
Sorghum Phenolic Compounds Are Associated with Cell Growth Inhibition through Cell Cycle Arrest and Apoptosis in Human Hepatocarcinoma and Colorectal Adenocarcinoma Cells

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sorghum Accessions
2.2. Reagents
2.3. Phenolic Extraction
2.4. Total Phenolic Content
2.5. Cell Culture
2.6. Cell Growth Inhibition Assay
2.7. Cytotoxicity Assay
2.8. Cell Cycle Analysis
2.9. Apoptosis Analysis
2.10. Statistics
3. Results
3.1. Total Phenolic Content
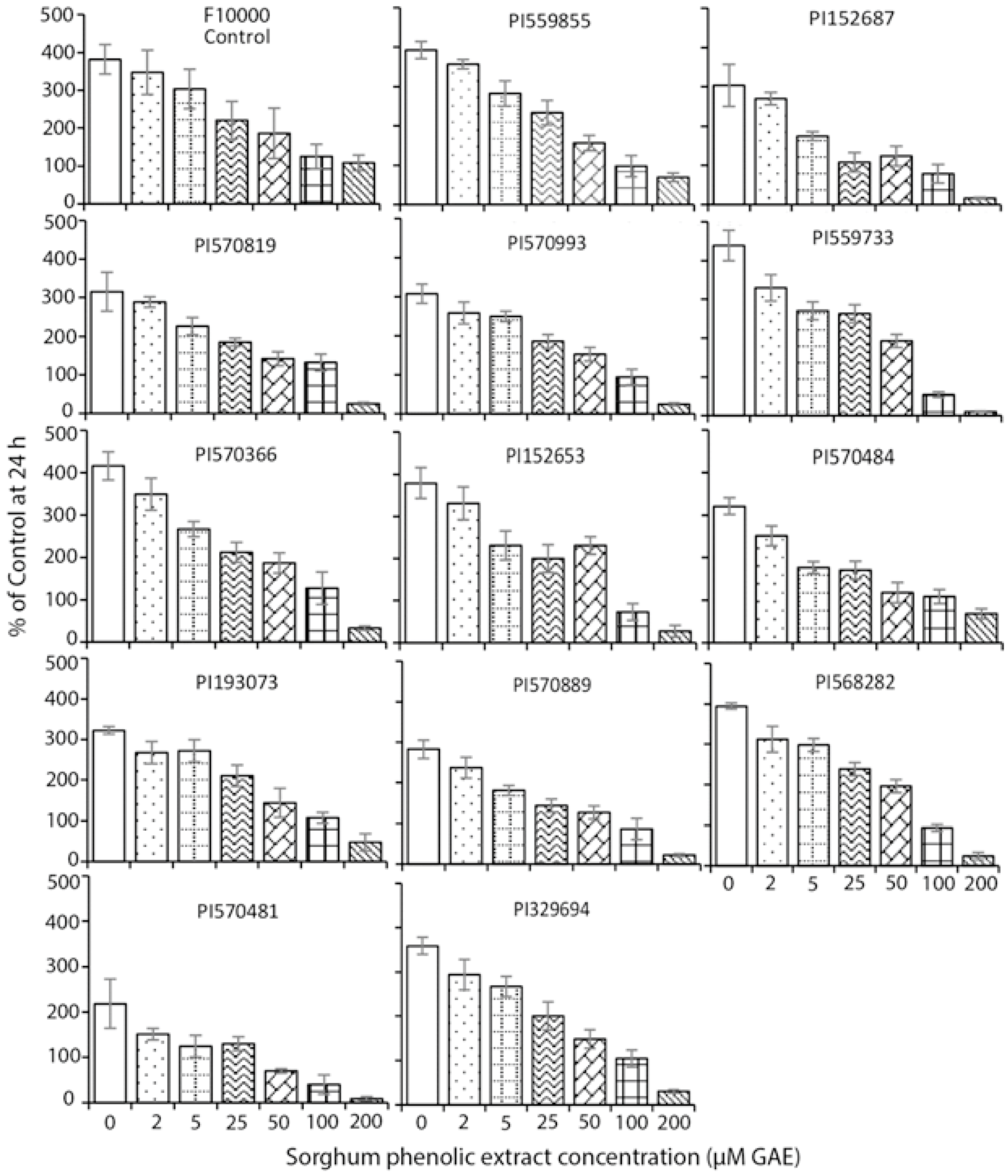
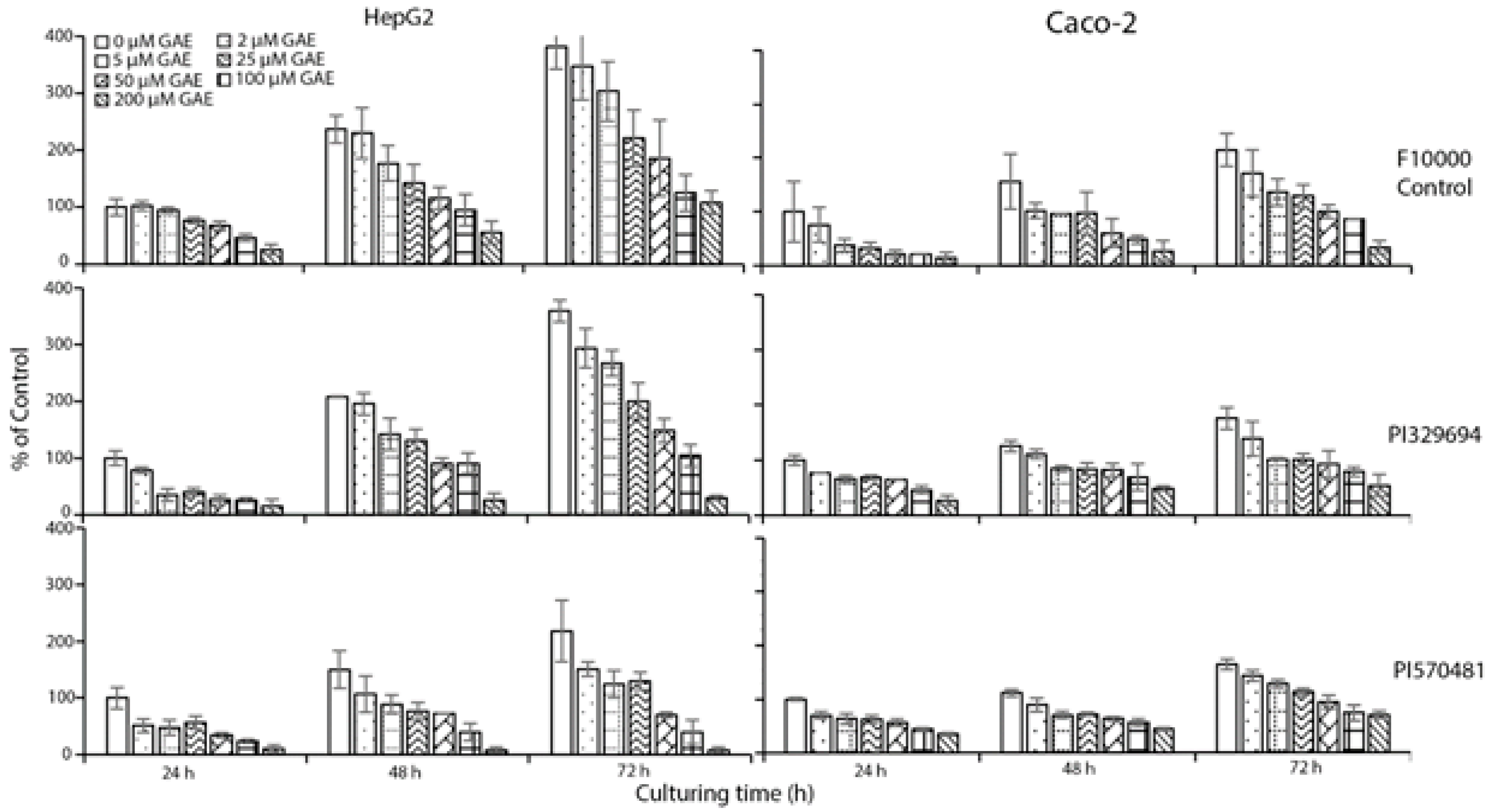
3.2. Cell Growth Inhibition
3.3. Cytotoxicity Assay
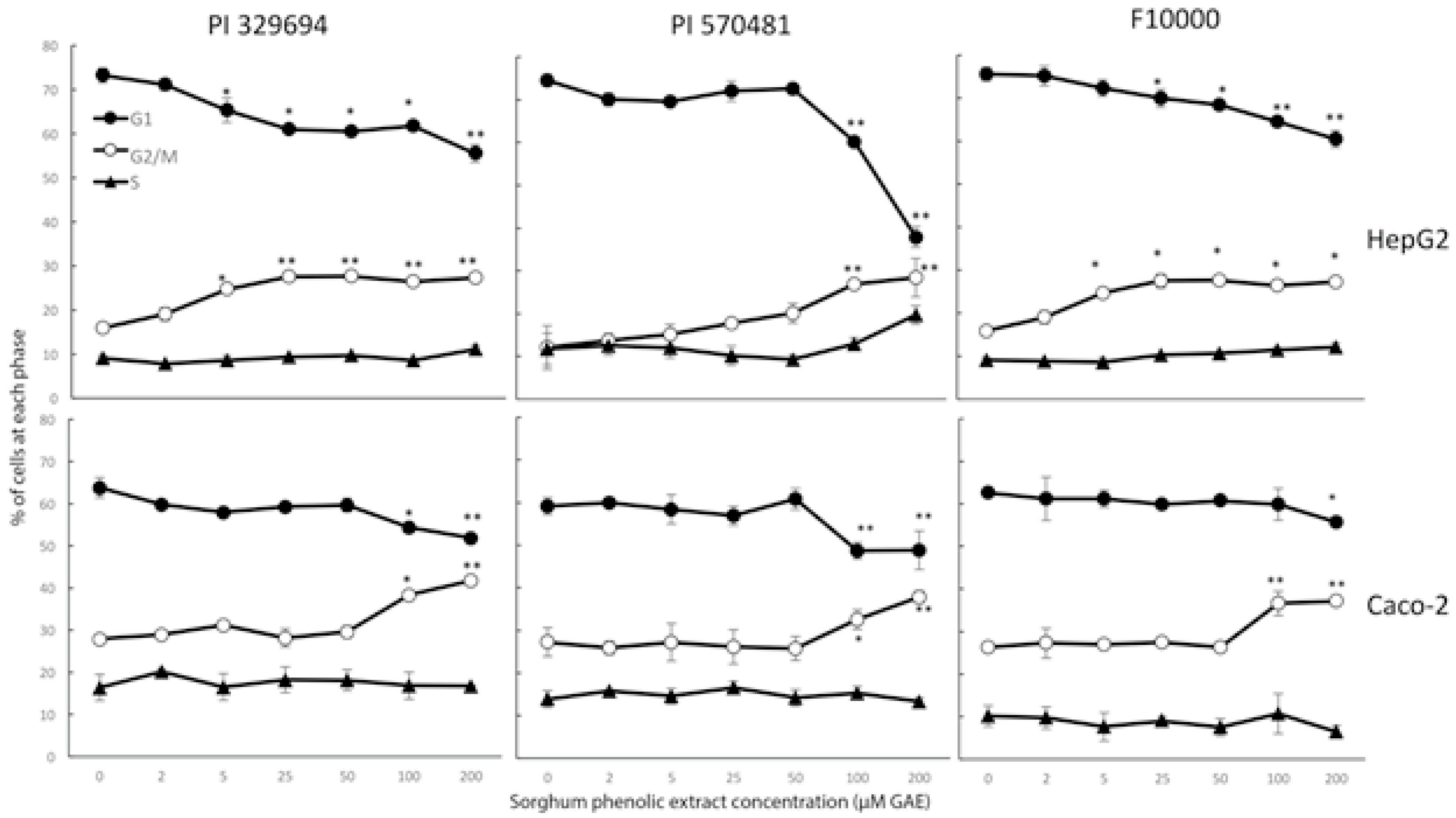
3.4. Cell Cycle Arrest
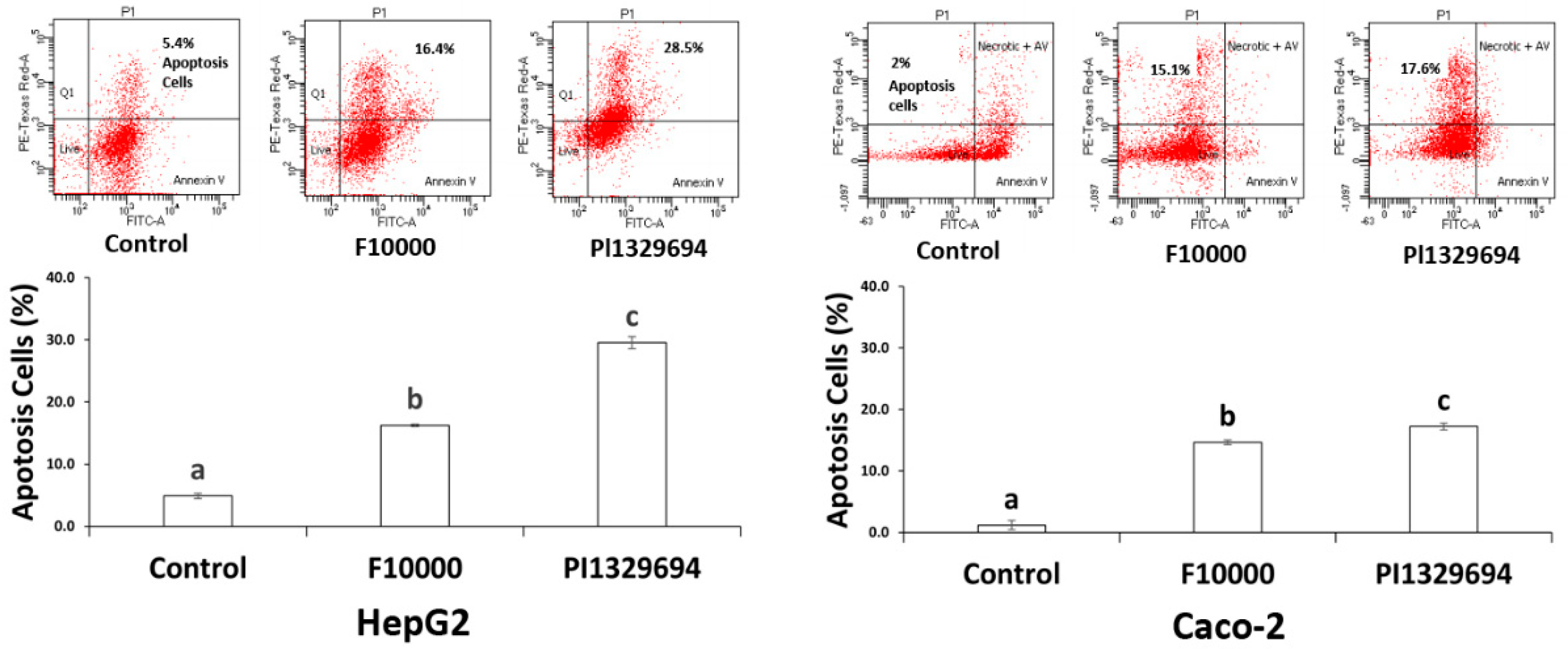
3.5. Apoptosis
3.6. Correlation Coefficient
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Disclaimer
References
- Stefoska-Needham, A.; Beck, E.J.; Johnson, S.K.; Tapsell, L.C. Sorghum: An Underutilized Cereal Whole Grain with the Potential to Assist in the Prevention of Chronic Disease. Food Rev. Int. 2015, 31, 401–437. [Google Scholar] [CrossRef]
- Hwang, K.; Weller, C.; Cuppett, S.L.; Hanna, M. Policosanol Contents and Composition of Grain Sorghum Kernels and Dried Distillers Grains. Cereal Chem. 2004, 81, 345–349. [Google Scholar] [CrossRef]
- Su, X.; Rhodes, D.; Xu, J.; Chen, X.; Davis, H.; Wang, D.; Herald, T.J.; Wang, W. Phenotypic Diversity of Anthocyanins in Sorghum Accessions with Various Pericarp Pigments. J. Nutr. Food Sci. 2017, 7, 1000610. [Google Scholar] [CrossRef]
