Nutritional and Rheological Features of Lentil Protein Isolate for Yoghurt-Like Application



Abstract

1. Introduction
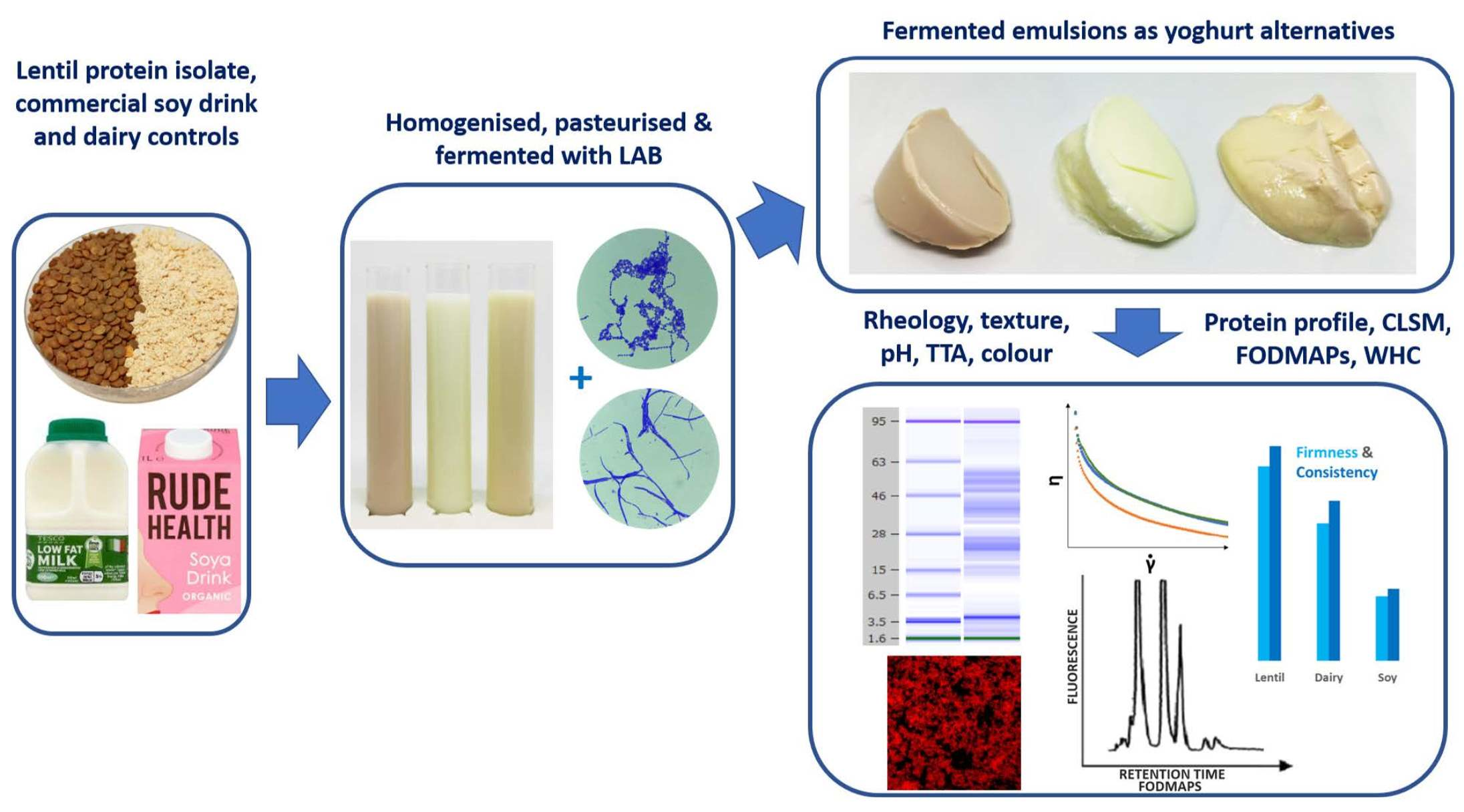
2. Materials and Methods
2.1. Materials
2.2. Preparation of Set Yoghurts
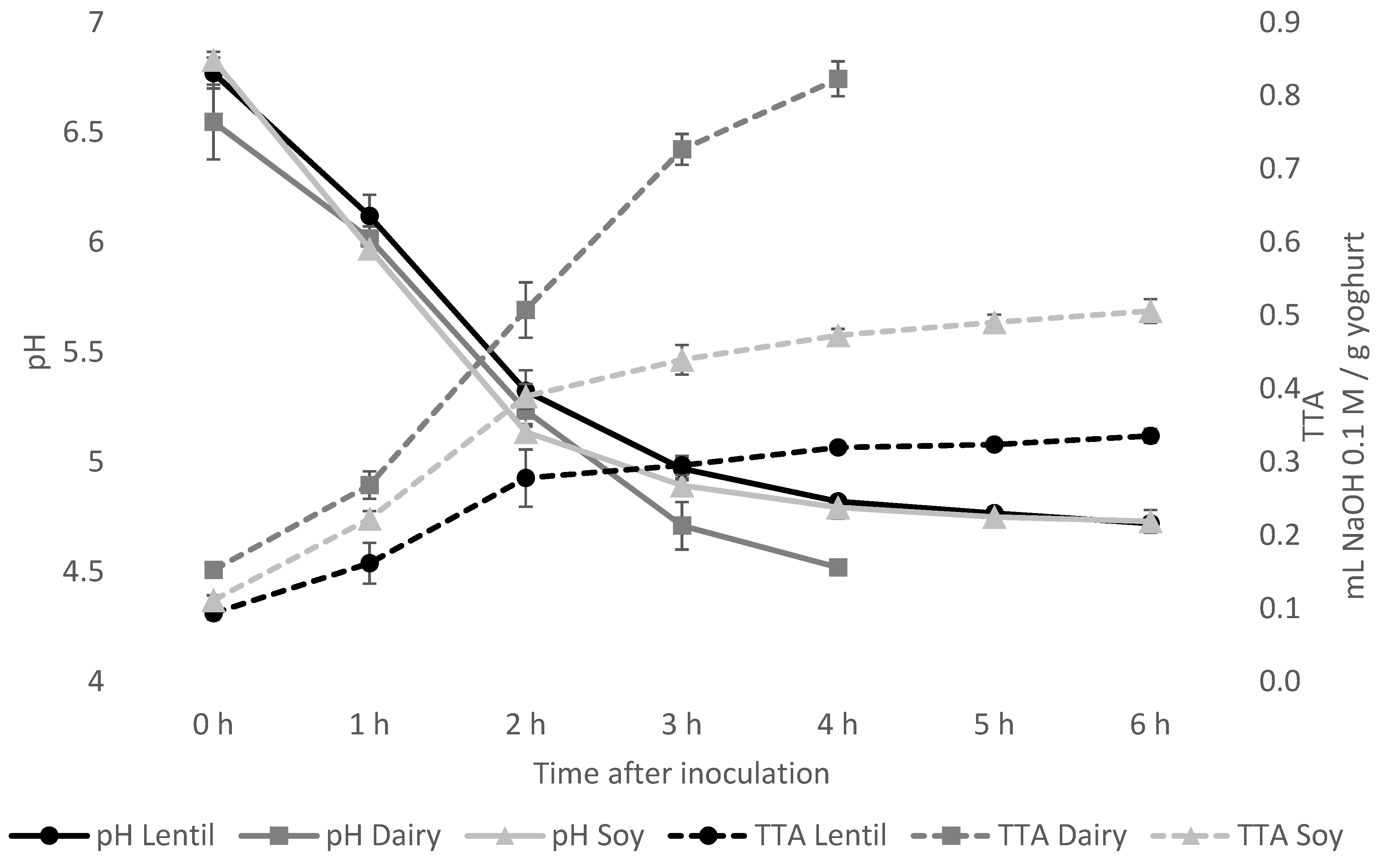
2.3. Fermentation Monitoring by pH and Total Titratable Acidity (TTA) Measurement
2.4. Viable Cell Count
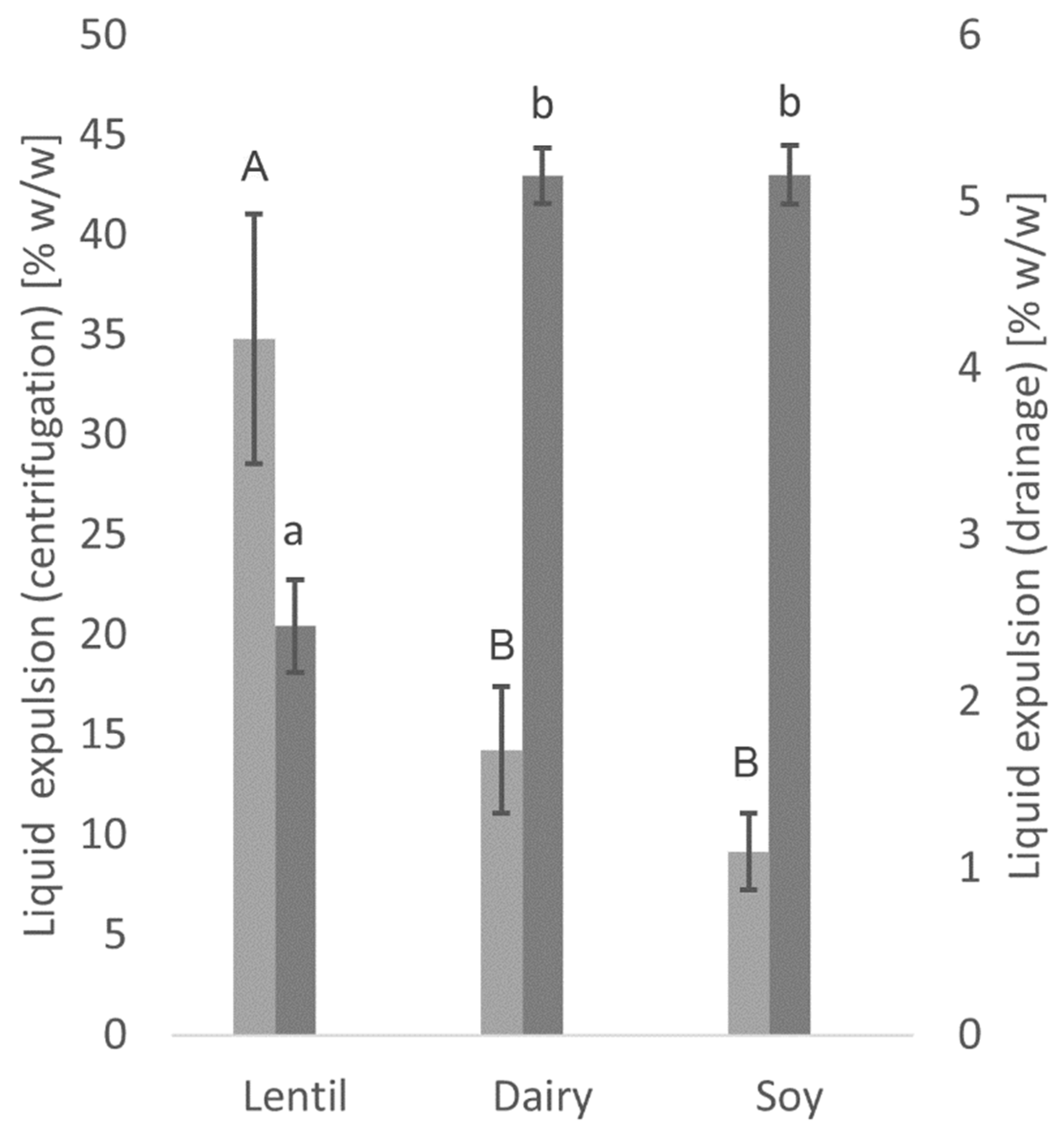
2.5. Water Holding Capacity
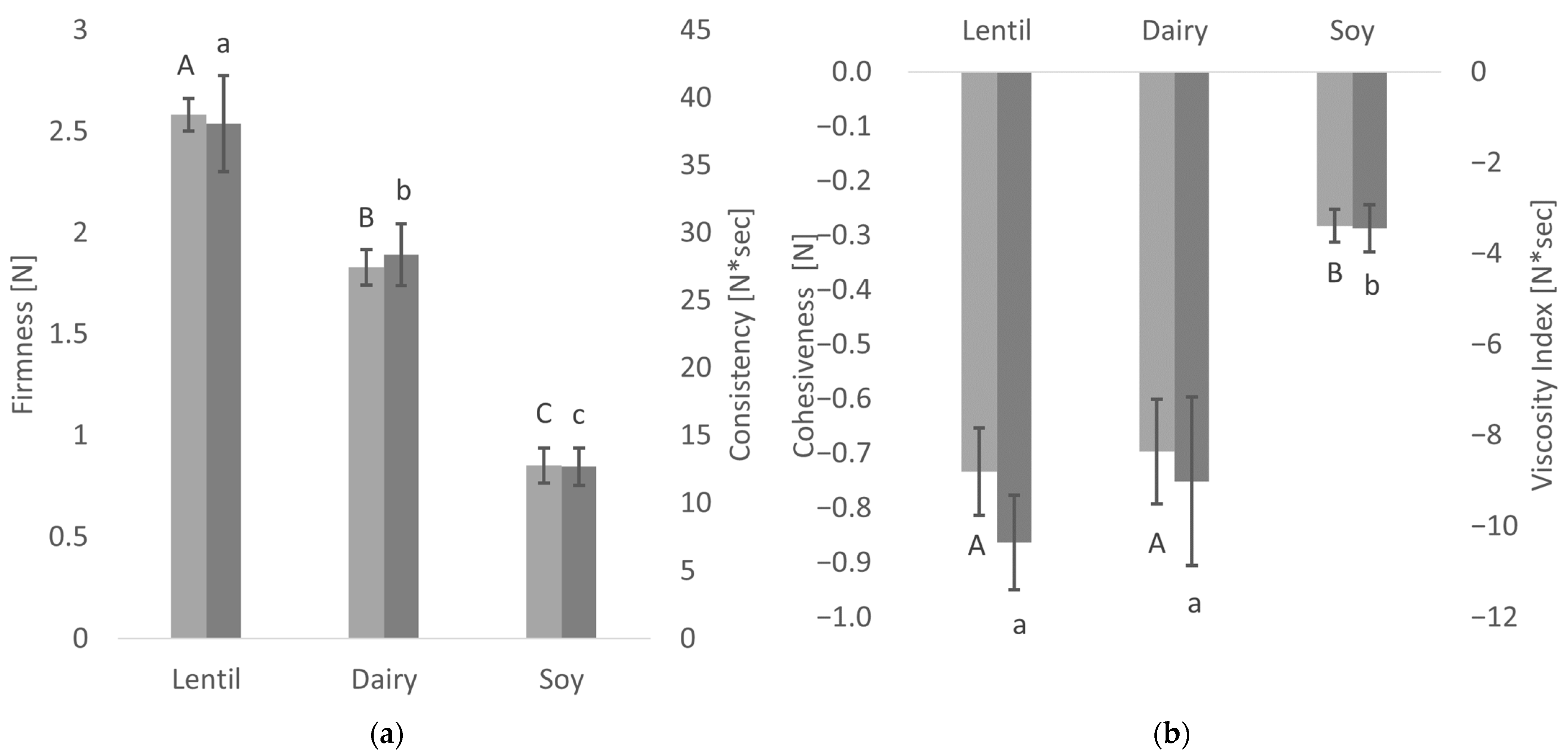
2.6. Uniaxial Compression Testing
2.7. Rheological Measurements
2.8. Confocal Laser Scanning Microscopy
2.9. Colour Measurements
2.10. Quantification of Sugars, Organic Acids and FODMAPs
