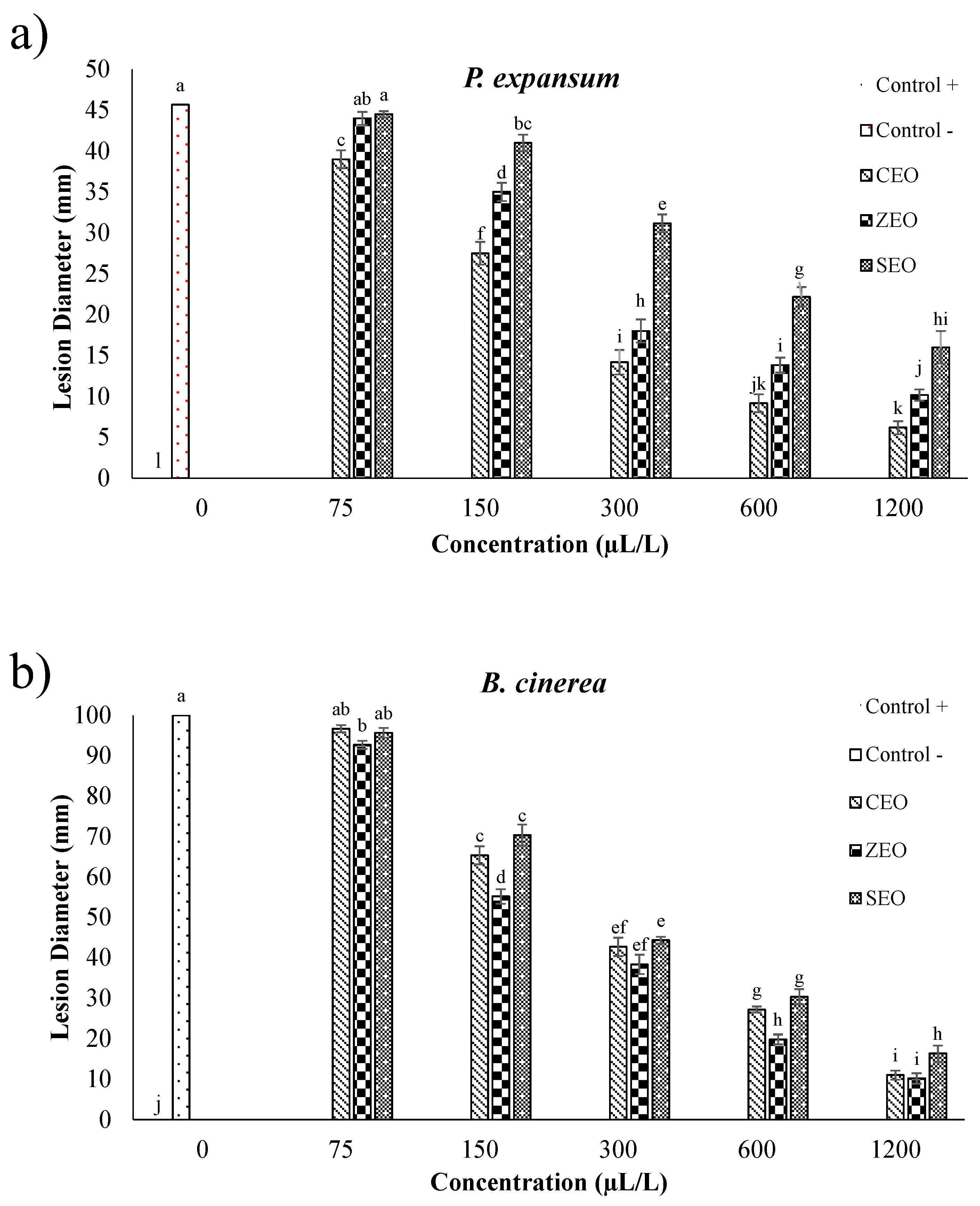
3.1. Antifungal Effects of EOs against P. expansum and B. cinerea (In Vitro Assay)
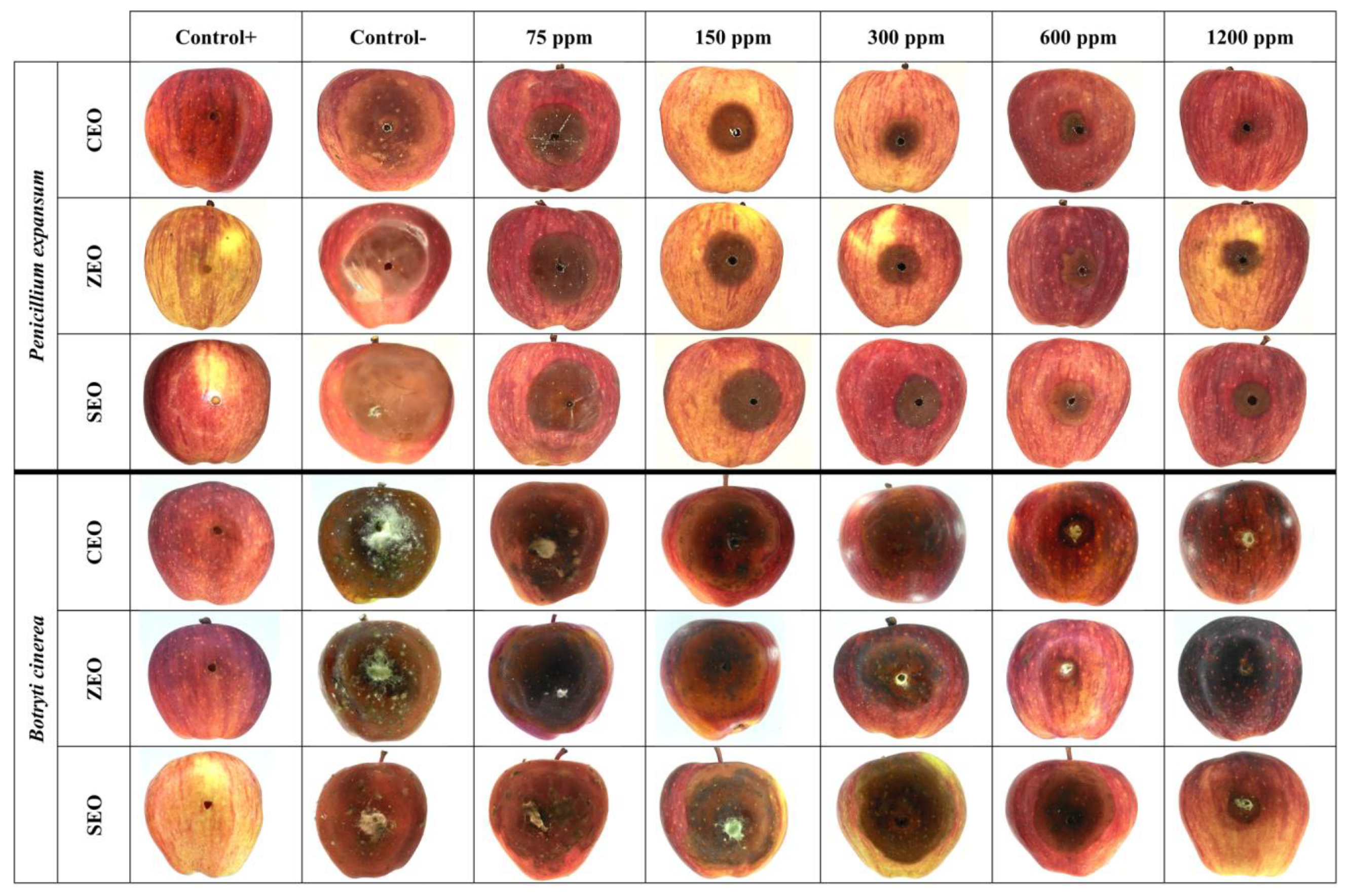
3.2. Antifungal Effects of EOs against P. expansum and B. cinerea (In Vivo Assay)
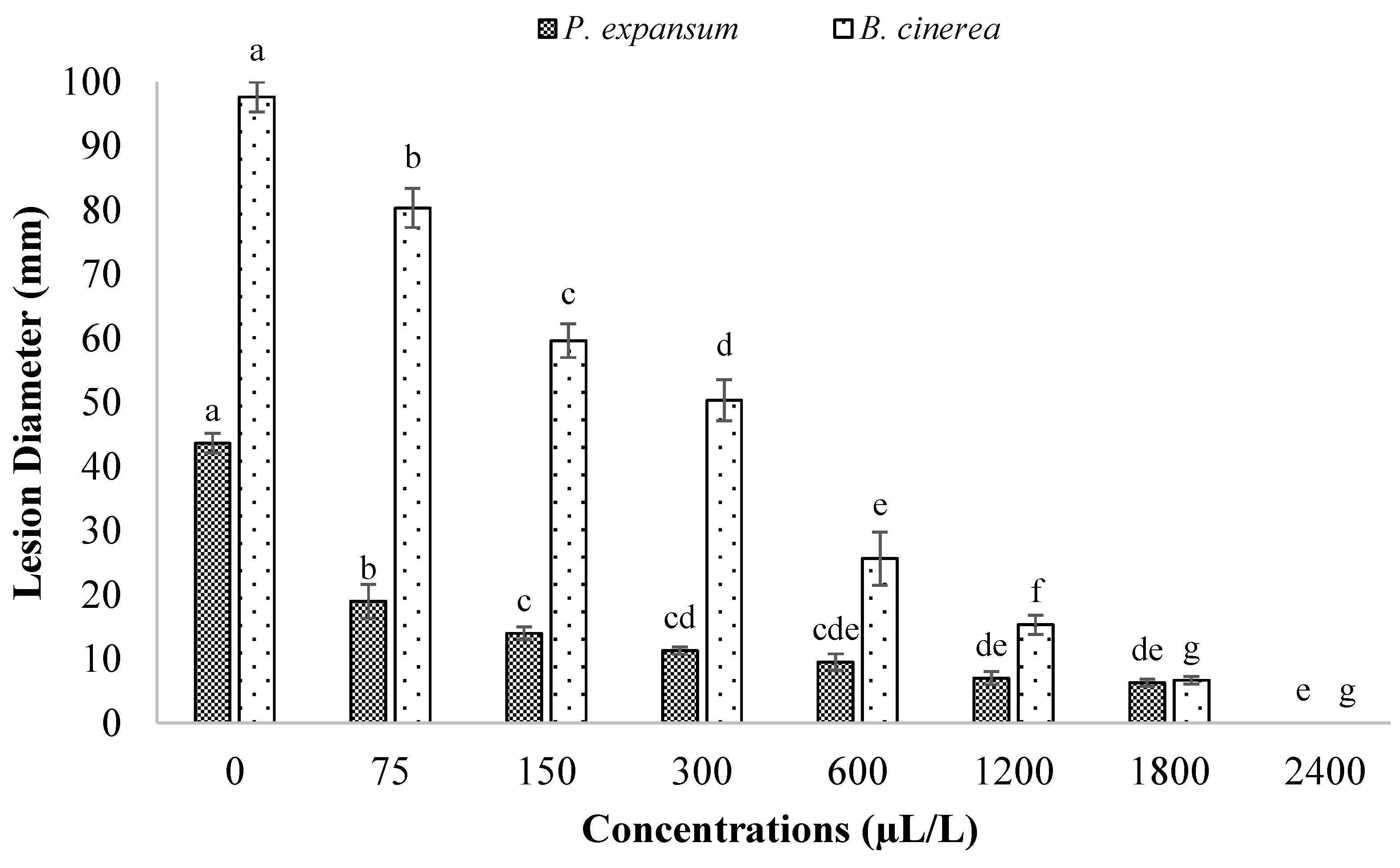
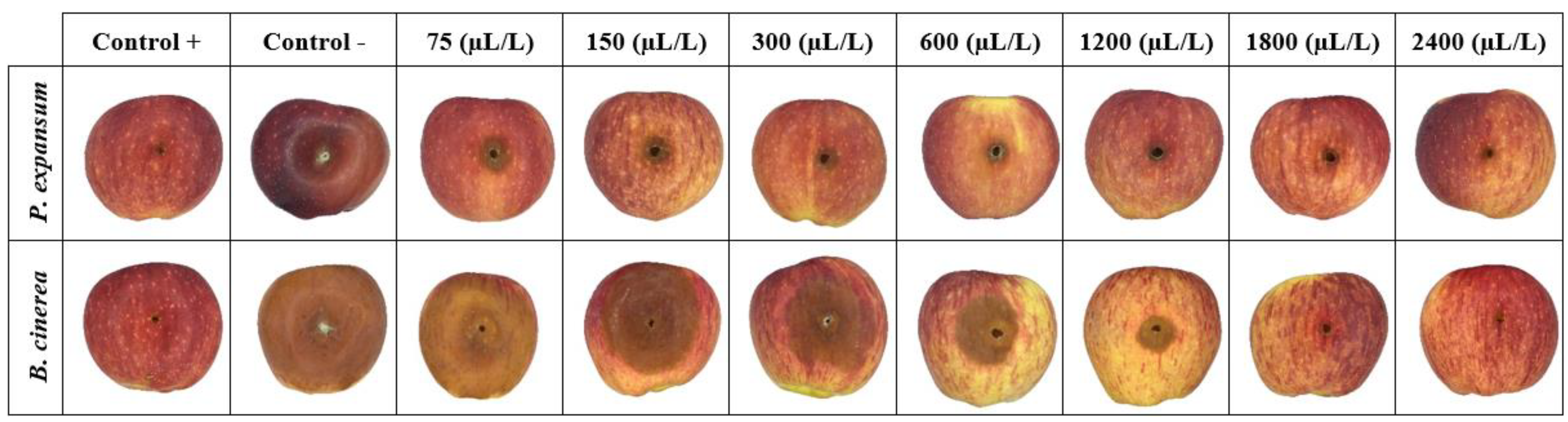
3.3. Antifungal Effects of BCNCs/GelA-CEO Emulsions against P. expansum and B. cinerea (In Vitro and In Vivo Assays)
3.4. Mechanism of Action of BCNCs/GelA-CEO Emulsion Coatings against P. expansum and B. cinerea
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dou, Y.; Routledge, M.N.; Gong, Y.; Godana, E.A.; Dhanasekaran, S.; Yang, Q.; Zhang, X.; Zhang, H. Efficacy of epsilon-poly-L-lysine inhibition of postharvest blue mold in apples and potential mechanisms. Postharvest Biol. Technol. 2021, 171, 111346. [Google Scholar] [CrossRef]
- Wallace, R.L.; Hirkala, D.L.; Nelson, L.M. Postharvest biological control of blue mold of apple by Pseudomonas fluorescens during commercial storage and potential modes of action. Postharvest Biol. Technol. 2017, 133, 1–11. [Google Scholar] [CrossRef]
- Xu, D.; Qiao, F.; Xi, P.; Lin, Z.; Jiang, Z.; Romanazzi, G.; Gao, L. Efficacy of pterostilbene suppression of postharvest gray mold in table grapes and potential mechanisms. Postharvest Biol. Technol. 2022, 183, 111745. [Google Scholar] [CrossRef]
- Becker, P.; Esker, P.; Umaña, G. Incorporation of microorganisms to reduce chemical fungicide usage in black sigatoka control programs in Costa Rica by use of biological fungicides. Crop Prot. 2021, 146, 105657. [Google Scholar] [CrossRef]
- Arras, G.; Usai, M. Fungitoxic Activity of 12 Essential Oils against Four Postharvest Citrus Pathogens: Chemical Analysis of Thymus capitatus oil and its effect in subatmospheric pressure conditions. J. Food Prot. 2001, 64, 1025–1029. [Google Scholar] [CrossRef]
