
Improvement of Must Fermentation from Late Harvest cv. Tempranillo Grapes Treated with Pulsed Light

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Pulsed Light Treatment
2.2. Atomic Force Microscopy
2.3. Yeast Strains and Microfermentations
2.4. Oenological Parameters
2.5. Aromatic Volatile Compounds
2.6. Anthocyanins Quantification
2.7. Statistical Analysis
3. Results
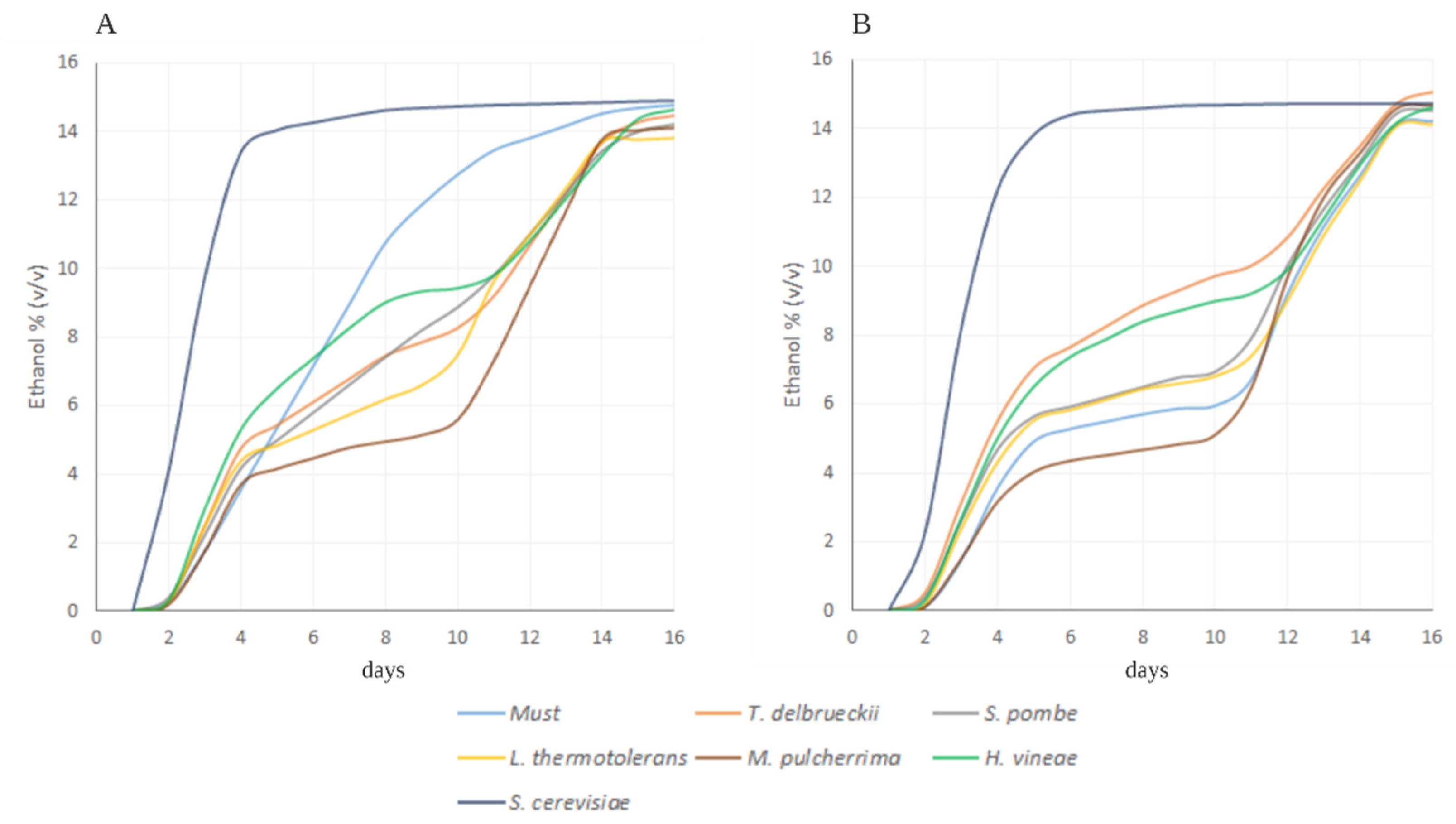
3.1. Microfermentations
3.2. Oenological Parameters
3.3. Volatile Fraction and Phenolic Profile
4. Discussion
4.1. Microfermentations
4.2. Oenological Parameters
4.3. Volatile Fraction and Anthocyanin Profile
4.3.1. Aromatic Volatile’s Fraction
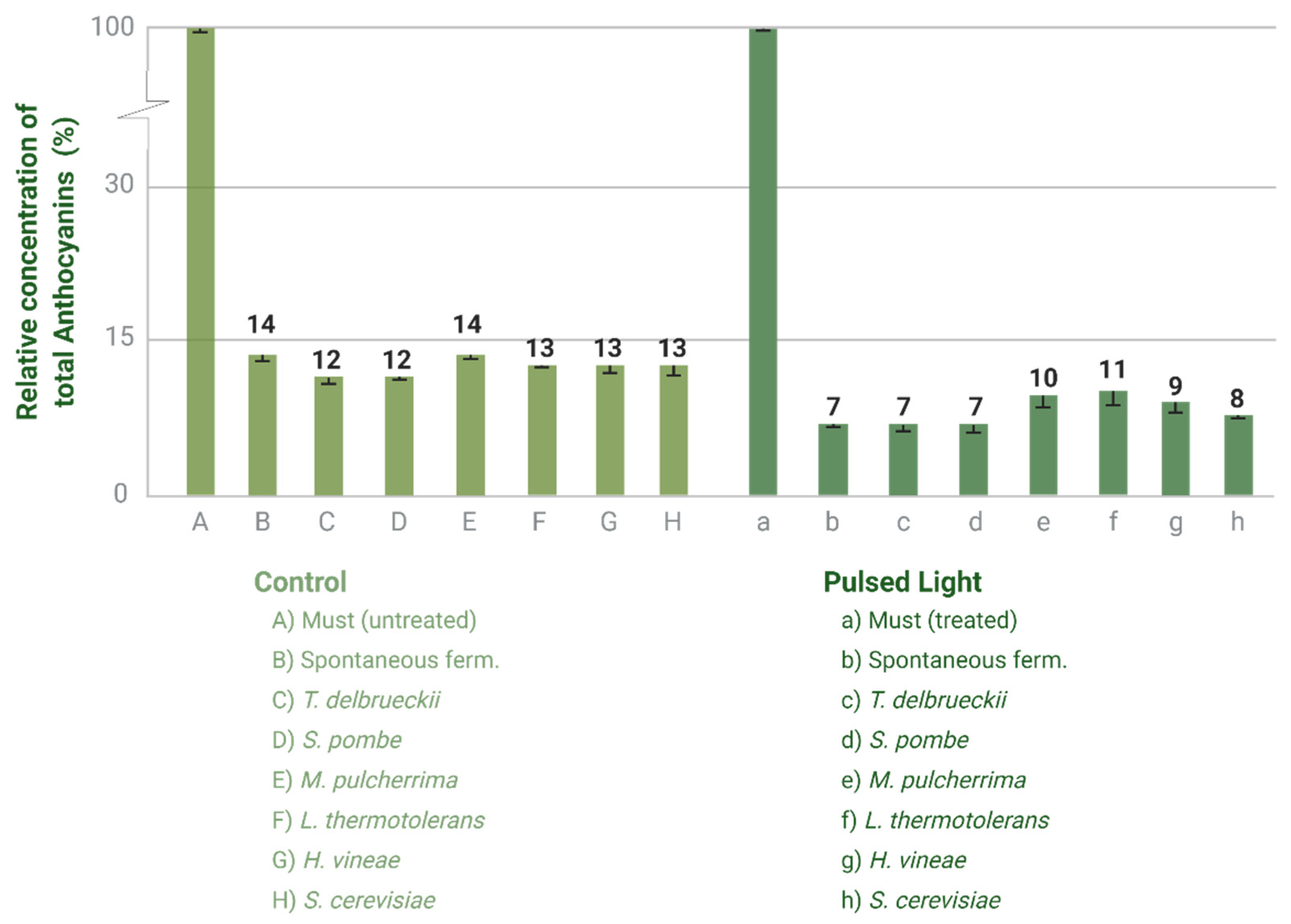
4.3.2. Anthocyanin’s Profile
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacGregor, S.J.; Rowan, N.J.; McIlvaney, L.; Anderson, J.G.; Fouracre, R.A.; Farish, O. Light inactivation of food-related pathogenic bacteria using a pulsed power source. Lett. Appl. Microbiol. 1998, 27, 67–70. [Google Scholar] [CrossRef]
- Rowan, N.J.; MacGregor, S.J.; Anderson, J.G.; Fouracre, R.A.; McIlvaney, L.; Farish, O. Pulsed-Light Inactivation of Food-Related Microorganisms. Appl. Environ. Microbiol. 1999, 65, 1312–1315. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J. Pulsed light and pulsed electric field for foods and eggs. Poult. Sci. 1996, 75, 1133–1136. [Google Scholar] [CrossRef]
- Anderson, J.G.; Rowan, N.J.; MacGregor, S.J.; Fouracre, R.A.; Parish, O. Inactivation of food-borne enteropathogenic bacteria and spoilage fungi using pulsed-light. IEEE Trans. Plasma Sci. 2000, 28, 83–88. [Google Scholar] [CrossRef]
- Kramer, B.; Wunderlich, J.; Muranyi, P. Recent findings in pulsed light disinfection. J. Appl. Microbiol. 2017, 122, 830–856. [Google Scholar] [CrossRef]
- Nannelli, F.; Claisse, O.; Gindreau, E.; De Revel, G.; Lonvaud-Funel, A.; Lucas, P.M. Determination of lactic acid bacteria producing biogenic amines in wine by quantitative PCR methods. Lett. Appl. Microbiol. 2008, 47, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Elmnasser, N.; Guillou, S.; Leroi, F.; Orange, N.; Bakhrouf, A.; Federighi, M. Pulsed-light system as a novel food decontamination technology: A review. Can. J. Microbiol. 2007, 53, 813–821. [Google Scholar] [CrossRef]
- Gómez-López, V.M.; Ragaert, P.; Debevere, J.; Devlieghere, F. Pulsed light for food decontamination: A review. Trends Food Sci. Technol. 2007, 18, 464–473. [Google Scholar] [CrossRef]
- Escott, C.; Vaquero, C.; del Fresno, J.M.; Bañuelos, M.A.; Loira, I.; Han, S.; Bi, Y.; Morata, A.; Suárez-Lepe, J.A. Pulsed light effect in red grape quality and fermentation. Food Bioprocess Technol. 2017, 10, 1540–1547. [Google Scholar] [CrossRef]