2.11. Protein Profile Analysis
2.12. Statistical Data Analysis
3. Results
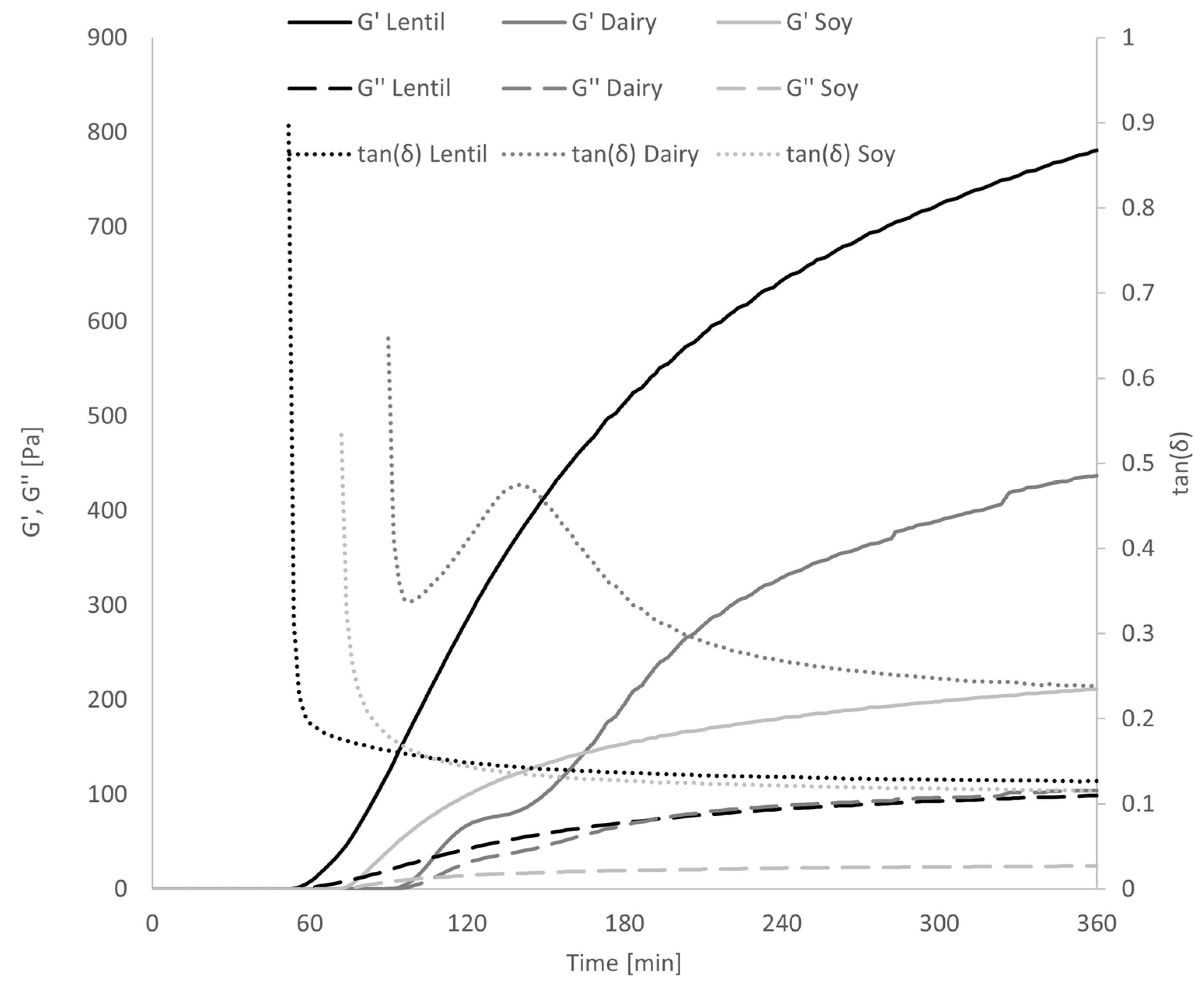
3.1. Fermentation Characteristics
3.2. Water Holding Capacity
3.3. Textural Properties
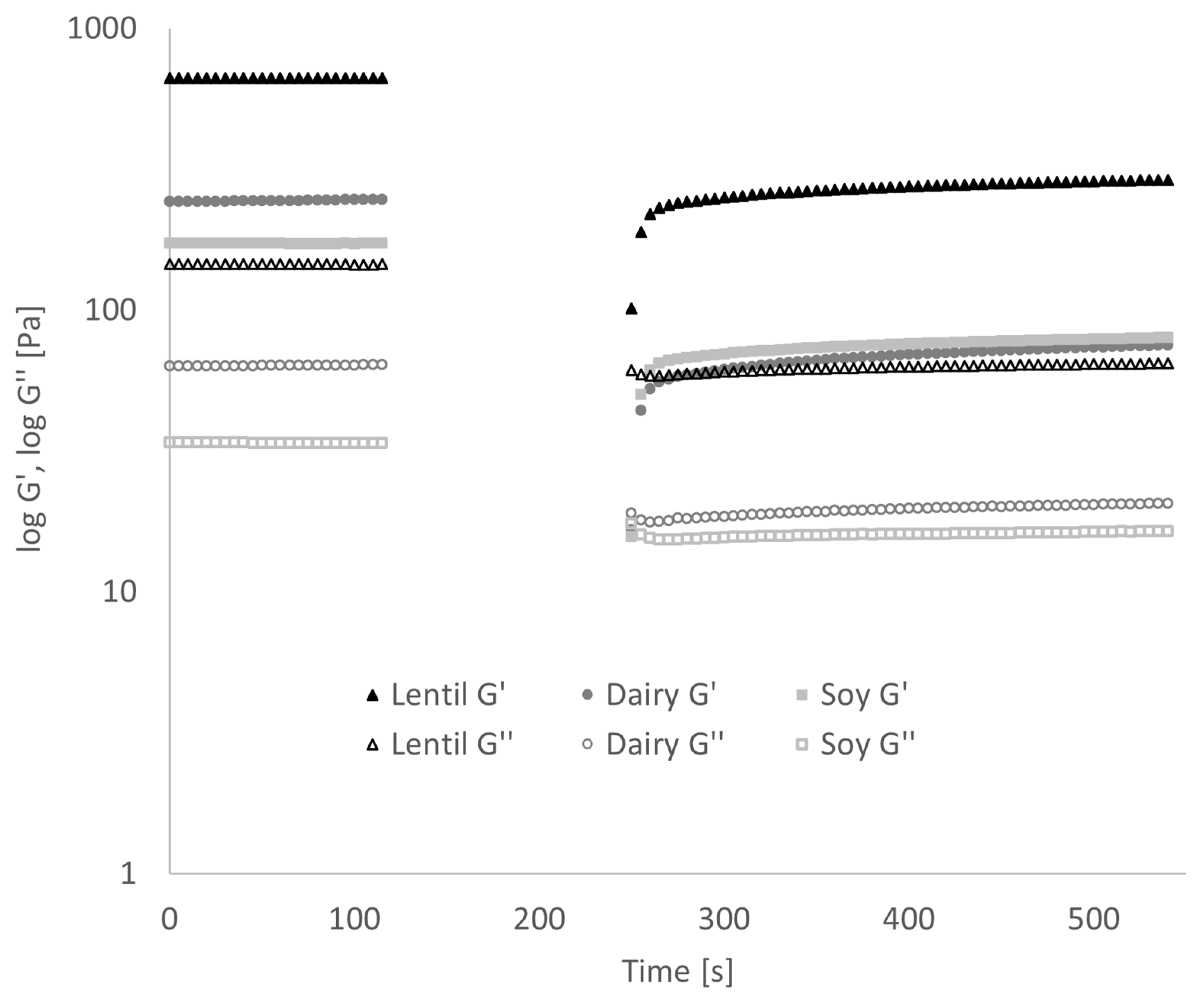
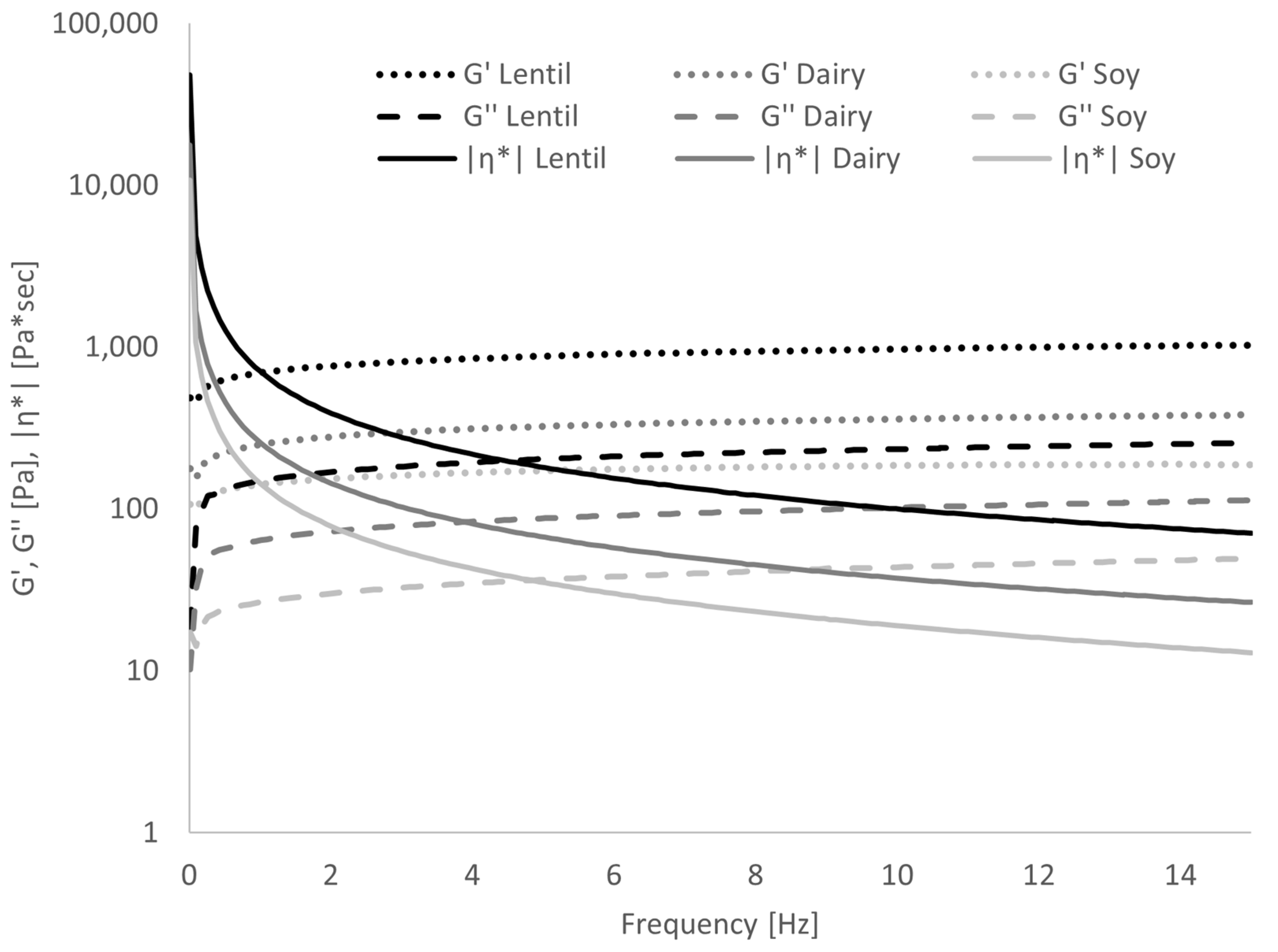
3.4. Gel Behaviour of Fermented Samples
3.5. Microscopic Protein Structure
3.6. Colour Results
3.7. FODMAP Content
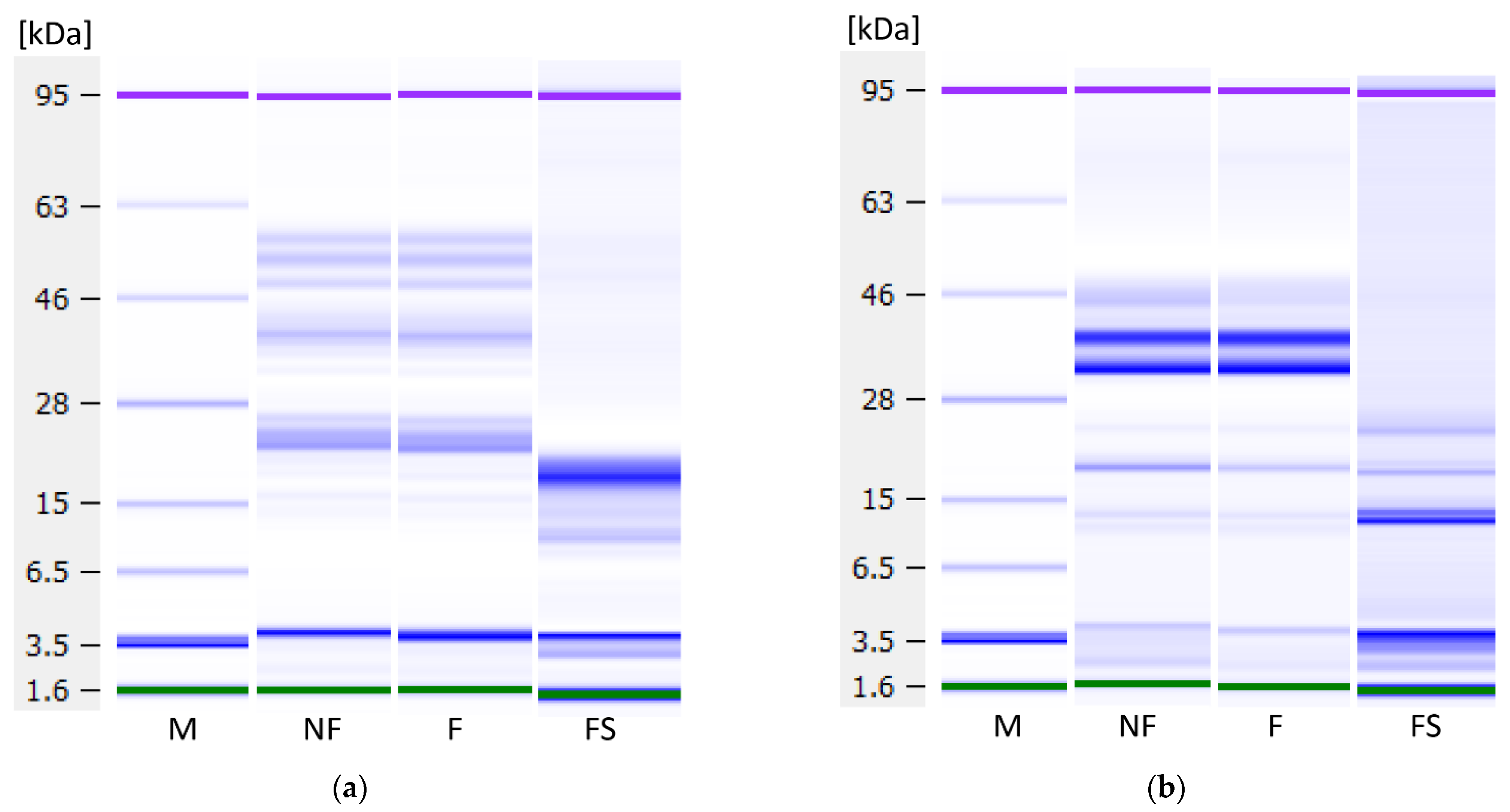
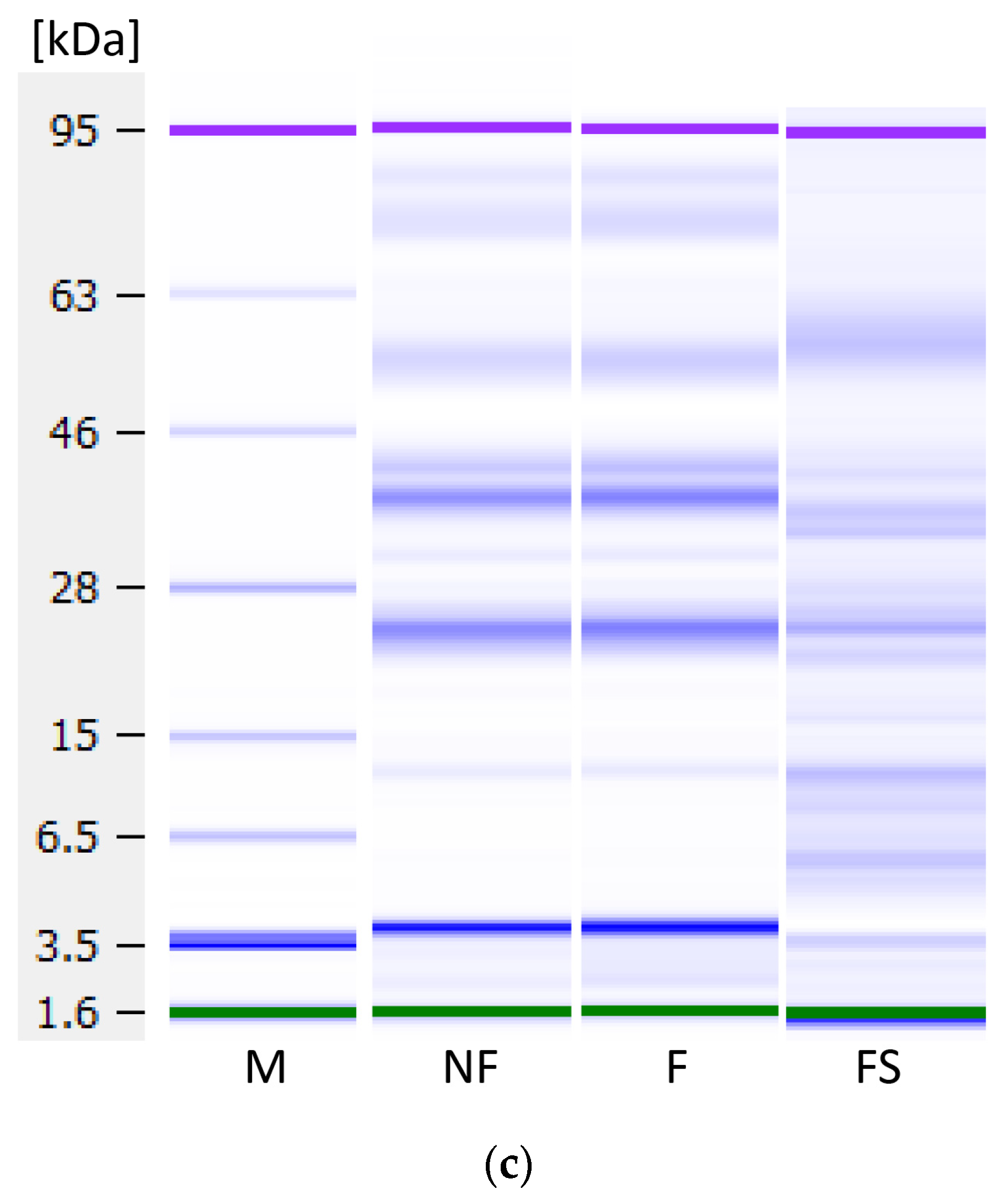
3.8. Protein Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vermeulen, S.J.; Campbell, B.M.; Ingram, J.S.I. Climate change and food systems. Annu. Rev. Environ. Resour. 2012, 37, 195–222. [Google Scholar] [CrossRef]
