
The 10,000-Year Success Story of Wheat!





Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
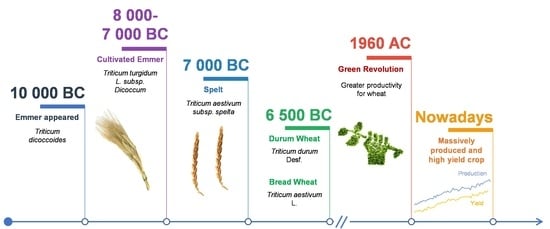
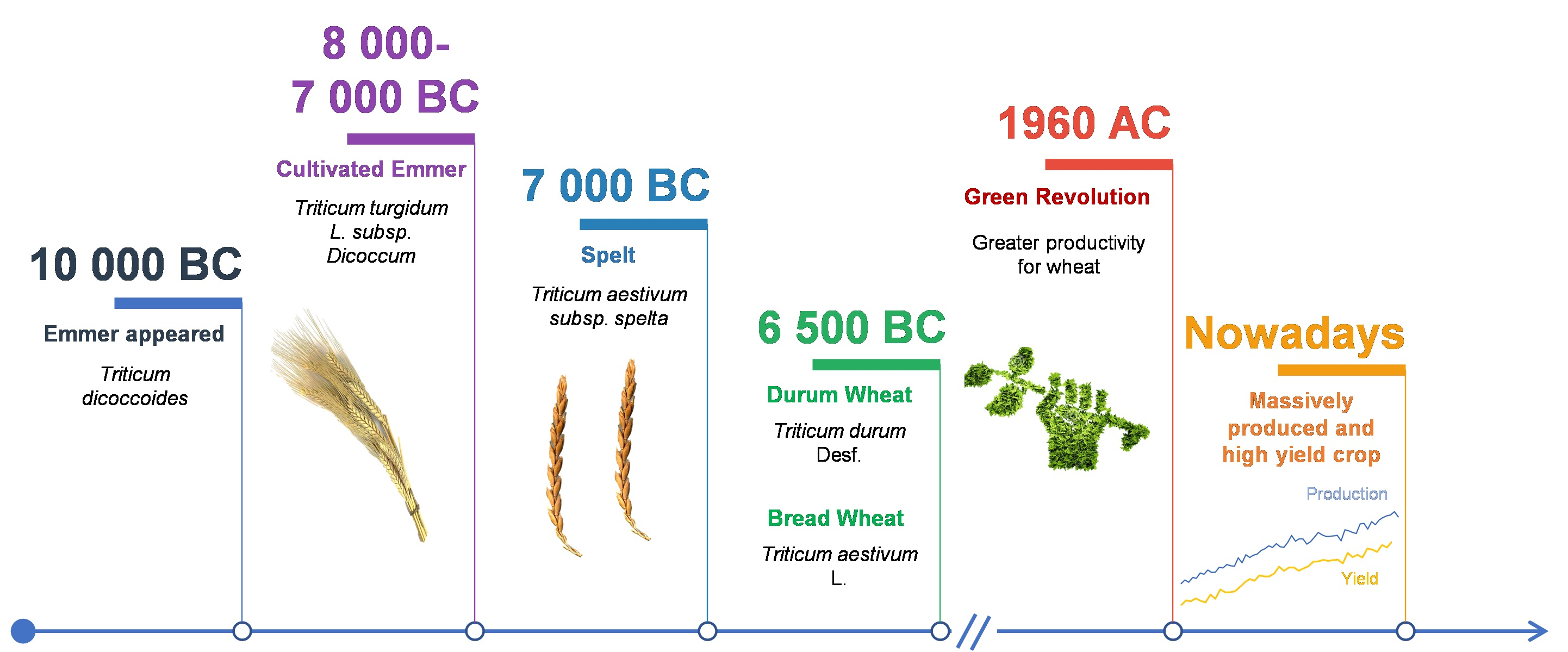
2. Origin and Evolution
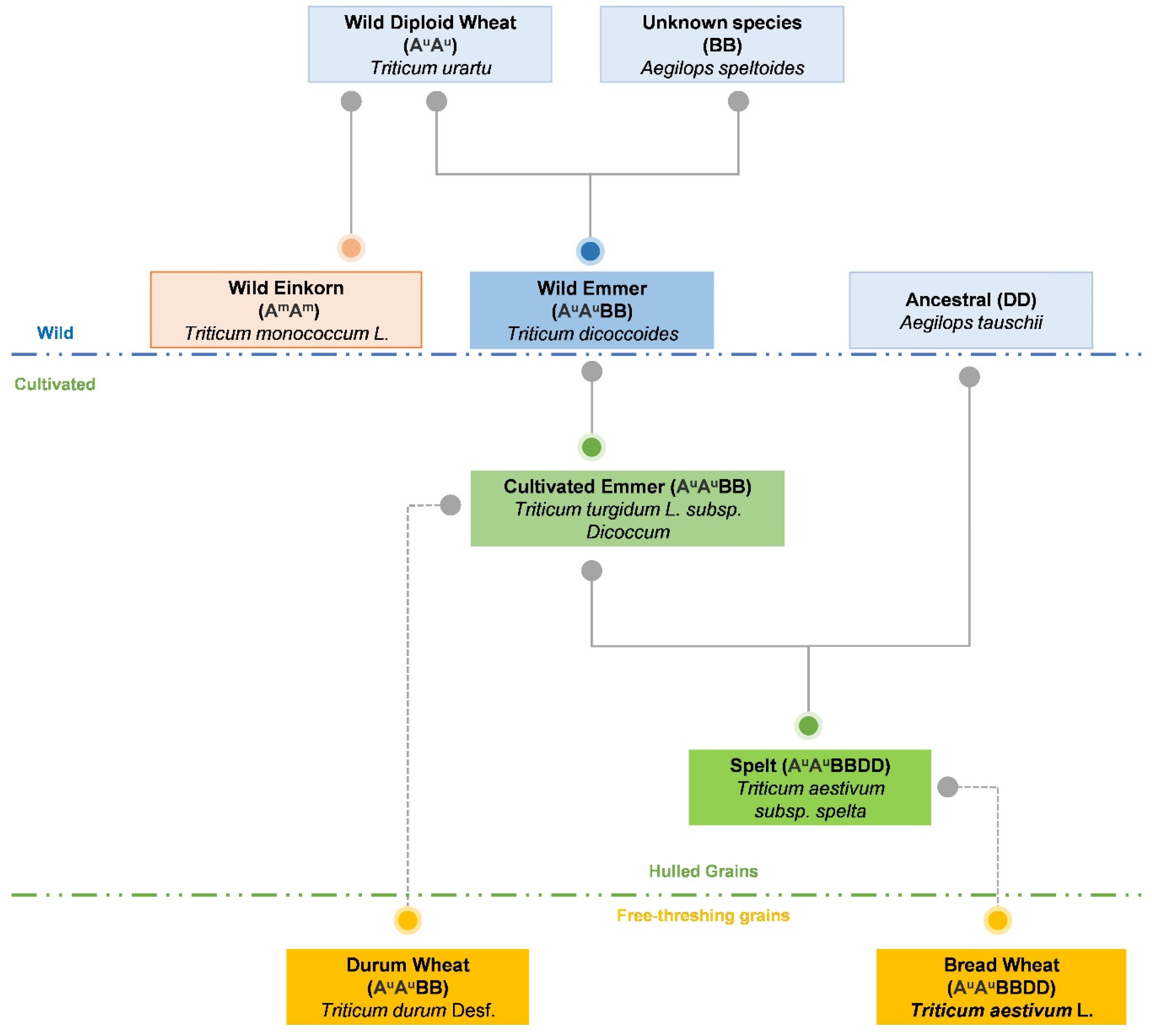
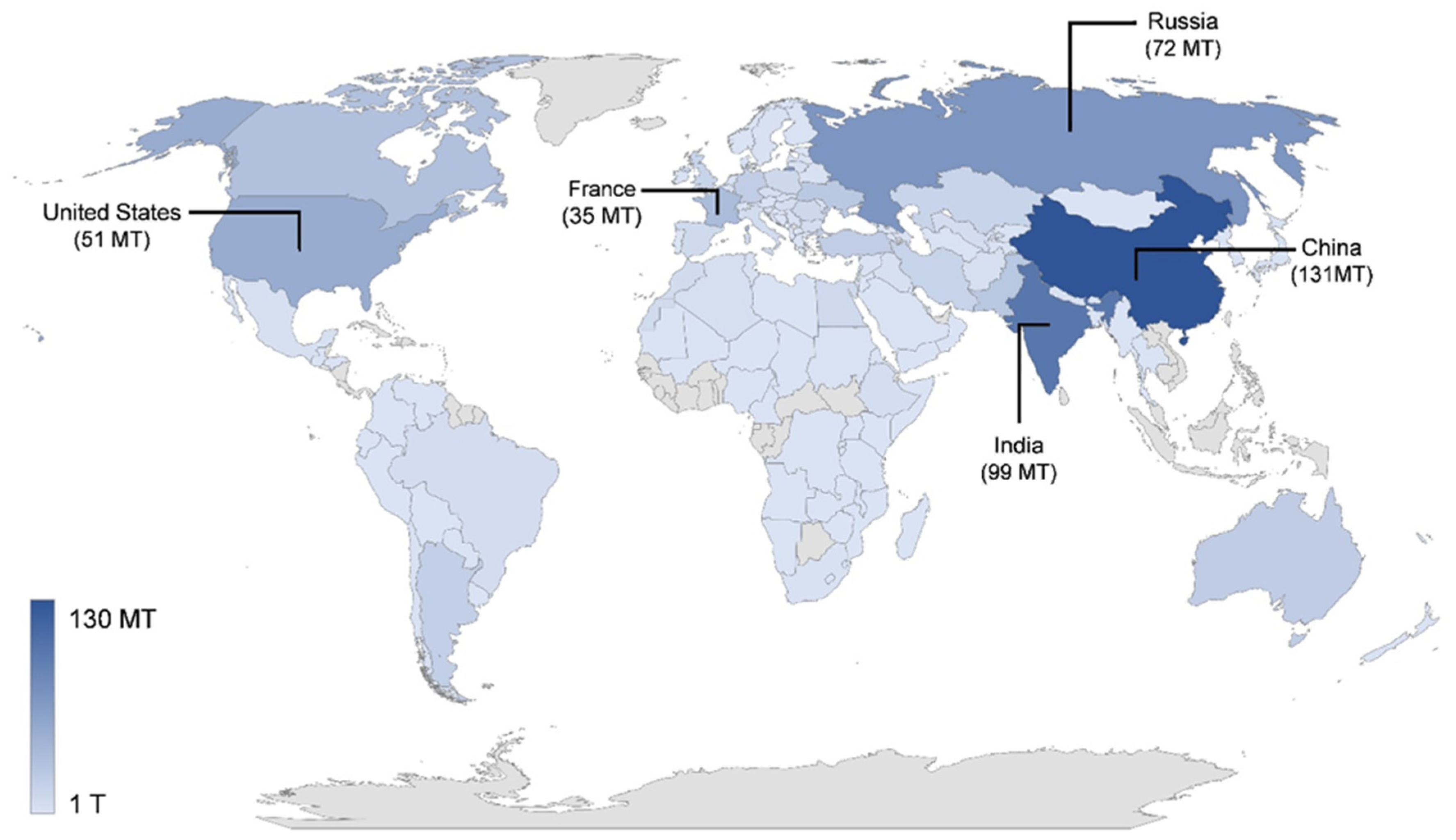
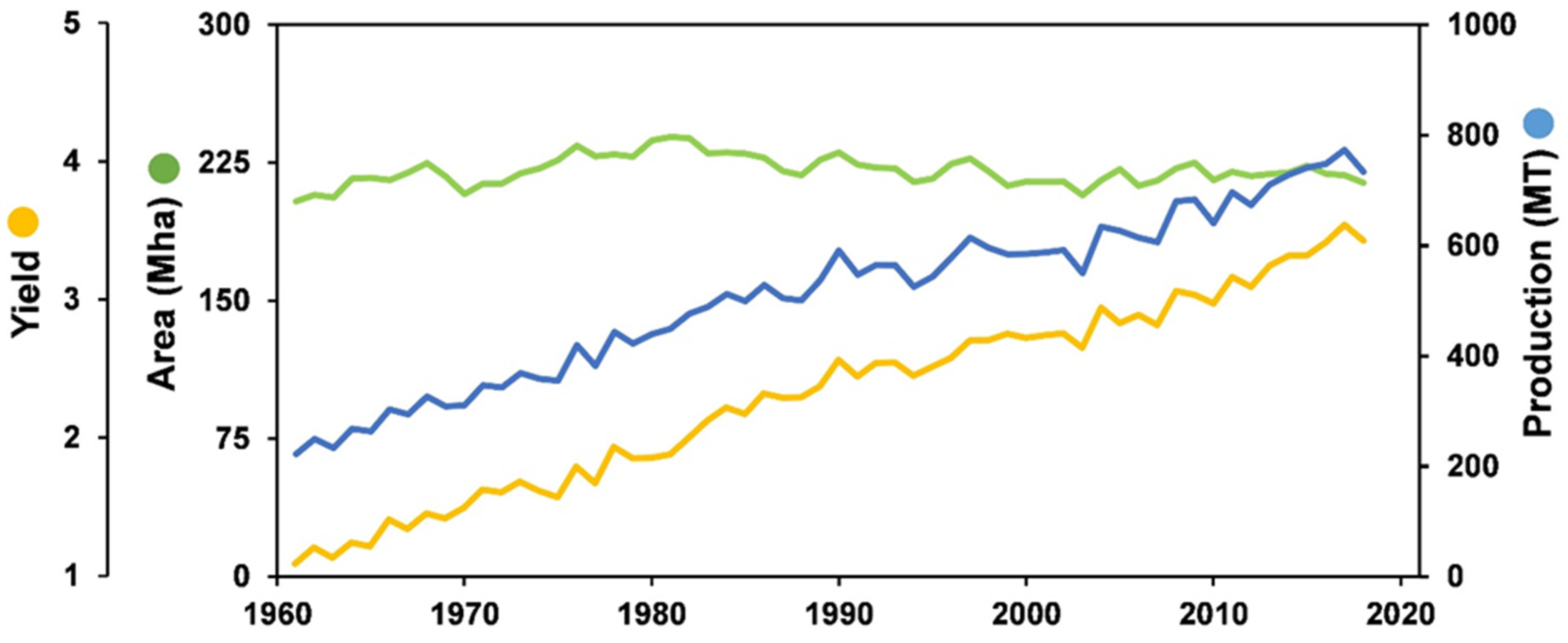
3. Production
4. Wheat Consumption Habits
5. Nutritional Value and Health Impact
6. Functional and Technological Properties
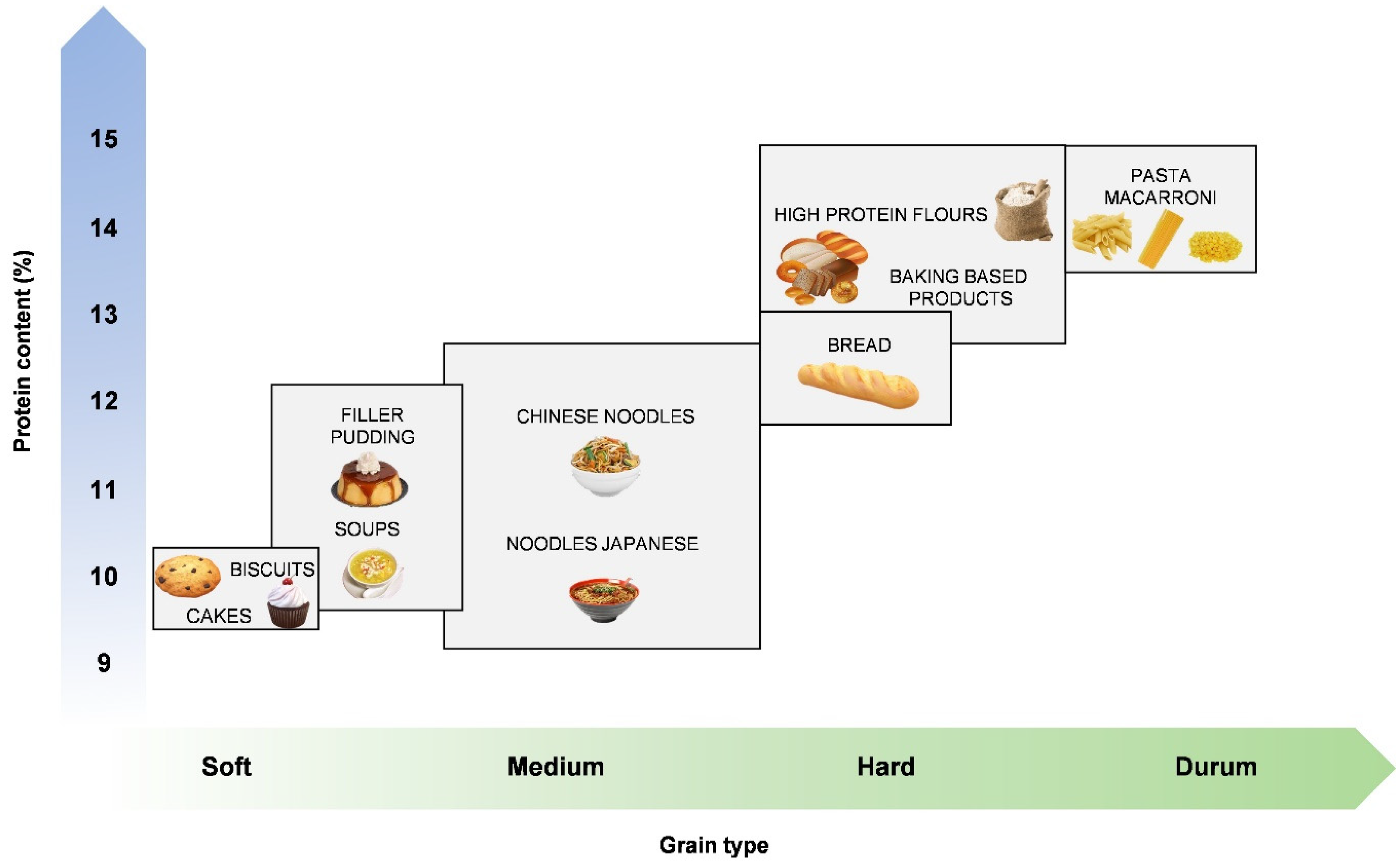
6.1. Grain Hardness
6.2. Wheat Proteins
6.2.1. Non-Gluten Proteins
6.2.2. Gluten
6.3. Effect of Gluten Proteins on Wheat Functionality
6.4. Models of Gluten Structure and Function in Dough
6.4.1. Pom-Pom Model
6.4.2. Loop-Train Model
6.4.3. Particle-Gel Model
6.4.4. Linear Glutenin Hypothesis
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization. Available online: http://www.fao.org/faostat/en/#home (accessed on 14 October 2020).
- Venske, E.; Dos Santos, R.S.; Busanello, C.; Gustafson, P.; Costa de Oliveira, A. Bread wheat: A role model for plant domestication and breeding. Hereditas 2019, 156, 16. [Google Scholar] [CrossRef] [PubMed]
- William, A.; Alain, B.; Maarten, V.G. The World Wheat Book: A History of Wheat Breeding; Lavoisier: Paris, France, 2011; Volume 2. [Google Scholar]
- Shewry, P.R. Do ancient types of wheat have health benefits compared with modern bread wheat? J. Cereal Sci. 2018, 79, 469–476. [Google Scholar] [CrossRef]
- Curtis, B. Wheat in the World. Bread Wheat: Improvement and Production; Food & Agriculture Organization of the UN: Rome, Italy, 2002. [Google Scholar]
- Dubcovsky, J.; Dvorak, J. Genome plasticity a key factor in the success of polyploid wheat under domestication. Science 2007, 316, 1862–1866. [Google Scholar] [CrossRef]
- Goncharov, N. Comparative-genetic analysis–a base for wheat taxonomy revision. Czech J. Genet. Plant Breed. 2005, 41, 52–55. [Google Scholar] [CrossRef]
- Peng, J.H.; Sun, D.; Nevo, E. Domestication evolution, genetics and genomics in wheat. Mol. Breed. 2011, 28, 281. [Google Scholar] [CrossRef]
- Dhaka, V.; Khatkar, B.J. Effects of gliadin/glutenin and HMW-GS/LMW-GS ratio on dough rheological properties and bread-making potential of wheat varieties. J. Food Qual. 2015, 38, 71–82. [Google Scholar] [CrossRef]
