
In Vivo Genotoxicity Evaluation of a Stilbene Extract Prior to Its Use as a Natural Additive: A Combination of the Micronucleus Test and the Comet Assay

 , ,
, ,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Supplies and Chemicals
2.2. Animal Hosting and Nourishing Conditions
2.3. Stilbene Enriched Extract
2.4. Experimental Design and Treatment
2.5. Sample Collection
2.6. MN Assay
2.7. Isolation of Single-Cell Suspensions for the Comet Assay
2.8. Standard and Enzyme-Modified Comet Assay
2.9. Determination of Stilbenes in Plasma and Tissues
2.9.1. Standards Solutions
2.9.2. Stilbenes Extraction from Plasma and Liver Samples
2.9.3. Extracts Analysis by UPLC-HESI-HRMS
2.10. Histopathogical Analysis
2.11. Statistical Analyses
3. Results
3.1. Micronucleus Test
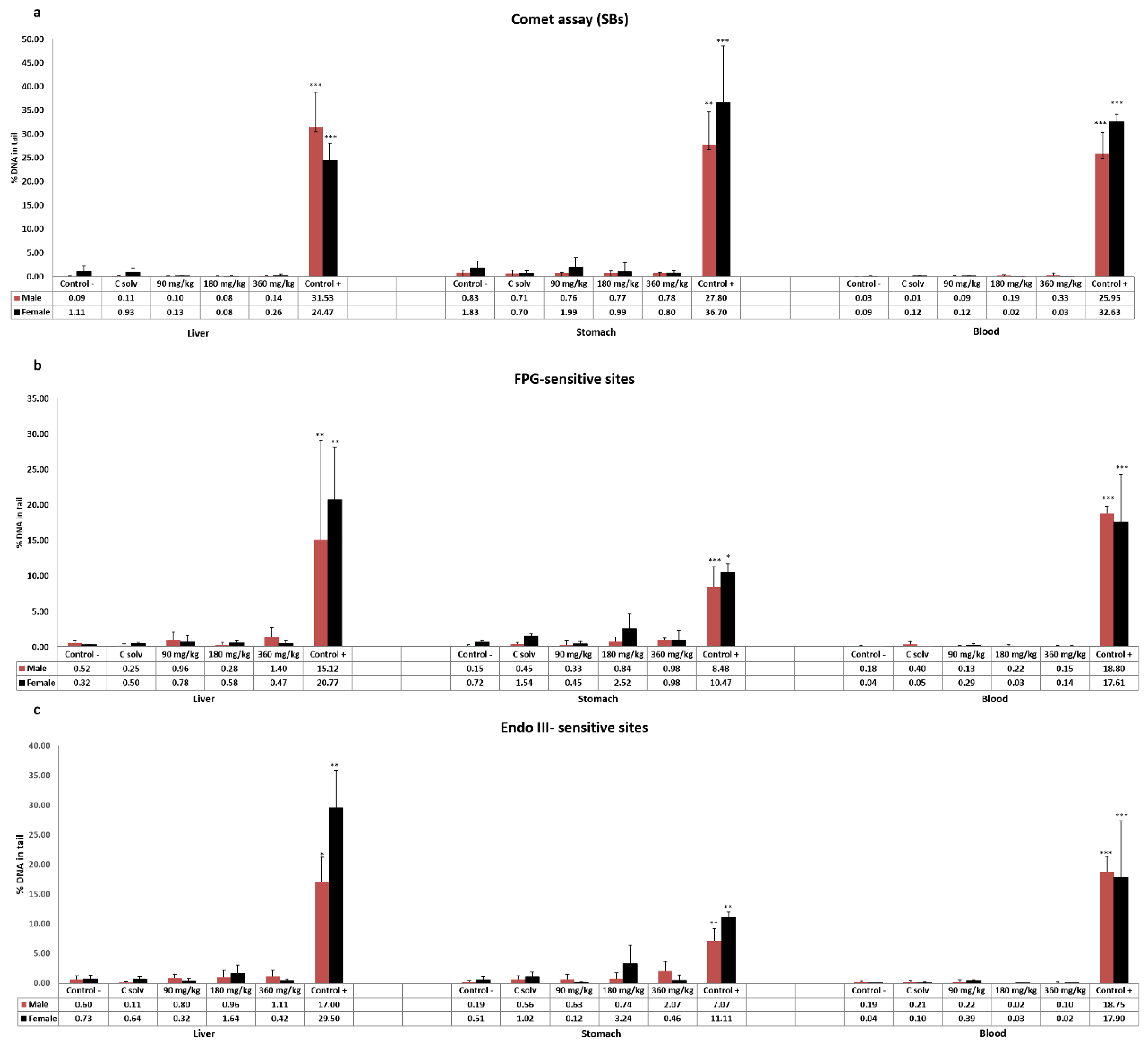
3.2. Standard and Enzyme-Modified Comet Assay
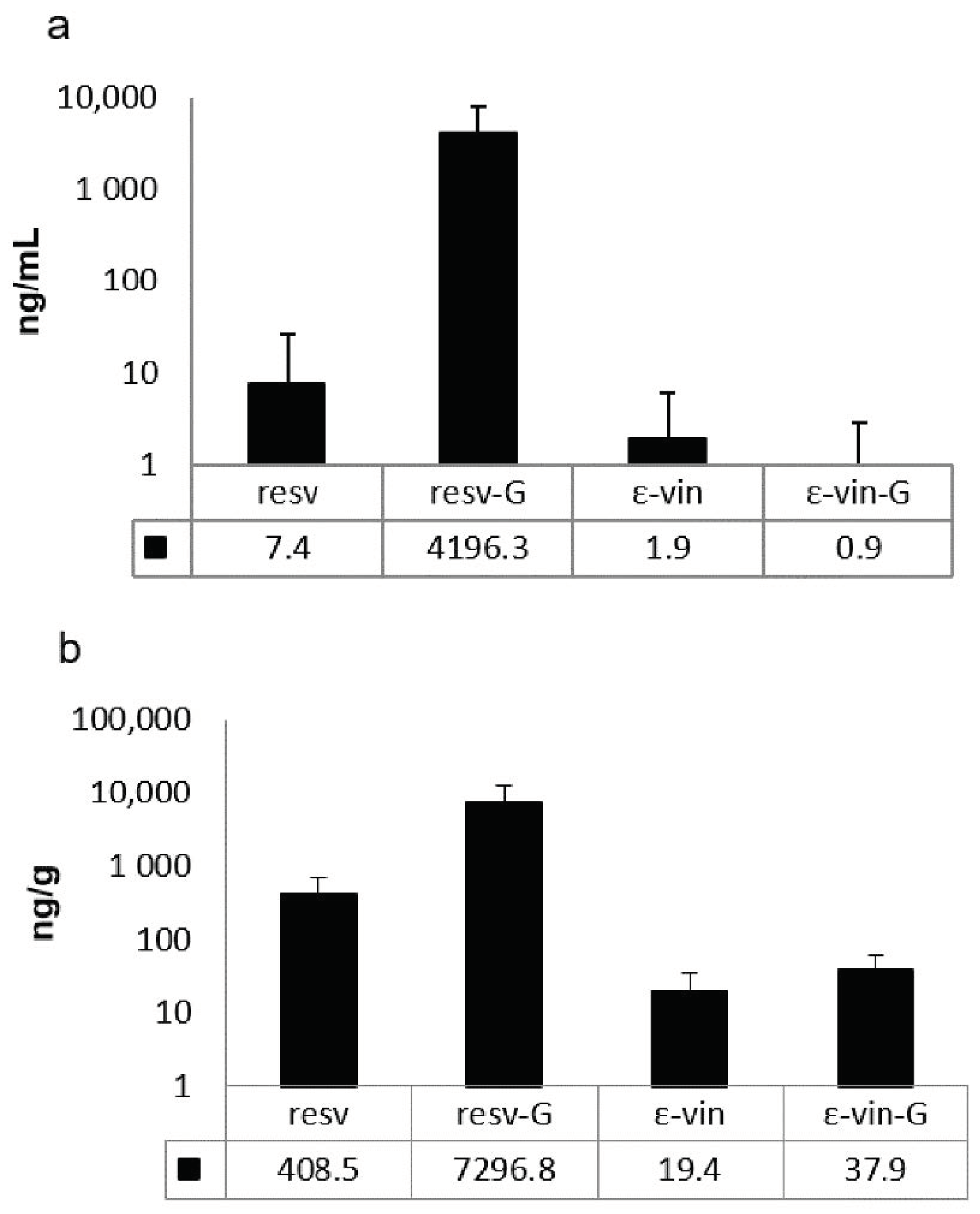
3.3. Presence of ε-Viniferin, Trans-Resveratrol, and Its Derived Compounds in Tissue
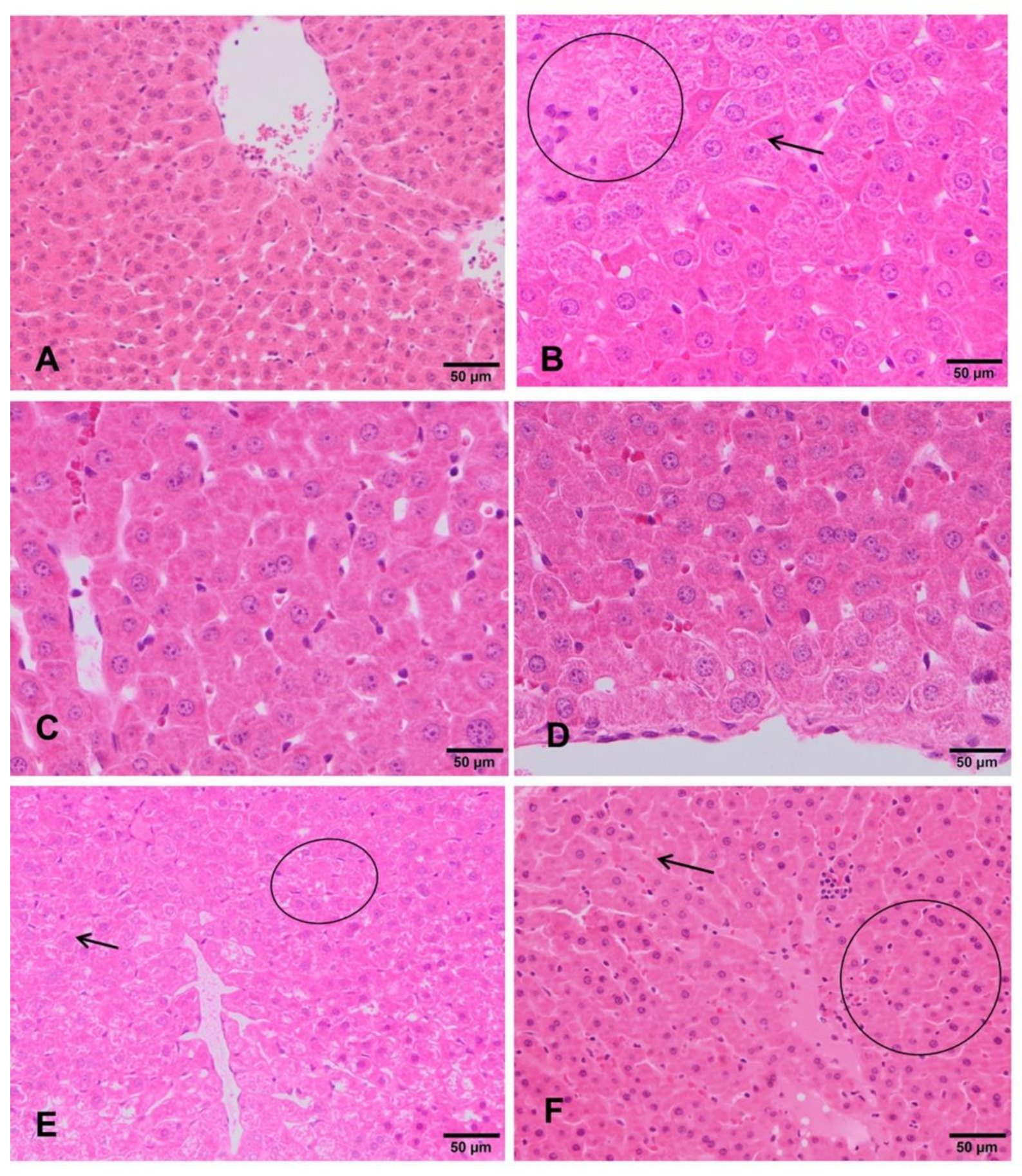
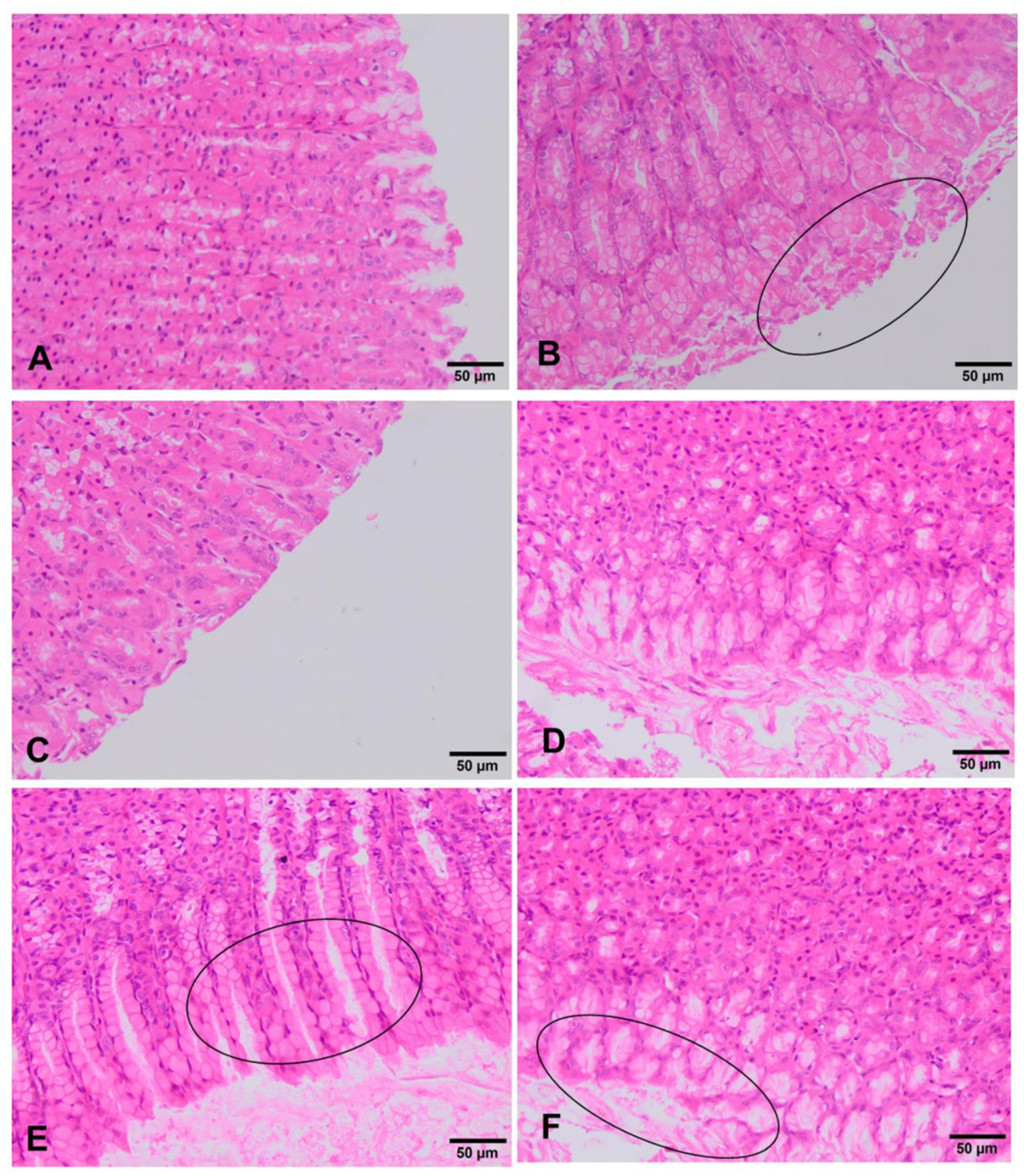
3.4. Clinical and Histopathological Analysis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carocho, M.; Morales, P.; Ferreira, I.C.F.R. Natural Food Additives: Quo Vadis? Trends Food Sci. Technol. 2015, 45, 284–295. [Google Scholar] [CrossRef]
- Schieber, A. Side Streams of Plant Food Processing as a Source of Valuable Compounds: Selected Examples. Annu. Rev. Food Sci. Technol. 2017, 8, 97–112. [Google Scholar] [CrossRef]
- EFSA. ANS Panel (EFSA Panel on Food Additives and Nutrient Sources Added to Foods). Scientific Opinion on the Re-evaluation of Sulfur Dioxide (E 220), Sodium Sulfite (E 221), Sodium Bisulfite (E 222), Sodium Metabisulfite (E 223), Potassium Metabisulfite (E 224), Calcium Sulfite (E 226), Calcium Bisulfite (E 227) and Potassium Bisulfite. EFSA J. 2016, 14. [Google Scholar] [CrossRef]
- Guerrero, R.F.; Cantos-Villar, E. Demonstrating the Efficiency of Sulphur Dioxide Replacements in Wine: A Parameter Review. Trends Food Sci. Technol. 2015, 42, 27–43. [Google Scholar] [CrossRef]
- Raposo, R.; Ruiz-Moreno, M.J.; Garde-Cerdán, T.; Puertas, B.; Moreno-Rojas, J.M.; Gonzalo-Diago, A.; Guerrero, R.; Ortíz, V.; Cantos-Villar, E. Grapevine-Shoot Stilbene Extract as a Preservative in Red Wine. Food Chem. 2016, 197, 1102–1111. [Google Scholar] [CrossRef]