- Castillo, S.; Pérez-Alfonso, C.O.; Martínez-Romero, D.; Guillén, F.; Serrano, M.; Valero, D. The essential oils thymol and carvacrol applied in the packing lines avoid lemon spoilage and maintain quality during storage. Food Control 2014, 35, 132–136. [Google Scholar] [CrossRef]
- Gniewosz, M.; Synowiec, A.; Kraśniewska, K.; Przybył, J.L.; Bączek, K.; Węglarz, Z. The antimicrobial activity of pullulan film incorporated with meadowsweet flower extracts (Filipendulae ulmariae flos) on postharvest quality of apples. Food Control 2014, 37, 351–361. [Google Scholar] [CrossRef]
- Calo, J.R.; Crandall, P.G.; O’Bryan, C.A.; Ricke, S.C. Essential oils as antimicrobials in food systems—A review. Food Control 2015, 54, 111–119. [Google Scholar] [CrossRef]
- Vergis, J.; Gokulakrishnan, P.; Agarwal, R.K.; Kumar, A. Essential oils as natural food antimicrobial agents: A Review. Crit. Rev. Food Sci. Nutr. 2013, 55, 1320–1323. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Sapper, M.; Wilcaso, P.; Santamarina, M.P.; Roselló, J.; Chiralt, A. Antifungal and functional properties of starch-gellan films containing thyme (Thymus zygis) essential oil. Food Control 2018, 92, 505–515. [Google Scholar] [CrossRef]
- Tahmasebi, M.; Golmohammadi, A.; Nematollahzadeh, A.; Davari, M.; Chamani, E. Control of nectarine fruits postharvest fungal rots caused by Botrytis cinerea and Rhizopus stolonifer via some essential oils. J. Food Sci. Technol. 2020, 57, 1647–1655. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Li, X.; Xu, Q.; Yun, J.; Lu, Y. Antifungal activities of cinnamon oil against Rhizopus nigricans, Aspergillus flavus and Penicillium expansum in vitro and in vivo fruit test. Int. J. Food Sci. Technol. 2010, 45, 1837–1842. [Google Scholar] [CrossRef]
- Behdani, M.; Pooyan, M.; Abbasi, S. Evaluation of antifungal activity of some medicinal plants essential oils against Botrytis cinerea, causal agent of postharvest apple rot, in vitro. IJACS 2012, 4, 1012–1016. [Google Scholar]
