
Survival of Escherichia coli and Listeria innocua on Lettuce after Irrigation with Contaminated Water in a Temperate Climate

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Generation of Marked Strains and Inoculum Preparation
2.2. Plant Growth Conditions
2.3. Experimental Plot Design
2.4. Inoculation of Lettuce Plants and Irrigation Water Microcosms
2.5. Sample Collection
2.6. Detection and Enumeration of Bacteria in Lettuce Plants via Culture
2.7. Detection and Enumeration of Bacteria in Irrigation Water via Culture
2.8. Detection and Enumeration of Bacteria in Lettuce Plants and Irrigation Water via qPCR
2.9. Live/Dead Staining
3. Results
3.1. Temperature Data
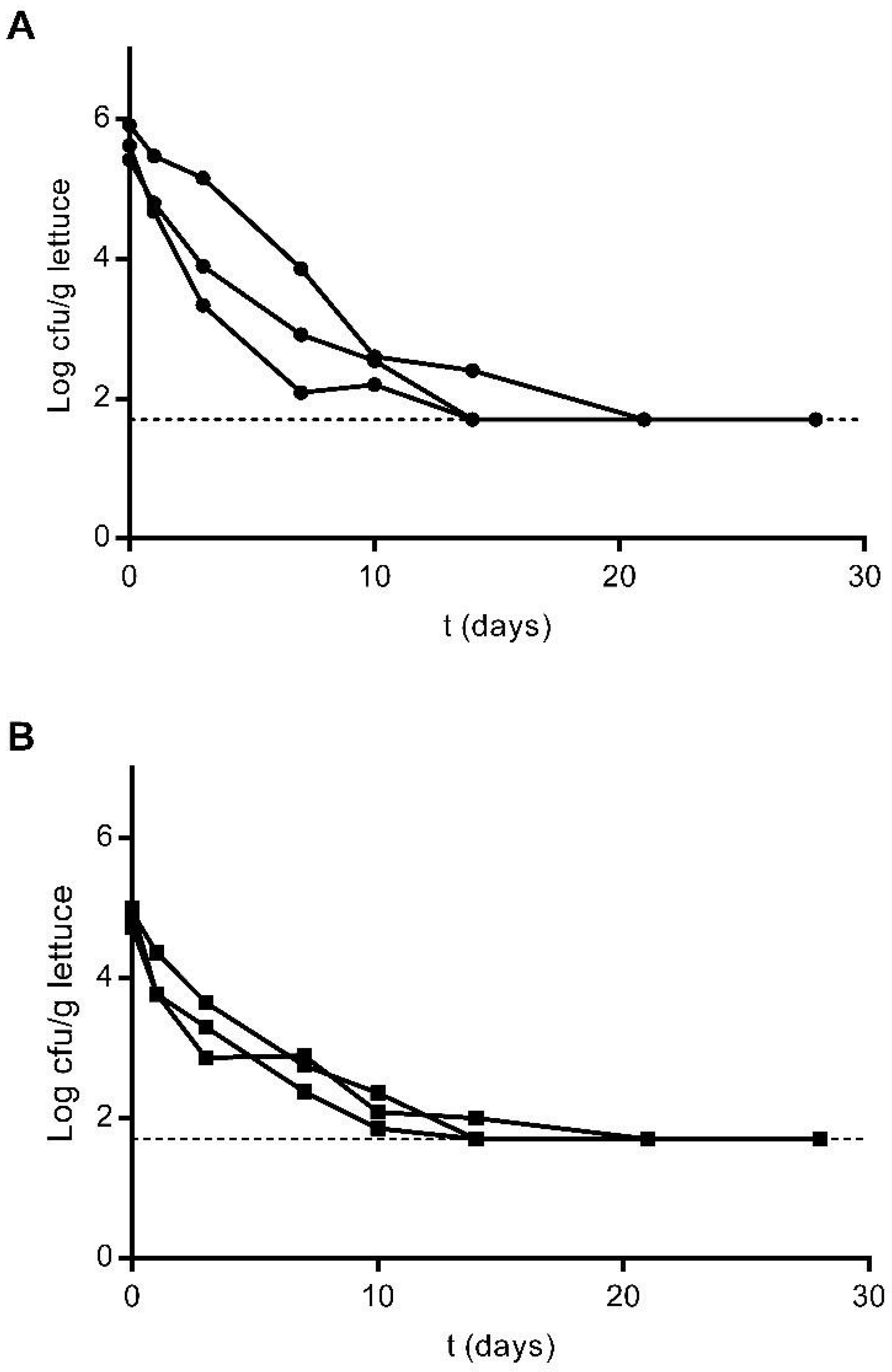
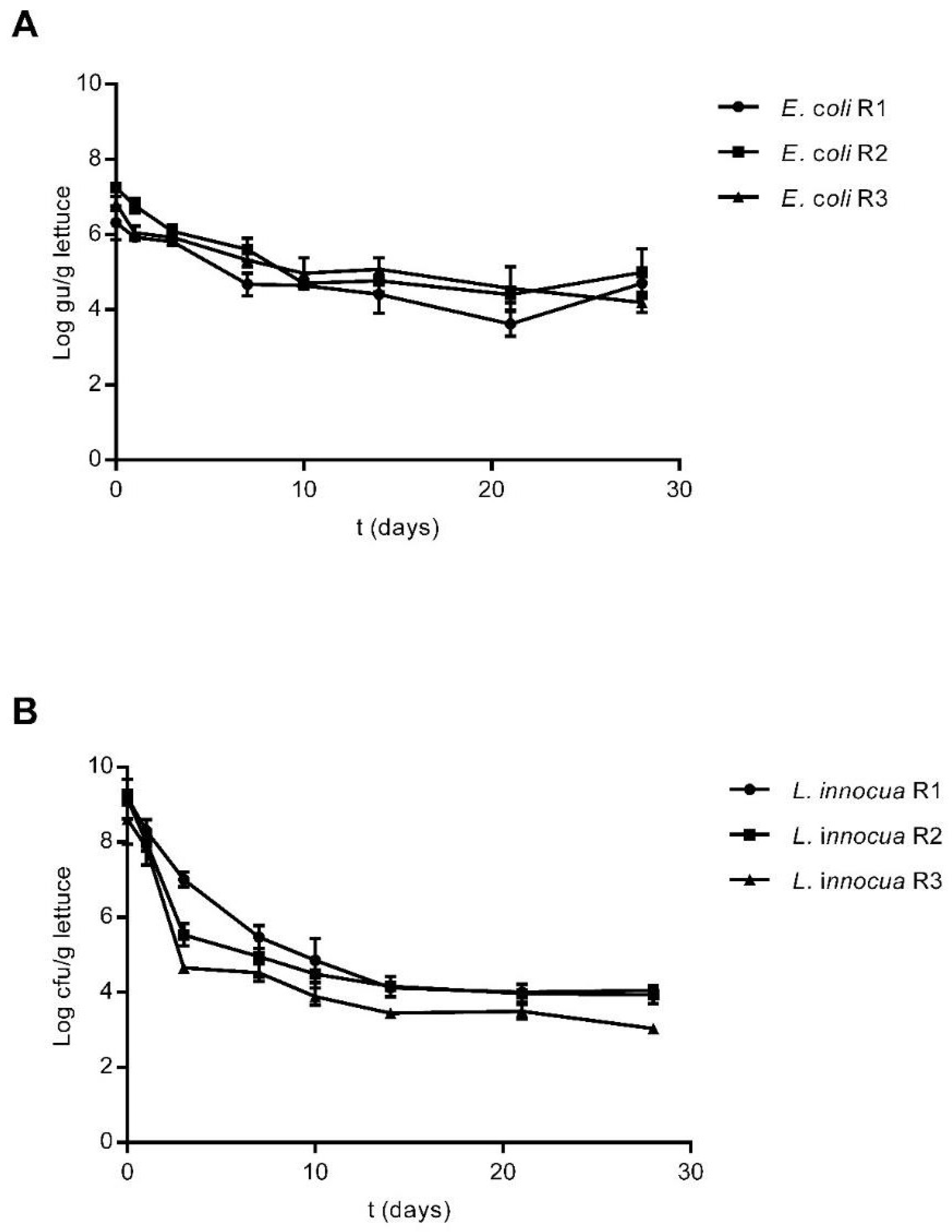
3.2. Survival of E. coli and L. innocua in Lettuce Plants
3.3. Survival of E. coli and L. innocua in Stored Irrigation Water
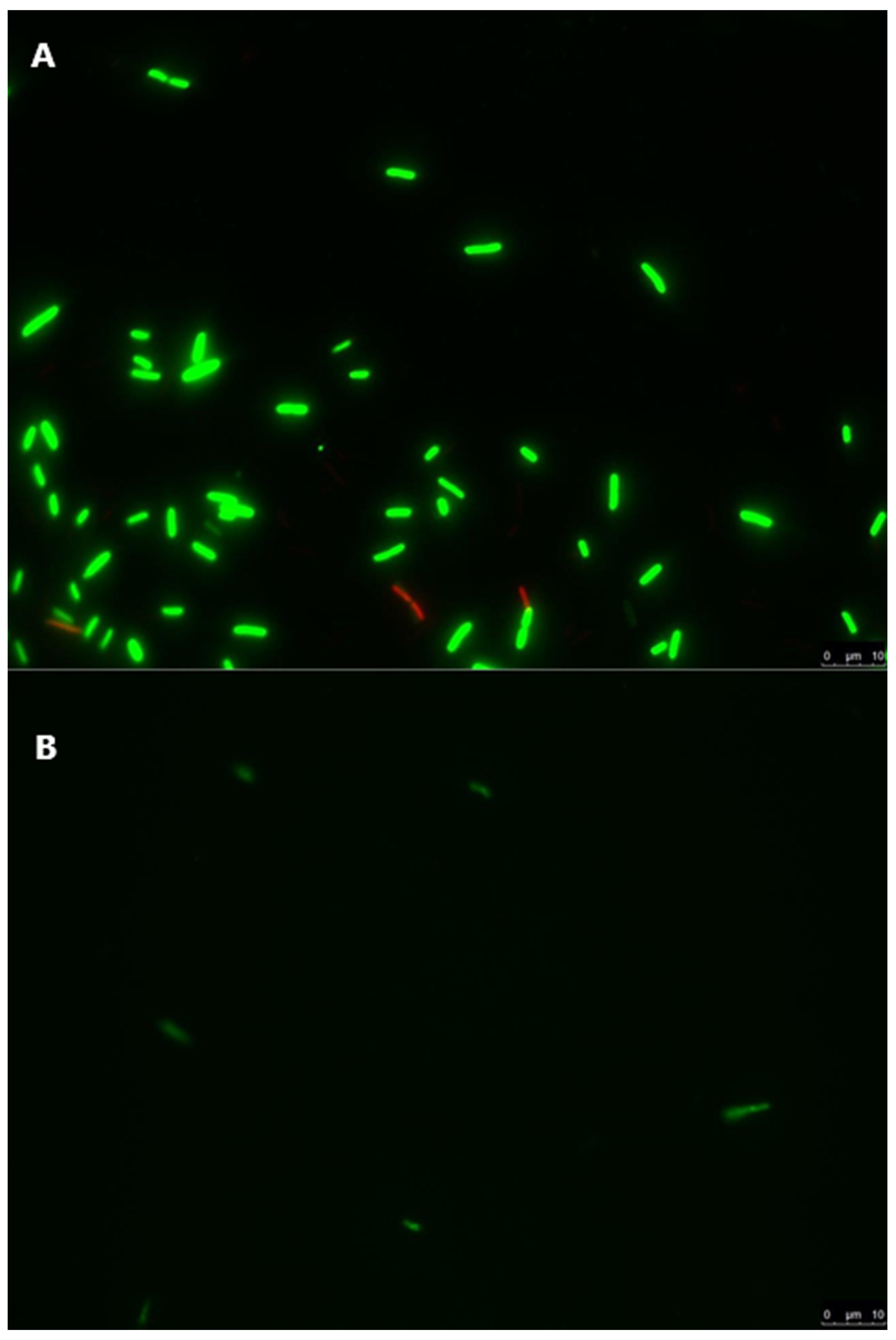
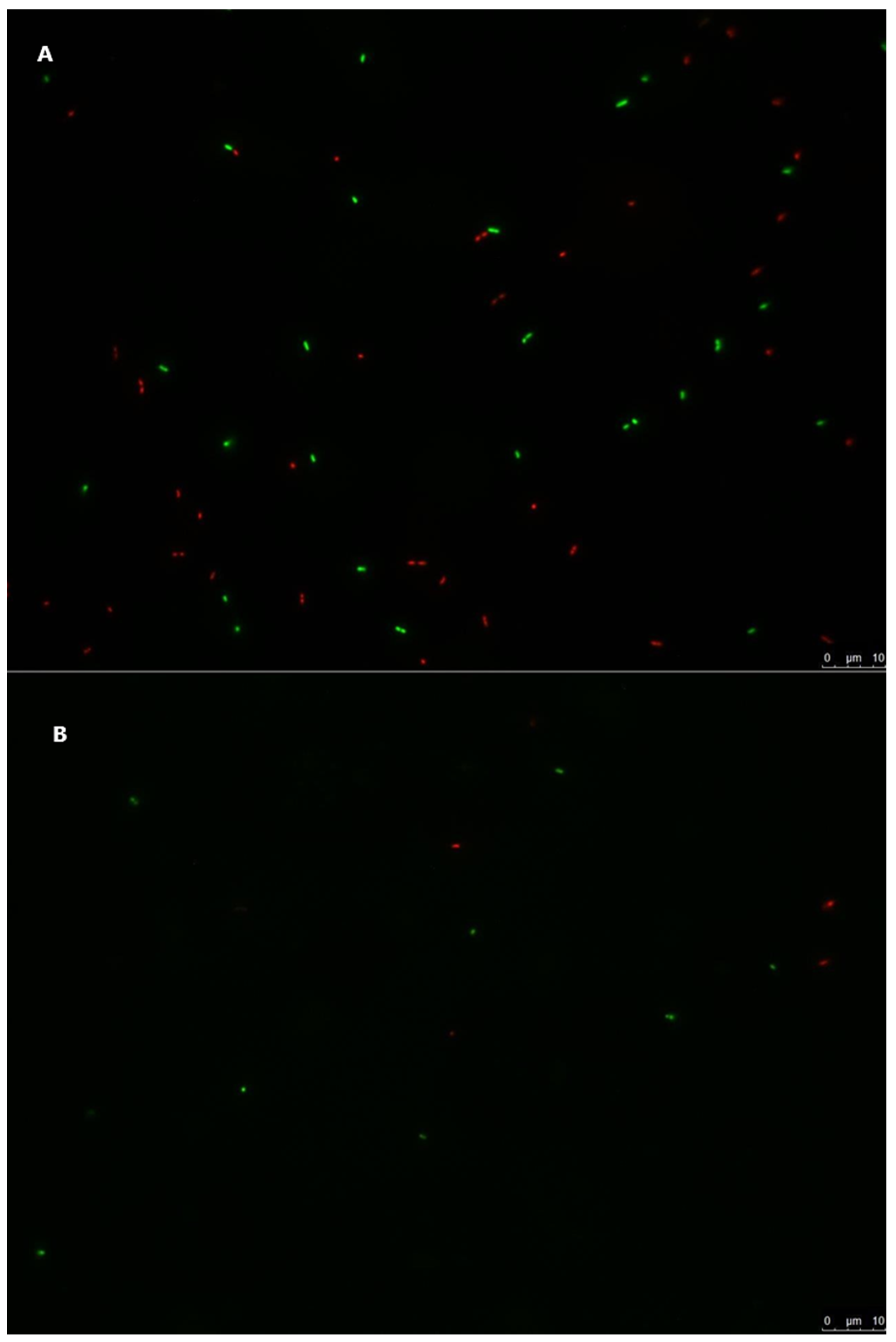
3.4. Live/Dead Staining
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Betts, R. Microbial update: Fruit & salad. Int. Food Hyg. 2014, 25, 9–12. [Google Scholar]
- Machado-Moreira, B.; Richards, K.; Brennan, F.; Abram, F.; Burgess, C.M. Microbial contamination of fresh produce: What, where, and how? Compr. Rev. Food Sci. Food Saf. 2019, 18, 1727–1750. [Google Scholar] [CrossRef] [PubMed]
- Hoelzer, K.; Pouillot, R.; Egan, K.; Dennis, S. Produce consumption in the United States: An analysis of consumption frequencies, serving sizes, processing forms, and high-consuming population subgroups for microbial risk assessments. J. Food Prot. 2012, 75, 328–340. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Holley, R.A. Factors influencing the microbial safety of fresh produce: A review. Food Microbiol. 2012, 32, 1–19. [Google Scholar]
- Abanyie, F.; Harvey, R.R.; Harris, J.R.; Wiegand, R.E.; Gaul, L.; Desvignes-Kendrick, M.; Irvin, K.; Williams, I.; Hall, R.L.; Herwaldt, B.; et al. 2013 multistate outbreaks of Cyclospora cayetanensis infections associated with fresh produce: Focus on the Texas investigations. Epidemiol. Infect. 2015, 143, 3451–3458. [Google Scholar] [CrossRef]
- Buss, B.F.; Joshi, M.V.; Dement, J.L.; Cantu, V.; Safranek, T.J. Multistate product traceforward investigation to link imported romaine lettuce to a US cyclosporiasis outbreak—Nebraska, Texas, and Florida, June-August 2013. Epidemiol. Infect. 2016, 144, 2709–2718. [Google Scholar] [CrossRef]
- Horby, P.W.; O’Brien, S.J.; Adak, G.K.; Graham, C.; Hawker, J.I.; Hunter, P.; Lane, C.; Lawson, A.J.; Mitchell, R.T.; Reacher, M.H.; et al. A national outbreak of multi-resistant Salmonella enterica serovar typhimurium definitive phage type (DT) 104 associated with consumption of lettuce. Epidemiol. Infect. 2003, 130, 169–178. [Google Scholar] [CrossRef]
