
Drought and Heat Stress Impacts on Phenolic Acids Accumulation in Durum Wheat Cultivars

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials/Agronomic Trials
2.2. Grain Traits
2.3. Phenolic Acids Analysis
2.4. Statistical Analysis
3. Results
3.1. Grain Yield, Grain Traits, and Phenolic Acid Profile of Durum Wheat Cultivars
3.2. Effects of Genotype, Growth Conditions, Year, and Their Interactions
3.3. Relationships between Phenolic Acids, Yield Components, and Protein Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Grains Council [IGC]. World Grain Statistics 2016. 2020. Available online: https://www.igc.int/en/subscriptions/subscription.aspx (accessed on 21 May 2020).
- Ciudad-Mulero, M.; Barros, L.; Fernandes, Â.; Ferreira, I.C.; Callejo, M.J.; Matallana-González, M.C.; Fernández-Ruiz, V.; Morales, P.; Carrillo, J.M. Potential Health Claims of Durum and Bread Wheat Flours as Functional Ingredients. Nutrients 2020, 12, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabriso, N.; Massaro, M.; Scoditti, E.; Pasqualone, A.; Laddomada, B.; Carluccio, M.A. Phenolic extracts from whole wheat biofortified bread dampen overwhelming inflammatory response in human endothelial cells and monocytes: Major role of VCAM-1 and CXCL-10. Eur. J. Nutr. 2020, 59, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Shewry, P.R.; Ward, J.L. Phenolic Acids in Wheat Varieties in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2008, 56, 9732–9739. [Google Scholar] [CrossRef] [PubMed]
- De Leonardis, A.M.; Fragasso, M.; Beleggia, R.; Ficco, D.B.M.; De Vita, P.; Mastrangelo, A.M. Effects of Heat Stress on Metabolite ccumulation and Composition, and Nutritional Properties of Durum Wheat Grain. Int. J. Mol. Sci. 2015, 16, 30382–30404. [Google Scholar] [CrossRef] [Green Version]
- Aberkane, H.; Belkadi, B.; Kehel, Z.; Filali-Maltouf, A.; Tahir, I.S.A.; Meheesi, S.; Amri, A. Assessment of drought and heat tol-erance of durum wheat lines derived from interspecific crosses using physiological parameters and stress indices. Agronomy 2021, 11, 695. [Google Scholar] [CrossRef]