- Baydar, H.; Saǧdiç, O.; Özkan, G.; Karadoǧan, T. Antibacterial Activity and Composition of Essential Oils from Origanum, Thymbra and Satureja Species with Commercial Importance in Turkey. Food Control 2004, 15, 169–172. [Google Scholar] [CrossRef]
- Baydar, N.G.; Sagdic, O.; Ozkan, G.; Cetin, S. Determination of Antibacterial Effects and Total Phenolic Contents of Grape (Vitis Vinifera L.) Seed Extracts. Int. J. Food Sci. Technol. 2006, 41, 799–804. [Google Scholar] [CrossRef]
- Sagdic, O.; Ozturk, I.; Ozkan, G.; Yetim, H.; Ekici, L.; Yilmaz, M.T. RP-HPLC-DAD Analysis of Phenolic Compounds in Pomace Extracts from Five Grape Cultivars: Evaluation of Their Antioxidant, Antiradical and Antifungal Activities in Orange and Apple Juices. Food Chem. 2011, 126, 1749–1758. [Google Scholar] [CrossRef] [PubMed]
- Anastasiadi, M.; Pratsinis, H.; Kletsas, D.; Skaltsounis, A.L.; Haroutounian, S.A. Grape Stem Extracts: Polyphenolic Content and Assessment of Their in Vitro Antioxidant Properties. LWT Food Sci. Technol. 2012, 48, 316–322. [Google Scholar] [CrossRef]
- Biais, B.; Krisa, S.; Cluzet, S.; Da Costa, G.; Waffo-Teguo, P.; Mérillon, J.M.; Richard, T. Antioxidant and Cytoprotective Activities of Grapevine Stilbenes. J. Agric. Food Chem. 2017, 65, 4952–4960. [Google Scholar] [CrossRef]
- Müller, C.; Ullmann, K.; Wilkens, A.; Winterhalter, P.; Toyokuni, S.; Steinberg, P. Potent Antioxidative Activity of Vineatrol®30 Grapevine-Shoot Extract. Biosci. Biotechnol. Biochem. 2009, 73, 1831–1836. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, A.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Bartolomé, B. Comparative Study of the Inhibitory Effects of Wine Polyphenols on the Growth of Enological Lactic Acid Bacteria. Int. J. Food Microbiol. 2011, 145, 426–431. [Google Scholar] [CrossRef]
- Raposo, R.; Ruiz-Moreno, M.J.; Garde-Cerdán, T.; Puertas, B.; Moreno-Rojas, J.M.; Gonzalo-Diago, A.; Guerrero, R.F.; Ortiz, V.; Cantos-Villar, E. Effect of Hydroxytyrosol on Quality of Sulfur Dioxide-Free Red Wine. Food Chem. 2016, 192, 25–33. [Google Scholar] [CrossRef]
- Raposo, R.; Chinnici, F.; Ruiz-Moreno, M.J.; Puertas, B.; Cuevas, F.J.; Carbú, M.; Guerrero, R.F.; Ortíz-Somovilla, V.; Moreno-Rojas, J.M.; Cantos-Villar, E. Sulfur Free Red Wines through the Use of Grapevine Shoots: Impact on the Wine Quality. Food Chem. 2018, 243, 453–460. [Google Scholar] [CrossRef]
