
Microbial Regulation of Deterioration and Preservation of Salted Kelp under Different Temperature and Salinity Conditions

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Kelp Collection and Setup
2.2. Detection of Bioenzyme Activity in Salted Kelp Samples
2.3. High-Throughput Sequencing of Amplicons and Metagenomics in Salted Kelp Samples
2.4. High-Throughput Sequencing Data Analysis
3. Results
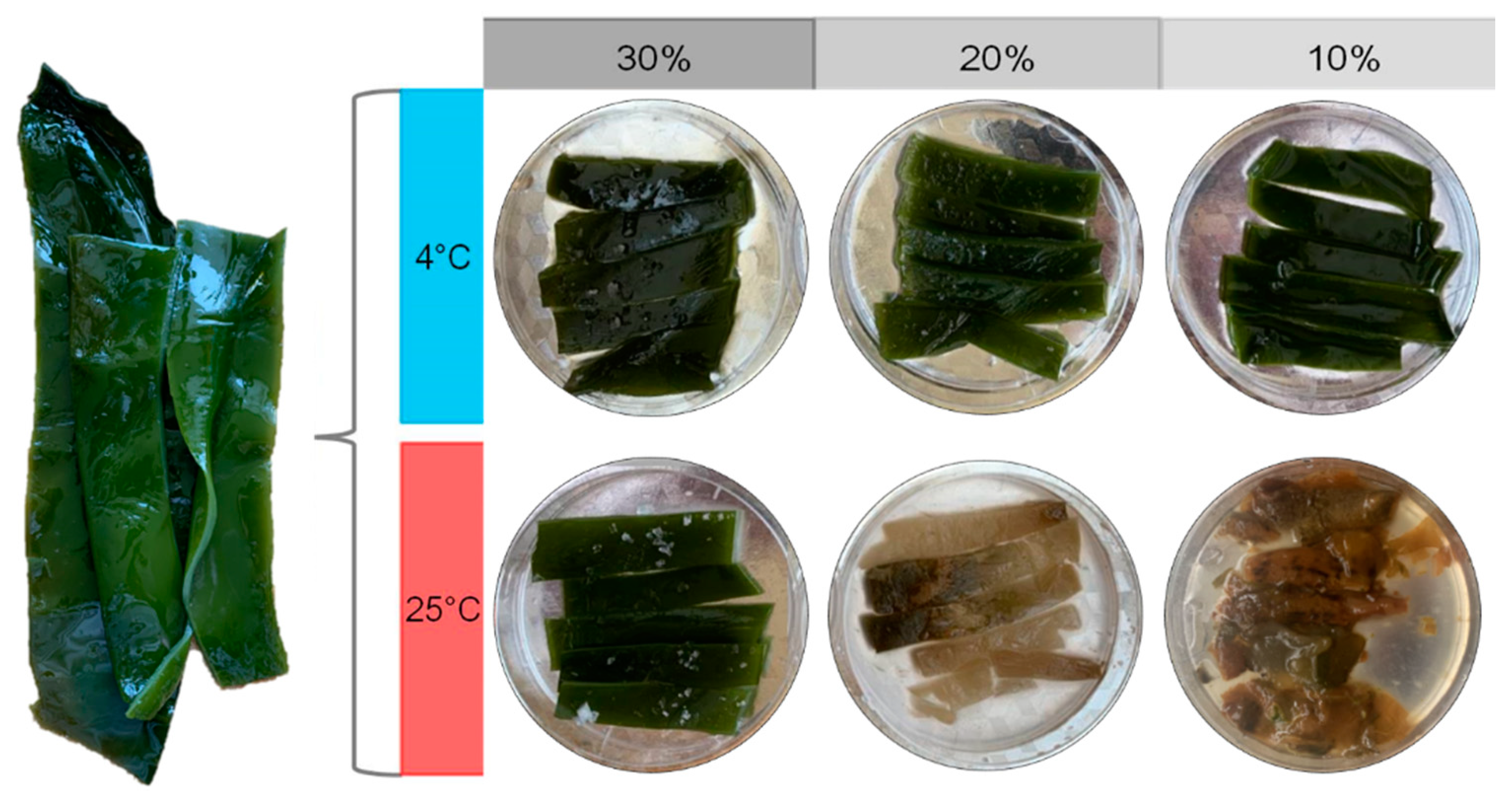
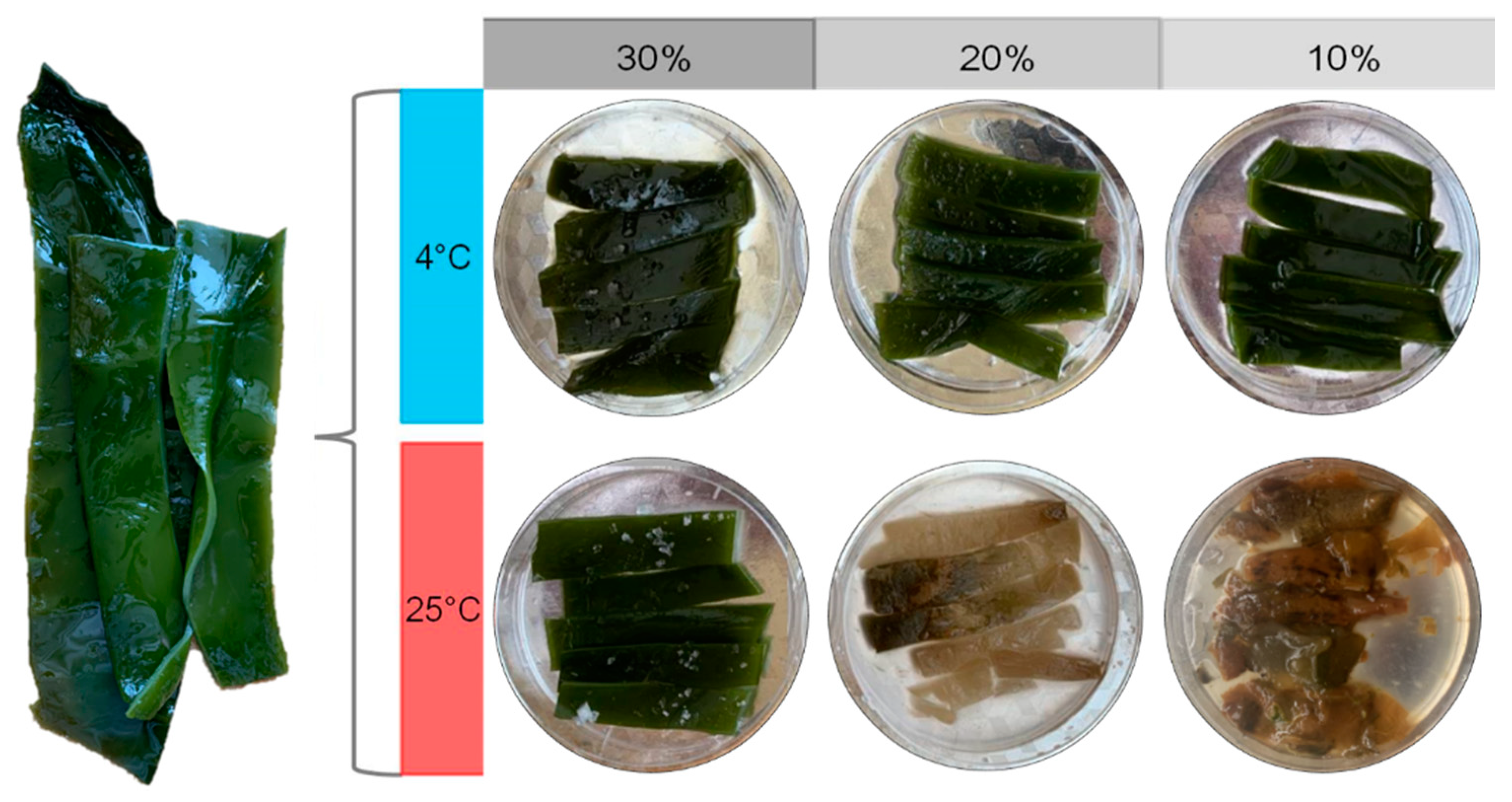
3.1. The States of Salted Kelp Samples after 30 Days of Preservation
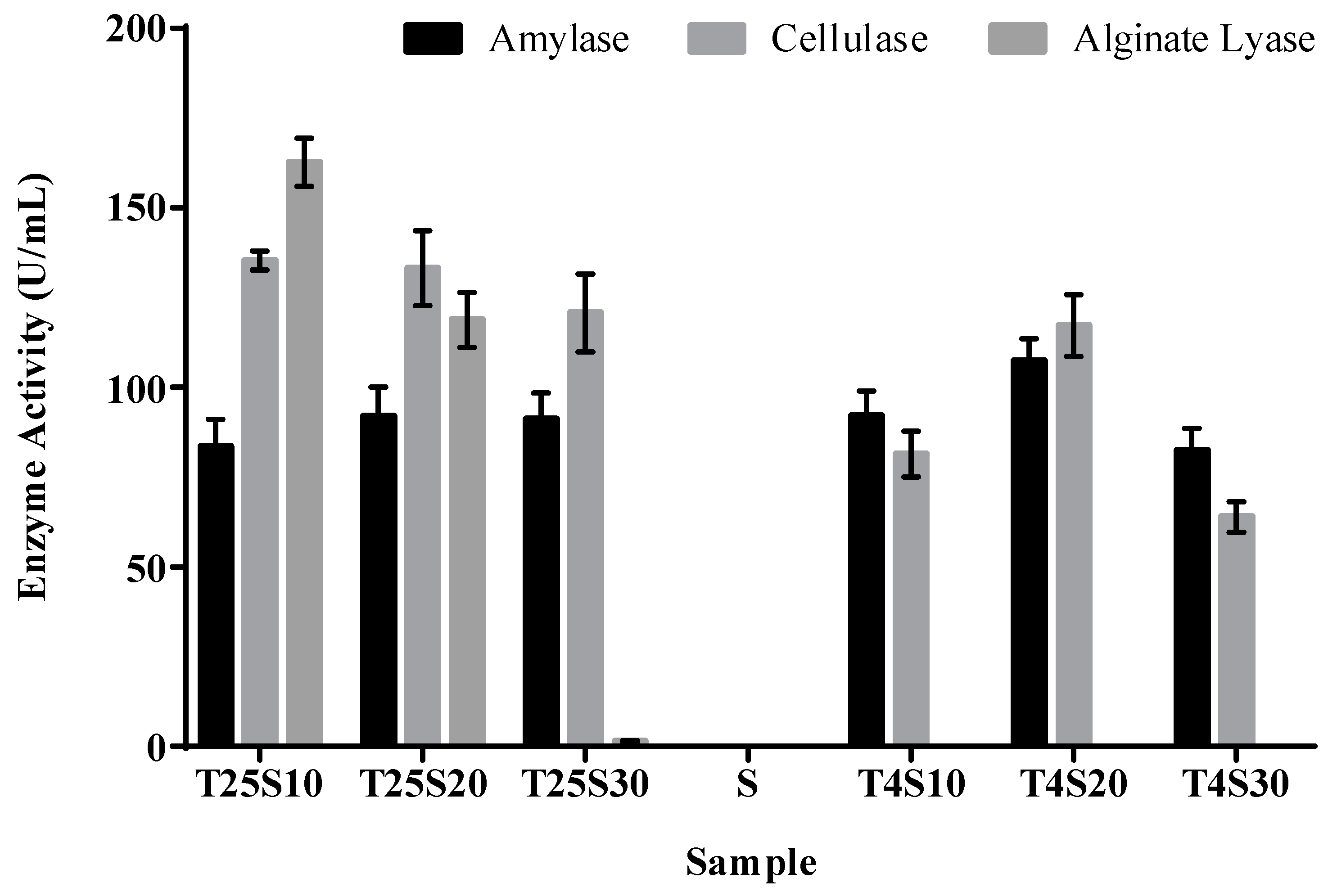
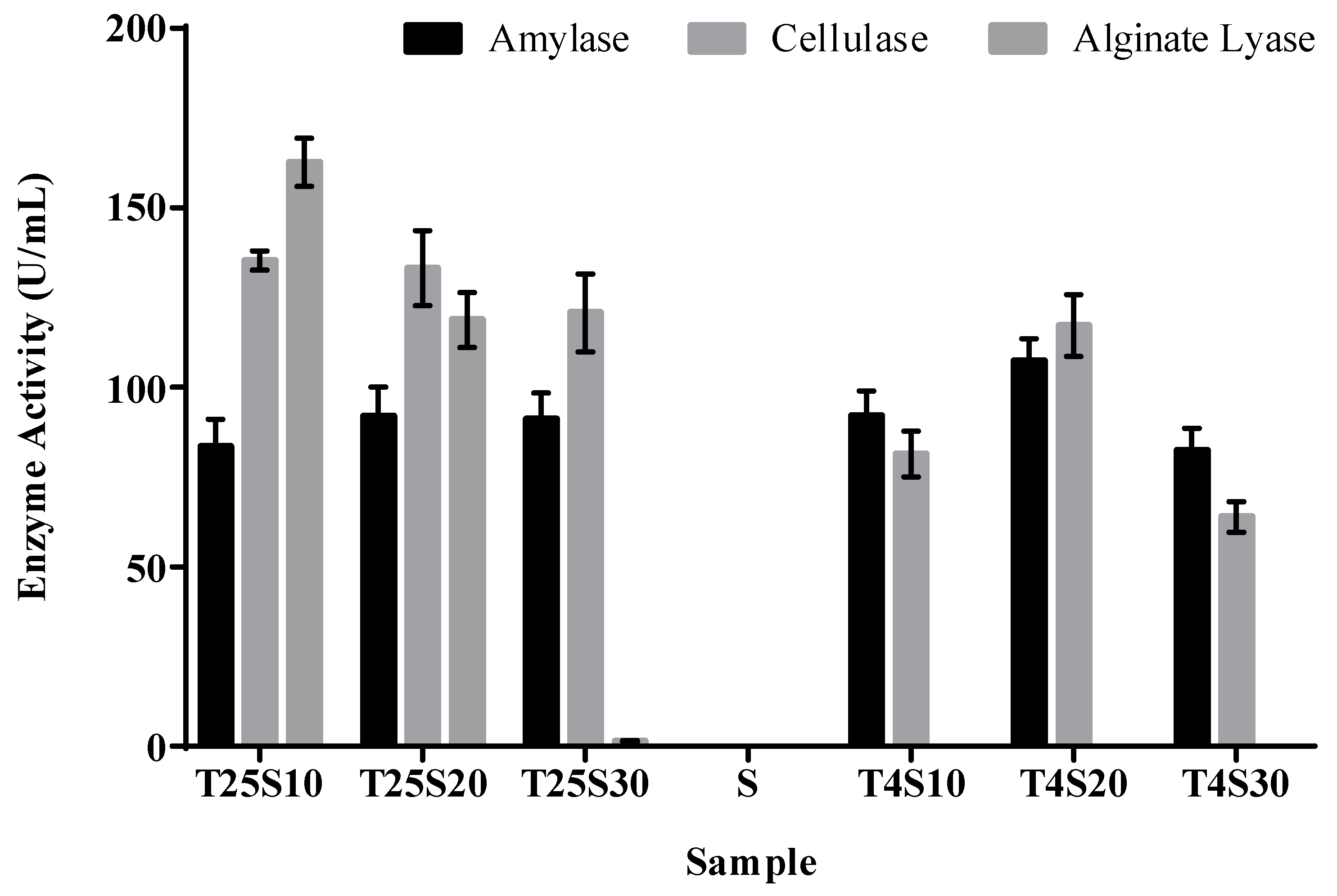
3.2. Activity of Algal Polysaccharide Lyase in Salted Kelp Samples
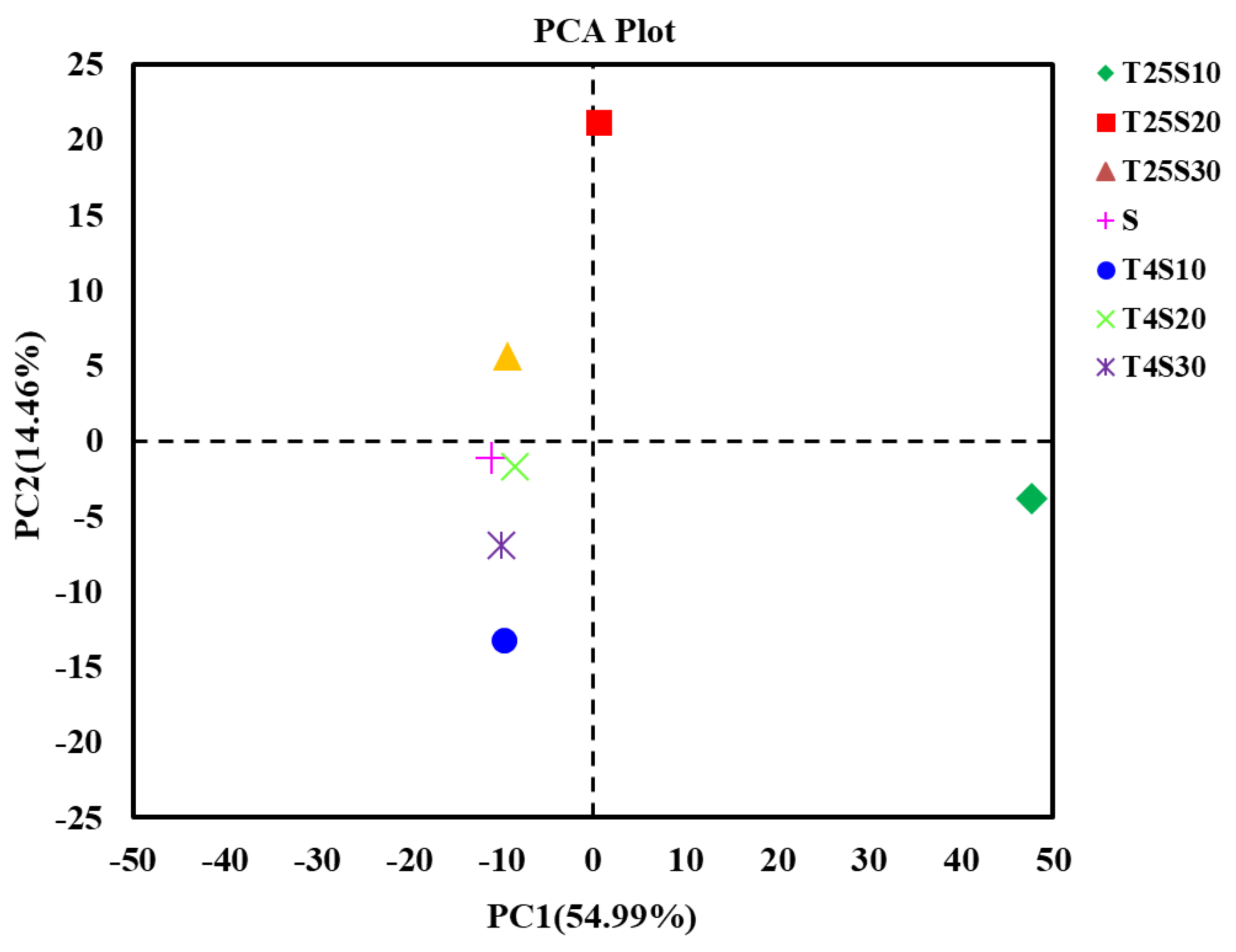
3.3. Microbial Community Diversity in Salted Kelp Samples
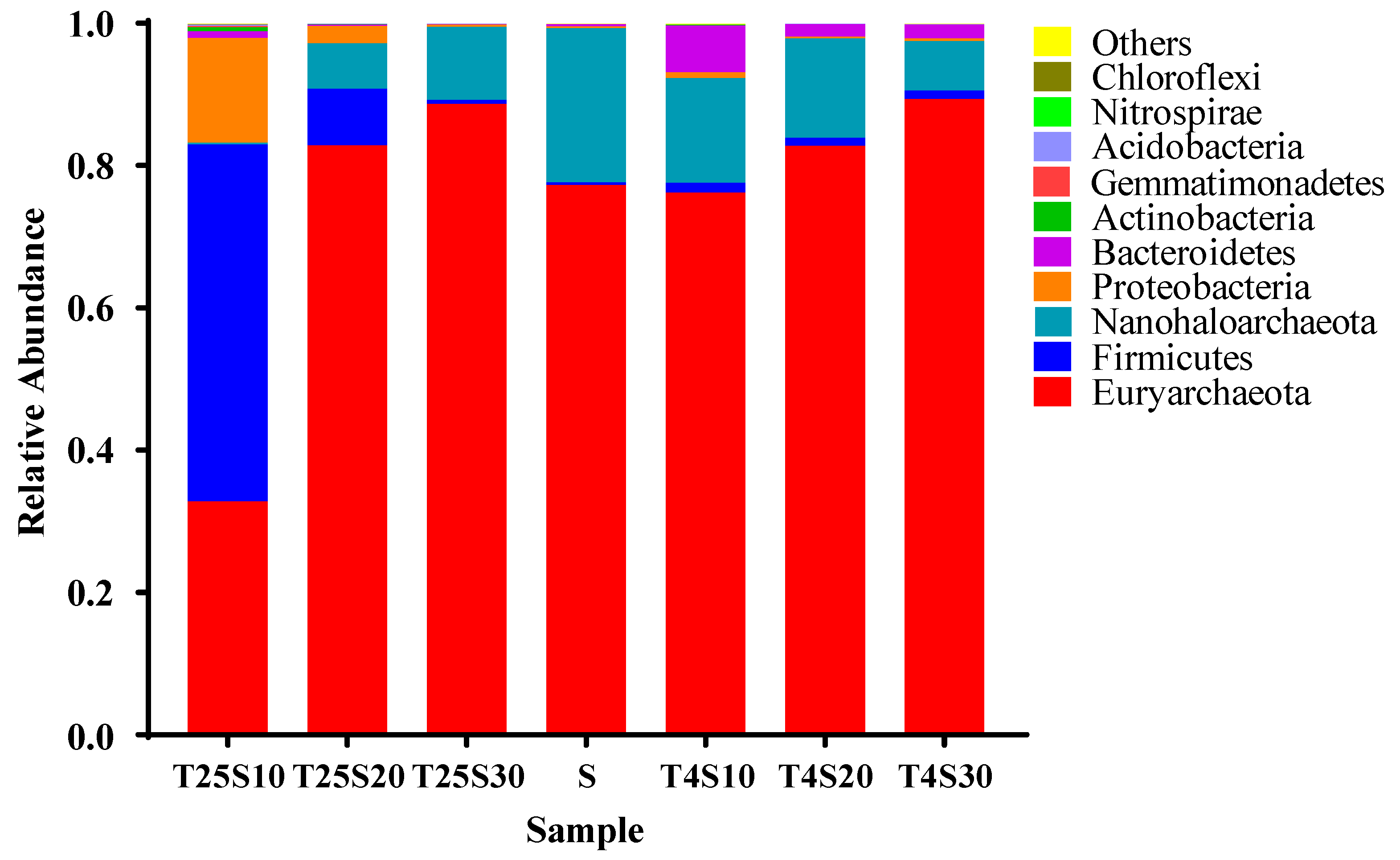
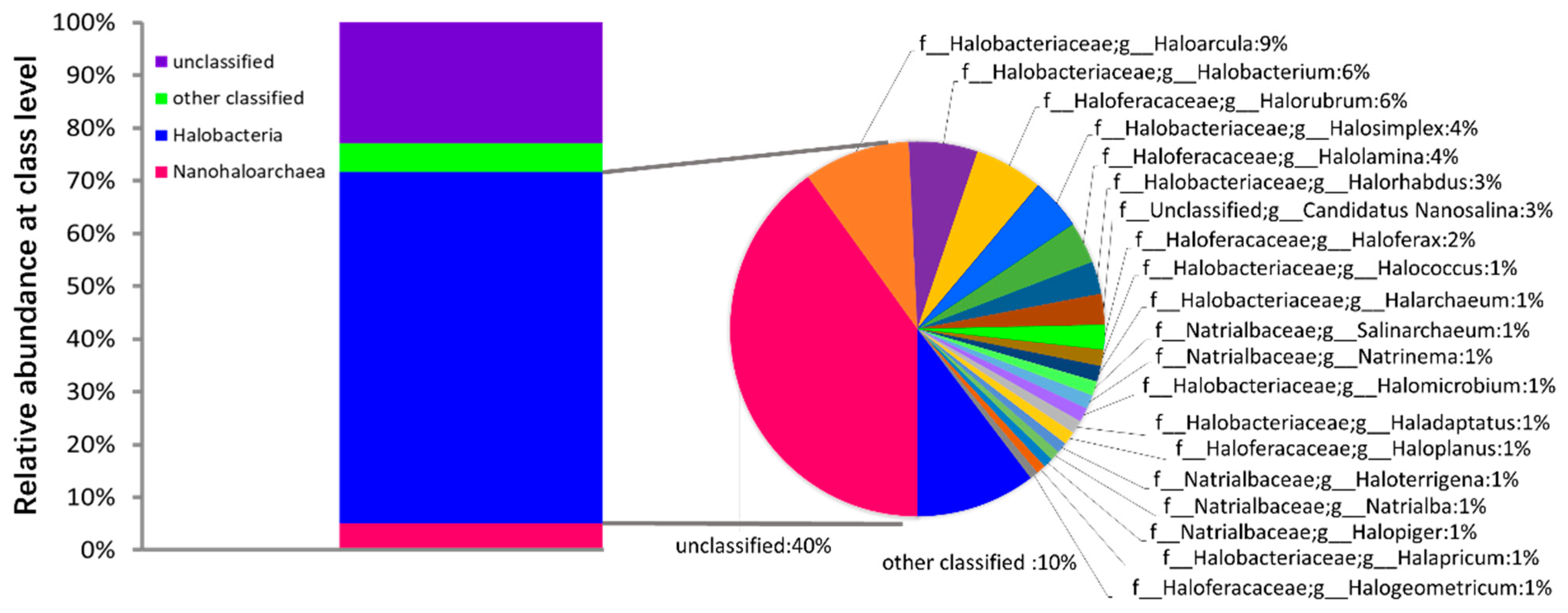
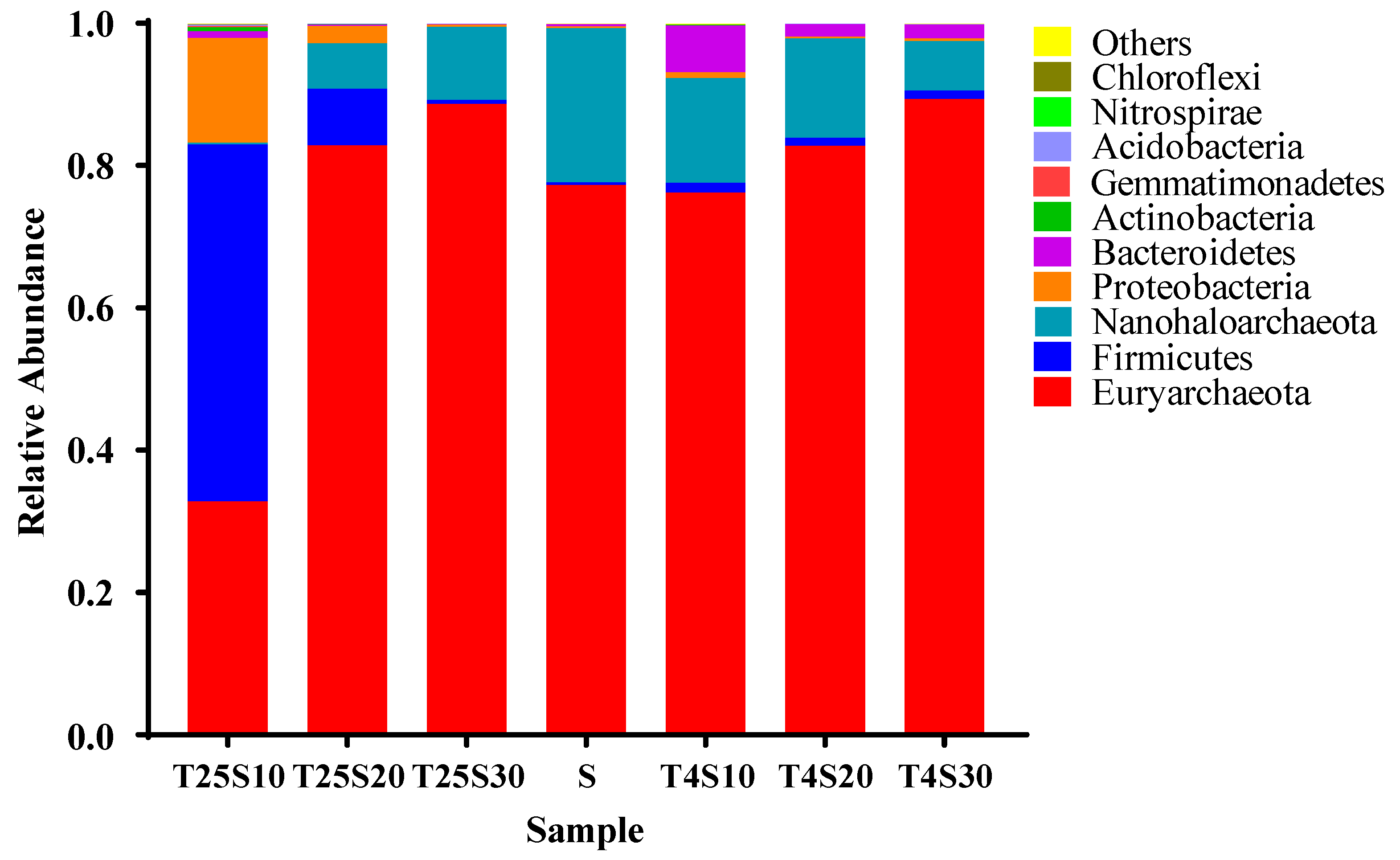
3.4. Composition and Source of Microbial Communities in Salted Kelp Samples
3.5. Microbial Function Analysis of Rotten and Undamaged Salted Kelp Based on the Metagenome
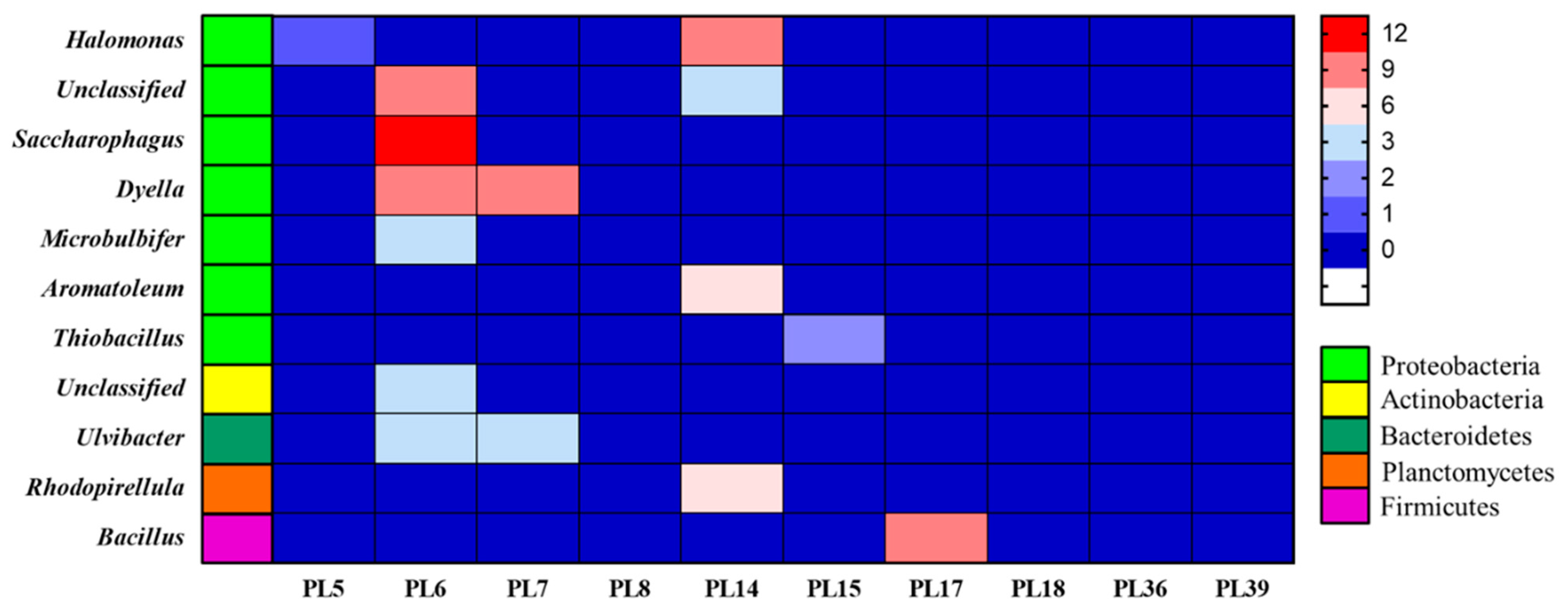
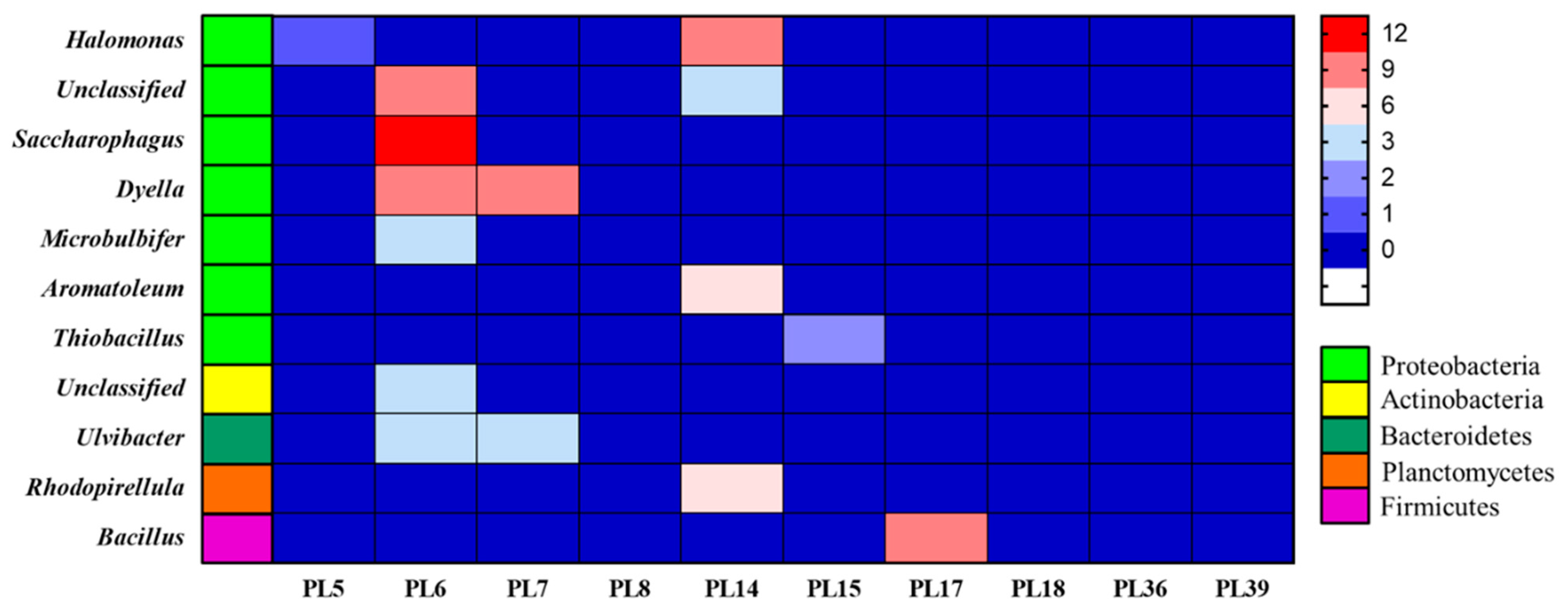
3.6. Species Annotation of Rotten and Undamaged Salted Kelp Microbial Metagenomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiao, L.; Feng, H.; Yang, Z.; Lu, X.Z.; Yu, W.G. A Novel Alginate Lyase with High Activity on Acetylated Alginate of Pseudomonas aeruginosa FRD1 from Pseudomonas sp. QD03. World J. Microbiol. Biotechnol. 2006, 22, 81–88. [Google Scholar] [CrossRef]
- An, Q.-D.; Zhang, G.-L.; Wu, H.-T.; Zhang, Z.-C.; Zheng, G.-S. Alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. J. Appl. Microbiol. 2009, 106, 161–170. [Google Scholar] [CrossRef]