- Pérez-López, A.J.; Rodríguez-López, M.I.; Burló, F.; Carbonell-Barrachina, Á.A.; Gabaldón, J.A.; Gómez-López, V.M. Evaluation of Pulsed Light to Inactivate Brettanomyces bruxellensis in White Wine and Assessment of Its Effects on Color and Aromatic Profile. Foods 2020, 9, 1903. [Google Scholar] [CrossRef] [PubMed]
- Santamera, A.; Escott, C.; Loira, I.; Del Fresno, J.M.; González, C.; Morata, A. Pulsed light: Challenges of a non-thermal sanitation technology in the winemaking industry. Beverages 2020, 6, 45. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Guamis, B.; Raso, J.; del Fresno, J.M.; Escott, C.; Bañuelos, M.A.; Álvarez, I.; Tesfaye, W.; González, C.; et al. Emerging Technologies to Increase Extraction, Control Microorganisms, and Reduce SO2. In Winemaking—Stabilization, Aging Chemistry and Biochemistry; Cosme, F., Ed.; IntechOpen: London, UK, 2020; pp. 1–20. [Google Scholar]
- Liu, P.T.; Lu, L.; Duan, C.Q.; Yan, G.L. The contribution of indigenous non-Saccharomyces wine yeast to improved aromatic quality of Cabernet Sauvignon wines by spontaneous fermentation. LWT—Food Sci. Technol. 2016, 71, 356–363. [Google Scholar] [CrossRef]
- Pinto, L.; Baruzzi, F.; Cocolin, L.; Malfeito-Ferreira, M. Emerging technologies to control Brettanomyces spp. in wine: Recent advances and future trends. Trends Food Sci. Technol. 2020, 99, 88–100. [Google Scholar] [CrossRef]
- Avalos-Llano, K.R.; Martín-Belloso, O.; Soliva-Fortuny, R. Effect of pulsed light treatments on quality and antioxidant properties of fresh-cut strawberries. Food Chem. 2018, 264, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Tibiru Apaliya, M.; Sackle Sackey, A.; Wu, M.; Xiao, L. Effect of pulsed light treatment on the phytochemical, volatile, and sensorial attributes of lactic-acid-fermented mulberry juice. Int. J. Food Prop. 2018, 21, 213–228. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Sackey, A.S.; Wu, M.; Xiao, L. Impact of ultrasonication and pulsed light treatments on phenolics concentration and antioxidant activities of lactic-acid-fermented mulberry juice. LWT—Food Sci. Technol. 2018, 92, 61–66. [Google Scholar] [CrossRef]
- Rodov, V.; Vinokur, Y.; Horev, B. Brief postharvest exposure to pulsed light stimulates coloration and anthocyanin accumulation in fig fruit (Ficus carica L.). Postharvest Biol. Technol. 2012, 68, 43–46. [Google Scholar] [CrossRef]
- Morata, A.; González, C.; Tesfaye, W.; Loira, I.; Suárez-Lepe, J.A. Maceration and Fermentation: New Technologies to Increase Extraction. In Red Wine Technology; Morata, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 35–49. ISBN 9780128144008. [Google Scholar]