- Gutiérrez-Escobar, R.; Fernández-Marín, M.I.; Richard, T.; Fernández-Morales, A.; Carbú, M.; Cebrian-Tarancón, C.; Torija, M.J.; Puertas, B.; Cantos-Villar, E. Development and Characterization of a Pure Stilbene Extract from Grapevine Shoots for Use as a Preservative in Wine. Food Control 2021, 121. [Google Scholar] [CrossRef]
- Medrano-Padial, C.; Puerto, M.; Richard, T.; Cantos-Villar, E.; Pichardo, S. Protection and reversion role of a pure stilbene extract from grapevine shoot and its major compounds against an induced oxidative stress. J. Funct. Foods 2021, in press. [Google Scholar]
- Medrano-Padial, C.; Puerto, M.; del Mar Merchán-Gragero, M.; Moreno, F.J.; Richard, T.; Cantos-Villar, E.; Pichardo, S. Cytotoxicity Studies of a Stilbene Extract and Its Main Components Intended to Be Used as Preservative in the Wine Industry. Food Res. Int. 2020, 137. [Google Scholar] [CrossRef]
- EFSA Scientific Committee. Scientific Opinion on Genotoxicity Testing Strategies Applicable to Food and Feed Safety Assessment. EFSA J. 2011, 9. [Google Scholar] [CrossRef]
- Medrano-Padial, C.; Prieto, A.; Puerto, M.; Pichardo, S. In vitro assessment of the mutagenic and genotoxic potential of a pure stilbene extract. Food Chem. Toxicol. 2021, in press. [Google Scholar] [CrossRef]
- EFSA Scientific Committee. Clarification of Some Aspects Related to Genotoxicity Assessment. EFSA J. 2017, 15. [Google Scholar] [CrossRef]
- Collins, A.R. The Comet Assay for DNA Damage and Repair: Principles, Applications, and Limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Tatefuji, T.; Yanagihara, M.; Fukushima, S.; Hashimoto, K. Safety Assessment of Melinjo (Gnetum Gnemon L.) Seed Extract: Acute and Subchronic Toxicity Studies. Food Chem. Toxicol. 2014, 67, 230–235. [Google Scholar] [CrossRef] [PubMed]
- EFSA. ANS Panel (EFSA Panel on Food Additives and Nutrient Sources added to Foods). Guidance for Submission for Food Additive Evaluations. EFSA J. 2012, 10. [Google Scholar] [CrossRef]
- Bowen, D.E.; Whitwell, J.H.; Lillford, L.; Henderson, D.; Kidd, D.; Mc Garry, S.; Pearce, G.; Beevers, C.; Kirkland, D.J. Evaluation of a Multi-Endpoint Assay in Rats, Combining the Bone-Marrow Micronucleus Test, the Comet Assay and the Flow-Cytometric Peripheral Blood Micronucleus Test. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2011, 722, 7–19. [Google Scholar] [CrossRef]