- Considine, M.J.; Siddique, K.H.M.; Foyer, C.H. Nature’s pulse power: Legumes, food security and climate change. J. Exp. Bot. 2017, 68, 1815–1818. [Google Scholar] [CrossRef]
- Clune, S.; Crossin, E.; Verghese, K. Systematic review of greenhouse gas emissions for different fresh food categories. J. Clean. Prod. 2017, 140, 766–783. [Google Scholar] [CrossRef]
- Davis, K.F.; Gephart, J.A.; Emery, K.A.; Leach, A.M.; Galloway, J.N.; D’Odorico, P. Meeting future food demand with current agricultural resources. Glob. Environ. Chang. 2016, 39, 125–132. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lam, H.M.; Nguyen, H.T.; Siddique, K.H.M.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M.; et al. Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef]
- Afshin, A.; Micha, R.; Khatibzadeh, S.; Mozaffarian, D. Consumption of nuts and legumes and risk of incident ischemic heart disease, stroke, and diabetes: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2014, 100, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Kushi, L.H.; Meyer, K.A.; Jacobs, D.R. Cereals, legumes, and chronic disease risk reduction: Evidence from epidemiologic studies. Am. J. Clin. Nutr. 1999, 70. [Google Scholar] [CrossRef]
- Bernstein, A.M.; Sun, Q.; Hu, F.B.; Stampfer, M.J.; Manson, J.E.; Willett, W.C. Major dietary protein sources and risk of coronary heart disease in women. Circulation 2010, 122, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Johansen, C.; Turner, N.C.; Jeuffroy, M.H.; Hashem, A.; Sakar, D.; Gan, Y.; Alghamdi, S.S. Innovations in agronomy for food legumes. A review. Agron. Sustain. Dev. 2012, 32, 45–64. [Google Scholar] [CrossRef]
- ICRISAT; CIAT; ICARDA; IITA. Grain Legumes—Leveraging Legumes to Combat Poverty, Hunger, Malnutrition and Environmental Degradation. CGIAR Research Program 3.5. 2012. Available online: https://hdl.handle.net/10947/5316 (accessed on 19 January 2021).