- Bintsis, T.; Litopoulou-Tzanetaki, E.; Robinson, R.K. Existing and potential applications of ultraviolet light in the food industry—A critical review. J. Sci. Food Agric. 2000, 80, 637–645. [Google Scholar] [CrossRef]
- Wright, J.R.; Sumner, S.S.; Hackney, C.R.; Pierson, M.D.; Zoecklein, B.W. Efficacy of Ultraviolet Light for Reducing Escherichia coli O157:H7 in Unpasteurized Apple Cider. J. Food Prot. 2000, 63, 563–567. [Google Scholar] [CrossRef]
- OIV. Analysis of Volatile Compounds in Wines by Gas Chromatography; OIV: Paris, France, 2016. [Google Scholar]
- Bañuelos, M.A.; Loira, I.; Buenaventura, G.; Escott, C.; del Fresno, J.M.; Codina-Torrella, I.; Quevedo, J.M.; Gervilla, R.; Rodríguez Chavarría, J.M.; de Lamo, S.; et al. White wine processing by UHPH without SO2. Elimination of microbial populations and effect in oxidative enzymes, colloidal stability and sensory quality. Food Chem. 2020, 332, 127417. [Google Scholar] [CrossRef] [PubMed]
- Wekhof, A. Disinfection with flash lamps. PDA J. Pharm. Sci. Technol. 2000, 54, 264–276. [Google Scholar]
- Fava, J.; Hodara, K.; Nieto, A.; Guerrero, S.; Alzamora, S.M.; Castro, M.A. Structure (micro, ultra, nano), color and mechanical properties of Vitis labrusca L. (grape berry) fruits treated by hydrogen peroxide, UV-C irradiation and ultrasound. Food Res. Int. 2011, 44, 2938–2948. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Vejarano, R.; González, C.; Callejo, M.J.; Suárez-Lepe, J.A. Emerging preservation technologies in grapes for winemaking. Trends Food Sci. Technol. 2017, 67, 36–43. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gómez, M.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef] [PubMed]
- Loira, I.; Morata, A.; Palomero, F.; González, C.; Suárez-Lepe, J.A. Schizosaccharomyces pombe: A promising biotechnology for modulating wine composition. Fermentation 2018, 4, 70. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.A.; Morata, A. New trends in yeast selection for winemaking. Trends Food Sci. Technol. 2012, 23, 39–50. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.A.; Palomero, F.; Benito, S.; Calderón, F.; Morata, A. Oenological versatility of Schizosaccharomyces spp. Eur. Food Res. Technol. 2012, 235, 375–383. [Google Scholar] [CrossRef]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez-Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Vilela, A. Lachancea thermotolerans, the Non-Saccharomyces Yeast that Reduces the Volatile Acidity of Wines. Fermentation 2018, 4, 56. [Google Scholar] [CrossRef]