- OECD. Test No. 474: Mammalian Erythrocyte Micronucleus Test; OECD Publishing: Paris, France, 2014. [Google Scholar]
- OECD. Test No. 489: In Vivo Mammalian Alkaline Comet Assay; OECD Publishing: Paris, France, 2014. [Google Scholar]
- Mughal, A.; Vikram, A.; Ramarao, P.; Jena, G.B. Micronucleus and Comet Assay in the Peripheral Blood of Juvenile Rat: Establishment of Assay Feasibility, Time of Sampling and the Induction of DNA Damage. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2010, 700, 86–94. [Google Scholar] [CrossRef]
- Mellado-García, P.; Puerto, M.; Prieto, A.I.; Pichardo, S.; Martín-Cameán, A.; Moyano, R.; Blanco, A.; Cameán, A.M. Genotoxicity of a Thiosulfonate Compound Derived from Allium Sp. Intended to Be Used in Active Food Packaging: In Vivo Comet Assay and Micronucleus Test. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2016, 800–801, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Llana-Ruiz-Cabello, M.; Maisanaba, S.; Puerto, M.; Prieto, A.I.; Pichardo, S.; Moyano, R.; González-Pérez, J.A.; Cameán, A.M. Genotoxicity Evaluation of Carvacrol in Rats Using a Combined Micronucleus and Comet Assay. Food Chem. Toxicol. 2016, 98, 240–250. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Medrano-Padial, C.; Llana-Ruiz-Cabello, M.; Catunescu, G.M.; Moyano, R.; Risalde, M.A.; Cameán, A.M.; Jos, Á. Cylindrospermopsin-Microcystin-LR Combinations May Induce Genotoxic and Histopathological Damage in Rats. Toxins 2020, 12, 348. [Google Scholar] [CrossRef]
- Lehman, A.J.; Fitzhugh, O.G. 100-fold margin of safety. Assoc. Food Drug Off. US Q Bull. 1954, 18, 33–35. [Google Scholar]
- Corcuera, L.A.; Vettorazzi, A.; Arbillaga, L.; Pérez, N.; Gil, A.G.; Azqueta, A.; González-Peñas, E.; García-Jalón, J.A.; López de Cerain, A. Genotoxicity of Aflatoxin B1 and Ochratoxin A after Simultaneous Application of the in Vivo Micronucleus and Comet Assay. Food Chem. Toxicol. 2015, 76, 116–124. [Google Scholar] [CrossRef]
- Llana-Ruiz-Cabello, M.; Puerto, M.; Maisanaba, S.; Guzmán-Guillén, R.; Pichardo, S.; Cameán, A.M. Use of Micronucleus and Comet Assay to Evaluate Evaluate the Genotoxicity of Oregano Essential Oil (Origanum Vulgare l. Virens) in Rats Orally Exposed for 90 Days. J. Toxicol. Environ. Heal. Part A Curr. Issues 2018, 81, 525–533. [Google Scholar] [CrossRef]