- Fathi, Z.; Hassani, A.; Ghosta, Y.; Abdollahi, A.; Meshkatalsadat, M.H. The potential of Thyme, Clove, Cinnamon and Ajowan essential oils in inhibiting the growth of Botrytis cinerea and Monilinia fructicola. J. Essent. Oil-Bear. Plants 2012, 15, 38–47. [Google Scholar] [CrossRef]
- Nikkhah, M.; Hashemi, M.; Habibi Najafi, M.B.; Farhoosh, R. Synergistic effects of some essential oils against fungal spoilage on pear fruit. Int. J. Food Microbiol. 2017, 257, 285–294. [Google Scholar] [CrossRef]
- Etemadi, N.A.; Behdad, M.; Zeinali, H. Antifungal effects of three plant essential oils against Botrytis cinerea: The cause of gray mold on strawberry. J. Res. Agric. Sci. 2011, 8, 165–170. [Google Scholar]
- Mohammadi, A.; Hashemi, M.; Hosseini, S.M. The control of Botrytis fruit rot in strawberry using combined treatments of Chitosan with Zataria multiflora or Cinnamomum zeylanicum essential oil. J. Food Sci. Technol. 2015, 52, 7441–7448. [Google Scholar] [CrossRef]
- Farzaneh, M.; Kiani, H.; Sharifi, R.; Reisi, M.; Hadian, J. Chemical composition and antifungal effects of three species of Satureja (S. hortensis, S. spicigera, and S. khuzistanica) essential oils on the main pathogens of strawberry fruit. Postharvest Biol. Technol. 2015, 109, 145–151. [Google Scholar] [CrossRef]
- Safari, N.; Hemmati, R.; Farzane, M.; Chegini, S. Study on three essential oils from Mentha piperita, Thymus daenensis and Satureja khuzistanica for controlling Penicillium expansum against of Apple Blue Mould. Appl. Res. Plant Prot. 2014, 3, 19–33. (In Persian) [Google Scholar]
- Razavi, M.S.; Golmohammadi, A.; Nematollahzadeh, A.; Fiori, F.; Rovera, C.; Farris, S. Preparation of cinnamon essential oil emulsion by bacterial cellulose nanocrystals and fish gelatin. Food Hydrocoll. 2020, 109, 106111. [Google Scholar] [CrossRef]