- Thomas, N.V.; Kim, S.K. Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Environ. Toxicol. Pharmacol. 2011, 32, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.H.; Kim, Y.M.; Kim, S.K. Antimicrobial effect of phlorotannins from marine brown algae. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2012, 50, 3251–3255. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C. Comprehensive utilization of fresh kelp processing in China. China Fish. 2012, 10, 22–23. (In Chinese) [Google Scholar]
- Jennifer, J.P.; Alison, B.; Denise, I.S. Influence of dry salting on quality attributes of farmed kelp (Alaria esculenta) during long-term refrigerated storage. LWT Food Sci. Technol. 2019, 114, 108362. [Google Scholar]
- Yang, Z.; Tang, X.X.; Gong, X.Z.; Fang, F.; Lu, W.T. Histological study on kelp disease caused by alginic acid degrading bacteria. Mar. Sci. 2000, 24, 1–2. (In Chinese) [Google Scholar]
- ØStgaard, K.; Knutsen, S.H.; Dyrset, N.; Aasen, I.M. Production and characterization of guluronate lyase from Klebsiella pneumoniae for applications in seaweed biotechnology. Enzym. Microb. Technol. 1993, 15, 756–763. [Google Scholar] [CrossRef]
- Iwamoto, Y.; Araki, R.; Iriyama, K.I.; Oda, T.; Fukuda, H.; Hayashida, S.; Muramatsu, T. Purification and characterization of bifunctional alginate lyase from Alteromonas sp. strain no. 272 and its action on saturated oligomeric substrates. J. Agric. Chem. Soc. Jpn. 2001, 65, 133–142. [Google Scholar] [CrossRef]
- Miyazaki, M.; Obata, J.; Iwamoto, Y.; Oda, T.; Muramatsu, T. Calcium-sensitive extracellular poly(α-L-guluronate) lyase from a marine bacterium Pseudomonas sp. strain F6: Purification and some properties. Fish. Sci. 2001, 67, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, W.; Miyake, O.; Ochiai, A.; Murata, K. Molecular identification of Sphingomonas sp. A1 Alginate lyase (A1-IV′) as a member of novel polysaccharide lyase family 15 and implications in alginate lyase evolution. J. Biosci. Bioeng. 