- Courtois, A.; Atgié, C.; Marchal, A.; Hornedo-Ortega, R.; Lapèze, C.; Faure, C.; Richard, T.; Krisa, S. Tissular Distribution and Metabolism of Trans-ε-Viniferin after Intraperitoneal Injection in Rat. Nutrients 2018, 10, 1660. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, D.; Pfuhler, S.; Tweats, D.; Aardema, M.; Corvi, R.; Darroudi, F.; Elhajouji, A.; Glatt, H.; Hastwell, P.; Hayashi, M.; et al. How to Reduce False Positive Results When Undertaking In Vitro Genotoxicity Testing and Thus Avoid Unnecessary Follow-up Animal Tests: Report of an ECVAM Workshop. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2007, 628, 31–55. [Google Scholar] [CrossRef] [PubMed]
- FDA. Guidance for Industry and Other Stakeholders Toxicological Principles for the Safety Assessment of Food Ingredients. Hum. Stud. 2007, 3, 1–286. [Google Scholar]
- Araldi, R.P.; de Melo, T.C.; Mendes, T.B.; de Sá Júnior, P.L.; Nozima, B.H.N.; Ito, E.T.; de Carvalho, R.F.; de Souza, E.B.; de Cassia Stocco, R. Using the Comet and Micronucleus Assays for Genotoxicity Studies: A Review. Biomed. Pharmacother. 2015, 72, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Hynes, G. Resveratrol Rat Micronucleus Test; DSM Nutritional Products Ltd.: Basel, Switzerland, 2007; DNO 0015/063572; Huntingdon Life Sciences Limited, 25 May 2007. [Google Scholar]
- EFSA. NDA Panel (EFSA Panel on Dietetic Products, Nutrition and Allergies). Safety of Synthetic Trans-resveratrol as a Novel Food Pursuant to Regulation (EC) No. 258/97. EFSA J. 2016, 14, 1–30. [Google Scholar] [CrossRef]
- Williams, L.D.; Burdock, G.A.; Edwards, J.A.; Beck, M.; Bausch, J. Safety Studies Conducted on High-Purity Trans-Resveratrol in Experimental Animals. Food Chem. Toxicol. 2009, 47, 2170–2182. [Google Scholar] [CrossRef]
- Møller, H.D.; Mohiyuddin, M.; Prada-Luengo, I.; Sailani, M.R.; Halling, J.F.; Plomgaard, P.; Maretty, L.; Hansen, A.J.; Snyder, M.P.; Pilegaard, H.; et al. Circular DNA Elements of Chromosomal Origin Are Common in Healthy Human Somatic Tissue. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.; Dušinská, M.; Franklin, M.; Somorovská, M.; Petrovská, H.; Duthie, S.; Fillion, L.; Panayiotidis, M.; Rašlová, K.; Vaughan, N. Comet Assay in Human Biomonitoring Studies: Reliability, Validation, and Applications. Environ. Mol. Mutagen. 1997, 30, 139–146. [Google Scholar] [CrossRef]