- Peña, R.; Trethowan, R.; Pfeiffer, W.; Ginkel, M.V. Quality (end-use) improvement in wheat: Compositional, genetic, and environmental factors. J. Crops Prod. 2002, 5, 1–37. [Google Scholar] [CrossRef]
- Peña, R. Wheat for bread and other foods. In Bread Wheat Improvement and Production; Food and Agriculture Organization of the United Nations: Rome, Italy, 2002; pp. 483–542. [Google Scholar]
- Callejo, M.J.; Vargas-Kostiuk, M.-E.; Ribeiro, M.; Rodríguez-Quijano, M. Triticum aestivum ssp. vulgare and ssp. spelta cultivars: 2. Bread-making optimisation. Eur. Food Res. Technol. 2019, 245, 1399–1408. [Google Scholar] [CrossRef]
- Rodríguez-Quijano, M.; Vargas-Kostiuk, M.-E.; Ribeiro, M.; Callejo, M.J. Triticum aestivum ssp. vulgare and ssp. spelta cultivars. 1. Functional evaluation. Eur. Food Res. Technol. 2019, 245, 1561–1570. [Google Scholar] [CrossRef]
- Arzani, A.; Ashraf, M. Cultivated ancient wheats (Triticum spp.): A potential source of health-beneficial food products. Compr. Rev. Food Sci. Food Saf. 2017, 16, 477–488. [Google Scholar] [CrossRef]
- Harlan, J.R.; Zohary, D. Distribution of wild wheats and barley. Science 1966, 153, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- Braun, H.-J.; Atlin, G.; Payne, T. Multi-location testing as a tool to identify plant response to global climate change. Clim. Chang. Crop Prod. 2010, 1, 115–138. [Google Scholar]
- Hawkesford, M.J.; Araus, J.L.; Park, R.; Calderini, D.; Miralles, D.; Shen, T.; Zhang, J.; Parry, M.A. Prospects of doubling global wheat yields. Food Energy Secur. 2013, 2, 34–48. [Google Scholar] [CrossRef]
- Sakamoto, S. Patterns of phylogenetic differentiation in the tribe Triticeae. Seiken Ziho 1973, 24, 11–31. [Google Scholar]
- Consortium, I.W. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar]
- Petersen, G.; Seberg, O.; Yde, M.; Berthelsen, K. Phylogenetic relationships of Triticum and Aegilops and evidence for the origin of the A, B, and D genomes of common wheat (Triticum aestivum). Mol. Phylogenet. Evol. 2006, 39, 70–82. [Google Scholar] [CrossRef]
- Feuillet, C.; Langridge, P.; Waugh, R. Cereal breeding takes a walk on the wild side. Trends Genet. 2008, 24, 24–32. [Google Scholar] [CrossRef]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin; Oxford University Press on Demand: Oxford, UK, 2012. [Google Scholar]
- Feldman, M.; Sears, E.R. The wild gene resources of wheat. Sci. Am. 1981, 244, 102–113. [Google Scholar] [CrossRef]