2005, 99, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Jagtap, S.; Nair, S.K. Structure of a PL17 family alginate lyase demonstrates functional similarities among exotype depolymerases. J. Biol. Chem. 2014, 289, 8645–8655. [Google Scholar] [CrossRef] [Green Version]
- Sheng, D.; Wei, T.D.; Chen, X.L.; Li, C.Y.; Peng, W.; Xie, B.B.; Qin, Q.L.; Zhang, X.Y.; Pang, X.H.; Zhou, B.C. Molecular insight into the role of the N-terminal extension in the maturation, substrate recognition, and catalysis of a bacterial alginate lyase from polysaccharide lyase family 18. J. Biol. Chem. 2014, 289, 29558–29569. [Google Scholar]
- Zhang, Z.; Su, H.; Wang, X.; Tang, L.; Han, F. Cloning and characterization of a novel chondroitinase ABC categorized into a new subfamily of polysaccharide lyase family 8. Int. J. Biol. Macromol. 2020, 164, 3762–3770. [Google Scholar] [CrossRef]
- Dong, F.; Xu, F.; Chen, X.L.; Li, P.Y.; Zhang, Y.Z. Alginate lyase Aly36B is a new bacterial member of the polysaccharide lyase family 36 and catalyzes by a novel mechanism with lysine as both the catalytic base and catalytic acid. J. Biol. Chem. 2019, 431, 4897–4909. [Google Scholar] [CrossRef]
- Ji, S.Q.; Dix Samuel, R.; Aziz Adli, A.; Sedelnikova, S.E.; Baker, P.J.; Rafferty, J.B.; Tzokov, S.B.; Agirre, J.; Li, F.-L.; Rice, D.W.; et al. The molecular basis of endolytic activity of a multidomain alginate lyase from Defluviitalea phaphyphila, a representative of a new lyase family, PLxx. J. Biol. Chem. 2019, 294, 18077–18091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D. Study on the Microbiological Nature of the Red Rot Phenomenon of Salted Kelp. Master’s Thesis, Jiangsu University, Zhenjiang, China, 2015. (In Chinese). [Google Scholar]