- Attia, S.M. Influence of Resveratrol on Oxidative Damage in Genomic DNA and Apoptosis Induced by Cisplatin. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2012, 741, 22–31. [Google Scholar] [CrossRef]
- Aydin, S.; Bacanli, M.; Taner, G.; Şahin, T.; Başaran, A.A.; Başaran, N. Protective Effects of Resveratrol on Sepsis-Induced DNA Damage in the Lymphocytes of Rats. Hum. Exp. Toxicol. 2013, 32, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- Alarcón De La Lastra, C.; Villegas, I. Resveratrol as an Antioxidant and Pro-Oxidant Agent: Mechanisms and Clinical Implications. Biochem. Soc. Trans. 2007, 35, 1156–1160. [Google Scholar] [CrossRef]
- Erdogan, A.; Ozkan, A. A Comparative Study of Cytotoxic, Membrane and DNA Damaging Effects of Origanum Majorana’s Essential Oil and Its Oxygenated Monoterpene Component Linalool on Parental and Epirubicin-Resistant H1299. Cells. Biol. 2013, 68, 754–761. [Google Scholar] [CrossRef]
- Ferguson, L.R. Role of Plant Polyphenols in Genomic Stability. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2001, 475, 89–111. [Google Scholar] [CrossRef]
- Mena, S.; Ortega, A.; Estrela, J.M. Oxidative Stress in Environmental-Induced Carcinogenesis. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2009, 674, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Plauth, A.; Geikowski, A.; Cichon, S.; Wowro, S.J.; Liedgens, L.; Rousseau, M.; Weidner, C.; Fuhr, L.; Kliem, M.; Jenkins, G.; et al. Hormetic Shifting of Redox Environment by Pro-Oxidative Resveratrol Protects Cells against Stress. Free Radic. Biol. Med. 2016, 99, 608–622. [Google Scholar] [CrossRef]
- Cottart, C.H.; Nivet-Antoine, V.; Laguillier-Morizot, C.; Beaudeux, J.L. Resveratrol Bioavailability and Toxicity in Humans. Mol. Nutr. Food Res. 2010, 54, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Kapetanovic, I.M.; Muzzio, M.; Huang, Z.; Tompson, T.N.; McCormick, D.L. Pharmacokinetics, oral bioavailability, and metabolic profile of resveratrol and its dimethylether analog, pterostilbene, in rats. Cancer Chemother. Pharmacol. 