- Clemente-Jimenez, J.M.; Mingorance-Cazorla, L.; Martínez-Rodríguez, S.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F. Molecular characterization and oenological properties of wine yeasts isolated during spontaneous fermentation of six varieties of grape must. Food Microbiol. 2004, 21, 149–155. [Google Scholar] [CrossRef]
- Mateo, J.J.; Jiménez, M.; Pastor, A.; Huerta, T. Yeast starter cultures affecting wine fermentation and volatiles. Food Res. Int. 2001, 34, 307–314. [Google Scholar] [CrossRef]
- Cappello, M.S.; Zapparoli, G.; Logrieco, A.; Bartowsky, E.J. Linking wine lactic acid bacteria diversity with wine aroma and flavour. Int. J. Food Microbiol. 2017, 243, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.; Velázquez, R. The Yeast Torulaspora delbrueckii: An Interesting But Difficult-To-Use Tool for Winemaking. Fermentation 2018, 4, 94. [Google Scholar] [CrossRef]
- Loira, I.; Vejarano, R.; Bañuelos, M.A.; Morata, A.; Tesfaye, W.; Uthurry, C.; Villa, A.; Cintora, I.; Suárez-Lepe, J.A. Influence of sequential fermentation with Torulaspora delbrueckii and Saccharomyces cerevisiae on wine quality. LWT—Food Sci. Technol. 2014, 59, 915–922. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Tesfaye, W.; Bañuelos, M.A.; González, C.; Suárez Lepe, J.A. Lachancea thermotolerans applications in wine technology. Fermentation 2018, 4, 53. [Google Scholar] [CrossRef]
- Martin, V.; Giorello, F.; Fariña, L.; Minteguiaga, M.; Salzman, V.; Boido, E.; Aguilar, P.S.; Gaggero, C.; Dellacassa, E.; Mas, A.; et al. De novo synthesis of benzenoid compounds by the yeast Hanseniaspora vineae increases the flavor diversity of wines. J. Agric. Food Chem. 2016, 64, 4574–4583. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Lu, Y.; Sun, F.; Wang, W.; Liu, Y.; Wang, J.; Sun, J.; Mu, J.; Gao, Z. Effects of spontaneous fermentation on the microorganisms diversity and volatile compounds during “Marselan” from grape to wine. LWT—Food Sci. Technol. 2020, 134, 110193. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]
- Xiaokang, W.; Brunton, N.P.; Lyng, J.G.; Harrison, S.M.; Carpes, S.T.; Papoutsis, K. Volatile and non-volatile compounds of shiitake mushrooms treated with pulsed light after twenty-four hour storage at different conditions. Food Biosci. 2020, 36, 100619. [Google Scholar] [CrossRef]
- Morata, A.; Gómez-Cordovés, M.C.; Suberviola, J.; Bartolomé, B.; Colomo, B.; Suárez, J.A. Adsorption of Anthocyanins by Yeast Cell Walls during the Fermentation of Red Wines. J. Agric. Food Chem. 2003, 51, 4084–4088. [Google Scholar] [CrossRef]