- Ling, H.-Q.; Zhao, S.; Liu, D.; Wang, J.; Sun, H.; Zhang, C.; Fan, H.; Li, D.; Dong, L.; Tao, Y. Draft genome of the wheat A-genome progenitor Triticum urartu. Nature 2013, 496, 87–90. [Google Scholar] [CrossRef]
- Gustafson, P.; Raskina, O.; Ma, X.; Nevo, E. Wheat evolution, domestication, and improvement. In Wheat: Science and Trade; Wiley: Danvers, MA, USA, 2009; pp. 5–30. [Google Scholar]
- Feldman, M.; Levy, A. Allopolyploidy—A shaping force in the evolution of wheat genomes. Cytogenet. Genome Res. 2005, 109, 250–258. [Google Scholar] [CrossRef]
- Chantret, N.; Salse, J.; Sabot, F.; Rahman, S.; Bellec, A.; Laubin, B.; Dubois, I.; Dossat, C.; Sourdille, P.; Joudrier, P.; et al. Molecular basis of evolutionary events that shaped the hardness locus in diploid and polyploid wheat species (Triticum and Aegilops). Plant Cell 2005, 17, 1033–1045. [Google Scholar] [CrossRef]
- Ikanović, J.; Popović, V.; Janković, S.; Živanović, L.; Rakić, S.; Dončić, D.J.G. Khorasan wheat population researching (triticum turgidum, ssp turanicum (mckey) in the minimum tillage conditions. Genetika 2014, 46, 105–115. [Google Scholar] [CrossRef]
- Brown, A.H. Variation under domestication in plants: 1859 and today. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2523–2530. [Google Scholar] [CrossRef][Green Version]
- Levy, A.A.; Feldman, M. The impact of polyploidy on grass genome evolution. Plant Physiol. 2002, 130, 1587–1593. [Google Scholar] [CrossRef]
- Sabot, F.; Guyot, R.; Wicker, T.; Chantret, N.; Laubin, B.; Chalhoub, B.; Leroy, P.; Sourdille, P.; Bernard, M. Updating of transposable element annotations from large wheat genomic sequences reveals diverse activities and gene associations. Mol. Genet. Genom. 2005, 274, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Nalam, V.J.; Vales, M.I.; Watson, C.J.; Kianian, S.F.; Riera-Lizarazu, O. Map-based analysis of genes affecting the brittle rachis character in tetraploid wheat (Triticum turgidum L.). Theor. Appl. Genet. 2006, 112, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Siddique, K.; Belford, R.; Perry, M.; Tennant, D. Growth, development and light interception of old and modern wheat cultivars in a Mediterranean-type environment. Aust. J. Agric. Res. 1989, 40, 473–487. [Google Scholar]