- Wadamori, Y.; Gooneratne, R.; Hussain, M.A. Outbreaks and factors influencing microbiological contamination of fresh produce. J. Sci. Food Agric. 2017, 97, 1396–1403. [Google Scholar] [CrossRef]
- Mikhail, A.F.W.; Jenkins, C.; Dallman, T.J.; Inns, T.; Douglas, A.; Martin, A.I.C.; Fox, A.; Cleary, P.; Elson, R.; Hawker, J. An outbreak of Shiga toxin-producing Escherichia coli O157:H7 associated with contaminated salad leaves: Epidemiological, genomic and food trace back investigations. Epidemiol. Infect. 2018, 146, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Bottichio, L.; Keaton, A.; Thomas, D.; Fulton, T.; Tiffany, A.; Frick, A.; Mattioli, M.; Kahler, A.; Murphy, J.; Otto, M.; et al. Shiga toxin-producing Escherichia coli infections associated with romaine Lettuce-United States, 2018. Clin. Infect. Dis. 2020, 71, e323–e330. [Google Scholar] [CrossRef] [PubMed]
- Self, J.L.; Conrad, A.; Stroika, S.; Jackson, A.; Burnworth, L.; Beal, J.; Wellman, A.; Jackson, K.A.; Bidol, S.; Gerhardt, T.; et al. Notes from the field: Outbreak of listeriosis associated with consumption of packaged salad—United States and Canada, 2015–2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 879–881. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Gooneratne, R.; Hussain, M.A. Listeria monocytogenes in fresh produce: Outbreaks, prevalence and contamination levels. Foods 2017, 6, 21. [Google Scholar] [CrossRef]
- Bolton, D.J.; Byrne, C.M.; Sheridan, J.J.; McDowell, D.A.; Blair, I.S. The survival characteristics of a non-toxigenic strain of Escherichia coli O157:H7. J. Appl. Microbiol. 1999, 86, 407–411. [Google Scholar] [CrossRef]
- Franz, E.; van Diepeningen, A.D.; de Vos, O.J.; van Bruggen, A.H. Effects of cattle feeding regimen and soil management type on the fate of Escherichia coli O157:H7 and Salmonella enterica serovar typhimurium in manure, manure-amended soil, and lettuce. Appl. Environ. Microbiol. 2005, 71, 6165–6174. [Google Scholar] [CrossRef]
- Hutchison, M.L.; Walters, L.D.; Moore, A.; Crookes, K.M.; Avery, S.M. Effect of length of time before incorporation on survival of pathogenic bacteria present in livestock wastes applied to agricultural soil. Appl. Environ. Microbiol. 2004, 70, 5111–5118. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, M.L.; Walters, L.D.; Moore, T.; Thomas, D.J.; Avery, S.M. Fate of pathogens present in livestock wastes spread onto fescue plots. Appl. Environ. Microbiol. 2005, 71, 691–696. [Google Scholar] [CrossRef]
- Zhang, G.; Ma, L.; Beuchat, L.R.; Erickson, M.C.; Phelan, V.H.; Doyle, M.P. Lack of internalization of Escherichia coli O157:H7 in lettuce (Lactuca sativa L.) after leaf surface and soil inoculation. J. Food Prot. 2009, 72, 2028–2037. [Google Scholar] [CrossRef]