- Echeverrigaray, S.; Scariot, F.J.; Menegotto, M.; Delamare, A.P.L. Anthocyanin adsorption by Saccharomyces cerevisiae during wine fermentation is associated to the loss of yeast cell wall/membrane integrity. Int. J. Food Microbiol. 2020, 314, 108383. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Escott, C.; Loira, I.; Del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Influence of Saccharomyces and non-Saccharomyces Yeasts in the Formation of Pyranoanthocyanins and Polymeric Pigments during Red Wine Making. Molecules 2019, 24, 4490. [Google Scholar] [CrossRef]
- Vasserot, Y.; Caillet, S.; Maujean, A. Study of Anthocyanin Adsorption by Yeast Lees. Effect of Some Physicochemical Parameters. Am. J. Enol. Vitic. 1997, 48, 433–437. [Google Scholar]
- Morata, A.; Gómez-Cordovés, M.C.; Colomo, B.; Suárez, J.A. Cell wall anthocyanin adsorption by different Saccharomyces strains during the fermentation of Vitis vinifera L. cv Graciano grapes. Eur. Food Res. Technol. 2005, 220, 341–346. [Google Scholar] [CrossRef]
- Gómez-Míguez, M.; González-Miret, M.L.; Heredia, F.J. Evolution of colour and anthocyanin composition of Syrah wines elaborated with pre-fermentative cold maceration. J. Food Eng. 2007, 79, 271–278. [Google Scholar] [CrossRef]
- Loira, I.; Morata, A.; Bañuelos, M.A.; Suárez-Lepe, J.A. Isolation, Selection, and Identification Techniques for Non-Saccharomyces Yeasts of Oenological Interest. In Biotechnological Progress and Beverage Consumption; Grumezescu, A.M., Holban, A.M., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 467–578. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Starter | Inocula d0 CFU/mL | Sequential Inocula d8 CFU/mL |
|---|---|---|
| T. delbrueckii | 1.1 × 108 | Sc 6.0 × 107 |
| S. pombe | 3.2 × 107 | Sc 6.0 × 107 |
| M. pulcherrima | 6.9 × 107 | Sc 6.0 × 107 |
| L. thermotolerans | 6.0 × 107 | Sc 6.0 × 107 |
| H. vineae | 1.3 × 108 | Sc 6.0 × 107 |
| S. cerevisiae | 1.0 × 108 | Sc 6.0 × 107 |
| Ethanol | pH | Total Acidity 1 | Volatile Acidity 2 | Malic Acid | Lactic Acid | Fructose | Glucose | |
|---|---|---|---|---|---|---|---|---|
| % v/v | g/L | g/L | g/L | g/L | g/L | g/L | ||
| A | ||||||||
| Spontaneous | 14.1 ± 0.1 a | 4.0 ± 0.0 a | 4.4 ± 0.1 b | 0.5 ± 0.0 bc | 0.7 ± 0.1 c | 1.3 ± 0.1 bc | 2.7 ± 0.1 ab | 0.0 ± 0.0 b |
| T. delbrueckii | 14.1 ± 0.1 a | 3.9 ± 0.0 a | 4.3 ± 0.2 b | 0.1 ± 0.0 d | 1.3 ± 0.0 b | 0.1 ± 0.0 c | 2.6 ± 1.0 ab | 0.4 ± 0.2 b |
| S. pombe | 13.7 ± 0.3 b | 4.0 ± 0.1 a | 5.5 ± 1.6 ab | 0.6 ± 0.1 b | 0.5 ± 0.2 c | 1.9 ± 0.9 bc | 1.3 ± 1.5 bc | 1.0 ± 0.8 b |
| M. pulcherrima | 13.7 ± 0.3 b | 3.9 ± 0.0 a | 6.8 ± 1.3 a | 0.9 ± 0.3 a | 0.1 ± 0.1 d | 3.1 ± 0.8 ab | 0.3 ± 0.3 c | 2.2 ± 1.3 a |
| L. thermotolerans | 13.8 ± 0.2 ab | 3.6 ± 0.2 b | 6.9 ± 1.1 a | 0.4 ± 0.0 bc | 0.2 ± 0.3 d | 4.6 ± 1.7 a | 0.0 ± 0.1 c | 0.1 ± 0.2 b |
| H. vineae | 14.2 ± 0.1 a | 3.9 ± 0.0 a | 4.1 ± 0.1 b | 0.3 ± 0.0 cd | 1.4 ± 0.1 b | 0.0 ± 0.0 c | 3.0 ± 0.6 a | 0.9 ± 0.3 b |