- Poland, J.; Endelman, J.; Dawson, J.; Rutkoski, J.; Wu, S.; Manes, Y.; Dreisigacker, S.; Crossa, J.; Sánchez-Villeda, H.; Sorrells, M. Genomic selection in wheat breeding using genotyping-by-sequencing. Plant Genome 2012, 5, 103–113. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hey, S. Do “ancient” wheat species differ from modern bread wheat in their contents of bioactive components? J. Cereal Sci. 2015, 65, 236–243. [Google Scholar] [CrossRef]
- Evans, J.R.; Lawson, T. From green to gold: Agricultural revolution for food security. J. Exp. Bot. 2020, 71, 2211–2215. [Google Scholar] [CrossRef] [PubMed]
- Khush, G.S. Green revolution: The way forward. Nat. Rev. Genet. 2001, 2, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P. The genes of the Green Revolution. Trends Genet. 2003, 19, 5–9. [Google Scholar] [CrossRef]
- Zhang, C.; Gao, L.; Sun, J.; Jia, J.; Ren, Z. Haplotype variation of Green Revolution gene Rht-D1 during wheat domestication and improvement. J. Integr. Plant Biol. 2014, 56, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, S.; van Ginkel, M. Increasing the Yield Potential in Wheat: Breaking the Barriers; Cimmyt: Mex, Mexico, 1996. [Google Scholar]
- Börner, A.; Plaschke, J.; Korzun, V.; Worland, A. The relationships between the dwarfing genes of wheat and rye. Euphytica 1996, 89, 69–75. [Google Scholar] [CrossRef]
- Na, T.; Jiang, Y.; He, B.-R.; Hu, Y.-G. The effects of dwarfing genes (Rht-B1b, Rht-D1b, and Rht8) with different sensitivity to GA3 on the coleoptile length and plant height of wheat. Agric. Sci. China 2009, 8, 1028–1038. [Google Scholar]
- Hooley, R. Gibberellins: Perception, transduction and responses. Plant Mol. Biol. 1994, 26, 1529–1555. [Google Scholar] [CrossRef]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F. ‘Green revolution’genes encode mutant gibberellin response modulators. Nature 1999, 400, 256–261. [Google Scholar] [CrossRef]
- Allan, R. Agronomic comparisons between Rht1 and Rht2 semidwarf genes in winter wheat. Crop Sci. 1989, 29, 1103–1108. [Google Scholar] [CrossRef]
- Rebetzke, G.; Richards, R.; Fettell, N.; Long, M.; Condon, A.G.; Forrester, R.; Botwright, T. Genotypic increases in coleoptile length improves stand establishment, vigour and grain yield of deep-sown wheat. Field Crops Res. 2007, 100, 10–23. [Google Scholar] [CrossRef]
- Khan, K. Wheat: Chemistry and Technology; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Indexmundi. Available online: https://www.indexmundi.com/ (accessed on 23 July 2021).