- Samperio, C.; Boyer, R.; Eigel, W.N., III; Holland, K.W.; McKinney, J.S.; O’Keefe, S.F.; Smith, R.; Marcy, J.E. Enhancement of plant essential oils’ aqueous solubility and stability using alpha and beta cyclodextrin. J. Agric. Food Chem. 2010, 58, 12950–12956. [Google Scholar] [CrossRef] [PubMed]
- Reineccius, G. Use of proteins for the delivery of flavours and other bioactive compounds. Food Hydrocoll. 2019, 86, 62–69. [Google Scholar] [CrossRef]
- Anal, A.K.; Shrestha, S.; Sadiq, M.B. Biopolymeric-based emulsions and their effects during processing, digestibility and bioaccessibility of bioactive compounds in food systems. Food Hydrocoll. 2019, 87, 691–702. [Google Scholar] [CrossRef]
- Razavi, M.S.; Golmohammadi, A.; Nematollahzadeh, A.; Rovera, C.; Farris, S. Cinnamon essential oil encapsulated into a fish gelatin-bacterial cellulose nanocrystals complex and active films thereof. Food Biophys. 2021, 17, 38–46. [Google Scholar] [CrossRef]
- Kraśniewska, K.; Gniewosz, M.; Synowiec, A.; Przybył, J.L.; Bączek, K.; Węglarz, Z. The application of pullulan coating enriched with extracts from Bergenia crassifolia to control the growth of food microorganisms and improve the quality of peppers and apples. Food Bioprod. Process. 2015, 94, 422–433. [Google Scholar] [CrossRef]
- Khalifa, I.; Barakat, H.; El-Mansy, H.A.; Soliman, S.A. Preserving apple (Malus domestica var. Anna) fruit bioactive substances using olive wastes extract-chitosan film coating. Inf. Process. Agric. 2017, 4, 90–99. [Google Scholar] [CrossRef]
- Yuan, G.; Chen, X.; Li, D. Chitosan films and coatings containing essential oils: The antioxidant and antimicrobial activity, and application in food systems. Food Res. Int. 2016, 89, 117–128. [Google Scholar] [CrossRef]
- Sapper, M.; Palou, L.; Pérez-Gago, M.B.; Chiralt, A. Antifungal starch-gellan edible coatings with Thyme Essential oil for the postharvest preservation of Apple and Persimmon. Coatings 2019, 9, 333. [Google Scholar] [CrossRef] [Green Version]
