
Antioxidant, Antidiabetic, and Antiobesity Properties, TC7-Cell Cytotoxicity and Uptake of Achyrocline satureioides (Marcela) Conventional and High Pressure-Assisted Extracts

 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Raw Material and Chemicals
2.2. TC7-Cells and Reagents for Cell Culture
2.3. Methods
2.3.1. Preparation of Marcela Extracts
2.3.2. Proximate Analysis
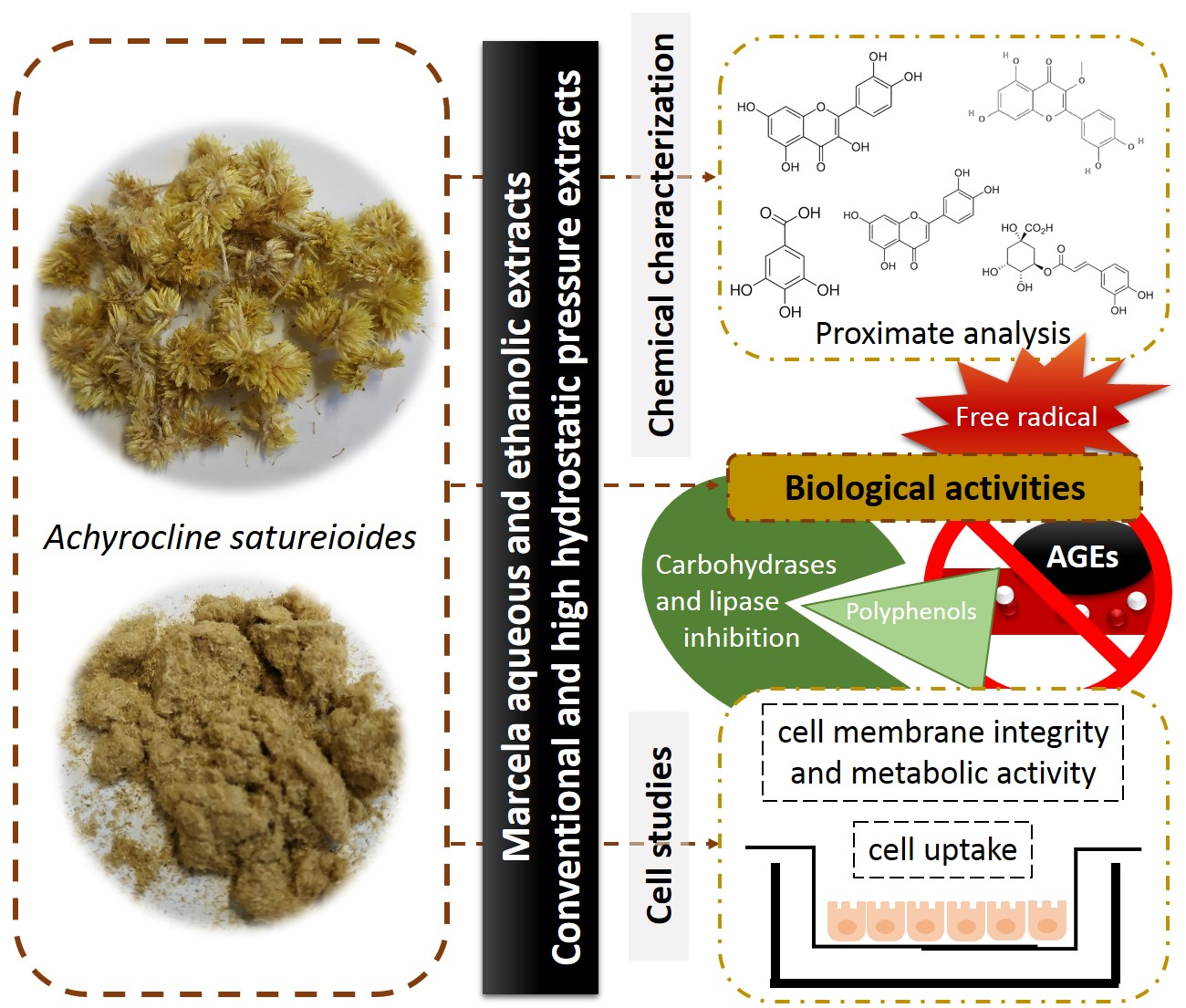
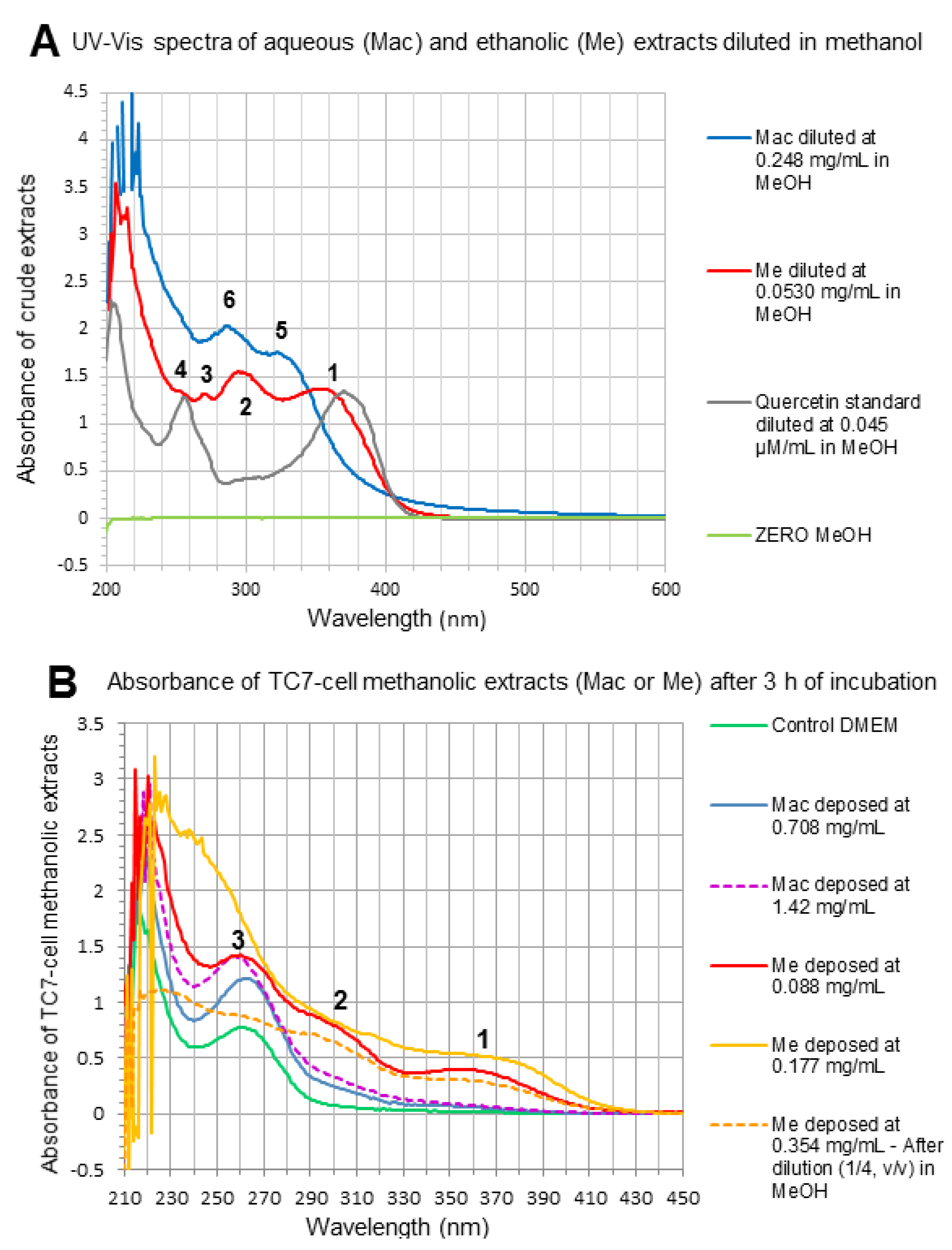
2.3.3. RP-HPLC and RP-UHPLC Analyses