- Hong, P.T.K.; Jang, C.H. Simple, Sensitive Technique for α-Amylase Detection Facilitated by Liquid Crystal-based Microcapillary Sensors—ScienceDirect. Microchem. J. 2020, 162, 105864. [Google Scholar] [CrossRef]
- Wei, T.; Xu, Q.; Zou, C.; He, Z.; Dai, Z. A boronate-modified renewable nanointerface for ultrasensitive electrochemical assay of cellulase activity. Chin. Chem. Lett. 2020, 32, 1470–1474. [Google Scholar] [CrossRef]
- Preiss, J.; Ashwell, G. Alginic acid metabolism in bacteria I. Enzymatic formation of unsaturated oligosaccharides and 4-deoxy-1-erythro-5-hexoseulose uronic acid. J. Biol. Chem. 1962, 237, 309–316. [Google Scholar] [CrossRef]
- Zhu, B.W.; Huang, L.; Tan, H.D.; Qin, Y.Q.; Du, Y.G.; Yin, H. Characterization of a new endo-type polyM-specific alginate lyase from Pseudomonas sp. Biotechnol. Lett. 2015, 37, 409–415. [Google Scholar] [CrossRef]
- Su, I.L.; Choi, S.H.; Lee, E.Y.; Kim, H.S. Molecular cloning, purification, and characterization of a novel polyMG-specific alginate lyase responsible for alginate MG block degradation in Stenotrophomas maltophilia KJ-2. Appl. Microbiol. Biotechnol. 2012, 95, 1643–1653. [Google Scholar]
- Ghadam, P.; Akhlaghi, F.; Abdi, A. One-step purification and characterization of alginate lyase from a clinical Pseudomonas aeruginosa with destructive activity on bacterial biofilm. Iran. J. Basic Med. Sci. 2017, 20, 467–473. [Google Scholar] [PubMed]
- Suzuki, H.; Suzuki, K.I.; Inoue, A.; Ojima, T. A novel oligoalginate lyase from abalone, Haliotis discus hannai, that releases disaccharide from alginate polymer in an exolytic manner. Carbohydr. Res. 2006, 341, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Ogura, K.; Yamasaki, M.; Yamada, T.; Mikami, B.; Hashimoto, W.; Murata, K. Crystal structure of family 14 polysaccharide lyase with pH-dependent modes of action. J. Biol. Chem. 2009, 284, 35572–35579. [Google Scholar] [CrossRef] [Green Version]