- Gram, L. Potential hazards in cold-smoked fish: Listeria monocytogenes. J. Food Sci. 2001, 66, S1072–S1081. [Google Scholar] [CrossRef]
- Zhu, M.; Du, M.; Cordray, J.; Ahn, D.U. Control of Listeria monocytogenes contamination in ready-to-eat meat products. Compr. Rev. Food Sci. Food Saf. 2005, 4, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Matthews, K.R. Sources of enteric pathogen contamination of fruits and vegetables: Future directions of research. Stewart Postharvest Rev. 2013, 9, 1–5. [Google Scholar] [CrossRef]
- Ackers, M.L.; Mahon, B.E.; Leahy, E.; Goode, B.; Damrow, T.; Hayes, P.S.; Bibb, W.F.; Rice, D.H.; Barrett, T.J.; Hutwagner, L.; et al. An outbreak of Escherichia coli O157:H7 infections associated with leaf lettuce consumption. J. Infect. Dis. 1998, 177, 1588–1593. [Google Scholar] [CrossRef]
- Bernstein, N.; Sela, S.; Neder-Lavon, S. Effect of irrigation regimes on persistence of Salmonella enterica serovar Newport in small experimental pots designed for plant cultivation. Irrig. Sci. 2007, 26, 1–8. [Google Scholar] [CrossRef]
- Kapperud, G.; Rorvik, L.M.; Hasseltvedt, V.; Hoiby, E.A.; Iversen, B.G.; Staveland, K.; Johnsen, G.; Leitao, J.; Herikstad, H.; Andersson, Y.; et al. Outbreak of Shigella sonnei infection traced to imported iceberg lettuce. J. Clin. Microbiol. 1995, 33, 609–614. [Google Scholar] [CrossRef]
- Mercanoglu Taban, B.; Halkman, A.K. Do leafy green vegetables and their ready-to-eat [RTE] salads carry a risk of foodborne pathogens? Anaerobe 2011, 17, 286–287. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, E.; Einöder-Moreno, M.; Borgen, K.; Brandal, L.T.; Diab, L.; Fossli, Ø.; Herrador, B.G.; Hassan, A.A.; Johannessen, G.S.; Johansen, E.J.; et al. National outbreak of Yersinia enterocolitica infections in military and civilian populations associated with consumption of mixed salad, Norway, 2014. Eurosurveillance 2016, 21, 30321. [Google Scholar] [CrossRef]
- Nuorti, J.P.; Niskanen, T.; Hallanvuo, S.; Mikkola, J.; Kela, E.; Hatakka, M.; Fredriksson-Ahomaa, M.; Lyytikainen, O.; Siitonen, A.; Korkeala, H.; et al. A widespread outbreak of Yersinia pseudotuberculosis O:3 infection from iceberg lettuce. J. Infect. Dis. 2004, 189, 766–774. [Google Scholar] [CrossRef]
- Parker, C.T.; Kyle, J.L.; Huynh, S.; Carter, M.Q.; Brandl, M.T.; Mandrell, R.E. Distinct transcriptional profiles and phenotypes exhibited by Escherichia coli O157:H7 isolates related to the 2006 spinach-associated outbreak. Appl. Environ. Microbiol. 2012, 78, 455–463. [Google Scholar] [CrossRef]