2.3.4. Antioxidant Capacity
2.3.5. Antidiabetic and Antiobesity Activities
2.3.6. TC7-Cell Culture and Sample Deposits
2.3.7. Determination of In Vitro TC7-Cell Membrane Integrity and Cell Metabolic Activity
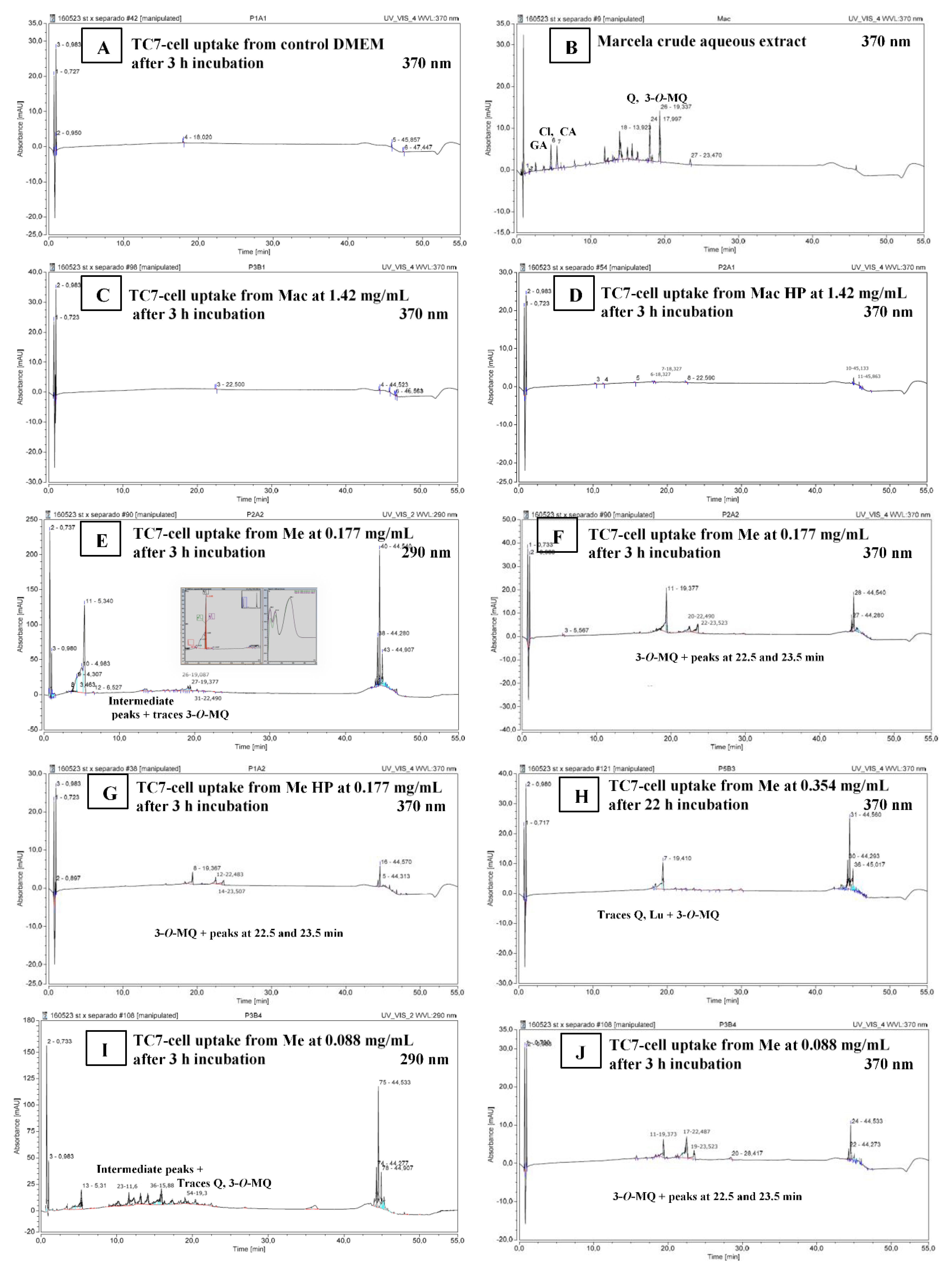