- Peltonen-Sainio, P.; Jauhiainen, L.; Hakala, K.; Ojanen, H. Climate change and prolongation of growing season: Changes in regional potential for field crop production in Finland. Agric. Food Sci. 2009, 18, 171–190. [Google Scholar] [CrossRef]
- Ramirez-Cabral, N.Y.Z.; Kumar, L.; Taylor, S. Crop niche modeling projects major shifts in common bean growing areas. Agric. For. Meteorol. 2016, 218–219, 102–113. [Google Scholar] [CrossRef]
- O’Toole, D.K. Soybean: Soymilk, Tofu, and Okara, 2nd ed.; Elsevier Ltd.: Oxford, UK, 2015; ISBN 9780123947864. [Google Scholar]
- Mäkinen, O.E.; Wanhalinna, V.; Zannini, E.; Arendt, E.K. Foods for special dietary needs: Non-dairy plant-based milk substitutes and fermented dairy-type products. Crit. Rev. Food Sci. Nutr. 2016, 56, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Schiano, A.N.; Harwood, W.S.; Gerard, P.D.; Drake, M.A. Consumer perception of the sustainability of dairy products and plant-based dairy alternatives. J. Dairy Sci. 2020, 103, 11228–11243. [Google Scholar] [CrossRef]
- ProVeg Plant-Based Foods in Europe: How Big Is the Market? Consumer Study on Plant-Based Food Sales in Europe, Performed by Nielsen MarketTrack. Available online: https://proveg.com/what-we-do/corporate-engagement/plant-based-food-sector-report/ (accessed on 16 April 2021).
- Grasso, N.; Alonso-Miravalles, L.; Mahony, J.A.O. Composition, physicochemical and sensorial propertes of commercial plant-based yogurts. Foods 2020, 9, 252. [Google Scholar] [CrossRef]
- Paul, A.A.; Kumar, S.; Kumar, V.; Sharma, R. Milk Analog: Plant based alternatives to conventional milk, production, potential and health concerns. Crit. Rev. Food Sci. Nutr. 2019, 60, 3005–3023. [Google Scholar] [CrossRef]
- Jeske, S.; Zannini, E.; Arendt, E.K. Evaluation of physicochemical and glycaemic properties of commercial plant-based milk substitutes. Plant Foods Hum. Nutr. 2017, 72, 26–33. [Google Scholar] [CrossRef]
- European Parliament & Council. Regulation (EC) 1924/2006 on Nutrition and Health Claims Made on Foods; European Parliament & Council: Brussels, Belgium, 2006; pp. 1–31. [Google Scholar]
- Chalupa-Krebzdak, S.; Long, C.J.; Bohrer, B.M. Nutrient density and nutritional value of milk and plant-based milk alternatives. Int. Dairy J. 2018, 87, 84–92. [Google Scholar] [CrossRef]
- Boeck, T.; Sahin, A.W.; Zannini, E.; Arendt, E.K. Nutritional properties and health aspects of pulses and their use in plant-based yogurt alternatives. Compr. Rev. Food Sci. Food Saf. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zuidmeer, L.; Goldhahn, K.; Rona, R.J.; Gislason, D.; Madsen, C.; Summers, C.; Sodergren, E.; Dahlstrom, J.; Lindner, T.; Sigurdardottir, S.T.; et al. The prevalence of plant food allergies: A systematic review. J. Allergy Clin. Immunol. 2008, 121. [Google Scholar] [CrossRef]
- Katz, Y.; Gutierrez-Castrellon, P.; González, M.G.; Rivas, R.; Lee, B.W.; Alarcon, P. A comprehensive review of sensitization and allergy to soy-based products. Clin. Rev. Allergy Immunol. 2014, 46, 272–281. [Google Scholar] [CrossRef]
- Hickisch, A.; Beer, R.; Vogel, R.F.; Toelstede, S. Influence of lupin-based milk alternative heat treatment and exopolysaccharide-producing lactic acid bacteria on the physical characteristics of lupin-based yogurt alternatives. Food Res. Int. 2016, 84, 180–188. [Google Scholar] [CrossRef]
- Lim, X.X.; Koh, W.Y.; Uthumporn, U.; Maizura, M.; Wan Rosli, W.I. The development of legume-based yogurt by using water kefir as starter culture. Int. Food Res. J. 2019, 26, 1219–1228. [Google Scholar]
- Jiang, Z.Q.; Wang, J.; Stoddard, F.; Salovaara, H.; Sontag-Strohm, T. Preparation and characterization of emulsion gels from whole faba bean flour. Foods 2020, 9, 755. [Google Scholar] [CrossRef]
- Jeske, S.; Bez, J.; Arendt, E.K.; Zannini, E. Formation, stability, and sensory characteristics of a lentil-based milk substitute as affected by homogenisation and pasteurisation. Eur. Food Res. Technol. 2019, 245, 1519–1531. [Google Scholar] [CrossRef]
- El Youssef, C.; Bonnarme, P.; Fraud, S.; Péron, A.C.; Helinck, S.; Landaud, S. Sensory improvement of a pea protein-based product using microbial co-cultures of lactic acid bacteria and yeasts. Foods 2020, 9, 349. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [PubMed]
- Damin, M.R.; Minowa, E.; Alcantara, M.R.; Oliveira, M.N. Effect of cold storage on culture viability and some rheological properties of fermented milk prepared with yogurt and probiotic bacteria. J. Text. Stud. 2008, 39, 40–55. [Google Scholar] [CrossRef]
- Alonso-Miravalles, L.; Jeske, S.; Bez, J.; Detzel, A.; Busch, M.; Krueger, M.; Wriessnegger, C.L.; O’Mahony, J.A.; Zannini, E.; Arendt, E.K. Membrane filtration and isoelectric precipitation technological approaches for the preparation of novel, functional and sustainable protein isolate from lentils. Eur. Food Res. Technol. 2019, 245, 1855–1869. [Google Scholar] [CrossRef]
- Zannini, E.; Jeske, S.; Lynch, K.; Arendt, E.K. Development of novel quinoa-based yoghurt fermented with dextran producer Weissella cibaria MG1. Int. J. Food Microbiol. 2018, 268, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Lucey, J.A. Formation and physical properties of yogurt. Asian Australas. J. Anim. Sci. 2010, 23, 1127–1136. [Google Scholar] [CrossRef]