- Kanmani, P.; Rhim, J.W. Physical, mechanical and antimicrobial properties of gelatin based active nanocomposite films containing AgNPs and nanoclay. Food Hydrocoll. 2014, 35, 644–652. [Google Scholar] [CrossRef]
- Tongnuanchan, P.; Benjakul, S.; Prodpran, T.; Pisuchpen, S.; Osako, K. Mechanical, thermal and heat sealing properties of fish skin gelatin film containing palm oil and basil essential oil with different surfactants. Food Hydrocoll. 2016, 56, 93–107. [Google Scholar] [CrossRef]
- Rostami, M.R.; Yousefi, M.; Khezerlou, A.; Mohammadi, M.A.; Jafari, S.M. Application of different biopolymers for nanoencapsulation of antioxidants via electrohydrodynamic processes. Food Hydrocoll. 2019, 97, 105170. [Google Scholar] [CrossRef]
- Karim, A.A.; Bhat, R. Fish gelatin: Properties, challenges, and prospects as an alternative to mammalian gelatins. Food Hydrocoll. 2009, 23, 563–576. [Google Scholar] [CrossRef]
- Uysal Unalan, I.; Wan, C.; Figiel, L.; Olsson, R.T.; Trabattoni, S.; Farris, S. Exceptional oxygen barrier performance of pullulan nanocomposites with ultra-low loading of graphene oxide. Nanomaterials 2015, 26, 275703–275713. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Romero, M.C.; Murphy, T.; Morris, M.; Cummins, E.; Kerry, J.P. Antimicrobial activity of chitosan, organic acids and nano-sized solubilisates for potential use in smart antimicrobially-active packaging for potential food applications. Food Control 2013, 34, 393–397. [Google Scholar] [CrossRef]
- Alboofetileh, M.; Rezaei, M.; Hosseini, H.; Abdollahi, M. Antimicrobial activity of alginate/clay nanocomposite films enriched with essential oils against three common foodborne pathogens. Food Control 2014, 36, 1–7. [Google Scholar] [CrossRef]
- Sharma, R.; Jafari, S.M.; Sharma, S. Antimicrobial bio-nanocomposites and their potential applications in food packaging. Food Control 2020, 112, 107086. [Google Scholar] [CrossRef]