- Abbas, T.; Rizwan, M.; Ali, S.; Adrees, M.; Mahmood, A.; Zia-ur-Rehman, M.; Ibrahim, M.; Arshad, M.; Qayyum, M.F. Biochar application increased the growth and yield and reduced cadmium in drought stressed wheat grown in an aged contam-inated soil. Ecotoxicol. Environ. Saf. 2018, 148, 825–833. [Google Scholar] [CrossRef]
- Raheem, A.; Shaposhnikov, A.; Belimov, A.A.; Dodd, I.C.; Ali, B. Auxin production by rhizobacteria was associated with im-proved yield of wheat (Triticum aestivum L.) under drought stress. Arch Agron. Soil Sci. 2018, 64, 574–587. [Google Scholar] [CrossRef]
- Porter, J.R.; Gawith, M. Temperatures and the growth and development of wheat: A review. Eur. J. Agron. 1999, 10, 23–36. [Google Scholar] [CrossRef]
- Poudel, P.B.; Poudel, M.R. Heat Sstress effects and tolerance in wheat: A review. J. Biol. Today’s World 2020, 9, 217. [Google Scholar]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.H. Whole grain phytochemicals and health. J. Cereal Sci. 2007, 46, 207–219. [Google Scholar] [CrossRef]
- Kikuzaki, H.; Hisamoto, M.; Hirose, K.; Akiyama, K.; Taniguchi, H. Antioxidant Properties of Ferulic Acid and Its Related Compounds. J. Agric. Food Chem. 2002, 50, 2161–2168. [Google Scholar] [CrossRef]
- Laddomada, B.; Durante, M.; Minervini, F.; Garbetta, A.; Cardinali, A.; D’Antuono, I.; Caretto, S.; Blanco, A.; Mita, G. Phyto-chemical characterization and anti-inflammatory activity of extracts from the whole-meal flour of Italian durum wheat cultivars. Int. J. Mol. Sci. 2015, 16, 3512–3527. [Google Scholar] [CrossRef]
- Lempereur, I.; Rouau, X.; Abecassis, J. Genetic and Agronomic Variation in Arabinoxylan and Ferulic Acid Contents of Durum Wheat (Triticum durumL.) Grain and Its Milling Fractions. J. Cereal Sci. 1997, 25, 103–110. [Google Scholar] [CrossRef]
- Nigro, D.; Grausgruberb, H.; Guzmàn, C.; Laddomada, B. Phenolic compounds in wheat kernels: Genetic and genomic studies of biosynthesis and regulation. In Wheat Quality for Improving Processing and Health; Igrejas, G., Ikeda, T., Guzmàn, C., Eds.; Springer Sciences+Business Media: Dordrecht, The Netherlands, 2020; pp. 223–251. ISBN 978-3-030-34162-6. [Google Scholar]