- Cauvain, S.P. Breadmaking: Improving Quality; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Vansteelandt, J.; Delcour, J. Physical behavior of durum wheat starch (Triticum durum) during industrial pasta processing. J. Agric. Food Chem. 1998, 46, 2499–2503. [Google Scholar] [CrossRef]
- Kaup, S.; Walker, C. Couscous in North Africa; Cereal Foods World: St. Paul, MN, USA, 1986. [Google Scholar]
- Elias, E. Durum wheat products. In Durum Wheat Improvement in the Mediterranean Region: New Challenges, Serie A: Séminaires Méditerranéennes; CIHEAM: Paris, France, 1995; Volume 40, pp. 23–31. [Google Scholar]
- Pauly, A.; Pareyt, B.; Fierens, E.; Delcour, J.A. Wheat (Triticum aestivum L. and T. turgidum L. ssp. durum) kernel hardness: II. Implications for end-product quality and role of puroindolines therein. Compr. Rev. Food Sci. Food Saf. 2013, 12, 427–438. [Google Scholar] [CrossRef]
- Qarooni, J. Wheat characteristics for flat breads: Hard or soft, white or red? Cereal Foods World 1996, 41, 391–395. [Google Scholar]
- Nagao, S. Quality characteristics of soft wheats and their utilization in Japan. II. Evaluation of wheats from the United States, Australia, France and Japan. Cereal Chem. 1977, 54, 198–204. [Google Scholar]
- Lee, M.R.; Swanson, B.G.; Baik, B.K.J. Influence of amylose content on properties of wheat starch and breadmaking quality of starch and gluten blends. Cereal Chem. 2001, 78, 701–706. [Google Scholar] [CrossRef]
- Peng, Q.; Wu, R.-L.; Kong, Z.-Y.; Zhang, B.-Q.J. Effect of waxy wheat flour blends on the quality of fresh and stale bread. Agric. Sci. China 2009, 8, 401–409. [Google Scholar]
- Graybosch, R.A.J. Technology. Waxy wheats: Origin, properties, and prospects. Trends Food Sci. Technol. 1998, 9, 135–142. [Google Scholar] [CrossRef]
- Arzani, A. Emmer (Triticum turgidum spp. dicoccum) flour and breads. In Flour and Breads and Their Fortification in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2011; pp. 69–78. [Google Scholar]
- Hidalgo, A.; Brandolini, A.J. Agriculture. Nutritional properties of einkorn wheat (Triticum monococcum L.). J. Sci. Food Agric. 2014, 94, 601–612. [Google Scholar] [CrossRef]
- Konvalina, P.; Moudrý, J., Jr.; Moudrý, J.J. Quality parametres of emmer wheat landraces. J. Cent. Eur. Agric. 2008, 9, 539–545. [Google Scholar]
- Geisslitz, S.; Scherf, K.A. Rediscovering ancient wheats. J. Cereal Sci. 2020, 65, 236–243. [Google Scholar]
- Benincasa, P.; Galieni, A.; Manetta, A.C.; Pace, R.; Guiducci, M.; Pisante, M.; Stagnari, F.J. Phenolic compounds in grains, sprouts and wheatgrass of hulled and non-hulled wheat species. J. Sci. Food Agric. 2015, 95, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Boukid, F.; Folloni, S.; Sforza, S.; Vittadini, E.; Prandi, B. Current Trends in Ancient Grains-Based Foodstuffs: Insights into Nutritional Aspects and Technological Applications. Compr. Rev. Food Sci. Food Saf. 2018, 17, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Pasqualone, A.; Piergiovanni, A.; Caponio, F.; Paradiso, V.M.; Summo, C.; Simeone, R. Evaluation of the technological characteristics and bread-making quality of alternative wheat cereals in comparison with common and durum wheat. Food Sci. Technol. Int. 2011, 17, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Frakolaki, G.; Giannou, V.; Topakas, E.; Tzia, C. Chemical characterization and breadmaking potential of spelt versus wheat flour. J. Cereal Sci. 2018, 79, 50–56. [Google Scholar] [CrossRef]
- Gomez-Becerra, H.F.; Erdem, H.; Yazici, A.; Tutus, Y.; Torun, B.; Ozturk, L.; Cakmak, I. Grain concentrations of protein and mineral nutrients in a large collection of spelt wheat grown under different environments. J. Cereal Sci. 2010, 52, 342–349. [Google Scholar] [CrossRef]
- Koenig, A.; Konitzer, K.; Wieser, H.; Koehler, P. Classification of spelt cultivars based on differences in storage protein compositions from wheat. Food Chem. 2015, 168, 176–182. [Google Scholar] [CrossRef]
- Sabença, C.; Ribeiro, M.; Sousa, T.d.; Poeta, P.; Bagulho, A.S.; Igrejas, G.J.F. Wheat/Gluten-Related Disorders and Gluten-Free Diet Misconceptions: A Review. Foods 2021, 10, 1765. [Google Scholar] [CrossRef]
- Bakery Products Market—Growth, Trends, COVID-19 Impact, and Forecasts (2021–2026); Mordor Intelligence: Telangana, India, 2021.