| S. cerevisiae | 13.8 ± 0.2 ab | 3.9 ± 0.0 a | 5.6 ± 0.1 ab | 0.2 ± 0.0 cd | 2.2 ± 0.1 a | 0.7 ± 0.2 c | 0.5 ± 0.1 c | 0.0 ± 0.0 b |
| B | ||||||||
| Spontaneous | 14.1 ± 0.1 a | 3.9 ± 0.0 c | 4.8 ± 0.0 c | 0.4 ± 0.1 ab | 1.6 ± 0.0 b | 0.0 ± 0.0 b | 0.9 ± 0.1 b | 0.1 ± 0.1 b |
| T. delbrueckii | 13.6 ± 0.2 cd | 3.9 ± 0.0 bc | 4.6 ± 0.1 c | 0.2 ± 0.1 d | 1.3 ± 0.1 bc | 0.2 ± 0.1 b | 1.7 ± 0.6 a | 0.3 ± 0.3 a |
| S. pombe | 13.8 ± 0.2 bc | 4.0 ± 0.0 a | 3.6 ± 0.2 d | 0.3 ± 0.0 c | 0.5 ± 0.1 d | 0.5 ± 0.1 b | 0.9 ± 0.2 b | 0.2 ± 0.1 ab |
| M. pulcherrima | 13.4 ± 0.2 d | 3.9 ± 0.0 b | 4.7 ± 0.3 c | 0.4 ± 0.0 a | 0.0 ± 0.1 e | 0.8 ± 0.1 b | 0.0 ± 0.0 d | 0.3 ± 0.1 a |
| L. thermotolerans | 13.7 ± 0.1 bc | 3.5 ± 0.0 d | 7.2 ± 0.5 a | 0.3 ± 0.0 bc | 1.1 ± 0.6 c | 4.9 ± 0.7 a | 0.7 ± 0.2 bc | 0.0 ± 0.0 b |
| H. vineae | 13.8 ± 0.1 ab | 3.9 ± 0.0 c | 4.4 ± 0.1 c | 0.3 ± 0.0 bc | 1.3 ± 0.1 bc | 0.2 ± 0.1 b | 1.0 ± 0.1 b | 0.1 ± 0.1 b |
| S. cerevisiae | 13.6 ± 0.1 cd | 3.9 ± 0.0 c | 5.8 ± 0.1 b | 0.2 ± 0.0 d | 2.1 ± 0.1 a | 0.7 ± 0.0 b | 0.3 ± 0.1 cd | 0.0 ± 0.0 b |
| A | Spontaneous | T. delbrueckii | S. pombe | L. thermotolerans | H. vineae | M. pulcherrima | S. cerevisiae |
|---|---|---|---|---|---|---|---|
| Acetaldehyde | 7.8 ± 0.9 a | 12.6 ± 8.4 a | 13.1 ± 3.3 a | 9.4 ± 1.0 a | 8.4 ± 0.5 a | 7.9 ± 0.3 a | 7.1 ± 1.2 a |
| Methanol | 26.9 ± 1.0 ab | 26.6 ± 4.2 b | 23.0 ± 0.3 ab | 23.1 ± 1.2 b | 24.9 ± 3.7 b | 23.7 ± 1.7 b | 29.7 ± 2.0 a |
| 1-propanol | 43.2 ± 4.0 b | 52.2 ± 1.3 a | 42.6 ± 1.2 b | 44.6 ± 1.6 b | 40.3 ± 4.6 b | 41.2 ± 1.6 b | 20.1 ± 1.0 c |
| Diacetyl | 1.5 ± 0.1 ab | 1.5 ± 0.1 b | 1.7 ± 0.1 ab | 1.6 ± 0.1 ab | 1.9 ± 0.2 a | 1.6 ± 0.1 b | 1.7 ± 0.4 ab |
| Ethyl acetate | 159.8 ± 17.9 a | 82.2 ± 16.2 c | 90.9 ± 10.3 c | 62.9 ± 7.6 d | 80.8 ± 1.4 c | 113.1 ± 7.4 b | 19.8 ± 1.5 e |
| 2-butanol | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Isobutanol | 45.0 ± 7.6 ab | 56.9 ± 12.9 a | 39.8 ± 3.2 b | 38.9 ± 5.1 b | 23.4 ± 1.7 c | 42.6 ± 5.7 b | 27.0 ± 0.5 c |
| 1-butanol | 4.3 ± 0.2 a | 4.8 ± 0.5 a | 4.4 ± 0.3 a | 4.6 ± 0.3 a | 4.9 ± 5.1 a | 4.8 ± 0.1 a | 4.8 ± 0.1 a |
| Acetoin | 6.0 ± 0.5 b | 5.4 ± 0.4 b | 19.2 ± 10.1 a | 6.2 ± 0.7 b | 7.3 ± 1.4 b | 7.1 ± 1.0 b | 5.8 ± 0.3 b |
| 3-methyl-1-butanol | 102.5 ± 4.7 c | 139.4 ± 18.1 b | 95.2 ± 6.3 c | 113.0 ± 7.0 bc | 106.4 ± 13.1 c | 93.6 ± 5.0 c | 192.4 ± 37.5 a |
| 2-methyl-1-butanol | 47.4 ± 2.6 a | 45.2 ± 2.7 a | 45.2 ± 1.2 a | 38.7 ± 3.2 a | 35.2 ± 1.9 a | 47.0 ± 2.8 a | 64.8 ± 56.1 a |
| Isobutyl acetate | 1.1 ± 1.5 a | 1.1 ± 1.9 a | 1.0 ± 1.7 a | 2.3 ± 0.9 a | 1.6 ± 2.8 a | 3.3 ± 0.8 a | 3.0 ± 0.6 a |