- Shewry, P.R. The HEALTHGRAIN programme opens new opportunities for improving wheat for nutrition and health. BNF 2009, 34, 225–231. [Google Scholar] [CrossRef]
- Laddomada, B.; Durante, M.; Mangini, G.; D’Amico, L.; Lenucci, M.S.; Simeone, R.; Piarulli, L.; Mita, G.; Blanco, A. Genetic var-iation for phenolic acids concentration and composition in a tetraploid wheat (Triticum turgidum L.) collection. Genet Resour. Crop Evol. 2017, 64, 587–597. [Google Scholar] [CrossRef]
- Rawat, N.; Laddomada, B.; Gill, B.S. Genomics of cereal-based functional foods. In Cereal Genomics, 2nd ed.; Varshney, R.K., Gupta, P.K., Eds.; Springer Sciences+Business Media: Dordrecht, The Netherlands, 2013; pp. 247–274. [Google Scholar] [CrossRef]
- Nigro, D.; Laddomada, B.; Mita, G.; Blanco, E.; Colasuonno, P.; Simeone, R.; Gadaleta, A.; Pasqualone, A.; Blanco, A. Genome-wide association mapping of phenolic acids in durum wheat. J. Cereal. Sci. 2017, 75, 25–34. [Google Scholar] [CrossRef]
- Branković, G.; Dragičević, V.; Dodig, D.; Zorić, M.; Knežević, D.; Žilić, S.; Denčić, S.; Šurlan, G. Genotype × Environment in-teraction for antioxidants and phytic acid contents in bread and durum wheat as influenced by climate. Chil. J. Agric. Res. 2015, 75, 139–146. [Google Scholar]
- Shamloo, M.; Babawale, E.A.; Furtado, A.; Henry, R.J.; Eck, P.K.; Jones, P.J.H. Effects of genotype and temperature on accumu-lation of plant secondary metabolites in Canadian and Australian wheat grown under controlled environments. Sci. Rep. 2017, 7, 9133. [Google Scholar] [CrossRef]
- Liu, H.; Bruce, D.R.; Sissons, M.; Able, A.J.; Able, J.A. Genotype-dependent changes in the phenolic content of durum under water-deficit stress. Cereal Chem. J. 2018, 95, 59–78. [Google Scholar] [CrossRef]
- Guzmán, C.; Autrique, J.E.; Mondal, S.; Singh, R.P.; Govindan, V.; Morales-Dorantes, A.; Posadas-Romano, G.; Crossa, J.; Ammar, K.; Peña, R.J. Response to drought and heat stress on wheat quality, with special emphasis on bread-making quality, in durum wheat. Field Crop. Res. 2016, 186, 157–165. [Google Scholar] [CrossRef]