2.3.8. Marcela Bioactive Compounds’ Uptake
2.3.9. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition
3.2. Antioxidant Capacity
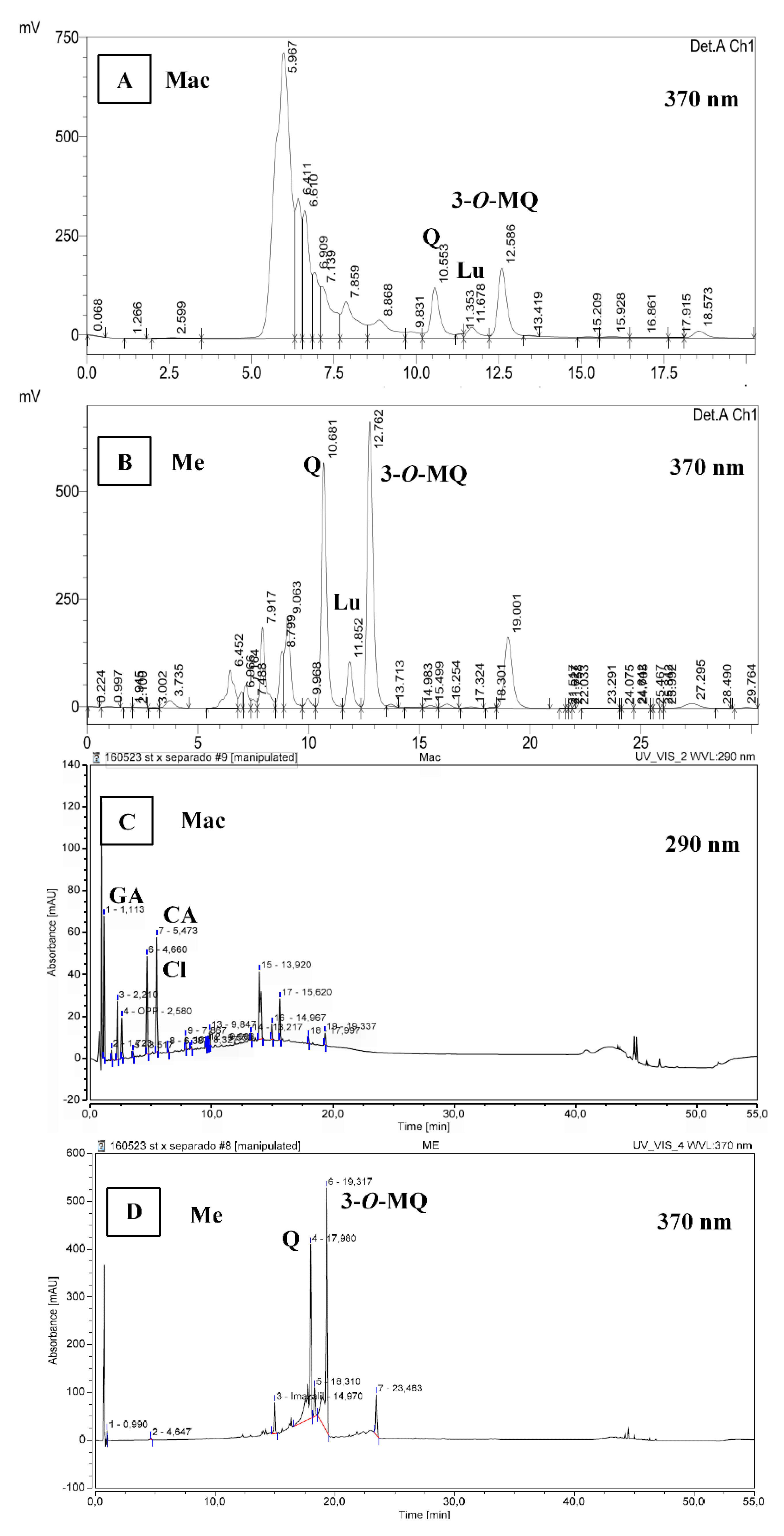
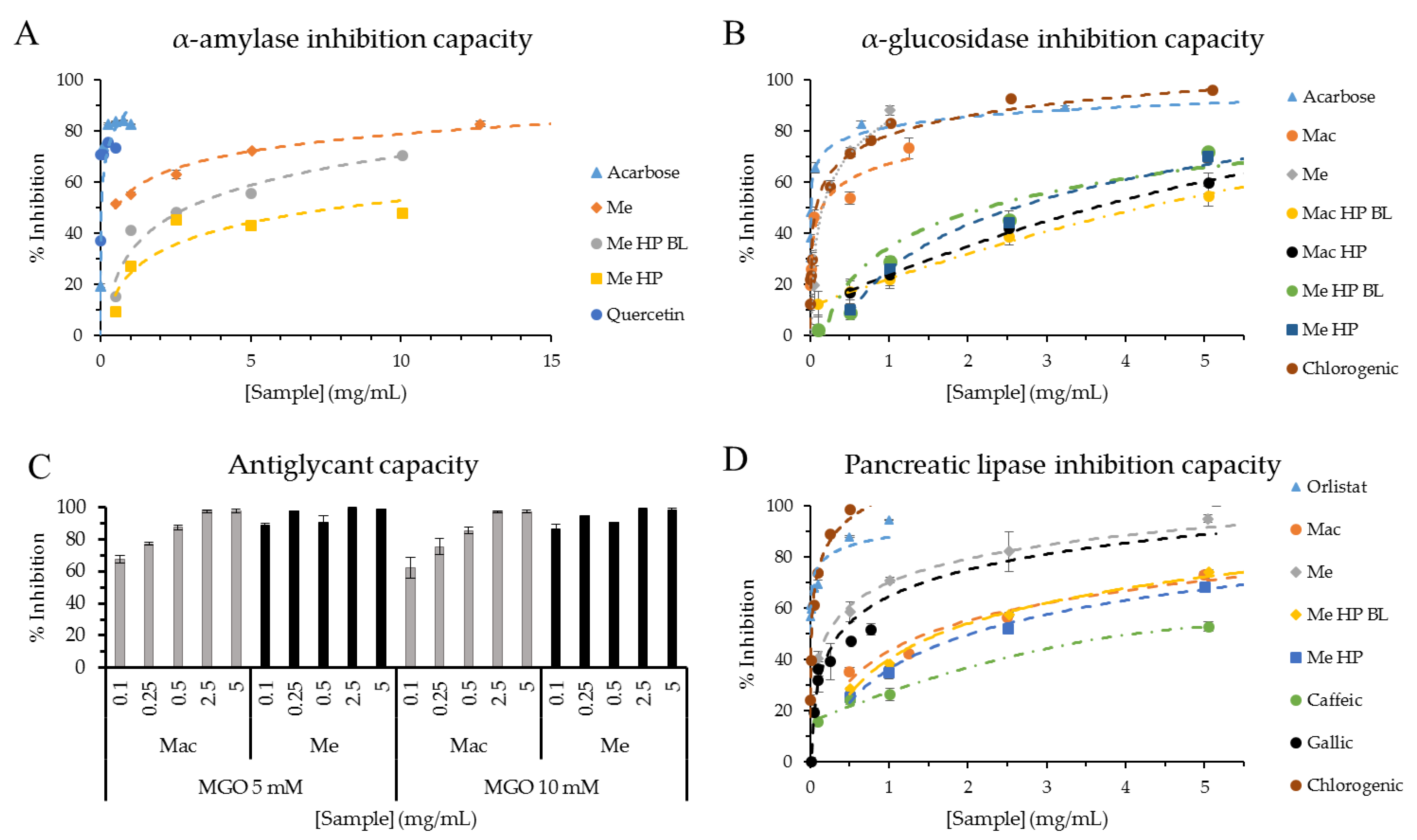
3.3. Antidiabetic Activities