- Mikami, B.; Ban, M.; Suzuki, S.; Yoon, H.J.; Miyake, O.; Yamasaki, M.; Ogura, K.; Maruyama, Y.; Hashimoto, W.; Murata, K. Induced-fit motion of a lid loop involved in catalysis in alginate lyase A1-III. Acta Crystallogr. 2012, 68, 1207–1216. [Google Scholar] [CrossRef] [Green Version]
- Jn, A.; Ai, A.; So, B.; To, A. Production of a novel dimeric 4-deoxy-L-erythro-5-hexoseulose uronic acid by a PL-17 exolytic alginate lyase from Hydrogenophaga sp. UMI-18. Biochem. Biophys. Res. Commun. 2020, 525, 982–988. [Google Scholar]
- Cheng, D.; Liu, Z.; Jiang, C.; Li, L.; Mao, X. Biochemical characterization and degradation pattern analysis of a novel PL-6 alginate lyase from Streptomyces coelicolor A3(2). Food Chem. 2020, 323, 126852. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Dong, S.; Li, F.L.; Ma, X.Q. Structural basis for the exolytic activity of polysaccharide lyase family 6 alginate lyase BcAlyPL6 from human gut microbe Bacteroides clarus. Biochem. Biophys. Res. Commun. 2021, 547, 111–117. [Google Scholar] [CrossRef]
- Tao, W.P. The halophilic mechanism of halophilic bacteria. Bull. Biol. 1996, 31, 23–24. (In Chinese) [Google Scholar]
- Zha, X.Q.; Lu, C.Q.; Cui, S.H.; Pan, L.H.; Zhang, H.L.; Wang, J.H.; Luo, J.P. Structural identification and immunostimulating activity of a Laminaria japonica polysaccharide. Int. J. Biol. Macromol. 2015, 78, 429–438. [Google Scholar] [CrossRef]