- Shen, Y.; Su, X.; Rhodes, D.; Herald, T.; Xu, J.; Chen, X.; Smith, J.S.; Wang, W. The pigments of sorghum pericarp are associated with the contents of carotenoids and pro-vitamin A. Int. J. Food Nutr. Sci. 2017, 6, 48–56. [Google Scholar]
- Davis, H.; Su, X.; Shen, Y.; Xu, J.; Wang, D.; Smith, J.S.; Aramouni, F.; Wang, W. Phenotypic diversity of colored phytochemicals in sorghum accessions with various pericarp pigments. In Polyphenols in Plants, 2nd ed.; Watson, R.R., Ed.; Academic Press: New York, NY, USA, 2019; pp. 123–131. [Google Scholar]
- Doll, R.; Peto, R. The causes of cancer: Quantitative estimates of avoidable risks of cancer in the United States today. J. Natl. Cancer Inst. 1981, 66, 1191–1308. [Google Scholar] [CrossRef]
- Ragaee, S.; Abdel-Aal, E.-S.M.; Noaman, M. Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 2006, 98, 32–38. [Google Scholar] [CrossRef]
- Awika, J.M.; Rooney, L.W. Sorghum phytochemicals and their potential impact on human health. Phytochemistry 2004, 65, 1199–1221. [Google Scholar] [CrossRef]
- Wang, W.; Goodman, M.T. Antioxidant properties of dietary phenolic agents in a human LDL-oxidation ex vivo model: Interaction of protein binding activity. Nutr. Res. 1999, 19, 191–202. [Google Scholar] [CrossRef]