- Li, T.; Liu, Y.; Qin, Q.; Zhao, L.; Wang, Y.; Wu, X.; Liao, X. Development of electrospun films enriched with ethyl lauroyl arginate as novel antimicrobial food packaging materials for fresh strawberry preservation. Food Control 2021, 130, 108371. [Google Scholar] [CrossRef]
- Dey, A.; Pandey, G.; Rawtani, D. Functionalized nanomaterials driven antimicrobial food packaging: A technological advancement in food science. Food Control 2022, 131, 10846. [Google Scholar] [CrossRef]
- George, J.; Hatna, S. High performance edible nanocomposite films containing bacterial cellulose nanocrystals. Carbohydr. Polym. 2012, 87, 2031–2037. [Google Scholar] [CrossRef]
- Rovera, C.; Ghaani, M.; Santo, N.; Trabattoni, S.; Olsson, R.T.; Romano, D.; Farris, S. Enzymatic hydrolysis in the green production of bacterial cellulose nanocrystals. ACS Sustain. Chem. Eng. 2018, 6, 7725–7734. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Zhang, Z.; Qin, G.; Li, B.; Tian, S. Infection assays of tomato and apple fruit by the fungal pathogen Botrytis cinerea. Bio-Protocol 2014, 4, 1311. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, H.; Farzaneh, M.; A’azami, Z.A. Effect of cinnamon nanoemulsion in reducing of postharvest diseases of strawberry. Biol. Control. Pests Plant Dis. 2015, 4, 57–64. (In Persian) [Google Scholar] [CrossRef]
- Ranjbar, H.; Farzaneh, M.; Sharifii, R.; Hadian, J.; Mirjalili, M.H. Classification of selected multi-cut Persian clover germplasm of National Plant Genebank based on agronomic traits. Pajouhesh Sazandegi 2009, 81, 54–60. (In Persian) [Google Scholar]
- Taş, L.; Karaca, G. Effects of some essential oils on mycelial growth of Penicillium expansum Link and Blue Mold Severity on Apple. AJAFS 2015, 3. Available online: https://www.ajouronline.com/index.php/AJAFS/article/view/3424 (accessed on 24 April 2022).