- Sharapov, U.M.; Wendel, A.M.; Davis, J.P.; Keene, W.E.; Farrar, J.; Sodha, S.; Hyytia-Trees, E.; Leeper, M.; Gerner-Smidt, P.; Griffin, P.M.; et al. Multistate outbreak of Escherichia coli O157:H7 infections associated with consumption of fresh spinach: United States, 2006. J. Food Prot. 2016, 79, 2024–2030. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Yang, B.; Wang, Z.; Wang, S.; Zhang, X.; Zhou, Y.; Pang, B.; Diao, B.; Yang, R.; Wu, S.; et al. A large-scale community-based outbreak of paratyphoid fever caused by hospital-derived transmission in southern china. PLoS Negl. Trop. Dis. 2015, 9, e0003859. [Google Scholar] [CrossRef] [PubMed]
- Allende, A.; Monaghan, J. Irrigation water quality for leafy crops: A perspective of risks and potential solutions. Int. J. Environ. Res. Public Health 2015, 12, 7457–7477. [Google Scholar] [CrossRef]
- Uyttendaele, M.; Jaykus, L.-A.; Amoah, P.; Chiodini, A.; Cunliffe, D.; Jacxsens, L.; Holvoet, K.; Korsten, L.; Lau, M.; McClure, P.; et al. Microbial hazards in irrigation water: Standards, norms, and testing to manage use of water in fresh produce primary production. Compr. Rev. Food Sci. Food Saf. 2015, 14, 336–356. [Google Scholar] [CrossRef]
- Makkaew, P.; Miller, M.; Fallowfield, H.J.; Cromar, N.J. Microbial risk in wastewater irrigated lettuce: Comparing Escherichia coli contamination from an experimental site with a laboratory approach. Water Sci. Technol. 2016, 74, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Chitarra, W.; Decastelli, L.; Garibaldi, A.; Gullino, M.L. Potential uptake of Escherichia coli O157:H7 and Listeria monocytogenes from growth substrate into leaves of salad plants and basil grown in soil irrigated with contaminated water. Int. J. Food Microbiol. 2014, 189, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Shouse, P.J.; Grieve, C.M. Quantification of survival of Escherichia coli O157:H7 on plants affected by contaminated irrigation water. Eng. Life Sci. 2006, 6, 566–572. [Google Scholar] [CrossRef]
- Erickson, M.; Liao, J.-Y.; Payton, A.S.; Cook, P.W.; Bakker, H.D.d.; Bautista, J.; Pérez, J.D. Pre-harvest internalization and surface survival of Salmonella and Escherichia coli O157:H7 sprayed onto different lettuce cultivars under field and growth chamber conditions. Int. J. Food Microbiol. 2019, 291, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Van der Linden, I.; Cottyn, B.; Uyttendaele, M.; Vlaemynck, G.; Heyndrickx, M.; Maes, M. Survival of enteric pathogens during butterhead lettuce growth: Crop stage, leaf age, and irrigation. Foodborne Pathog. Dis. 2013, 10, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Usall, J.; Viñas, I.; Solsona, C.; Abadias, M. Transfer of Listeria innocua from contaminated compost and irrigation water to lettuce leaves. Food Microbiol. 2011, 28, 590–596. [Google Scholar] [CrossRef]