- Van Rensburg, S.J. Epidemiologic and dietary evidence for a specific nutritional predisposition to esophageal cancer. J. Natl. Cancer Inst. 1981, 67, 243–251. [Google Scholar] [PubMed]
- Chen, F.; Cole, P.; Mi, Z.; Xing, L.Y. Corn and wheat-flour consumption and mortality from esophageal cancer in shanxi, China. Int. J. Cancer 1993, 53, 902–906. [Google Scholar] [CrossRef]
- Xu, S.; Shen, Y.; Xu, J.; Qi, G.; Chen, G.; Wang, W.; Sun, X.; Li, Y. Antioxidant and anticancer effects in human hepatocarcinoma (HepG2) cells of papain-hydrolyzed sorghum kafirin hydrolysates. J. Funct. Foods 2019, 58, 374–382. [Google Scholar] [CrossRef]
- Cox, S.; Noronha, L.; Herald, T.; Bean, S.; Lee, S.H.; Perumal, R.; Wang, W.; Smolensky, D. Evaluation of ethanol-based extraction conditions of sorghum bran bioactive compounds with downstream anti-proliferative properties in human cancer cells. Heliyon 2019, 5, e01589. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, C. The change of the staple diet of black South Africans from sorghum to maize (corn) is the cause of the epidemic of squamous carcinoma of the oesophagus. Med. Hypotheses 2005, 64, 658–660. [Google Scholar] [CrossRef]
- Birt, D.F.; Hendrich, S.; Wang, W. Dietary agents in cancer prevention: Flavonoids and isoflavonoids. Pharmacol. Ther. 2001, 90, 157–177. [Google Scholar] [CrossRef]