2011, 68, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, E.; Gibbs, S.T.; South, N.; Pierfelice, J.; Burback, B.; Germolec, D.; Waidyanatha, S. Comparative Toxicokinetics of Trans-Resveratrol and Its Major Metabolites in Harlan Sprague Dawley Rats and B6C3F1/N Mice Following Oral and Intravenous Administration. Toxicol. Appl. Pharmacol. 2020, 394, 114962. [Google Scholar] [CrossRef]
- Rege, S.D.; Geetha, T.; Griffin, G.D.; Broderick, T.L.; Babu, J.R. Neuroprotective Effects of Resveratrol in Alzheimer Disease Pathology. Front. Aging Neurosci. 2014, 6, 1–27. [Google Scholar] [CrossRef]
- Kim, J.; Min, J.S.; Kim, D.; Zheng, Y.F.; Mailar, K.; Choi, W.J.; Lee, C.; Bae, S.K. A Simple and Sensitive Liquid Chromatography–Tandem Mass Spectrometry Method for Trans-ε-Viniferin Quantification in Mouse Plasma and Its Application to a Pharmacokinetic Study in Mice. J. Pharm. Biomed. Anal. 2017, 134, 116–121. [Google Scholar] [CrossRef] [PubMed]
- EFSA Scientific Committee. Guidance on Safety Assessment of Botanicals and Botanical Preparations Intended for Use as Ingredients in Food Supplements. EFSA J. 2009, 7. [Google Scholar] [CrossRef]
- Andres-Lacueva, C.; MacArulla, M.T.; Rotches-Ribalta, M.; Boto-Ordóñez, M.; Urpi-Sarda, M.; Rodríguez, V.M.; Portillo, M.P. Distribution of Resveratrol Metabolites in Liver, Adipose Tissue, and Skeletal Muscle in Rats Fed Different Doses of This Polyphenol. J. Agric. Food Chem. 2012, 60, 4833–4840. [Google Scholar] [CrossRef]
- Böhmdorfer, M.; Szakmary, A.; Schiestl, R.H.; Vaquero, J.; Riha, J.; Brenner, S.; Thalhammer, T.; Szekeres, T.; Jäger, W. Involvement of UDP-Glucuronosyltransferases and Sulfotransferases in the Excretion and Tissue Distribution of Resveratrol in Mice. Nutrients 2017, 9, 1347. [Google Scholar] [CrossRef]
- Courtois, A.; Jourdes, M.; Dupin, A.; Lapèze, C.; Renouf, E.; Biais, B.; Teissedre, P.L.; Mérillon, J.M.; Richard, T.; Krisa, S. In Vitro Glucuronidation and Sulfation of ϵ-Viniferin, a Resveratrol Dimer, in Humans and Rats. Molecules 2017, 22, 733. [Google Scholar] [CrossRef]
- Marier, J.F.; Vachon, P.; Gritsas, A.; Zhang, J.; Moreau, J.P.; Ducharme, M.P. Metabolism and Disposition of Resveratrol in Rats: Extent of Absorption, Glucuronidation, and Enterohepatic Recirculation Evidenced by a Linked-Rat Model. J. Pharmacol. Exp. Ther. 2002, 302, 369–373. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Sex | n | Doses | PCE/NCE | PCE/Total | % MN |