- Oliveira, M.; Viñas, I.; Usall, J.; Anguera, M.; Abadias, M. Presence and survival of Escherichia coli O157:H7 on lettuce leaves and in soil treated with contaminated compost and irrigation water. Int. J. Food Microbiol. 2012, 156, 133–140. [Google Scholar] [CrossRef]
- Erickson, M.C.; Webb, C.C.; Diaz-Perez, J.C.; Phatak, S.C.; Silvoy, J.J.; Davey, L.; Payton, A.S.; Liao, J.; Ma, L.; Doyle, M.P. Surface and internalized Escherichia coli O157:H7 on field-grown spinach and lettuce treated with spray-contaminated irrigation water. J. Food Prot. 2010, 73, 1023–1029. [Google Scholar] [CrossRef]
- Bardsley, C.A.; Boyer, R.R.; Rideout, S.L.; Strawn, L.K. Survival of Listeria monocytogenes on the surface of basil, cilantro, dill, and parsley plants. Food Control. 2019, 95, 90–94. [Google Scholar] [CrossRef]
- Bezanson, G.; Delaquis, P.; Bach, S.; McKellar, R.; Topp, E.; Gill, A.; Blais, B.; Gilmour, M. Comparative examination of Escherichia coli O157:H7 survival on romaine lettuce and in soil at two independent experimental sites. J. Food Prot. 2012, 75, 480–487. [Google Scholar] [CrossRef]
- Moyne, A.L.; Sudarshana, M.R.; Blessington, T.; Koike, S.T.; Cahn, M.D.; Harris, L.J. Fate of Escherichia coli O157:H7 in field-inoculated lettuce. Food Microbiol. 2011, 28, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Pauwelyn, E.; Vanhouteghem, K.; Cottyn, B.; De Vos, P.; Maes, M.; Bleyaert, P.; Höfte, M. Epidemiology of Pseudomonas cichorii, the cause of lettuce midrib rot. J. Phytopathol. 2011, 159, 298–305. [Google Scholar] [CrossRef]
- Ashbolt, N.J.; Grabow, W.O.K.; Snozzi, M. Indicators of microbial water quality. In Water Quality: Guidelines, Standards & Health; Fewtrell, L., Bartram, B., Eds.; IWA Publishing: London, UK, 2001; pp. 289–315. [Google Scholar]
- Lee, S.; Khoo, V.S.L.; Medriano, C.A.D.; Lee, T.; Park, S.Y.; Bae, S. Rapid and in-situ detection of fecal indicator bacteria in water using simple DNA extraction and portable loop-mediated isothermal amplification (LAMP) PCR methods. Water Res. 2019, 160, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Pachepsky, Y.; Shelton, D.; Dorner, S.; Whelan, G. Can E. coli or thermotolerant coliform concentrations predict pathogen presence or prevalence in irrigation waters? Crit. Rev. Microbiol. 2016, 42, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Slavik, M.F. A multiplex polymerase chain reaction assay for rapid detection of Escherichia coli O157:H7, Listeria monocytogenes, Salmonella Typhimurium and Camplylobacter jejuni in artifically contaminated food samples. J. Rapid Met. Aut. Microbiol. 2005, 13, 213–223. [Google Scholar] [CrossRef]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors—Occurrence, properties and removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef]