3.4. Antiobesity Activity
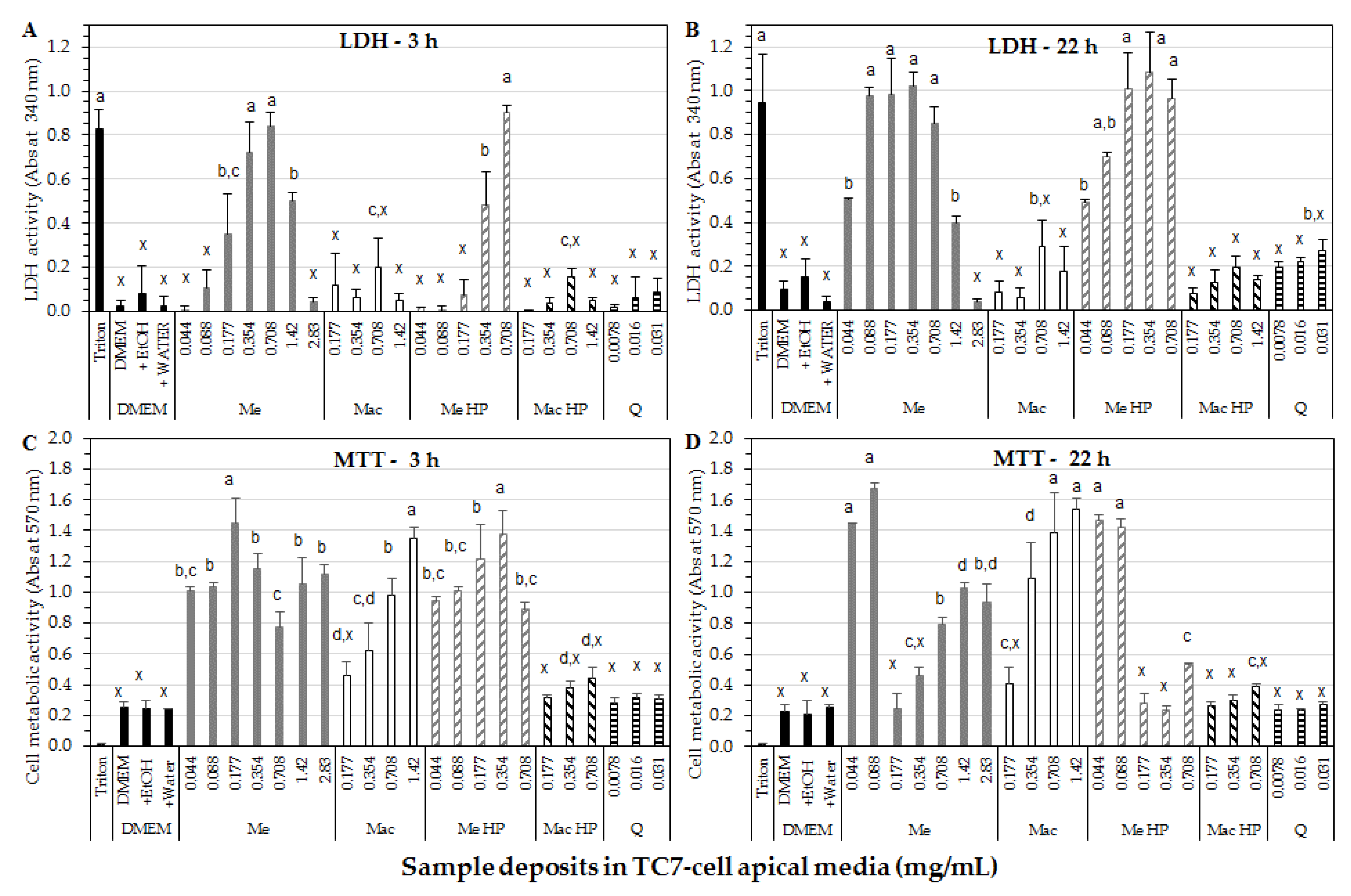
3.5. Cell Studies
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Noncommunicable Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/noncommunicable-diseases (accessed on 25 November 2019).