- Hernandez-Espinosa, N.; Laddomada, B.; Payne, T.; Huerta-Espino, J.; Govindan, V.; Ammar, K.; Ibba, M.I.; Pasqualone, A.; Guzman, C. Nutritional quality characterization of a set of durum wheat landraces from Iran and Mexico. LWT Food Sci. Technol. 2020, 124, 109198. [Google Scholar] [CrossRef]
- Cooper, M.; Delacy, I.H. Relationships among analytical methods used to study genotypic variation and geno-type-by-environment interaction in plant breeding multi environment experiments. Theor. Appl. Genet 1994, 88, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Saleem, M.F.; Anjum, S.A.; Shahid, M.; Afzal, I. Effect of terminal heat stress on proline, secondary metabolites and yield components of wheat (Triticum aestivum L.) genotypes. Philipp. Agric. Sci. 2017, 100, 278–286. [Google Scholar]
- FAO. The Impact of Disasters and Crises on Agriculture and Food Security; FAO: Rome, Italy, 2021. [Google Scholar]
- Fernandez-Orozco, R.; Li, L.; Harflett, C.; Shewry, P.R.; Ward, J.L. Effects of Environment and Genotype on Phenolic Acids in Wheat in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2010, 58, 9341–9352. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreasen, M.F.; Kroon, P.; Williamson, G.; Conesa, M.T.G. Intestinal release and uptake of phenolic antioxidant diferulic acids. Free. Radic. Biol. Med. 2001, 31, 304–314. [Google Scholar] [CrossRef]
- Vitaglione, P.; Napolitano, A.; Fogliano, V. Cereal dietary fibre: A natural functional ingredient to deliver phenolic com-pounds into the gut. Trends Food Sci. Tech. 2008, 19, 451–463. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Cardona, F.; Andres-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shewry, P.R.; Piironen, V.; Lampi, A.-M.; Edelmann, M.; Kariluoto, S.; Nurmi, T.; Fernandez-Orozco, R.; Ravel, C.; Charmet, G.; Andersson, A.A.M.; et al. The HEALTHGRAIN Wheat Diversity Screen: Effects of Genotype and Environment on Phytochemicals and Dietary Fiber Components. J. Agric. Food Chem. 2010, 58, 9291–9298. [Google Scholar] [CrossRef] [PubMed]
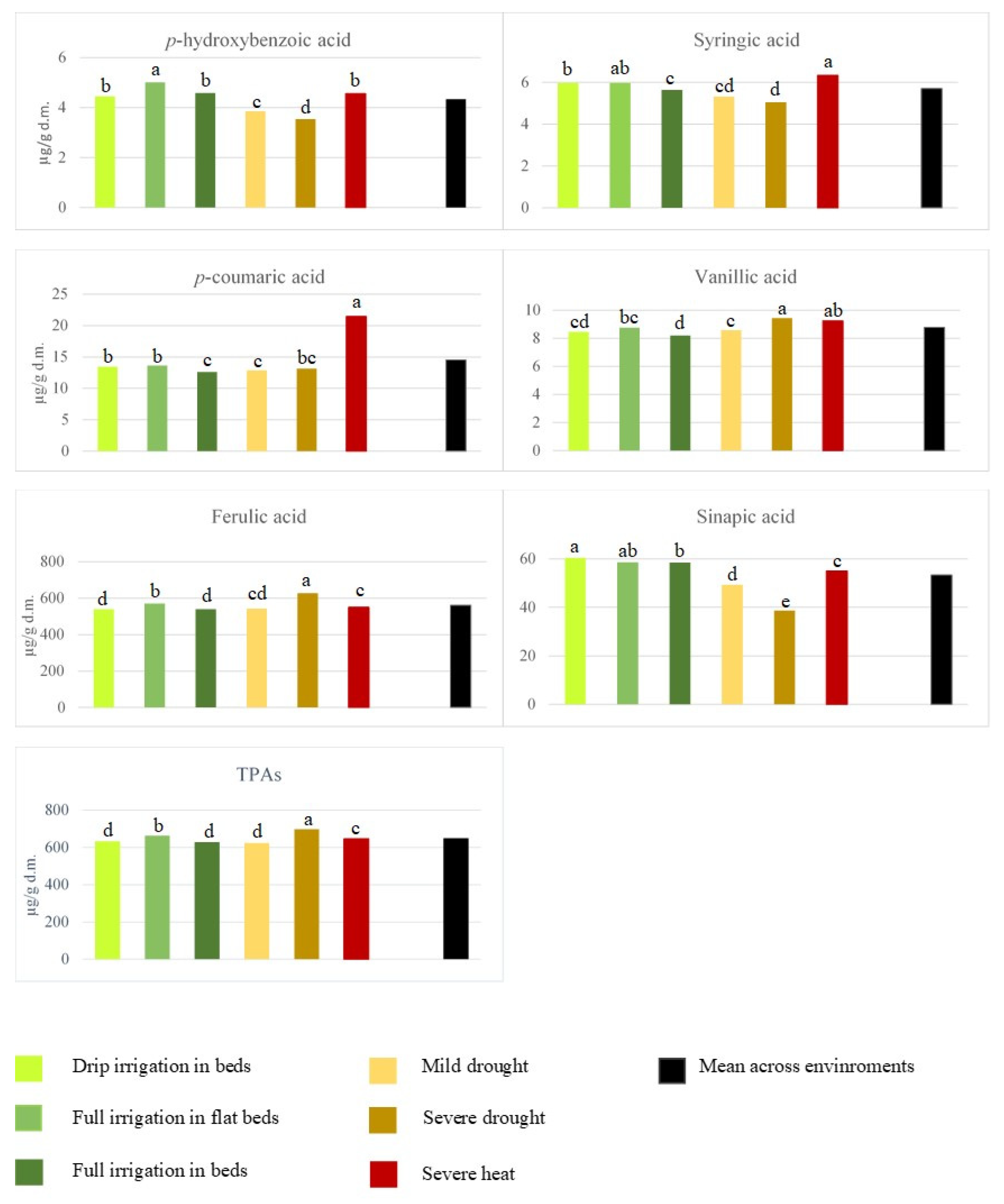

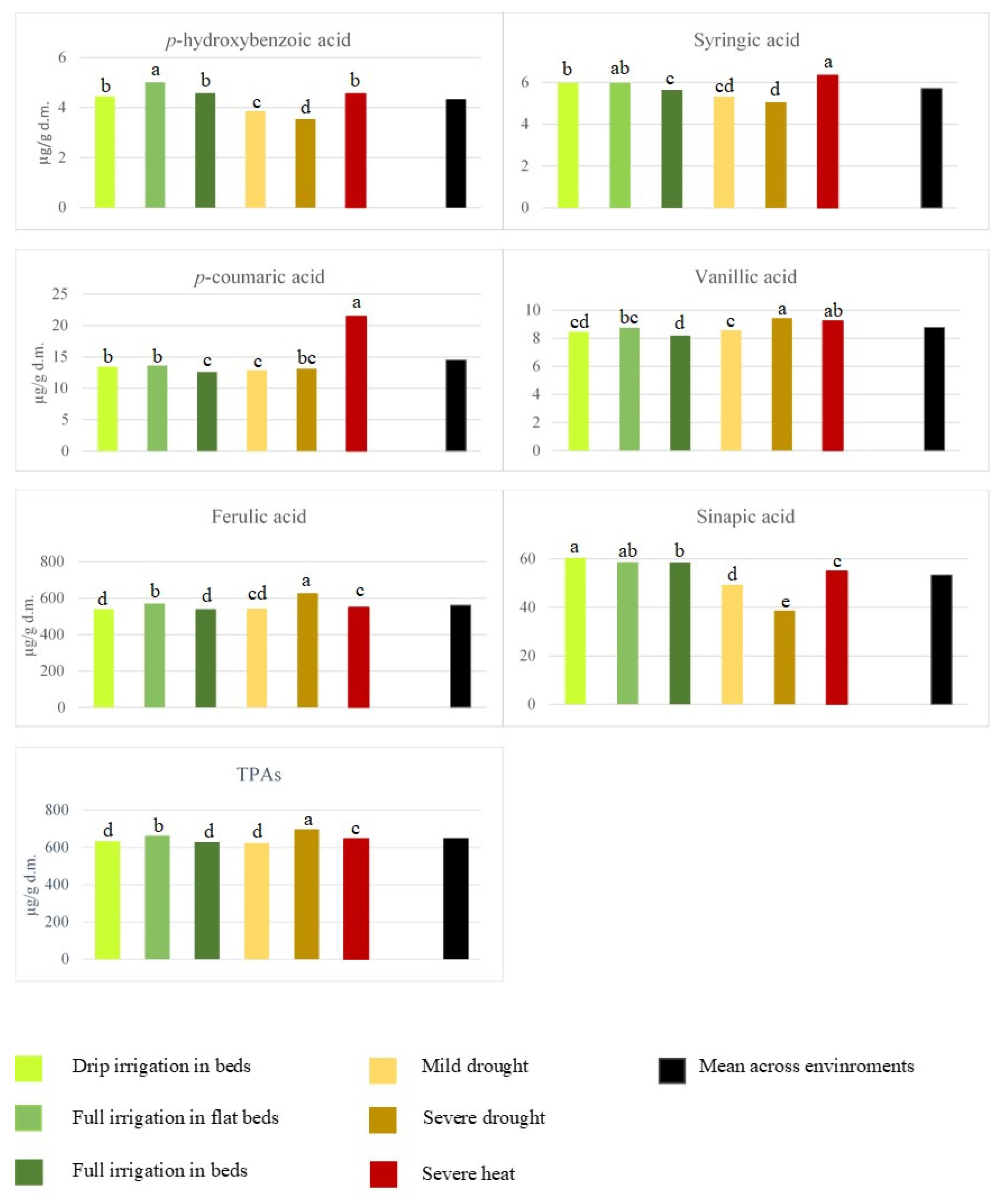{kind=link}
| p-Hydroxy-benzoic Acid | Syringic Acid | Vanillic Acid | p-Coumaric Acid | Ferulic Acid | Sinapic Acid | TPAs | |
|---|---|---|---|---|---|---|---|