- Truchado, P.; Gil, M.I.; Kostic, T.; Allende, A. Optimization and validation of a PMA qPCR method for Escherichia coli quantification in primary production. Food Control. 2016, 62, 150–156. [Google Scholar] [CrossRef]
- Varma, M.; Field, R.; Stinson, M.; Rukovets, B.; Wymer, L.; Haugland, R. Quantitative real-time PCR analysis of total and propidium monoazide-resistant fecal indicator bacteria in wastewater. Water Res. 2009, 43, 4790–4801. [Google Scholar] [CrossRef]
- Dinu, L.-D.; Bach, S. Detection of viable but non-culturable Escherichia coli O157:H7 from vegetable samples using quantitative PCR with propidium monoazide and immunological assays. Food Control 2013, 31, 268–273. [Google Scholar] [CrossRef]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef]
- Brennan, F.P.; Abram, F.; Chinalia, F.A.; Richards, K.G.; O’Flaherty, V. Characterization of environmentally persistent Escherichia coli isolates leached from an Irish soil. Appl. Environ. Microbiol. 2010, 76, 2175–2180. [Google Scholar] [CrossRef] [PubMed]
- Brennan, F.P.; Grant, J.; Botting, C.H.; O’Flaherty, V.; Richards, K.G.; Abram, F. Insights into the low-temperature adaptation and nutritional flexibility of a soil-persistent Escherichia Coli. FEMS Microbiol. Ecol. 2013, 84, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Brennan, F.P.; O’Flaherty, V.; Kramers, G.; Grant, J.; Richards, K.G. Long-term persistence and leaching of Escherichia coli in temperate maritime soils. Appl. Environ. Microbiol. 2010, 76, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, C.D.; Davies, A.R. Development of antibiotic-resistant strains for the enumeration of foodborne pathogenic bacteria in stored foods. Int. J. Food Microbiol. 1994, 24, 125–136. [Google Scholar] [CrossRef]
- Abberton, C.L.; Bereschenko, L.; van der Wielen, P.W.; Smith, C.J. Survival, biofilm formation, and growth potential of environmental and enteric Escherichia coli strains in drinking water microcosms. Appl. Environ. Microbiol. 2016, 82, 5320–5331. [Google Scholar] [CrossRef] [PubMed]
- Chern, E.C.; Brenner, K.P.; Wymer, L.; Haugland, R.A. Comparison of fecal indicator bacteria densities in marine recreational waters by QPCR. Water Qual. Expo Health 2009, 1, 203–214. [Google Scholar] [CrossRef]
- Rodríguez-Lázaro, D.; Hernández, M.; Scortti, M.; Esteve, T.; Vázquez-Boland, J.A.; Pla, M. Quantitative detection of Listeria monocytogenes and Listeria innocua by real-time PCR: Assessment of hly, iap, and lin02483 targets and AmpliFluor technology. Appl. Environ. Microbiol. 2004, 70, 1366–1377. [Google Scholar] [CrossRef]