- Arredondo, M.F.; Blasina, F.; Echeverry, C.; Morquio, A.; Ferreira, M.; Abin-Carriquiry, J.A.; Lafon, L.; Dajas, F. Cytoprotection by Achyrocline satureioides (Lam) D.C. and some of its main flavonoids against oxidative stress. J. Ethnopharmacol. 2004, 91, 13–20. [Google Scholar] [CrossRef]
- Rivera, F.; Gervaz, E.; Sere, C.; Dajas, F. Toxicological studies of the aqueous extract from Achyrocline satureioides (Lam.) DC (Marcela). J. Ethnopharmacol. 2004, 95, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Retta, D.; Dellacassa, E.; Villamil, J.; Suárez, S.A.; Bandoni, A.L. Marcela, a promising medicinal and aromatic plant from Latin America: A review. Ind. Crops Prod. 2012, 38, 27–38. [Google Scholar] [CrossRef]
- De Souza, P.O.; Bianchi, S.E.; Figueiró, F.; Heimfarth, L.; Moresco, K.S.; Gonçalves, R.M.; Hoppe, J.B.; Klein, C.P.; Salbego, G.; Pens Gelain, D.; et al. Anticancer activity of flavonoids isolated from Achyrocline satureioides in gliomas cell lines. Toxicol. Vitr. 2018, 51, 23–33. [Google Scholar] [CrossRef] [PubMed]
- De Souza, K.C.B.; Schapoval, E.E.S.; Bassani, V.L. LC determination of flavonoids: Separation of quercetin, luteolin and 3-O-methylquercetin in Achyrocline satureioides preparations. J. Pharm. Biomed. Anal. 2002, 28, 771–777. [Google Scholar] [CrossRef]
- Galanakis, C.M. Emerging technologies for the production of nutraceuticals from agricultural by-products: A viewpoint of opportunities and challenges. Food Bioprod. Process. 2013, 91, 575–579. [Google Scholar] [CrossRef]
- Ignat, I.; Volf, I.; Popa, V.I. A critical review of methods for characterisation of polyphenolis compounds in fruits and vegetables. Food Chem. 2011, 126, 1821–1835. [Google Scholar] [CrossRef]
- Deliza, R.; Rosenthal, A.; Abadio, F.B.D.; Silva, C.H.O.; Castillo, C. Application of high pressure technology in the fruit juice processing: Benefits perceived by consumers. J. Food Eng. 2005, 67, 241–246. [Google Scholar] [CrossRef]
- Barba, F.J.; Zhu, Z.; Koubaa, M.; de Souza Sant’Ana, A.; Orlien, V. Green alternative methods for the extraction of antioxidant bioactive compounds from winery wastes and by-products: A review. Trends Food Sci. Technol. 2016, 49, 96–109. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1999. [Google Scholar]
- Reza, H.M.; Gias, Z.T.; Islam, P.; Sabnam, S.; Jain, P.; Hossain, M.H.; Alam, M.A. HPLC-DAD system-based phenolic content analysis and in vitro antioxidant activities of rice bran obtained from aush dhan (Oryza sativa) of Bangladesh. J. Food Biochem. 2015, 39, 462–470. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total Phenol Analysis: Automation and Comparison with Manual Methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Fernández-Fernández, A.M.; Iriondo-DeHond, A.; Dellacassa, E.; Medrano-Fernandez, A.; del Castillo, M.D. Assessment of antioxidant, antidiabetic, antiobesity, and anti-inflammatory properties of a Tannat winemaking by-product. Eur. Food Res. Technol. 2019, 245, 1539–1551. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Bartolomé, B.; Gómez-Cordovés, C. Antioxidant properties of commercial grape juices and vinegars. Food Chem. 2005, 93, 325–330. [Google Scholar] [CrossRef]
- Li, K.; Yao, F.; Du, J.; Deng, X.; Li, C. Persimmon Tannin Decreased the Glycemic Response through Decreasing the Digestibility of Starch and Inhibiting α-Amylase, α-Glucosidase, and Intestinal Glucose Uptake. J. Agric. Food Chem. 2018, 66, 1629–1637. [Google Scholar] [CrossRef]
- Fernández-Fernández, A.M.; Iriondo-DeHond, A.; Nardin, T.; Larcher, R.; Dellacassa, E.; Medrano-Fernandez, A.; del Castillo, M.D. In Vitro Bioaccessibility of Extractable Compounds from Tannat Grape Skin Possessing Health Promoting Properties with Potential to Reduce the Risk of Diabetes. Foods 2020, 9, 1575. [Google Scholar] [CrossRef]
- Starowicz, M.; Zieliński, H. Inhibition of Advanced Glycation End-Product Formation by High Antioxidant-Leveled Spices Commonly Used in European Cuisine. Antioxidants 2019, 8, 100. [Google Scholar] [CrossRef] [PubMed]
- Benzaria, A.; Maresca, M.; Taieb, N.; Dumay, E. Interaction of curcumin with phosphocasein micelles processed or not by dynamic high-pressure. Food Chem. 2013, 138, 2327–2337. [Google Scholar] [CrossRef]