- Park, J.H.; Darvin, P.; Lim, E.J.; Joung, Y.H.; Hong, D.Y.; Park, E.U.; Park, S.H.; Choi, S.K.; Moon, E.S.; Cho, B.W.; et al. Hwanggeumchal sorghum induces cell cycle arrest, and suppresses tumor growth and metastasis through Jak2/STAT pathways in breast cancer xenografts. PLoS ONE 2012, 7, e40531. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.J.; Oh, I.T.; Lee, J.Y.; Jun, D.Y.; Seu, M.C.; Woo, K.S.; Nam, M.H.; Kim, Y.H. Apigeninidin induces apoptosis through activation of Bak and Bax and subsequent mediation of mitochondrial damage in human promyelocytic leukemia HL-60 cells. Process Biochem. 2012, 47, 1861–1871. [Google Scholar] [CrossRef]
- Devi, P.S.; Kumar, M.S.; Das, S.M. Evaluation of antiproliferative activity of red sorghum bran anthocyanin on a human breast cancer cell line (mcf-7). Int. J. Breast Cancer 2011, 2011, 891481. [Google Scholar] [CrossRef]
- Devi, P.S.; Saravanakumar, M.; Mohandas, S. Identification of 3- deoxyanthocyanins from red sorghum (Sorghum bicolor) bran and its biological properties. Afr. J. Pure Appl. Chem. 2011, 5, 181–193. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Ayella, A.; Lim, S.; Jiang, Y.; Iwamoto, T.; Lin, D.; Tomich, J.; Wang, W. Cytostatic inhibition of cancer cell growth by lignan secoisolariciresinol diglucoside. Nutr. Res. 2010, 30, 762–769. [Google Scholar] [CrossRef]
- Qu, H.; Madl, R.L.; Takemoto, D.J.; Baybutt, R.C. Lignans Are Involved in the Antitumor Activity of Wheat Bran in Colon Cancer SW480 Cells. J. Nutr. 2005, 135, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Smolensky, D.; Rhodes, D.; McVey, D.S.; Fawver, Z.; Perumal, R.; Herald, T.; Noronha, L. High-Polyphenol Sorghum Bran Extract Inhibits Cancer Cell Growth Through ROS Induction, Cell Cycle Arrest, and Apoptosis. J. Med. Food 2018, 21, 990–998. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sorghum Accession No. | Phenolic Contents (mg GAE/g DW) |
|---|---|
| F10000 (control) | 2.3 ± 0.2 |
| PI 559855 | 31.0 ± 0.2 |
| PI 152687 | 44.6 ± 2.9 |
| PI 570819 | 48.2 ± 4.4 |
| PI 570993 | 51.0 ± 1.9 |
| PI 559733 | 51.5 ± 1.4 |
| PI 570366 | 54.3 ± 1.7 |
| PI 152653 | 54.6 ± 1.2 |
| PI 570484 | 54.8 ± 0.8 |
| PI 193073 | 55.1 ± 4.5 |
| PI 570889 | 58.0 ± 2.0 |
| PI 568282 | 58.3 ± 2.5 |
| PI 570481 | 58.5 ± 2.5 |
| PI 329694 | 63.7 ± 2.5 |
| Sorghum Accession No. | IC50 a (mg DW/mL) | |
|---|---|---|
| HepG2 | Caco-2 | |
| F10000 (control) | 1275.6 | 1131.3 |
| PI 559855 | 221.8 | |
| PI 152687 | 138.9 | |
| PI 570819 | 192.1 | |
| PI 570993 | 146.2 | |
| PI 559733 | 90.8 | |
| PI 570366 | 120.9 | |
| PI 152653 | 127.9 | |
| PI 570484 | 158.0 | |
| PI 193073 | 177.3 | |
| PI 570889 | 120.9 | |
| PI 568282 | 113.3 | |
| PI 570481 | 85.8 | 115.6 |
| PI 329694 | 126.8 | 102.4 |
| IC50 | |||
|---|---|---|---|
| Total phenolics | −0.6806 * | ||
| Cell Arrest at G2/M | Apoptosis Cells | ||
| HepG2 | |||
| F10000 control | −0.8608 * | −0.9377 ** | |
| PI329694 | Cell number | −0.8281 * | −0.9764 ** |
| PI570481 | −0.9469 ** | ||
| Caco-2 | |||