- Solomon, E.B.; Pang, H.J.; Matthews, K.R. Persistence of Escherichia coli O157:H7 on lettuce plants following spray irrigation with contaminated water. J. Food Prot. 2003, 66, 2198–2202. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.C.; Liao, J.Y.; Payton, A.S.; Cook, P.W.; Bakker, H.C.D.; Bautista, J.; Díaz-Pérez, J.C. Survival of Salmonella enterica and Escherichia coli O157:H7 sprayed onto the foliage of field-grown cabbage plants. J. Food Prot. 2019, 82, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Callejas, A.; López-Velasco, G.; Camacho, A.B.; Artés, F.; Artés-Hernández, F.; Suslow, T.V. Survival and distribution of Escherichia coli on diverse fresh-cut baby leafy greens under preharvest through postharvest conditions. Int. J. Food Microbiol. 2011, 151, 216–222. [Google Scholar] [CrossRef]
- Weller, D.L.; Kovac, J.; Roof, S.; Kent, D.J.; Tokman, J.I.; Kowalcyk, B.; Oryang, D.; Ivanek, R.; Aceituno, A.; Sroka, C.; et al. Survival of Escherichia coli on lettuce under field conditions encountered in the Northeastern United States. J. Food Prot. 2017, 80, 1214–1221. [Google Scholar] [CrossRef]
- Wood, J.D.; Bezanson, G.S.; Gordon, R.J.; Jamieson, R. Population dynamics of Escherichia coli inoculated by irrigation into the phyllosphere of spinach grown under commercial production conditions. Int. J. Food Microbiol. 2010, 143, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Van der Linden, I.; Eriksson, M.; Uyttendaele, M.; Devlieghere, F. Is there a relation between the microscopic leaf morphology and the association of Salmonella and Escherichia coli O157:H7 with iceberg lettuce leaves? J. Food Prot. 2016, 79, 1784–1788. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.H.; Vogel, B.F.; Gram, L. Prevalence and survival of Listeria monocytogenes in Danish aquatic and fish-processing environments. J. Food Prot. 2006, 69, 2113–2122. [Google Scholar] [CrossRef] [PubMed]
- Budzinska, K.; Wronski, G.; Szejniuk, B. Survival time of bacteria Listeria monocytogenes in water environment and sewage. Pol. J. Environ. Stud. 2012, 21, 31–37. [Google Scholar]
- Van Der Linden, I.; Cottyn, B.; Uyttendaele, M.; Berkvens, N.; Vlaemynck, G.; Heyndrickx, M.; Maes, M. Enteric pathogen survival varies substantially in irrigation water from Belgian lettuce producers. Int. J. Environ. Res. Public Health 2014, 11, 10105–10124. [Google Scholar] [CrossRef]
- Rudi, K.; Moen, B.; Drømtorp, S.M.; Holck, A.L. Use of ethidium monoazide and PCR in combination for quantification of viable and dead cells in complex samples. Appl. Environ. Microbiol. 2005, 71, 1018–1024. [Google Scholar] [CrossRef]
- Dinu, L.D.; Bach, S. Induction of viable but nonculturable Escherichia coli O157:H7 in the phyllosphere of lettuce: A food safety risk factor. Appl. Environ. Microbiol. 2011, 77, 8295–8302. [Google Scholar] [CrossRef]