- Di Cagno, R.; Barbato, M.; Di Camillo, C.; Rizzello, C.G.; De Angelis, M.; Giuliani, G.; De Vincenzi, M.; Gobbetti, M.; Cucchiara, S.J. Gluten-free sourdough wheat baked goods appear safe for young celiac patients: A pilot study. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 777–783. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; Auricchio, S.; Greco, L.; Clarke, C.; De Vincenzi, M.; Giovannini, C.; D’Archivio, M.; Landolfo, F.; Parrilli, G.J.A.; et al. Sourdough bread made from wheat and nontoxic flours and started with selected lactobacilli is tolerated in celiac sprue patients. Appl. Environ. Microbiol. 2004, 70, 1088–1096. [Google Scholar] [CrossRef]
- Laatikainen, R.; Koskenpato, J.; Hongisto, S.-M.; Loponen, J.; Poussa, T.; Huang, X.; Sontag-Strohm, T.; Salmenkari, H.; Korpela, R.J.N. Pilot study: Comparison of sourdough wheat bread and yeast-fermented wheat bread in individuals with wheat sensitivity and irritable bowel syndrome. Nutrition 2017, 9, 1215. [Google Scholar] [CrossRef]
- Meziani, S.; Jasniewski, J.; Gaiani, C.; Ioannou, I.; Muller, J.-M.; Ghoul, M.; Desobry, S.J. Effects of freezing treatments on viscoelastic and structural behavior of frozen sweet dough. J. Food Eng. 2011, 107, 358–365. [Google Scholar] [CrossRef]
- Wang, P.; Xu, L.; Nikoo, M.; Ocen, D.; Wu, F.; Yang, N.; Jin, Z.; Xu, X.J. Effect of frozen storage on the conformational, thermal and microscopic properties of gluten: Comparative studies on gluten-, glutenin-and gliadin-rich fractions. Food Hydrocoll. 2014, 35, 238–246. [Google Scholar] [CrossRef]
- Wang, P.; Chen, H.; Mohanad, B.; Xu, L.; Ning, Y.; Xu, J.; Wu, F.; Yang, N.; Jin, Z.; Xu, X. Effect of frozen storage on physico-chemistry of wheat gluten proteins: Studies on gluten-, glutenin-and gliadin-rich fractions. Food Hydrocoll. 2014, 39, 187–194. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, X.; Hu, Z.; Li, L.; Li, B. Molecular structure evaluation of wheat gluten during frozen storage. Food Biophys. 2017, 12, 60–68. [Google Scholar] [CrossRef]
- Phimolsiripol, Y.; Siripatrawan, U.; Tulyathan, V.; Cleland, D.J. Effects of freezing and temperature fluctuations during frozen storage on frozen dough and bread quality. J. Food Eng. 2008, 84, 48–56. [Google Scholar] [CrossRef]
- Loveday, S.M.; Huang, V.T.; Reid, D.S.; Winger, R.J. Water dynamics in fresh and frozen yeasted dough. Crit. Rev. Food Sci. Nutr. 2012, 52, 390–409. [Google Scholar] [CrossRef]
- Kahleova, H.; Dort, S.; Holubkov, R.; Barnard, N.D. A plant-based high-carbohydrate, low-fat diet in overweight individuals in a 16-week randomized clinical trial: The role of carbohydrates. Nutrition 2018, 10, 1302. [Google Scholar] [CrossRef]
- Guragain, Y.N.; Ganesh, K.; Bansal, S.; Sathish, R.S.; Rao, N.; Vadlani, P.V. Low-lignin mutant biomass resources: Effect of compositional changes on ethanol yield. Ind. Crops Prod. 2014, 61, 1–8. [Google Scholar] [CrossRef]
- Khan, K.; Shewry, P.R. Wheat: Chemistry and Technology; AACC International: Eagan, MN, USA, 2009. [Google Scholar]
- Bushuk, W.; Rasper, V.F. Wheat: Production, Properties and Quality; Springer Science & Business Media: Berlin, Germany, 1994. [Google Scholar]
- Poutanen, K.J. Past and future of cereal grains as food for health. Trends Food Sci. Technol. 2012, 25, 58–62. [Google Scholar] [CrossRef]
- Escarnot, E.; Dornez, E.; Verspreet, J.; Agneessens, R.; Courtin, C.M. Quantification and visualization of dietary fibre components in spelt and wheat kernels. J. Cereal Sci. 2015, 62, 124–133. [Google Scholar] [CrossRef]
- Finnie, S.; Jeannotte, R.; Faubion, J.M. Quantitative characterization of polar lipids from wheat whole meal, flour, and starch. Cereal Chem. J. 2009, 86, 637–645. [Google Scholar] [CrossRef]
- Uthayakumaran, S.; Wrigley, C. Wheat: Grain-quality characteristics and management of quality requirements. In Cereal Grains; Elsevier: Amsterdam, The Netherlands, 2017; pp. 91–134. [Google Scholar]
- Righetti, L.; Rubert, J.; Galaverna, G.; Folloni, S.; Ranieri, R.; Stranska-Zachariasova, M.; Hajslova, J.; Dall’Asta, C. Characterization and Discrimination of Ancient Grains: A Metabolomics Approach. Int. J. Mol. Sci. 2016, 17, 1217. [Google Scholar] [CrossRef]
- Hidalgo, A.; Brandolini, A.; Pompei, C. Carotenoids evolution during pasta, bread and water biscuit preparation from wheat flours. Food Chem. 2010, 121, 746–751. [Google Scholar] [CrossRef]
- Hidalgo, A.; Brandolini, A.; Ratti, S.J. Influence of genetic and environmental factors on selected nutritional traits of Triticum monococcum. J. Agric. Food Chem. 2009, 57, 6342–6348. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Functional amino acids in growth, reproduction, and health. Adv. Nutr. 2010, 1, 31–37. [Google Scholar] [CrossRef]
- Ribeiro, M.; Sousa, T.d.; Sabença, C.; Poeta, P.; Bagulho, A.S.; Igrejas, G. Advances in quantification and analysis of the celiac-related immunogenic potential of gluten. Compr. Rev. Food Sci. Food Saf. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.; Korner, R.; Ren, O.K. The intes-final T cell response to a gliadin in adult celiac disease is fo-cused on a single deamidated glutamine targeted by tissue transglutaminase. JEM 2000, 191, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Molberg, Ø.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Sollid, L.M.; Khosla, C. Structural basis for gluten intolerance in celiac sprue. Science 2002, 297, 2275–2279. [Google Scholar] [CrossRef] [PubMed]
- Ronga, D.; Laviano, L.; Catellani, M.; Milc, J.; Prandi, B.; Boukid, F.; Sforza, S.; Dossena, A.; Graziano, S.; Gullì, M.; et al. Influence of environmental and genetic factors on content of toxic and immunogenic wheat gluten peptides. Eur. J. Agron. 2020, 118, 126091. [Google Scholar] [CrossRef]
- Ribeiro, M.; Nunes, F.M.; Guedes, S.; Domingues, P.; Silva, A.M.; Carrillo, J.M.; Rodriguez-Quijano, M.; Branlard, G.; Igrejas, G. Efficient chemo-enzymatic gluten detoxification: Reducing toxic epitopes for celiac patients improving functional properties. Sci. Rep. 2015, 5, 1–17. [Google Scholar] [CrossRef]
- Van Herpen, T.W.; Goryunova, S.V.; Van Der Schoot, J.; Mitreva, M.; Salentijn, E.; Vorst, O.; Schenk, M.F.; Van Veelen, P.A.; Koning, F.; Van Soest, L.J.; et al. Alpha-gliadin genes from the A, B, and D genomes of wheat contain different sets of celiac disease epitopes. BMC Genom. 2006, 7, 1–13. [Google Scholar] [CrossRef]
- Malalgoda, M.; Simsek, S. Celiac disease and cereal proteins. Food Hydrocoll. 2017, 68, 108–113. [Google Scholar] [CrossRef]