- Tesson, B.; Charrier, B. Brown algal morphogenesis: Atomic force microscopy as a tool to study the role of mechanical forces. Front. Plant Sci. 2014, 5, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.W.; Ni, F.; Ning, L.M.; Sun, Y.; Yao, Z. Cloning and characterization of a new pH-stable alginate lyase with high salt tolerance from marine Vibrio sp. NJ-04. Int. J. Biol. Macromol. 2018, 115, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.Y.; Preston, L.A.; Schiller, N.L. ALGINATE LYASE: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu. Rev. Microbiol. 2000, 54, 289–340. [Google Scholar] [CrossRef]
- Kim, H.T.; Ko, H.J.; Kim, N.; Kim, D.; Lee, D.; Choi, I.G.; Woo, H.C.; Kim, M.; Kim, K.H. Characterization of a recombinant endo-type alginate lyase (Alg7D) from Saccharophagus degradans. Biotechnol. Lett. 2012, 34, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Chung, J.H.; Wang, D.; Lee, J.; Woo, H.C.; Choi, I.G.; Kim, K.H. Depolymerization of alginate into a monomeric sugar acid using Alg17C, an exo-oligoalginate lyase cloned from Saccharophagus degradans 2-40. Appl. Microbiol. Biotechnol. 2012, 93, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Kam, N.; Lee, E.Y.; Kim, H.S. Cloning and Characterization of a Novel Oligoalginate Lyase from a Newly Isolated Bacterium Sphingomonas sp. MJ-3. Mar. Biotechnol. 2012, 14, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, S.S.; Hehemann, J.H.; Polz, M.F.; Lee, J.K.; Zhao, H. Comparative biochemical characterization of three exolytic oligoalginate lyases from Vibrio splendidus reveals complementary substrate scope, temperature, and pH adaptations. Appl. Environ. Microbiol. 2014, 80, 4207–4214. [Google Scholar] [CrossRef] [Green Version]
- Kixmuller, D.; Greie, J.C. Construction and characterization of a gradually inducible expression vector for Halobacterium salinarum, based on the kdp Promoter. Appl. Environ. Microbiol. 2012, 78, 2100–2105. [Google Scholar] [CrossRef]