- Machado-Moreira, B.; Monteiro, S.; Santos, R.; Martinez-Murcia, A.; Rajkovic, A.; Smigic, N.; Richards, K.G.; Abram, F.; Burgess, C.M. Impact of beef extract used for sample concentration on the detection of Escherichia coli DNA in water samples via qPCR. J. Microbiol. Methods 2020, 168, 105786. [Google Scholar] [CrossRef]
- Boulos, L.; Prévost, M.; Barbeau, B.; Coallier, J.; Desjardins, R. LIVE/DEAD BacLight: Application of a new rapid staining method for direct enumeration of viable and total bacteria in drinking water. J. Microbiol. Methods 1999, 37, 77–86. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E. coli | L. innocua | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t (Days) | Culture (Log10 cfu/mL) | qPCR (Log10 gu/mL) | Culture (Log10 cfu/mL) | qPCR (Log10 gu/mL) | ||||||||
| Rep 1 | Rep 2 | Rep 3 | Rep 1 | Rep 2 | Rep 3 | Rep 1 | Rep 2 | Rep 3 | Rep 1 | Rep 2 | Rep 3 | |
| 0 | 3.78 | 3.74 | 4.60 | 4.98 | 5.06 | 4.58 | 4.54 | 5.88 | 5.77 | 6.07 | 6.34 | 6.14 |
| 1 | −0.38 | 0.29 | 2.31 | 4.94 | 4.79 | 4.84 | 4.84 | 3.62 | 2.75 | 6.55 | 6.23 | 6.61 |
| 3 | −1.77 | −2.35 | −1.39 | 5.02 | 4.95 | 4.62 | −0.06 | −2.43 a | −1.23 | 6.38 | 6.36 | 6.34 |
| 7 | −2.32 a | 0.35 | −2.32 a | 4.59 | 4.67 | 4.73 | ND | ND | ND | 6.60 | 6.33 | 6.39 |
| 10 | ND b | −2.26 a | −2.26 a | 4.68 | 4.85 | 4.69 | 0.75 | ND | ND | 6.68 | 6.60 | 6.66 |
| 14 | −2.32 a | −2.32 a | −2.32 a | 4.71 | 4.87 | 4.64 | −0.35 | ND | ND | 6.76 | 6.35 | 6.74 |
| 21 | ND | −2.35 a | −2.35 a | 4.26 | 4.55 | 4.58 | ND | −0.97 | −0.06 | 6.68 | 6.66 | 6.53 |
| 28 | ND | −2.35 a | −2.35 a | 4.59 | 4.68 | 4.69 | ND | −2.35 a | 2.01 | 6.85 | 6.69 | 6.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machado-Moreira, B.; Richards, K.; Abram, F.; Brennan, F.; Gaffney, M.; Burgess, C.M. Survival of Escherichia coli and Listeria innocua on Lettuce after Irrigation with Contaminated Water in a Temperate Climate. Foods 2021, 10, 2072. https://doi.org/10.3390/foods10092072
Machado-Moreira B, Richards K, Abram F, Brennan F, Gaffney M, Burgess CM. Survival of Escherichia coli and Listeria innocua on Lettuce after Irrigation with Contaminated Water in a Temperate Climate. Foods. 2021; 10(9):2072. https://doi.org/10.3390/foods10092072
Chicago/Turabian StyleMachado-Moreira, Bernardino, Karl Richards, Florence Abram, Fiona Brennan, Michael Gaffney, and Catherine M. Burgess. 2021. "Survival of Escherichia coli and Listeria innocua on Lettuce after Irrigation with Contaminated Water in a Temperate Climate" Foods 10, no. 9: 2072. https://doi.org/10.3390/foods10092072
APA StyleMachado-Moreira, B., Richards, K., Abram, F., Brennan, F., Gaffney, M., & Burgess, C. M. (2021). Survival of Escherichia coli and Listeria innocua on Lettuce after Irrigation with Contaminated Water in a Temperate Climate. Foods, 10(9), 2072. https://doi.org/10.3390/foods10092072