| F10000 control | −0.7599 * | −0.9719 ** | |
| PI329694 | Cell number | −0.7655 * | −0.8199 * |
| PI570481 | −0.6840 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Shen, J.; Xu, J.; Herald, T.; Smolensky, D.; Perumal, R.; Wang, W. Sorghum Phenolic Compounds Are Associated with Cell Growth Inhibition through Cell Cycle Arrest and Apoptosis in Human Hepatocarcinoma and Colorectal Adenocarcinoma Cells. Foods 2021, 10, 993. https://doi.org/10.3390/foods10050993
Chen X, Shen J, Xu J, Herald T, Smolensky D, Perumal R, Wang W. Sorghum Phenolic Compounds Are Associated with Cell Growth Inhibition through Cell Cycle Arrest and Apoptosis in Human Hepatocarcinoma and Colorectal Adenocarcinoma Cells. Foods. 2021; 10(5):993. https://doi.org/10.3390/foods10050993
Chicago/Turabian StyleChen, Xi, Jiamin Shen, Jingwen Xu, Thomas Herald, Dmitriy Smolensky, Ramasamy Perumal, and Weiqun Wang. 2021. "Sorghum Phenolic Compounds Are Associated with Cell Growth Inhibition through Cell Cycle Arrest and Apoptosis in Human Hepatocarcinoma and Colorectal Adenocarcinoma Cells" Foods 10, no. 5: 993. https://doi.org/10.3390/foods10050993
APA StyleChen, X., Shen, J., Xu, J., Herald, T., Smolensky, D., Perumal, R., & Wang, W. (2021). Sorghum Phenolic Compounds Are Associated with Cell Growth Inhibition through Cell Cycle Arrest and Apoptosis in Human Hepatocarcinoma and Colorectal Adenocarcinoma Cells. Foods, 10(5), 993. https://doi.org/10.3390/foods10050993