|---|---|---|---|---|---|---|
| Negative Control | ♂ | 5 | 1.15 ± 0.17 | 0.53 ± 0.04 | 0.87 ± 0.08 | |
| ♀ | 5 | 1.78 ± 0.10 | 0.64 ± 0.03 | 0.99 ± 0.08 | ||
| Solvent Control | ♂ | 5 | 1.09 ± 0.19 | 0.51 ± 0.04 | 0.91 ± 0.16 | |
| ♀ | 5 | 2.35 ± 0.73 | 0.69 ± 0.07 | 1.03 ± 0.07 | ||
| Positive Control | ♂ | 3 | 200 mg/kg | 0.50 ± 0.08 ** | 0.33 ± 0.03 * | 2.35 ± 0.06 *** |
| ♀ | 3 | 0.68 ± 0.36 ** | 0.39 ± 0.13 * | 2.39 ± 0.05 *** | ||
| ST-99 extract | ♂ | 5 | 90 mg/kg b.w. | 0.94 ± 0.49 | 0.46 ± 0.12 | 0.86 ± 0.12 |
| ♀ | 5 | 1.12 ± 0.39 | 0.51 ± 0.08 | 0.89 ± 0.14 | ||
| ♂ | 5 | 180 mg/kg b.w. | 0.90 ± 0.17 | 0.46 ± 0.04 | 0.85 ± 0.05 | |
| ♀ | 5 | 1.16 ± 0.57 | 0.51 ± 0.10 | 0.93 ± 0.09 | ||
| ♂ | 5 | 360 mg/kg b.w. | 0.84 ± 0.38 | 0.43 ± 0.12 | 0.89 ± 0.19 | |
| ♀ | 5 | 1.32 ± 0.22 | 0.56 ± 0.04 | 0.94 ± 0.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medrano-Padial, C.; Puerto, M.; Prieto, A.I.; Ayala, N.; Beaumont, P.; Rouger, C.; Krisa, S.; Pichardo, S. In Vivo Genotoxicity Evaluation of a Stilbene Extract Prior to Its Use as a Natural Additive: A Combination of the Micronucleus Test and the Comet Assay. Foods 2021, 10, 439. https://doi.org/10.3390/foods10020439
Medrano-Padial C, Puerto M, Prieto AI, Ayala N, Beaumont P, Rouger C, Krisa S, Pichardo S. In Vivo Genotoxicity Evaluation of a Stilbene Extract Prior to Its Use as a Natural Additive: A Combination of the Micronucleus Test and the Comet Assay. Foods. 2021; 10(2):439. https://doi.org/10.3390/foods10020439
Chicago/Turabian StyleMedrano-Padial, Concepción, María Puerto, Ana Isabel Prieto, Nahúm Ayala, Pauline Beaumont, Caroline Rouger, Stéphanie Krisa, and Silvia Pichardo. 2021. "In Vivo Genotoxicity Evaluation of a Stilbene Extract Prior to Its Use as a Natural Additive: A Combination of the Micronucleus Test and the Comet Assay" Foods 10, no. 2: 439. https://doi.org/10.3390/foods10020439
APA StyleMedrano-Padial, C., Puerto, M., Prieto, A. I., Ayala, N., Beaumont, P., Rouger, C., Krisa, S., & Pichardo, S. (2021). In Vivo Genotoxicity Evaluation of a Stilbene Extract Prior to Its Use as a Natural Additive: A Combination of the Micronucleus Test and the Comet Assay. Foods, 10(2), 439. https://doi.org/10.3390/foods10020439