| Altar | 4.87 b | 5.37 d | 8.15 c | 14.24 bc | 540.96 c | 51.58 c | 625.18 cd |
| Atil | 4.80 b | 5.16 d | 8.14 c | 12.94 d | 563.69 b | 55.04 b | 649.75 b |
| Cirno | 5.12 a | 5.81 bc | 8.16 c | 13.85 c | 611.00 a | 59.13 a | 703.09 a |
| Jupare | 3.85 c | 6.10 ab | 9.30 b | 14.84 b | 530.09 c | 54.62 b | 618.80 d |
| Mexicali | 3.30 d | 6.14 a | 9.83 a | 14.09 b | 568.34 b | 54.74 b | 657.29 b |
| Yavaros | 3.99 c | 5.73 c | 9.06 b | 16.38 a | 559.15 b | 45.21 d | 639.52 bc |
| Grand Mean | 4.34 | 5.72 | 8.79 | 14.52 | 563.07 | 53.2 | 649.87 |
| Range | 1.9–6.8 | 2.4–8.5 | 5.4–12.8 | 7.4–38.3 | 390.1–785.6 | 29.4–92.3 | 444.9–902.2 |
| CV | 6.9 | 9.87 | 7.48 | 8.3 | 4.92 | 5.16 | 4.86 |
| Heritability | 0.95 | 0.75 | 0.89 | 0.23 | 0.69 | 0.85 | 0.65 |
| LSD | 0.17 | 0.33 | 0.38 | 0.74 | 17.1 | 2.21 | 19.51 |
| Source | DF | p-Hydroxy Benzoic Acid | Syringic Acid | Vanillic Acid | p-Coumaric Acid | Ferulic Acid | Sinapic Acid | TPAs 1 |
|---|---|---|---|---|---|---|---|---|
| Year (Y) | 1 | 0.61 * | 59.82 *** | 83.31 *** | 0.21 *** | 220,684.87 *** | 6149.96 *** | 320,586.19 *** |
| Growth conditions (E) | 5 | 7.08 *** | 5.51 *** | 5.26 *** | 276.34 *** | 27,885.92 *** | 1650.95 *** | 18,322.21 *** |
| E x Y | 5 | 4.01 *** | 5.11 *** | 3.10 *** | 32.38 *** | 19,815.92 *** | 540.06 *** | 27,821.01 *** |
| Rep (E x Y) | 12 | 0.08 ns | 0.26 ns | 0.44 ns | 0.45 ns | 214.82 ns | 9.10 ns | 284.14 ns |
| Genotype (G) | 5 | 12.07 *** | 3.73 *** | 12.13 *** | 32.43 *** | 20507.17 *** | 545.30 *** | 24,087.22 *** |
| G × Y | 5 | 0.28 *** | 0.214 *** | 1.380 * | 4.117 * | 9046.34 *** | 109.01 *** | 11,060.94 *** |
| G × E | 25 | 0.41 * | 1.28 *** | 1.46 *** | 45.12 *** | 6831.27 *** | 82.69 *** | 9043.75 *** |
| G × E × Y | 25 | 0.95 *** | 0.74 ** | 1.26 *** | 9.16 *** | 5768.96 *** | 95.32 *** | 7721.88 *** |
| Error | 60 | 0.09 | 0.33 *** | 0.43 *** | 1.65 *** | 876.77 *** | 14.71 *** | 1141.52 *** |
| Grain Yield | 1000 K. Weight | Test Weight | Protein Content | Grain Length | Grain Width | Grain Thickness | p-Hydroxy Benzoic Acid | Syringic Acid | Vanillic Acid | p-Coumaric Acid | Ferulic Acid | Sinapic Acid | TPAs | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grain yield | 1 | |||||||||||||