- Ribeiro, M.; Nunes, F.M. We might have got it wrong: Modern wheat is not more toxic for celiac patients. Food Chem. 2019, 278, 820–822. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.; Rodriguez-Quijano, M.; Nunes, F.M.; Carrillo, J.M.; Branlard, G.; Igrejas, G. New insights into wheat toxicity: Breeding did not seem to contribute to a prevalence of potential celiac disease’s immunostimulatory epitopes. Food Chem. 2016, 213, 8–18. [Google Scholar] [CrossRef]
- Prandi, B.; Tedeschi, T.; Folloni, S.; Galaverna, G.; Sforza, S. Peptides from gluten digestion: A comparison between old and modern wheat varieties. Food Res. Int. 2017, 91, 92–102. [Google Scholar] [CrossRef]
- Malalgoda, M.; Meinhardt, S.W.; Simsek, S. Detection and quantitation of immunogenic epitopes related to celiac disease in historical and modern hard red spring wheat cultivars. Food Chem. 2018, 264, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Spaenij–Dekking, L.; Kooy–Winkelaar, Y.; van Veelen, P.; Drijfhout, J.W.; Jonker, H.; van Soest, L.; Smulders, M.J.; Bosch, D.; Gilissen, L.J.; Koning, F. Natural variation in toxicity of wheat: Potential for selection of nontoxic varieties for celiac disease patients. Gastroenterology 2005, 129, 797–806. [Google Scholar] [CrossRef]
- Pasha, I.; Anjum, F.; Morris, C. Grain hardness: A major determinant of wheat quality. Food Sci. Technol. Int. 2010, 16, 511–522. [Google Scholar] [CrossRef]
- Gazza, L.; Nocente, F.; Ng, P.; Pogna, N. Genetic and biochemical analysis of common wheat cultivars lacking puroindoline a. Theor. Appl. Genet. 2005, 110, 470–478. [Google Scholar] [CrossRef]
- Ribeiro, M.; Rodríguez-Quijano, M.; Giraldo, P.; Pinto, L.; Vázquez, J.F.; Carrillo, J.M.; Igrejas, G. Effect of allelic variation at glutenin and puroindoline loci on bread-making quality: Favorable combinations occur in less toxic varieties of wheat for celiac patients. Eur. Food Res. Technol. 2017, 243, 743–752. [Google Scholar] [CrossRef]
- Greenblatt, G.; Bettge, A.; Morris, C. Relationship between Endosperm Texture and the Occurrence of Friabilin and Bound Polar Lipids on Wheat Starch; American Association of Cereal Chemists: St. Paul, MN, USA, 1995. [Google Scholar]
- Pomeranz, Y. Composition and functionality of wheat-flour components. Wheat Chem. Technol. 1971, 3, 385–674. [Google Scholar]
- Guerrieri, N.; Cavaletto, M. Cereals proteins. In Proteins in Food Processing; Elsevier: Amsterdam, The Netherlands, 2018; pp. 223–244. [Google Scholar]
- Gao, L.; Wang, A.; Li, X.; Dong, K.; Wang, K.; Appels, R.; Ma, W.; Yan, Y. Wheat quality related differential expressions of albumins and globulins revealed by two-dimensional difference gel electrophoresis (2-D DIGE). J. Proteom. 2009, 73, 279–296. [Google Scholar] [CrossRef]
- Boukid, F.; Prandi, B.; Faccini, A.; Sforza, S.J. A complete mass spectrometry (MS)-based peptidomic description of gluten peptides generated during in vitro gastrointestinal digestion of durum wheat: Implication for celiac disease. J. Am. Soc. Mass Spectrom. 2019, 30, 1481–1490. [Google Scholar] [CrossRef]
- Platzer, B.; Baker, K.; Vera, M.P.; Singer, K.; Panduro, M.; Lexmond, W.S.; Turner, D.; Vargas, S.O.; Kinet, J.P.; Maurer, D.; et al. Dendritic cell-bound IgE functions to restrain allergic inflammation at mucosal sites. Mucosal Immunol. 2015, 8, 516–532. [Google Scholar] [CrossRef] [PubMed]
- Veraverbeke, W.S.; Delcour, J.A. Wheat protein composition and properties of wheat glutenin in relation to breadmaking functionality. Crit. Rev. Food Sci. Nutr. 2002, 42, 179–208. [Google Scholar] [CrossRef] [PubMed]
- Tomić, J.; Torbica, A.; Popović, L.; Strelec, I.; Vaštag, Ž.; Pojić, M.; Rakita, S.J. Albumins characterization in relation to rheological properties and enzymatic activity of wheat flour dough. JAST 2015, 17, 805–816. [Google Scholar]
- Chen, X.; Cao, X.; Zhang, Y.; Islam, S.; Zhang, J.; Yang, R.; Liu, J.; Li, G.; Appels, R.; Keeble-Gagnere, G.; et al. Genetic characterization of cysteine-rich type-b avenin-like protein coding genes in common wheat. Sci. Rep. 2016, 6, 30692. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, Y.; Hu, X.; Islam, S.; She, M.; Peng, Y.; Yu, Z.; Wylie, S.; Juhasz, A.; Dowla, M.; Yang, R.; et al. New insights into the evolution of wheat avenin-like proteins in wild emmer wheat (Triticum dicoccoides). Proc. Natl. Acad. Sci. USA 2018, 115, 13312–13317. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, C.; Xiao, Y.; Crossa, J.; González-Santoyo, H.; Huerta, J.; Singh, R.; Dreisigacker, S. Sources of the highly expressed wheat bread making (wbm) gene in CIMMYT spring wheat germplasm and its effect on processing and bread-making quality. Euphytica 2016, 209, 689–692. [Google Scholar] [CrossRef]
- Beccari, J.B. De Bononiensi Scientiarum et Artium Atque Academia Commentarii; Part I; Nabu Press: Charleston, SC, USA, 1745; pp. 122–127. [Google Scholar]
- Osborne, T.B. The Vegetable Proteins; Longmans, Green and Company: London, UK, 1924. [Google Scholar]
- Shewry, P.R.; Tatham, A.S. The characteristics, structures and evolutionary relationships of prolamins. In Seed Proteins; Springer: Berlin/Heidelberg, Germany, 1999; pp. 11–33. [Google Scholar]
- Osborne, T.B. The Proteins of the Wheat Kernel; Carnegie Institution: Washington, DC, USA, 1907. [Google Scholar]
- Ribeiro, M.; Nunes-Miranda, J.D.; Branlard, G.; Carrillo, J.M.; Rodriguez-Quijano, M.; Igrejas, G. One hundred years of grain omics: Identifying the glutens that feed the world. J. Proteome Res. 2013, 12, 4702–4716. [Google Scholar] [CrossRef]
- Ribeiro, M.; Nunes, F.M.; Rodriguez-Quijano, M.; Carrillo, J.M.; Branlard, G.; Igrejas, G. Next-generation therapies for celiac disease: The gluten-targeted approaches. Trends Food Sci. Technol. 2018, 75, 56–71. [Google Scholar] [CrossRef]
- Tatham, A.S.; Shewry, P.R. The S-poor prolamins of wheat, barley and rye: Revisited. J. Cereal Sci. 2012, 55, 79–99. [Google Scholar] [CrossRef]
- Payne, P.; Holt, L.M.; Jackson, E.A. Wheat storage proteins: Their genetics and their potential for manipulation by plant breeding. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1984, 304, 359–371. [Google Scholar]
- Payne, P.I. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu. Rev. Plant Physiol. 1987, 38, 141–153. [Google Scholar] [CrossRef]
- Shewry, P.; Tatham, A. Disulphide bonds in wheat gluten proteins. J. Cereal Sci. 1997, 25, 207–227. [Google Scholar] [CrossRef]