- Brückner-Gühmann, M.; Benthin, A.; Drusch, S. Enrichment of yoghurt with oat protein fractions: Structure formation, textural properties and sensory evaluation. Food Hydrocoll. 2019, 86, 146–153. [Google Scholar] [CrossRef]
- Harwlakar, V.R.; Kalab, M. Susceptibility of yoghurt to syneresis. Comparison of centrifugation and drainage methods. Milchwissenschaft 1983, 38, 517–522. [Google Scholar]
- Silva, J.V.C.; O’Mahony, J.A. Microparticulated whey protein addition modulates rheological and microstructural properties of high-protein acid milk gels. Int. Dairy J. 2018, 78, 145–151. [Google Scholar] [CrossRef]
- Klost, M.; Drusch, S. Structure formation and rheological properties of pea protein-based gels. Food Hydrocoll. 2019, 94, 622–630. [Google Scholar] [CrossRef]
- Hickisch, A.; Bindl, K.; Vogel, R.F.; Toelstede, S. Thermal treatment of lupin-based milk alternatives—Impact on lupin proteins and the network of respective lupin-based yogurt alternatives. Food Res. Int. 2016, 89, 850–859. [Google Scholar] [CrossRef]
- Ispiryan, L.; Heitmann, M.; Hoehnel, A.; Zannini, E.; Arendt, E.K. Optimization and Validation of an HPAEC-PAD Method for the Quantification of FODMAPs in Cereals and Cereal-Based Products. J. Agric. Food Chem. 2019, 67, 4384–4392. [Google Scholar] [CrossRef] [PubMed]
- Ispiryan, L.; Zannini, E.; Arendt, E.K. Characterization of the FODMAP-profile in cereal-product ingredients. J. Cereal Sci. 2020, 92, 102916. [Google Scholar] [CrossRef]
- Dwyer, M.V.-O.; Bez, J.; Petersen, I.L.; Joehnke, M.S.; Detzel, A.; Busch, M.; Krueger, M.; Ispiryan, L.; O’Mahony, J.A.; Arendt, E.K.; et al. Techno-functional, nutritional, and environmental performance of protein isolates from blue lupin and white lupin. Foods 2019, 9, 230. [Google Scholar] [CrossRef]
- Joehnke, M.S.; Jeske, S.; Ispiryan, L.; Zannini, E.; Arendt, E.K.; Bez, J.; Sørensen, J.C.; Petersen, I.L. Nutritional and anti-nutritional properties of lentil (Lens culinaris) protein isolates prepared by pilot-scale processing. Food Chem. X 2020, 100112. [Google Scholar] [CrossRef]
- Steffe, J.F. Rheological Methods in Food Process Engineering; Freeman Press: East Lansing, MI, USA, 1996; ISBN 0963203614. [Google Scholar]
- Lee, S.-Y.; Yeo, Y.-S.; Park, S.-Y.; Lee, S.-G.; Lee, S.-M.; Cho, H.-S.; Chung, N.-J.; Oh, S.-W. Compositional Analysis of Lentil (Lens culinaris) Cultivars Related to Colors and Their Antioxidative Activity. Plant Breed. Biotechnol. 2017, 5, 192–203. [Google Scholar] [CrossRef]
- Ladjal-Ettoumi, Y.; Boudries, H.; Chibane, M.; Romero, A. Pea, Chickpea and Lentil Protein Isolates: Physicochemical Characterization and Emulsifying Properties. Food Biophys. 2016, 11, 43–51. [Google Scholar] [CrossRef]
- Sparvoli, F.; Bollini, R.; Cominelli, E. Nutritional Value. In Grain Legumes; De Ron, A.M., Ed.; Springer: Pontevedra, Spain, 2015; pp. 291–325. ISBN 9781493927975. [Google Scholar]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Swanson, B.G. Pea and lentil protein extraction and functionality. J. Am. Oil Chem. Soc. 1990, 67, 276–280. [Google Scholar] [CrossRef]
- Costa, F.F.; Brito, M.A.V.P.; Furtado, M.A.M.; Martins, M.F.; De Oliveira, M.A.L.; De Castro Barra, P.M.; Garrido, L.A.; De Oliveira Dos Santos, A.S. Microfluidic chip electrophoresis investigation of major milk proteins: Study of buffer effects and quantitative approaching. Anal. Methods 2014, 6, 1666–1673. [Google Scholar] [CrossRef]
- Nishinari, K.; Fang, Y.; Guo, S.; Phillips, G.O. Soy proteins: A review on composition, aggregation and emulsification. Food Hydrocoll. 2014, 39, 301–318. [Google Scholar] [CrossRef]
- Bashi, Z.; McCullough, R.; Ong, L.; Ramirez, M. Alternative Proteins: The Race for Market Share Is On; McKinsey & Company: Denver, CO, USA, 2019; pp. 1–11. [Google Scholar]
- Carr, F.J.; Chill, D.; Maida, N. The lactic acid bacteria: A literature survey. Crit. Rev. Microbiol. 2002, 28, 281–370. [Google Scholar] [CrossRef]
- Angioloni, A.; Collar, C. Small and large deformation viscoelastic behaviour of selected fibre blends with gelling properties. Food Hydrocoll. 2009, 23, 742–748. [Google Scholar] [CrossRef]
- Malhotra, A.; Coupland, J.N. The effect of surfactants on the solubility, zeta potential, and viscosity of soy protein isolates. Food Hydrocoll. 2004, 18, 101–108. [Google Scholar] [CrossRef]
- Bi, C.H.; Li, D.; Wang, L.J.; Adhikari, B. Viscoelastic properties and fractal analysis of acid-induced SPI gels at different ionic strength. Carbohydr. Polym. 2013, 92, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Staudacher, H.M.; Whelan, K.; Irving, P.M.; Lomer, M.C.E. Comparison of symptom response following advice for a diet low in fermentable carbohydrates (FODMAPs) versus standard dietary advice in patients with irritable bowel syndrome. J. Hum. Nutr. Diet. 2011, 24, 487–495. [Google Scholar] [CrossRef]