- Kraegeloh, A.; Amendt, B.; Kunte, H.J. Potassium transport in a halophilic member of the bacteria domain: Identification and characterization of the K+ uptake systems TrkH and TrkI from Halomonas elongata DSM 2581T. J. Bacteriol. 2005, 187, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.X.; Liang, M.; Fei, Z.W.; Xie, Z.X. Research on the method of determining the minimum NaCl concentration for extreme halophilic archaea to maintain intact cell morphology. Microbiol. Bull. 2008, 35, 1332–1334. (In Chinese) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | OTU | Unique OTU | Shannon Index | Simpson Index | Chao1 | Goods_Coverage |
|---|---|---|---|---|---|---|
| T25S10 | 554 | 177 | 3.58 | 0.77 | 699.10 | 99.6% |
| T25S20 | 331 | 12 | 4.35 | 0.88 | 393.58 | 99.7% |
| T25S30 | 233 | 4 | 4.91 | 0.91 | 268.00 | 99.9% |
| S | 287 | 15 | 5.63 | 0.96 | 346.43 | 99.8% |
| T4S10 | 305 | 9 | 5.63 | 0.96 | 401.28 | 99.8% |
| T4S20 | 334 | 4 | 5.88 | 0.97 | 469.30 | 99.7% |
| T4S30 | 296 | 8 | 5.49 | 0.95 | 445.50 | 99.7% |
| Item | T25S10 | T4S10 |
|---|---|---|
| Total Len. (bp) | 389,089,268 | 236,304,057 |
| Max Len. (bp) | 43,046 | 94,615 |
| Average Len. (bp) | 998.89 | 1069.79 |
| N50 Len. (bp) | 1023 | 1128 |
| N90 Len. (bp) | 558 | 571 |
| Total Num. | 389,522 | 220,889 |
| Archaeal Num. | 12,231 | 193,350 |
| Bacterial Num. | 301,263 | 9161 |
| Eukaryota Num. | 344 | 144 |
| Virus Num. | 948 | 722 |
| Others | 74,736 | 17,512 |
| Total Num. of Carbohydrate-Active enzymes | 18,712 | 4634 |
| PL | 654 (87) | 56 (0) |
| GT | 5641 | 2504 |
| GH | 6247 | 1393 |
| CE | 2527 | 98 |
| CBM | 3420 | 511 |
| AA | 223 | 72 |
| PL Family | Enzyme | Action Mode | Source of First Report | Environment for Sample Isolation | References |
|---|---|---|---|---|---|
| 5 | AlyA1-III | Endo- | Pseudomonas sp. E03 | Mud | [21] |
| 6 | AlyMG | Endo- | Stenotrophomonas maltophilia KJ-2 | - | [22] |