- Branlard, G.; Dardevet, M.; Saccomano, R.; Lagoutte, F.; Gourdon, J. Genetic diversity of wheat storage proteins and bread wheat quality. Euphytica 2001, 119, 59–67. [Google Scholar] [CrossRef]
- Agarwal, M.; Shrivastava, N.; Padh, H. Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Rep. 2008, 27, 617–631. [Google Scholar] [CrossRef]
- Payne, P.I.; Lawrence, G.J. Catalogue of alleles for the complex gene loci, Glu-A1, Glu-B1, and Glu-D1 which code for high-molecular-weight subunits of glutenin in hexaploid wheat. Cereal Res. Commun. 1983, 11, 29–35. [Google Scholar]
- Payne, P.I.; Holt, L.M.; Lawrence, G.J.; Law, C.N. The genetics of gliadin and glutenin, the major storage proteins of the wheat endosperm. Plant Foods Hum. Nutr. 1982, 31, 229–241. [Google Scholar] [CrossRef]
- Jackson, E.A.; Holt, L.M.; Payne, P.I. Characterisation of high molecular weight gliadin and low-molecular-weight glutenin subunits of wheat endosperm by two-dimensional electrophoresis and the chromosomal localisation of their controlling genes. Theor. Appl. Genet. 1983, 66, 29–37. [Google Scholar] [CrossRef]
- Rasheed, A.; Xia, X.; Yan, Y.; Appels, R.; Mahmood, T.; He, Z. Wheat seed storage proteins: Advances in molecular genetics, diversity and breeding applications. J. Cereal Sci. 2014, 60, 11–24. [Google Scholar] [CrossRef]
- Ribeiro, M.; Sousa, T.; Poeta, P.; Bagulho, A.S.; Igrejas, G. Review of Structural Features and Binding Capacity of Polyphenols to Gluten Proteins and Peptides In Vitro: Relevance to Celiac Disease. Antioxidants 2020, 9, 463. [Google Scholar] [CrossRef] [PubMed]
- Biesiekierski, J.R. What is gluten? J. Gastroenterol. Hepatol. 2017, 32 (Suppl. 1), 78–81. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- Dupont, F.M.; Vensel, W.H.; Tanaka, C.K.; Hurkman, W.J.; Altenbach, S.B. Deciphering the complexities of the wheat flour proteome using quantitative two-dimensional electrophoresis, three proteases and tandem mass spectrometry. Proteome Sci. 2011, 9, 1–29. [Google Scholar] [CrossRef]
- Payne, P.I.; Corfield, K.G.; Holt, L.M.; Blackman, J.A. Correlations between the inheritance of certain high-molecular weight subunits of glutenin and bread-making quality in progenies of six crosses of bread wheat. J. Sci. Food Agric. 1981, 32, 51–60. [Google Scholar] [CrossRef]
- Payne, P.I.; Nightingale, M.A.; Krattiger, A.F.; Holt, L.M. The relationship between HMW glutenin subunit composition and the bread-making quality of British-grown wheat varieties. J. Sci. Food Agric. 1987, 40, 51–65. [Google Scholar] [CrossRef]
- Igrejas, G.; Guedes-Pinto, H.; Carnide, V.; Clement, J.; Branlard, G. Genetical, biochemical and technological parameters associated with biscuit quality. II. Prediction using storage proteins and quality characteristics in a soft wheat population. J. Cereal Sci. 2002, 36, 187–197. [Google Scholar] [CrossRef]
- Metakovsky, E.; Branlard, G.; Chernakov, V.; Upelniek, V.; Redaelli, R.; Pogna, N. Recombination mapping of some chromosome 1A-, 1B-, 1D-and 6B-controlled gliadins and low-molecular-weight glutenin subunits in common wheat. Theor. Appl. Genet. 1997, 94, 788–795. [Google Scholar] [CrossRef]
- Masci, S.; Rovelli, L.; Kasarda, D.; Vensel, W.; Lafiandra, D. Characterisation and chromosomal localisation of C-type low-molecular-weight glutenin subunits in the bread wheat cultivar Chinese Spring. Theor. Appl. Genet. 2002, 104, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhen, S.; Luo, N.; Han, C.; Lu, X.; Li, X.; Xia, X.; He, Z.; Yan, Y. Low molecular weight glutenin subunit gene Glu-B3h confers superior dough strength and breadmaking quality in wheat (Triticum aestivum L.). Sci. Rep. 2016, 6, 27182. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-Q.; Cloutier, S. Molecular characterization and genomic organization of low molecular weight glutenin subunit genes at the Glu-3 loci in hexaploid wheat (Triticum aestivum L.). Theor. Appl. Genet. 2008, 116, 953–966. [Google Scholar] [CrossRef] [PubMed]
- Wrigley, C.; Békés, F.; Bushuk, W. Gluten: A balance of gliadin and glutenin. In Gliadin and Glutenin: The Unique Balance of Wheat Quality; Amer Assn of Cereal Chemists: St. Paul, MN, USA, 2006; pp. 3–32. [Google Scholar]
- Geisslitz, S.; Longin, C.F.H.; Scherf, K.A.; Koehler, P. Comparative Study on Gluten Protein Composition of Ancient (Einkorn, Emmer and Spelt) and Modern Wheat Species (Durum and Common Wheat). Foods 2019, 8, 409. [Google Scholar] [CrossRef]
- Dobraszczyk, B.; Morgenstern, M. Rheology and the breadmaking process. J. Cereal Sci. 2003, 38, 229–245. [Google Scholar] [CrossRef]
- McLeish, T.; Larson, R. Molecular constitutive equations for a class of branched polymers: The pom-pom polymer. J. Rheol. 1998, 42, 81–110. [Google Scholar] [CrossRef]
- Belton, P. Mini review: On the elasticity of wheat gluten. J. Cereal Sci. 1999, 29, 103–107. [Google Scholar] [CrossRef]
- Don, C.; Lichtendonk, W.; Plijter, J.; Hamer, R. Glutenin macropolymer: A gel formed by glutenin particles. J. Cereal Sci. 2003, 37, 1–7. [Google Scholar] [CrossRef]
- Hamer, R.; Vliet, T.v. Understanding the structure and properties of gluten: An overview. In Proceedings of the 7th International Workshop Gluten 2000, Bristol, UK, 2–6 April 2000; pp. 125–131. [Google Scholar]
- Greenwood, C.; Ewart, J. Hypothesis for the structure of glutenin in relation to rheological properties of gluten and dough [Wheat]. Cereal Chem. 1975, 52, 146–153. [Google Scholar]
- Ewart, J. A hypothesis for the structure and rheology of glutenin. J. Sci. Food Agric. 1968, 19, 617–623. [Google Scholar] [CrossRef]
- Lindsay, M.P.; Skerritt, J.H. The glutenin macropolymer of wheat flour doughs: Structure–function perspectives. Trends Food Sci. Technol. 1999, 10, 247–253. [Google Scholar] [CrossRef]
- Ewart, J.A. Calculated molecular weight distribution for glutenin. J. Sci. Food Agric. 1987, 38, 277–289. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa, T.; Ribeiro, M.; Sabença, C.; Igrejas, G. The 10,000-Year Success Story of Wheat! Foods 2021, 10, 2124. https://doi.org/10.3390/foods10092124
de Sousa T, Ribeiro M, Sabença C, Igrejas G. The 10,000-Year Success Story of Wheat! Foods. 2021; 10(9):2124. https://doi.org/10.3390/foods10092124
Chicago/Turabian Stylede Sousa, Telma, Miguel Ribeiro, Carolina Sabença, and Gilberto Igrejas. 2021. "The 10,000-Year Success Story of Wheat!" Foods 10, no. 9: 2124. https://doi.org/10.3390/foods10092124
APA Stylede Sousa, T., Ribeiro, M., Sabença, C., & Igrejas, G. (2021). The 10,000-Year Success Story of Wheat! Foods, 10(9), 2124. https://doi.org/10.3390/foods10092124