- Varney, J.; Barrett, J.; Scarlata, K.; Catsos, P.; Gibson, P.R.; Muir, J.G. FODMAPs: Food composition, defining cutoff values and international application. J. Gastroenterol. Hepatol. 2017, 32, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Bartkiene, E.; Sakiene, V.; Bartkevics, V.; Rusko, J.; Lele, V.; Juodeikiene, G.; Wiacek, C.; Braun, P.G. Lupinus angustifolius L. lactofermentation and protein isolation: Effects on phenolic compounds and genistein, antioxidant properties, trypsin inhibitor activity, and protein digestibility. Eur. Food Res. Technol. 2018, 244, 1521–1531. [Google Scholar] [CrossRef]
- Coda, R.; Melama, L.; Rizzello, C.G.; Curiel, J.A.; Sibakov, J.; Holopainen, U.; Pulkkinen, M.; Sozer, N. Effect of air classification and fermentation by Lactobacillus plantarum VTT E-133328 on faba bean (Vicia faba L.) flour nutritional properties. Int. J. Food Microbiol. 2015, 193, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Lucey, J.A.; Munro, P.A.; Singh, H. Whey separation in acid skim milk gels made with glucono-delta-lactone: Effects of heat treatment and gelation temperature. J. Texture Stud. 1998, 29, 413–426. [Google Scholar] [CrossRef]
- Zulkurnain, M.; Goh, M.H.; Karim, A.A.; Liong, M.T. Development of a soy-based cream cheese. J. Texture Stud. 2008, 39, 635–654. [Google Scholar] [CrossRef]
- Li, Q.; Xia, Y.; Zhou, L.; Xie, J. Evaluation of the rheological, textural, microstructural and sensory properties of soy cheese spreads. Food Bioprod. Process. 2013, 91, 429–439. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lactic Acid | Sucrose | CFU/mL | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| [mg/100 g] | [mg/100 g] | |||||||||
| Non-fermented | Lentil | n.d. | 780 | ± | 32 A | 1.36 × 107 | ± | 5.64 × 106 A | ||
| Dairy | n.d. | 748 | ± | 144 A | 2.65 × 107 | ± | 1.61 × 106 A | |||
| Soy | n.d. | 704 | ± | 8 A | 2.12 × 107 | ± | 1.45 × 107 A | |||
| Fermented | Lentil | 244 | ± | 8 a | 483 | ± | 24 a | 3.54 × 107 | ± | 2.98 × 107 a |
| Dairy | 603 | ± | 16 b | n.d. | 7.09 × 108 | ± | 6.50 × 108 a | |||
| Soy | 339 | ± | 9 c | 328 | ± | 5 b | 3.02 × 107 | ± | 2.98 × 107 a | |
| ΔG′ [%] | ΔG″ [%] | Dlog G′/Dlog ω | Dlog G″/Dlog ω | Dlog|η*|/Dlogω | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lentil | 55.30 | ± | 3.57 a | 54.97 | ± | 1.95 a | 0.147 | ± | 0.003 a | 0.208 | ± | 0.007 ab | −0.850 | ± | 0.003 a |
| Dairy | 69.63 | ± | 3.48 b | 67.84 | ± | 3.63 b | 0.164 | ± | 0.019 a | 0.204 | ± | 0.009 a | −0.831 | ± | 0.018 a |
| Soy | 53.73 | ± | 0.63 a | 51.44 | ± | 0.56 a | 0.113 | ± | 0.003 b | 0.221 | ± | 0.003 b | −0.882 | ± | 0.003 b |
| L* | a* | b* | WI | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Non-fermented | Lentil | 81.21 | ± | 0.24 A | 1.36 | ± | 0.22 A | 6.30 | ± | 0.17 A | 80.14 | ± | 0.23 A |
| Dairy | 93.83 | ± | 0.02 B | −4.70 | ± | 0.64 B | 6.93 | ± | 0.01 B | 89.59 | ± | 0.30 B | |
| Soy | 88.74 | ± | 0.19 C | 1.61 | ± | 0.11 C | 13.44 | ± | 0.02 C | 82.40 | ± | 0.15 A | |
| Fermented | Lentil | 87.90 | ± | 0.24 a | 1.65 | ± | 0.28 a | 7.57 | ± | 0.28 a | 85.63 | ± | 0.25 a |
| Dairy | 96.20 | ± | 0.10 b | −3.79 | ± | 0.10 b | 9.35 | ± | 0.43 b | 89.22 | ± | 0.37 b | |
| Soy | 92.88 | ± | 0.03 c | −0.77 | ± | 0.07 c | 13.26 | ± | 0.20 c | 84.93 | ± | 0.17 c | |
| Lactose | Glucose | Fructose | EF | ||
|---|---|---|---|---|---|
| Non-fermented | Lentil | n.d. | 6 ± 1 A | n.d. | - |
| Dairy | 5257 ± 148 | n.d. | n.d. | - | |
| Soy | n.d. | 7 ± 0 A | n.d. | - | |
| Fermented | Lentil | n.d. | 19 ± 4 ab | 26 ± 7 ab | 8 ± 4 a |
| Dairy | 4807 ± 23 | 23 ± 10 a | 40 ± 10 a | 17 ± 20 a | |
| Soy | n.d. | 10 ± 0 b | 15 ± 1 b | 5 ± 1 a | |
| Polyols | |||||
| Xylitol | Sorbitol | Mannitol | ƩPolyols | ||
| Non-fermented | Lentil | 9 ± 0 | 22 ± 10 A | n.d. | 30 ± 11 A |
| Dairy | n.d. | n.d. | n.d. | n.d. | |
| Soy | n.d. | 7 ± 2 B | n.d. | 11 ± 2 B | |
| Fermented | Lentil | n.d. | n.d. | n.d. | n.d. |
| Dairy | n.d. | n.d. | n.d. | n.d. | |
| Soy | n.d. | 9 ± 0 | n.d. | 13 ± 0 | |
| GOS | |||||
| Raffinose | Stachyose | ||||
| Non-fermented | Lentil | n.d. | 62 ± 2 A | ||
| Dairy | n.d. | n.d. | |||
| Soy | 102 ± 2 | 310 ± 8 B | |||
| Fermented | Lentil | n.d. | 53 ± 2 a | ||
| Dairy | n.d. | n.d. | |||
| Soy | 89 ± 28 | 293 ± 2 b | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boeck, T.; Zannini, E.; Sahin, A.W.; Bez, J.; Arendt, E.K. Nutritional and Rheological Features of Lentil Protein Isolate for Yoghurt-Like Application. Foods 2021, 10, 1692. https://doi.org/10.3390/foods10081692
Boeck T, Zannini E, Sahin AW, Bez J, Arendt EK. Nutritional and Rheological Features of Lentil Protein Isolate for Yoghurt-Like Application. Foods. 2021; 10(8):1692. https://doi.org/10.3390/foods10081692
Chicago/Turabian StyleBoeck, Theresa, Emanuele Zannini, Aylin W. Sahin, Juergen Bez, and Elke K. Arendt. 2021. "Nutritional and Rheological Features of Lentil Protein Isolate for Yoghurt-Like Application" Foods 10, no. 8: 1692. https://doi.org/10.3390/foods10081692
APA StyleBoeck, T., Zannini, E., Sahin, A. W., Bez, J., & Arendt, E. K. (2021). Nutritional and Rheological Features of Lentil Protein Isolate for Yoghurt-Like Application. Foods, 10(8), 1692. https://doi.org/10.3390/foods10081692