| Ethyl butyrate | 0.8 ± 1.1 ab | 3.0 ± 2.4 a | 0.9 ± 0.7 b | 1.8 ± 0.9 ab | 0.9 ± 0.8 ab | 0.9 ± 0.8 ab | 1.4 ± 0.4 ab |
| Ethyl lactate | 46.5 ± 8.8 ab | 31.9 ± 17.0 bc | 13.7 ± 4.5 d | 21.8 ± 8.3 cd | 51.2 ± 6.6 a | 15.6 ± 1.5 d | 11.6 ± 1.2 d |
| 2,3-butanediol | 529.2 ± 17.4 bcd | 547.4 ± 67.8 bc | 672.6 ± 35.7 a | 586.7 ± 63.5 b | 470.2 ± 53.7 cd | 614.5 ± 33.4 ab | 445.9 ± 24.4 d |
| Isoamyl acetate | 1.9 ± 0.1 abc | 2.7 ± 0.3 a | 1.8 ± 0.1 bc | 2.5 ± 0.1 ab | 1.9 ± 0.1 ab | 1.9 ± 0.0 b | 1.2 ± 1.0 c |
| Hexanol | 4.1 ± 0.8 b | 4.4 ± 0.1 b | 4.6 ± 0.1 ab | 4.4 ± 0.1 b | 4.3 ± 0.1 b | 4.6 ± 0.1 ab | 5.0 ± 0.6 a |
| 2-phenyl ethanol | 29.2 ± 0.8 cd | 35.7 ± 2.8 bc | 29.8 ± 1.2 cd | 41.5 ± 7.3 b | 25.6 ± 1.5 d | 34.5 ± 2.3 bc | 76.6 ± 6.5 a |
| 2-phenyl ethyl acetate | 6.5 ± 1.5 a | 5.3 ± 0.2 a | 5.1 ± 0.1 a | 5.0 ± 0.3 a | 6.2 ± 1.9 a | 5.2 ± 0.3 a | 5.2 ± 0.4 a |
| B | Spontaneous | T. delbrueckii | S. pombe | L. thermotolerans | H. vineae | M. pulcherrima | S. cerevisiae |
| Acetaldehyde | 14.1 ± 0.9 a | 10.0 ± 1.2 b | 10.0 ± 3.0 b | 8.2 ± 1.8 b | 13.5 ± 0.7 a | 7.8 ± 1.2 b | 8.4 ± 0.1 b |
| Methanol | 21.7 ± 1.2 ab | 24.0 ± 2.0 a | 24.0 ± 3.5 a | 22.4 ± 3.3 ab | 17.9 ± 1.3 b | 26.0 ± 0.2 a | 22.5 ± 3.6 a |
| 1-propanol | 54.0 ± 1.2 bc | 71.5 ± 5.0 a | 40.6 ± 15.1 c | 52.8 ± 1.3 b | 48.7 ± 2.2 bc | 40.5 ± 7.1 c | 26.6 ± 0.6 d |
| Diacetyl | 1.5 ± 0.1 b | 1.7 ± 0.3 ab | 1.6 ± 0.1 ab | 1.5 ± 0.1 b | 1.8 ± 0.1 a | 1.5 ± 0.0 b | 1.5 ± 0.1 b |
| Ethyl acetate | 147.3 ± 3.2 a | 118.5 ± 17.8 b | 120.3 ± 3.2 b | 96.5 ± 4.6 c | 81.2 ± 3.2 c | 134.1 ± 16.3 ab | 24.6 ± 2.8 d |
| 2-butanol | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Isobutanol | 64.4 ± 2.7 a | 53.5 ± 6.1 ab | 39.2 ± 6.5 c | 42.2 ± 11.0 c | 19.5 ± 0.4 d | 45.3 ± 2.6 bc | 25.5 ± 0.8 d |
| 1-butanol | 4.4 ± 0.1 bc | 4.5 ± 0.2 abc | 4.4 ± 0.5 bc | 4.9 ± 0.4 a | 4.5 ± 0.1 abc | 4.1 ± 0.1 c | 4.8 ± 0.1 ab |
| Acetoin | 4.8 ± 0.2 a | 7.2 ± 1.4 a | 8.0 ± 3.9 a | 7.1 ± 1.5 a | 8.4 ± 2.5 a | 6.3 ± 0.7 a | 5.8 ± 1.0 a |
| 3-methyl-1-butanol | 89.5 ± 3.2 cd | 132.9 ± 16.4 b | 64.2 ± 14.8 e | 113.0 ± 16.5 bc | 100.0 ± 4.6 cd | 80.6 ± 14.2 de | 167.3 ± 3.0 a |
| 2-methyl-1-butanol | 51.2 ± 0.3 b | 44.1 ± 4.5 bc | 33.3 ± 7.9 de | 36.4 ± 8.7 cd | 25.4 ± 1.0 e | 31.4 ± 2.7 de | 81.5 ± 1.9 a |
| Isobutyl acetate | 2.6 ± 0.1 ab | 3.1 ± 0.9 a | 2.3 ± 1.0 ab | 1.0 ± 0.9 b | 1.8 ± 1.8 ab | 3.2 ± 0.9 a | 0.8 ± 1.5 b |
| Ethyl butyrate | 2.7 ± 0.1 a | 1.3 ± 0.0 ab | 1.1 ± 1.8 ab | 1.4 ± 0.2 ab | 0.5 ± 0.8 b | 1.1 ± 1.1 ab | 1.6 ± 0.5 ab |
| Ethyl lactate | 36.1 ± 2.2 a | 25.6 ± 16.2 a | 31.2 ± 3.2 a | 42.2 ± 3.9 a | 22.5 ± 8.1 a | 12.2 ± 0.7 a | 28.7 ± 13.3 a |