- Fieira, C.; Oliveira, F.; Calegari, R.P.; Machado, A.; Coelho, A.R. In vitro and in vivo antifungal activity of natural inhibitors against Penicillium expansum. Ciênc. Tecnol. Aliment. 2013, 33, 40–46. [Google Scholar] [CrossRef]
- Rupasinghe, H.P.V.; Jayasankar, S.; Lay, W. Variation in total phenolics and antioxidant capacity among European plum genotypes. Sci. Hortic. 2006, 108, 243–246. [Google Scholar] [CrossRef]
- Lopez-Reyes, J.G.; Spadaro, D.; Gullino, M.L.; Garibaldi, A. Efficacy of plant essential oils on postharvest control of rot caused by fungi on four cultivars of apples in vivo. Flavour Fragr. J. 2010, 25, 171–177. [Google Scholar] [CrossRef]
- Moreno, M.A.; Vallejo, A.M.; Ballester, A.-R.; Zampini, C.; Isla, M.I.; López-Rubio, A.; Fabra, M.J. Antifungal edible coatings containing Argentinian propolis extract and their application in raspberries. Food Hydrocoll. 2020, 107, 105973. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The fungal cell wall: Candida, Cryptococcus, and Aspergillus species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef] [PubMed]
- Edwards, I.P.; Upchurch, R.A.; Zak, D.R. Isolation of fungal Cellobiohydrolase I genes from sporocarps and forest soils by PCR. Appl. Environ. Appl. Environ. Microbiol. 2008, 74, 3481–3489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, R.S. Physical Interactions of Filamentous Fungal Spores and Unicellular Fungi. Master’s Thesis, University of Stellenbosch, April 2006. Available online: https://core.ac.uk/download/pdf/37344386.pdf (accessed on 24 April 2022).
- Wardana, A.A.; Koga, A.; Tanaka, F.; Tanaka, F. Antifungal features and properties of Chitosan/Sandalwood oil Pickering emulsion coating stabilized by appropriate cellulose nanofiber dosage for fresh fruit application. Sci. Rep. 2021, 11, 18412. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; Feo, V.D. Essential Oils and Antifungal Activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Grande-Tovar, C.D.; Chaves-Lopez, C.; Serio, A.; Rossi, C.; Paparella, A. Chitosan coatings enriched with essential oils: Effects on fungi involved in fruit decay and mechanisms of action. Trends Food Sci. Technol. 2018, 78, 61–71. [Google Scholar] [CrossRef]
- Skandamis, P.N.; Nychas, G.-J.E. Effect of oregano essential oil on microbiological and physico-chemical attributes of minced meat stored in air and modified atmospheres. J. Appl. Microbiol. 2001, 91, 1011–1022. [Google Scholar] [CrossRef]
- Da Rocha Neto, A.C.; Navarro, B.B.; Canton, L.; Maraschin, M.; Di Piero, R.M. Antifungal activity of palmarosa (Cymbopogon martinii), tea tree (Melaleuca alternifolia) and star anise (Illicium verum) essential oils against Penicillium expansum and their mechanisms of action. LWT-Food Sci. Technol. 2019, 105, 385–392. [Google Scholar] [CrossRef]
- Selvakumaran, J.; Jell, G. A guide to basic cell culture and applications in biomaterials and tissue engineering. In Biomaterials, Artificial Organs and Tissue Engineering, 1st ed.; Woodhead Publishing Series in Biomaterials; Hench, L.L., Jones, J.R., Eds.; Woodhead Publishing: Cambridge, UK, 2005; pp. 215–226. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. expansum | B. cinerea | ||||||
|---|---|---|---|---|---|---|---|
| Concentration [(μL/L) | Time (Days) | Time (Days) | |||||
| 3 | 9 | 15 | 2 | 5 | 7 | ||
| CEO | 75 | b 44.0 ± 2.6 a,C | b 38.7 ± 2.5 a,B | a 29.3 ± 1.8 a,C | b 18.1 ± 3.5 a,AB | a 5.4 ± 1.2 a,A | a 3.4 ± 0.5 a,A |
| 150 | b 67.2 ± 4.6 b,C | b 64.7 ± 5.3 b,C | a 55.4 ± 1.1 b,C | b 100.00 ± 0.0 b,B | a 90.4 ± 0.3 b,B | a 89.6 ± 1.3 b,B | |