- Benzaria, A.; Gràcia-Julià, A.; Picart-Palmade, L.; Hue, P.; Chevalier-Lucia, D.; Marti-Mestres, G.; Hodor, N.; Dumay, E. UHPH-processed O/W submicron emulsions stabilised with a lipid-based surfactant: Physicochemical characteristics and behaviour on in vitro TC7-cell monolayers and ex vivo pig’s ear skin. Colloids Surf. B Biointerfaces 2014, 116, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Zorzi, G.K.; Caregnato, F.; Cláudio, J.; Moreira, F.; Ferreira Teixeira, H.; Luis, E.; Carvalho, S. Antioxidant Effect of Nanoemulsions Containing Extract of Achyrocline satureioides (Lam) D.C.—Asteraceae. AAPS PharmSciTech 2015, 17, 844–850. [Google Scholar] [CrossRef]
- Martínez-Busi, M.; Arredondo, F.; González, D.; Echeverry, C.; Vega-Teijido, M.A.; Carvalho, D.; Rodríguez-Haralambides, A.; Rivera, F.; Dajas, F.; Abin-Carriquiry, J.A. Purification, structural elucidation, antioxidant capacity and neuroprotective potential of the main polyphenolic compounds contained in Achyrocline satureioides (Lam) D.C. (Compositae). Bioorg. Med. Chem. 2019, 27, 2579–2591. [Google Scholar] [CrossRef]
- Polydoro, M.; de Souza, K.C.; Andrades, M.; Da Silva, E.; Bonatto, F.; Heydrich, J.; Dal-Pizzol, F.; Schapoval, E.E.; Bassani, V.; Moreira, J.C. Antioxidant, a pro-oxidant and cytotoxic effects of Achyrocline satureioides extracts. Life Sci. 2004, 74, 2815–2826. [Google Scholar] [CrossRef]
- Guss, K.L.; Pavanni, S.; Prati, B.; Dazzi, L.; De Oliveira, J.P.; Nogueira, B.V.; Pereira, T.M.C.; Fronza, M.; Endringer, D.C.; Scherer, R. Ultrasound-assisted extraction of Achyrocline satureioides prevents contrast-induced nephropathy in mice. Ultrason. Sonochem. 2017, 37, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, G.E.; Norbedo, C.; Coussio, J.D. Polyphenols from Achyrocline satureioides. Phytochemistry 1981, 20, 2053–2054. [Google Scholar] [CrossRef]
- Grassi-Zampieron, R.; França, L.V.; Carollo, C.A.; do Carmo Vieira, M.; Oliveros-Bastidas, A.; de Siqueira, J.M. Comparative profiles of Achyrocline alata (Kunth) DC. and A. satureioides (Lam.) DC., Asteraceae, applying HPLC-DAD-MS. Braz. J. Pharmacogn. 2010, 20, 575–579. [Google Scholar] [CrossRef]
- Ferraro, G.; Anesini, C.; Ouviña, A.; Retta, D.; Filip, R.; Gattuso, M.; Gattuso, S.; Hnatyszyn, O.; Bandoni, A. Total Phenolic Content and Antioxidant Activity of Extracts of Achyrocline satureoides Flowers from Different Zones in Argentina. Lat. Am. J. Pharm. 2008, 27, 626–628. [Google Scholar]
- Irazusta, A.; Caccavello, R.; Panizzolo, L.; Gugliucci, A.; Medrano, A. The potential use of Mentha x piperita L., Peumus boldus Mol. and Baccharis trimera Iless. extracts as functional food ingredients. Int. J. Food Nutr. Res. 2018, 2, 14. [Google Scholar]
- Tang, K.S.C.; Konczak, I.; Zhao, J. Identification and quantification of phenolics in Australian native mint (Mentha australis R. Br.). Food Chem. 2016, 192, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Nyambe-Silavwe, H.; Villa-Rodriguez, J.A.; Ifie, I.; Holmes, M.; Aydin, E.; Jensen, J.M.; Williamson, G. Inhibition of human α-amylase by dietary polyphenols. J. Funct. Foods 2015, 19, 723–732. [Google Scholar] [CrossRef]
- Trinh, B.T.D.; Staerk, D.; Jäger, A.K. Screening for potential α-glucosidase and α-amylase inhibitory constituents from selected Vietnamese plants used to treat type 2 diabetes. J. Ethnopharmacol. 2016, 186, 189–195. [Google Scholar] [CrossRef]
- Figueiredo-González, M.; Grosso, C.; Valentão, P.; Andrade, P.B. α-Glucosidase and α-amylase inhibitors from Myrcia spp.: A stronger alternative to acarbose? J. Pharm. Biomed. Anal. 2016, 118, 322–327. [Google Scholar] [CrossRef]
- Kashchenko, N.I.; Olennikov, D.N. Phenolome of asian agrimony tea (agrimonia asiatica juz., rosaceae): LC-MS profile, α-glucosidase inhibitory potential and stability. Foods 2020, 9, 1348. [Google Scholar] [CrossRef]
- Tran, N.; Tran, M.; Truong, H.; Le, L. Spray-Drying Microencapsulation of High Concentration of Bioactive Compounds Fragments from Euphorbia hirta L. Extract and Their Effect on Diabetes Mellitus. Foods 2020, 9, 881. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Tomar, M.; Amarowicz, R.; Saurabh, V.; Nair, M.S.; Maheshwari, C.; Sasi, M.; Prajapati, U.; Hasan, M.; Singh, S.; et al. Guava (Psidium guajava L.) Leaves: Nutritional Composition, Phytochemical Profile, and Health-Promoting Bioactivities. Foods 2021, 10, 752. [Google Scholar] [CrossRef]
- Ramkissoon, J.; Mahomoodally, M.; Ahmed, N.; Subratty, A. Antioxidant and anti-glycation activities correlates with phenolic composition of tropical medicinal herbs. Asian Pac. J. Trop. Med. 2013, 6, 561–569. [Google Scholar] [CrossRef]