| 2,3-butanediol | 766.5 ± 8.5 a | 588.3 ± 22.0 bc | 775.9 ± 182.3 a | 687.5 ± 73.7 ab | 459.7 ± 38.1 c | 638.7 ± 54.0 ab | 596.7 ± 41.4 bc |
| Isoamyl acetate | 1.7 ± 0.0 cd | 8.2 ± 0.4 a | 1.7 ± 0.0 d | 2.3 ± 0.6 bc | 2.3 ± 0.3 b | 1.8 ± 0.1 bcd | 2.0 ± 0.1 bcd |
| Hexanol | 4.2 ± 0.0 a | 3.8 ± 0.0 b | 4.1 ± 0.2 ab | 3.9 ± 0.2 ab | 4.1 ± 0.2 ab | 4.1 ± 0.1 a | 4.1 ± 0.2 ab |
| 2-phenyl ethanol | 24.6 ± 1.6 cd | 35.2 ± 8.1 b | 20.5 ± 4.1 d | 29.6 ± 0.9 bc | 25.5 ± 2.0 cd | 24.1 ± 2.6 cd | 62.9 ± 2.1 a |
| 2-phenyl ethyl acetate | 5.5 ± 0.1 a | 5.5 ± 0.4 a | 6.3 ± 2.1 a | 4.2 ± 3.8 a | 6.6 ± 0.8 a | 6.0 ± 1.5 a | 5.1 ± 0.1 a |
| TPI | CI | Hue | |
|---|---|---|---|
| A | |||
| Spontaneous | 11.1 ± 0.1 bc | 0.8 ± 0.0 b | 1.2 ± 0.0 bc |
| T. delbrueckii | 10.4 ± 0.6 c | 0.6 ± 0.1 c | 1.3 ± 0.1 b |
| S. pombe | 11.2 ± 0.2 bc | 0.7 ± 0.0 bc | 1.4 ± 0.0 a |
| M. pulcherrima | 10.7 ± 0.1 bc | 0.8 ± 0.0 b | 0.9 ± 0.0 d |
| L. thermotolerans | 11.3 ± 0.3 b | 0.8 ± 0.0 b | 1.3 ± 0.0 b |
| H. vineae | 10.9 ± 0.1 bc | 0.6 ± 0.0 c | 1.3 ± 0.0 ab |
| S. cerevisiae | 13.0 ± 0.1 a | 1.0 ± 0.0 a | 1.1 ± 0.0 cd |
| B | |||
| Spontaneous | 8.7 ± 0.0 b | 0.6 ± 0.0 a | 2.0 ± 0.0 ab |
| T. delbrueckii | 8.2 ± 0.1 b | 0.3 ± 0.0 c | 2.6 ± 0.1 a |
| S. pombe | 8.9 ± 0.1 b | 0.5 ± 0.1 ab | 2.0 ± 0.3 ab |
| M. pulcherrima | 8.4 ± 0.3 b | 0.4 ± 0.1 abc | 2.1 ± 0.1 ab |
| L. thermotolerans | 8.5 ± 0.4 b | 0.3 ± 0.1 bc | 1.5 ± 0.1 b |
| H. vineae | 8.7 ± 0.2 b | 0.3 ± 0.1 c | 2.4 ± 0.5 a |
| S. cerevisiae | 10.7 ± 0.9 a | 0.4 ± 0.0 abc | 1.5 ± 0.1 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escott, C.; López, C.; Loira, I.; González, C.; Bañuelos, M.A.; Tesfaye, W.; Suárez-Lepe, J.A.; Morata, A. Improvement of Must Fermentation from Late Harvest cv. Tempranillo Grapes Treated with Pulsed Light. Foods 2021, 10, 1416. https://doi.org/10.3390/foods10061416
Escott C, López C, Loira I, González C, Bañuelos MA, Tesfaye W, Suárez-Lepe JA, Morata A. Improvement of Must Fermentation from Late Harvest cv. Tempranillo Grapes Treated with Pulsed Light. Foods. 2021; 10(6):1416. https://doi.org/10.3390/foods10061416
Chicago/Turabian StyleEscott, Carlos, Carmen López, Iris Loira, Carmen González, María Antonia Bañuelos, Wendu Tesfaye, José Antonio Suárez-Lepe, and Antonio Morata. 2021. "Improvement of Must Fermentation from Late Harvest cv. Tempranillo Grapes Treated with Pulsed Light" Foods 10, no. 6: 1416. https://doi.org/10.3390/foods10061416
APA StyleEscott, C., López, C., Loira, I., González, C., Bañuelos, M. A., Tesfaye, W., Suárez-Lepe, J. A., & Morata, A. (2021). Improvement of Must Fermentation from Late Harvest cv. Tempranillo Grapes Treated with Pulsed Light. Foods, 10(6), 1416. https://doi.org/10.3390/foods10061416