| 7 | Pa1167 | Endo- | Pseudomonas aeruginosa PAO1 | Coastal marine habitats | [23] |
| 8 | Vpa_0049 | Endo- | Vibrio sp. QY108 | Marine | [14] |
| 14 | vAL-1 | Endo- | Halitos discus hannai | Abalone | [24] |
| Chlorella virus ATCV-1 | Heliozoon A. turfacea | [25] | |||
| 15 | Atu3025 | Exo- | Sphingomonas sp. A1 | Soil | [11] |
| 17 | Alg17C | Exo- | Saccharophagus degradans | Marine | [26] |
| 18 | Aly-SJ02 | Endo- | Pseudoalteromonas sp. SM0524 | Marine rotten kelp | [13] |
| 36 | Aly36B | Endo- | Chitinophaga sp. MD30 | Air conditioner condensate pipe | [15] |
| 39 | Dp0100 | Endo- | Defluviitalea phaphyphila | Coastal sediment | [16] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, W.; Zhang, X.; Hou, Z.; Hu, X.; Wang, Y.; Wang, C.; Yang, S.; Cui, H.; Zhu, L. Microbial Regulation of Deterioration and Preservation of Salted Kelp under Different Temperature and Salinity Conditions. Foods 2021, 10, 1723. https://doi.org/10.3390/foods10081723
Wei W, Zhang X, Hou Z, Hu X, Wang Y, Wang C, Yang S, Cui H, Zhu L. Microbial Regulation of Deterioration and Preservation of Salted Kelp under Different Temperature and Salinity Conditions. Foods. 2021; 10(8):1723. https://doi.org/10.3390/foods10081723
Chicago/Turabian StyleWei, Wei, Xin Zhang, Zhaozhi Hou, Xinyu Hu, Yuan Wang, Caizheng Wang, Shujing Yang, Henglin Cui, and Lin Zhu. 2021. "Microbial Regulation of Deterioration and Preservation of Salted Kelp under Different Temperature and Salinity Conditions" Foods 10, no. 8: 1723. https://doi.org/10.3390/foods10081723
APA StyleWei, W., Zhang, X., Hou, Z., Hu, X., Wang, Y., Wang, C., Yang, S., Cui, H., & Zhu, L. (2021). Microbial Regulation of Deterioration and Preservation of Salted Kelp under Different Temperature and Salinity Conditions. Foods, 10(8), 1723. https://doi.org/10.3390/foods10081723