| 300 | 100.0 ± 0.0 c,B | 100.0 ± 0.0 c,B | 100.0 ± 0.0 c,B | 100.0 ± 0.0 b,B | 100.0 ± 0.0 c,B | 100.0 ± 0.0 c,B | |
| 600 | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | |
| 1200 | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | |
| ZEO | 75 | b 22.1 ± 1.8 a,B | ab 17.2 ± 2.5 a,A | a 15.2 ± 1.7 a,B | 11.9 ± 6.4 a,A | 14.6 ± 2.1 a,B | 11.8 ± 1.7 a,C |
| 150 | c 46.2 ± 1.2 b,B | b 40.1 ± 1.0 b,B | a 34.1 ± 1.3 b,B | 100.0 ± 0.0 b,B | 100.0 ± 0.0 b,C | 100.0 ± 0.0 b,C | |
| 300 | 100.0 ± 0.0 c,B | 100.0 ± 0.0 c,B | 100.0 ± 0.0 c,B | 100.0 ± 0.0 b,B | 100.0 ± 0.0 b,B | 100.0 ± 0.0 b,B | |
| 600 | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 b,A | |
| 1200 | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 b,A | |
| SEO | 75 | b 13.8 ± 1.2 a,A | ab 12.1 ± 2.5 a,A | a 8.5 ± 1.7 a,A | c 25.4 ± 1.7 b,B | b 19.2 ± 1.0 a,C | a 7.4 ± 0.4 a,B |
| 150 | b 33.8 ± 1.2 b,A | b 31.5 ± 1.7 b,A | a 25.5 ± 2.2 b,A | a 52.5 ± 6.1 c,A | b 65.4 ± 4.2 b,A | a 48.5 ± 3.9 b,A | |
| 300 | 78.0 ± 4.3 c,A | 74.6 ± 4.0 c,A | 69.2 ± 3.9 c,A | a 7.4 ± 0.4 a,A | c 83.8 ± 1.0 c,A | b 81.3 ± 1.1 c,A | |
| 600 | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | |
| 1200 | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | 100.0 ± 0.0 d,A | |
| Concentration (μL/L) | Time (Days) | |||||||
|---|---|---|---|---|---|---|---|---|
| 2 | 5 | 7 | 9 | 13 | 15 | 20 | ||
| P. expansum | 75 | 82.4 ± 2.0 a,E | 74.1 ± 17.1 a,DE | 56.7 ± 1.5 a,CD | 44.8 ± 1.5 a,BC | 28.9 ± 5.4 a,AB | 19.4 ± 3.9 a,A | 12.9 ± 0.5 a,A |
| 150 | 95.9 ± 6.3 b,E | 76.5 ± 6.7 ab,D | 67.8 ± 6.9 a,CD | 54.6 ± 7.6 b,C | 39.2 ± 2.0 a,B | 23.7 ± 3.2 a,A | 20.0 ± 1.0 ab,A | |
| 300 | 100.0 ± 0.0 c,D | 94.4 ± 8.6 ab,D | 92.6 ± 11.5 b,D | 87.4 ± 9.9 c,CD | 62.9 ± 9.6 b,BC | 45.8 ± 9.0 b,AB | 31.1 ± 10.6 b,A | |
| 600 | 100.0 ± 0.0 c,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | |
| 1200 | 100.0 ± 0.0 c,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | |
| B. cinerea | 75 | 69.3 ± 3.0 a,D | 54.3 ± 4.6 a,CD | 44.6 ± 7.2 a,BC | 31.7 ± 6.1 a,B | 5.2 ± 0.5 a,A | - | - |
| 150 | 95.4 ± 7.2 b,B | 92.8 ± 11.2 b,B | 90.7 ± 14.4 b,B | 72.6 ± 16.1 b,B | 12.6 ± 3.2 b,A | - | - | |
| 300 | 100.0 ± 0.0 b,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | - | - | |
| 600 | 100.0 ± 0.0 b,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | - | - | |
| 1200 | 100.0 ± 0.0 b,A | 100.0 ± 0.0 b,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | 100.0 ± 0.0 c,A | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadat Razavi, M.; Golmohammadi, A.; Nematollahzadeh, A.; Ghanbari, A.; Davari, M.; Carullo, D.; Farris, S. Production of Innovative Essential Oil-Based Emulsion Coatings for Fungal Growth Control on Postharvest Fruits. Foods 2022, 11, 1602. https://doi.org/10.3390/foods11111602
Sadat Razavi M, Golmohammadi A, Nematollahzadeh A, Ghanbari A, Davari M, Carullo D, Farris S. Production of Innovative Essential Oil-Based Emulsion Coatings for Fungal Growth Control on Postharvest Fruits. Foods. 2022; 11(11):1602. https://doi.org/10.3390/foods11111602
Chicago/Turabian StyleSadat Razavi, Mahsa, Abdollah Golmohammadi, Ali Nematollahzadeh, Alireza Ghanbari, Mahdi Davari, Daniele Carullo, and Stefano Farris. 2022. "Production of Innovative Essential Oil-Based Emulsion Coatings for Fungal Growth Control on Postharvest Fruits" Foods 11, no. 11: 1602. https://doi.org/10.3390/foods11111602
APA StyleSadat Razavi, M., Golmohammadi, A., Nematollahzadeh, A., Ghanbari, A., Davari, M., Carullo, D., & Farris, S. (2022). Production of Innovative Essential Oil-Based Emulsion Coatings for Fungal Growth Control on Postharvest Fruits. Foods, 11(11), 1602. https://doi.org/10.3390/foods11111602