| 1000 K. weight | 0.78 | 1 | ||||||||||||
| Test weight | −0.16 | −0.60 | 1 | |||||||||||
| Protein content | NS | −0.13 | 0.43 | 1 | ||||||||||
| Grain length | −0.39 | −0.62 | 0.88 | 0.69 | 1 | |||||||||
| Grain width | NS | −0.32 | 0.64 | −0.38 | 0.24 | 1 | ||||||||
| Grain thickness | −0.60 | −0.91 | 0.64 | −0.17 | 0.48 | 0.66 | 1 | |||||||
| p-Hydroxybenzoic | 0.55 | NS | 0.24 | 0.46 | NS | NS | NS | 1 | ||||||
| Syringic | 0.17 | 0.39 | NS | −0.62 | −0.25 | 0.47 | NS | −0.64 | 1 | |||||
| Vanillic | −0.42 | NS | −0.18 | −0.51 | NS | 0.12 | NS | −0.98 | 0.78 | 1 | ||||
| p-Coumaric | −0.49 | −0.36 | NS | −0.84 | −0.23 | 0.53 | 0.59 | −0.64 | 0.54 | 0.65 | 1 | |||
| Ferulic | 0.32 | −0.26 | 0.87 | 0.47 | 0.64 | 0.59 | 0.35 | 0.62 | −0.15 | −0.52 | −0.34 | 1 | ||
| Sinapic | 0.77 | 0.65 | 0.13 | 0.55 | 0.13 | −0.13 | −0.66 | 0.43 | NS | −0.33 | −0.79 | 0.46 | 1 | |
| TPAs | 0.41 | −0.14 | 0.83 | 0.49 | 0.61 | 0.56 | 0.24 | 0.61 | NS | −0.50 | −0.40 | 0.99 | 0.56 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laddomada, B.; Blanco, A.; Mita, G.; D’Amico, L.; Singh, R.P.; Ammar, K.; Crossa, J.; Guzmán, C. Drought and Heat Stress Impacts on Phenolic Acids Accumulation in Durum Wheat Cultivars. Foods 2021, 10, 2142. https://doi.org/10.3390/foods10092142
Laddomada B, Blanco A, Mita G, D’Amico L, Singh RP, Ammar K, Crossa J, Guzmán C. Drought and Heat Stress Impacts on Phenolic Acids Accumulation in Durum Wheat Cultivars. Foods. 2021; 10(9):2142. https://doi.org/10.3390/foods10092142
Chicago/Turabian StyleLaddomada, Barbara, Antonio Blanco, Giovanni Mita, Leone D’Amico, Ravi P. Singh, Karim Ammar, Jose Crossa, and Carlos Guzmán. 2021. "Drought and Heat Stress Impacts on Phenolic Acids Accumulation in Durum Wheat Cultivars" Foods 10, no. 9: 2142. https://doi.org/10.3390/foods10092142
APA StyleLaddomada, B., Blanco, A., Mita, G., D’Amico, L., Singh, R. P., Ammar, K., Crossa, J., & Guzmán, C. (2021). Drought and Heat Stress Impacts on Phenolic Acids Accumulation in Durum Wheat Cultivars. Foods, 10(9), 2142. https://doi.org/10.3390/foods10092142