- Zheng, C.D.; Duan, Y.Q.; Gao, J.M.; Ruan, Z.G. Screening for Anti-lipase Properties of 37 Traditional Chinese Medicinal Herbs. J. Chin. Med. Assoc. 2010, 73, 319–324. [Google Scholar] [CrossRef]
- Tian, X.-J.; Yang, X.-W.; Yang, X.; Wang, K. Studies of intestinal permeability of 36 flavonoids using Caco-2 cell monolayer model. Int. J. Pharm. 2009, 367, 58–64. [Google Scholar] [CrossRef]
- Walle, T. Methylation of dietary flavones increases their metabolic stability and chemopreventive effects. Int. J. Mol. Sci. 2009, 10, 5002–5019. [Google Scholar] [CrossRef]
- Santos, J.; Oliveira, M.B.P.P.; Ibáñez, E.; Herrero, M. Phenolic profile evolution of different ready-to-eat baby-leaf vegetables during storage. J. Chromatogr. A 2014, 1327, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Bruno, R.S. Endogenous and exogenous mediators of quercetin bioavailability. J. Nutr. Biochem. 2015, 26, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Najgebauer-Lejko, D.; Sady, M.; Grega, T.; Walczycka, M. The impact of tea supplementation on microflora, pH and antioxidant capacity of yoghurt. Int. Dairy J. 2011, 21, 568–574. [Google Scholar] [CrossRef]
- Dabija, A.; Codinǎ, G.G.; Ropciuc, S.; Gâtlan, A.M.; Rusu, L. Assessment of the antioxidant activity and quality attributes of yogurt enhanced with wild herbs extracts. J. Food Qual. 2018, 2018. [Google Scholar] [CrossRef]
- Alirezalu, K.; Hesari, J.; Yaghoubi, M.; Khaneghah, A.M.; Alirezalu, A.; Pateiro, M.; Lorenzo, J.M. Combined effects of ε-polylysine and ε-polylysine nanoparticles with plant extracts on the shelf life and quality characteristics of nitrite-free frankfurter-type sausages. Meat Sci. 2021, 172, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Namal Senanayake, S.P.J. Green tea extract: Chemistry, antioxidant properties and food applications—A review. J. Funct. Foods 2013, 5, 1529–1541. [Google Scholar] [CrossRef]
- Quirós-Sauceda, A.E.; Palafox-Carlos, H.; Sáyago-Ayerdi, S.G.; Ayala-Zavala, J.F.; Bello-Perez, L.A.; Álvarez-Parrilla, E.; De La Rosa, L.A.; González-Córdova, A.F.; González-Aguilar, G.A. Dietary fiber and phenolic compounds as functional ingredients: Interaction and possible effect after ingestion. Food Funct. 2014, 5, 1063–1072. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total Polyphenol Content (mg GAE/g Extract) | ABTS (µmol TE/mg Extract) | ORAC-FL (µmol TE/mg Extract) |
|---|---|---|---|
| Mac | 83.36 ± 6.69 c | 2.71 ± 0.27 a | 2.17 ± 0.10 d |
| Me | 108.79 ± 16.47 d | 4.19 ± 0.43 b | 2.30 ± 0.07 e |
| Mac HP BL | 44.07 ± 1.97 a | 2.06 ± 0.17 a | 0.34 ± 0.08 a |
| Mac HP | 42.68 ± 3.55 a | 2.30 ± 0.18 a | 0.54 ± 0.04 b |
| Me HP BL | 63.00 ± 3.12 b | 1.84 ± 0.20 a | 1.11 ± 0.08 c |
| Me HP | 58.23 ± 4.51 b | 1.88 ± 0.13 a | 1.08 ± 0.07 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Fernández, A.M.; Dumay, E.; Lazennec, F.; Migues, I.; Heinzen, H.; Lema, P.; López-Pedemonte, T.; Medrano-Fernandez, A. Antioxidant, Antidiabetic, and Antiobesity Properties, TC7-Cell Cytotoxicity and Uptake of Achyrocline satureioides (Marcela) Conventional and High Pressure-Assisted Extracts. Foods 2021, 10, 893. https://doi.org/10.3390/foods10040893
Fernández-Fernández AM, Dumay E, Lazennec F, Migues I, Heinzen H, Lema P, López-Pedemonte T, Medrano-Fernandez A. Antioxidant, Antidiabetic, and Antiobesity Properties, TC7-Cell Cytotoxicity and Uptake of Achyrocline satureioides (Marcela) Conventional and High Pressure-Assisted Extracts. Foods. 2021; 10(4):893. https://doi.org/10.3390/foods10040893
Chicago/Turabian StyleFernández-Fernández, Adriana Maite, Eliane Dumay, Françoise Lazennec, Ignacio Migues, Horacio Heinzen, Patricia Lema, Tomás López-Pedemonte, and Alejandra Medrano-Fernandez. 2021. "Antioxidant, Antidiabetic, and Antiobesity Properties, TC7-Cell Cytotoxicity and Uptake of Achyrocline satureioides (Marcela) Conventional and High Pressure-Assisted Extracts" Foods 10, no. 4: 893. https://doi.org/10.3390/foods10040893
APA StyleFernández-Fernández, A. M., Dumay, E., Lazennec, F., Migues, I., Heinzen, H., Lema, P., López-Pedemonte, T., & Medrano-Fernandez, A. (2021). Antioxidant, Antidiabetic, and Antiobesity Properties, TC7-Cell Cytotoxicity and Uptake of Achyrocline satureioides (Marcela) Conventional and High Pressure-Assisted Extracts. Foods, 10(4), 893. https://doi.org/10.3390/foods10040893